Caveolin-1 Regulates P2Y2 Receptor Signaling during Mechanical Injury in Human 1321N1 Astrocytoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Cell Culture
2.3. Mechanical Injury
2.4. Mechanical Injury
2.5. Cell Viability Assay
2.6. Caspase-9 Activity
2.7. Protein Extraction
2.8. SDS-PAGE and Immunoblot Analysis
2.9. Statistical Analysis
3. Results
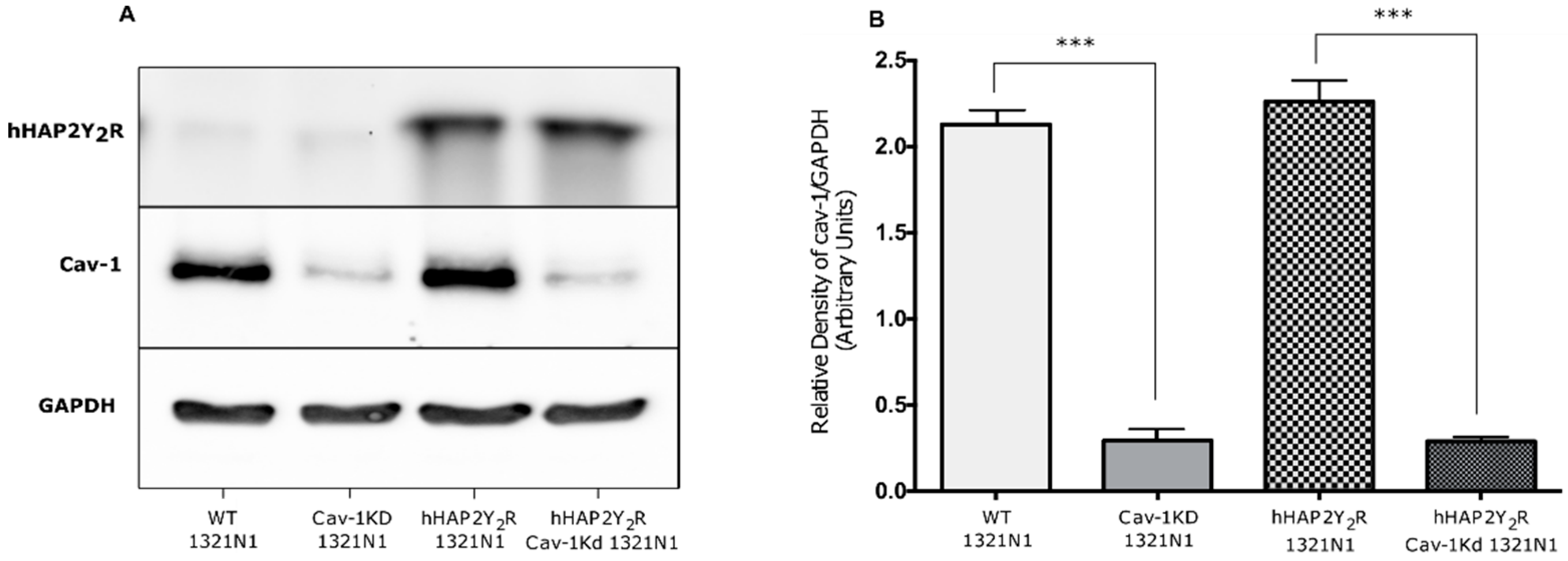
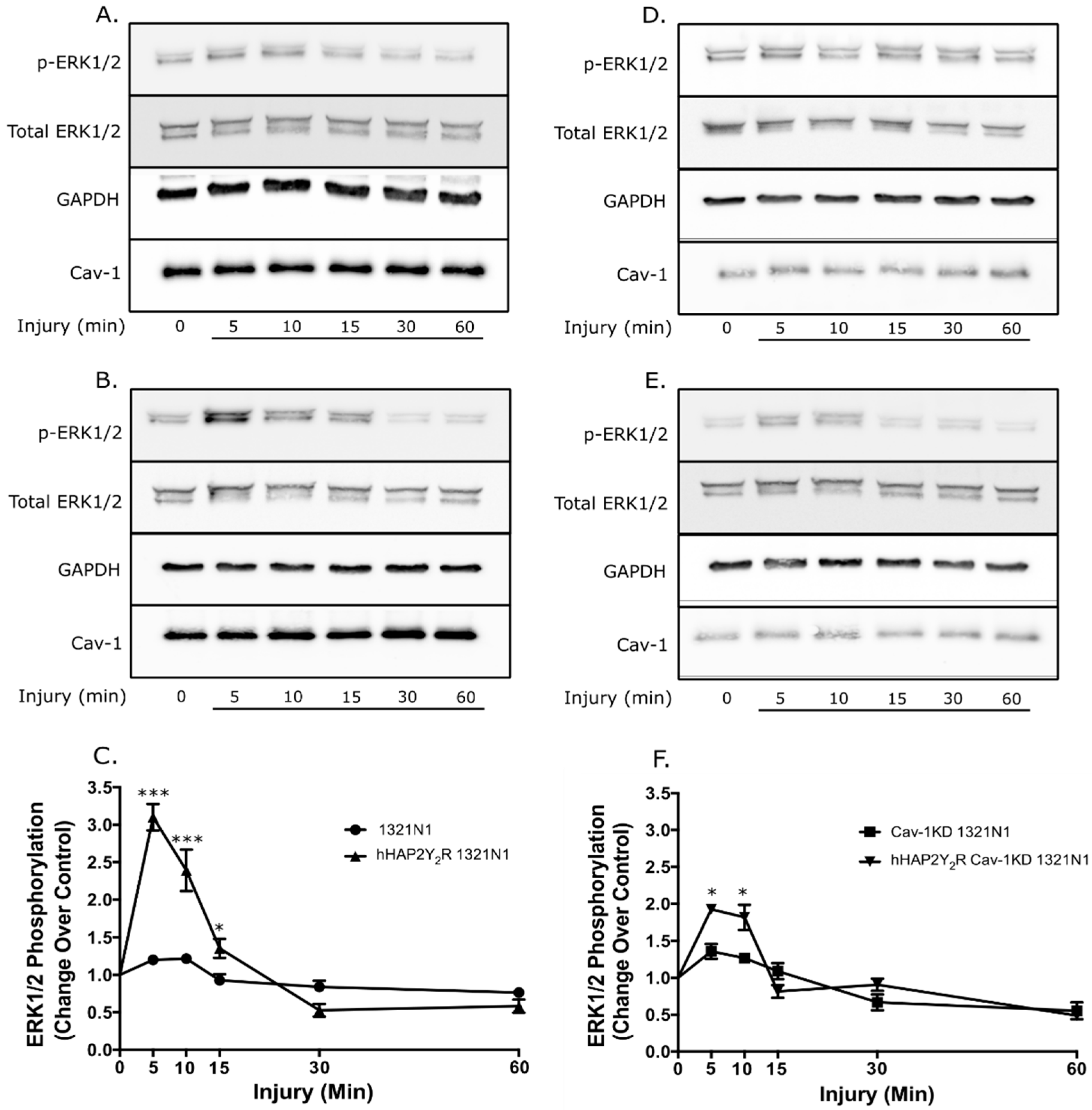
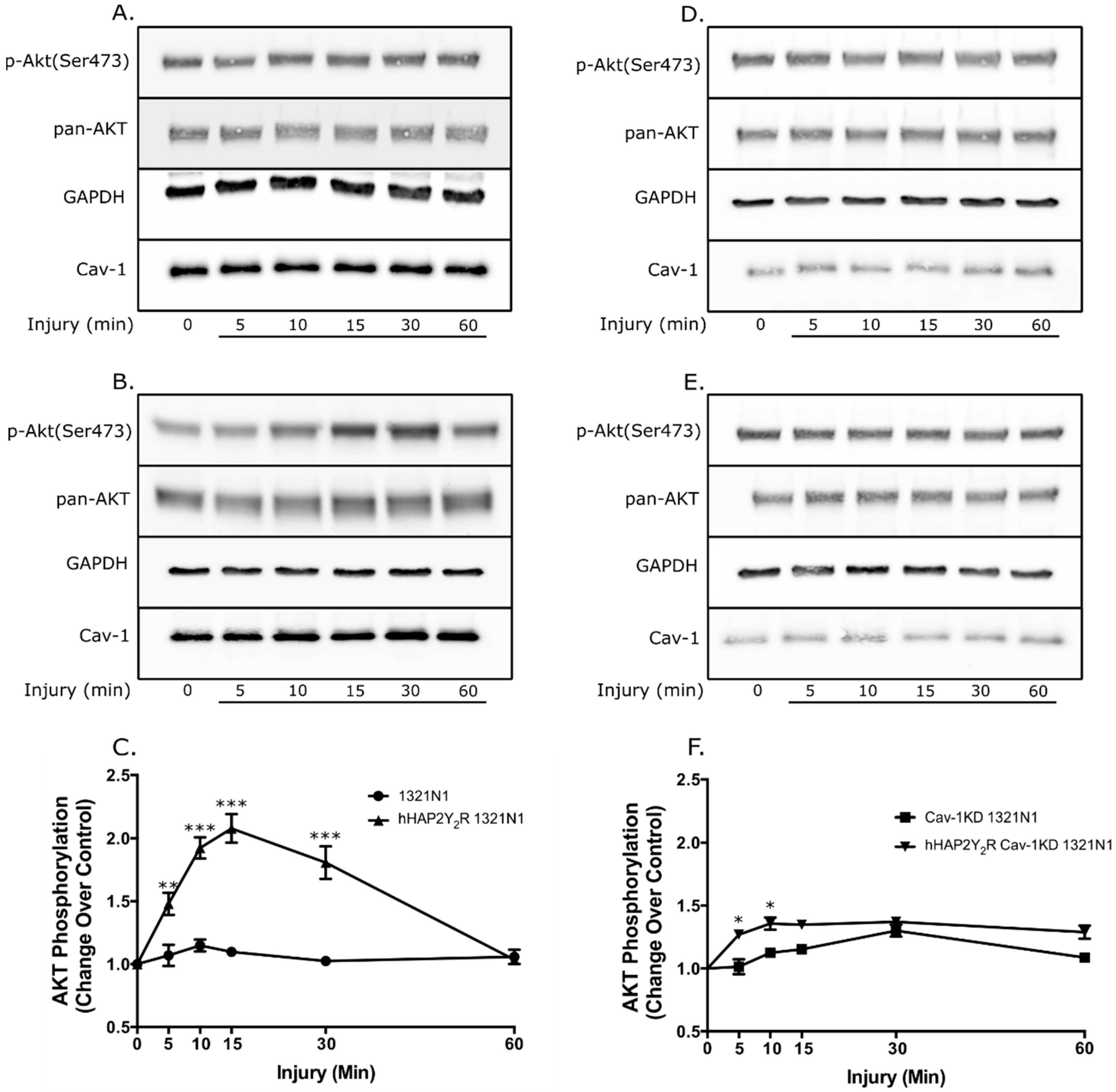
3.1. Cav-1 Knockdown Modulates P2Y2R Signaling Behavior after Mechanical Injury
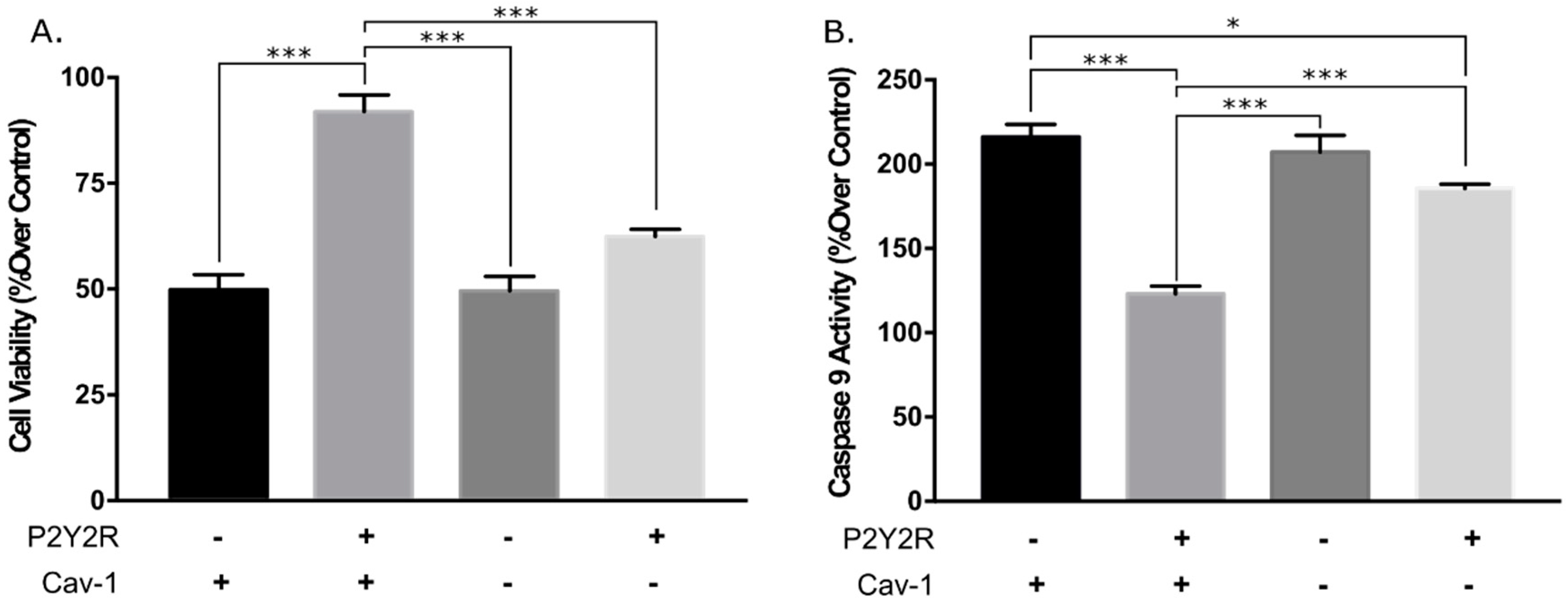
3.2. Knockdown of Caveolin-1 Expression Inhibits hP2Y2R-Mediated Increased Cell Viability and Anti-Apoptotic Actions after Mechanical Injury
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and pathology. Acta Neuropathol. 2009, 119, 7–35. [Google Scholar] [CrossRef]
- Buffo, A.; Rolando, C.; Ceruti, S. Astrocytes in the damaged brain: Molecular and cellular insights into their reactive response and healing potential. Biochem. Pharmacol. 2010, 79, 77–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weisman, G.A.; Ajit, D.; Garrad, R.; Peterson, T.S.; Woods, L.T.; Thebeau, C.; Erb, L. Neuroprotective roles of the P2Y2 receptor. Purinergic Signal. 2012, 8, 559–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neary, J.T.; Kang, Y.; Willoughby, K.A.; Ellis, E.F. Activation of Extracellular Signal-Regulated Kinase by Stretch-Induced Injury in Astrocytes Involves Extracellular ATP and P2 Purinergic Receptors. J. Neurosci. 2012, 23, 2348–2356. [Google Scholar] [CrossRef]
- Neary, J.T.; Zimmermann, H. Trophic functions of nucleotides in the central nervous system. Trends Neurosci. 2009, 32, 189–198. [Google Scholar] [CrossRef]
- Weisman, G.A.; Camden, J.M.; Peterson, T.S.; Ajit, D.; Woods, L.T.; Erb, L. P2 Receptors for Extracellular Nucleotides in the Central Nervous System: Role of P2 × 7 and P2Y2 Receptor Interactions in Neuroinflammation. Mol. Neurobiol. 2012, 46, 96–113. [Google Scholar] [CrossRef]
- Lazarowski, E.R.; Homolya, L.; Boucher, R.C.; Harden, T.K. Direct Demonstration of Mechanically Induced Release of Cellular UTP and Its Implication for Uridine Nucleotide Receptor Activation. J. Biol. Chem. 1997, 272, 24348–24354. [Google Scholar] [CrossRef] [Green Version]
- Peterson, T.S.; Camden, J.M.; Wang, Y.; Seye, C.I.; Wood, W.G.; Sun, G.Y.; Weisman, G.A. P2Y2 Nucleotide Receptor-Mediated Responses in Brain Cells. Mol. Neurobiol. 2010, 41, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Peterson, T.S.; Thebeau, C.N.; Ajit, D.; Camden, J.M.; Woods, L.T.; Wood, W.G.; Weisman, G.A. Up-regulation and activation of the P2Y2nucleotide receptor mediate neurite extension in IL-1β-treated mouse primary cortical neurons. J. Neurochem. 2013, 125, 885–896. [Google Scholar] [CrossRef]
- Burgos, M.; Neary, J.T.; González, F.A. P2Y2nucleotide receptors inhibit trauma-induced death of astrocytic cells. J. Neurochem. 2007, 103, 1785–1800. [Google Scholar] [CrossRef]
- Chorna, N.E.; Santiago-Perez, L.I.; Erb, L.; Seye, C.I.; Neary, J.T.; Sun, G.Y.; Gonzalez, F.A. P2Y2 receptors activate neuroprotective mechanisms in astrocytic cells. J. Neurochem. 2014, 91, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Bastiani, M.; Parton, R.G. Caveolae at a glance. J. Cell Sci. 2010, 123, 3831–3836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, W.; Maldonado, H.; Velázquez, G.; Rubio-Dávila, M.; Miranda, J.; Aquino, E.; Salgado-Villanueva, I. Caveolin isoform expression during differentiation of C6 glioma cells. Int. J. Dev. Neurosci. 2005, 23, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Boscher, C.; Nabi, I.R. CAVEOLIN-1: Role in Cell Signaling. Adv. Exp. Med. Biol. Caveolins Caveolae 2012, 29–50. [Google Scholar]
- Martinez, N.A.; Ayala, A.M.; Martinez, M.; Martinez-Rivera, F.J.; Miranda, J.D.; Silva, W.I. Caveolin-1 Regulates the P2Y2Receptor Signaling in Human 1321N1 Astrocytoma Cells. J. Biol. Chem. 2016, 291, 12208–12222. [Google Scholar] [CrossRef]
- Hsieh, T.; Kang, J.; Lai, J.; Huang, Y.; Rotenberg, A.; Chen, K.; Peng, C. Relationship of mechanical impact magnitude to neurologic dysfunction severity in a rat traumatic brain injury model. PLoS ONE 2017, 12, e0178186. [Google Scholar] [CrossRef]
- Prins, M.; Greco, T.; Alexander, D.; Giza, C.C. The pathophysiology of traumatic brain injury at a glance. Dis. Models Mech. 2012, 6, 1307–1315. [Google Scholar] [CrossRef]
- Cantu, R.C.; Cantu, R.V. Moderate to Severe Traumatic Brain Injury in Sports. Oxf. Handb. Online 2014. [Google Scholar] [CrossRef]
- Miras-Portugal, M.T.; Gomez-Villafuertes, R.; Gualix, J.; Diaz-Hernandez, J.I.; Artalejo, A.R.; Ortega, F.; Delicado, E.G.; Perez-Sen, R. Nucleotides in neuroregeneration and neuroprotection. Neuropharmacology 2016, 104, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signaling during inflammation. Nature 2014, 509, 310–317. [Google Scholar] [CrossRef]
- Mikolajewicz, N.; Zimmermann, E.A.; Willie, B.M.; Komarova, S.V. Mechanically stimulated ATP release from murine bone cells is regulated by a balance of injury and repair. ELife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Erb, L.; Weisman, G.A. Coupling of P2Y receptors to G proteins and other signaling pathways. Wiley Interdiscip. Rev.: Membr. Transport. Signal. 2012, 1, 789–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Qiu, Y.; Zhang, H.; Tian, X.; Fang, W. P2Y2 Receptor and EGFR Cooperate to Promote Prostate Cancer Cell Invasion via ERK1/2 Pathway. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Arthur, D.B.; Georgi, S.; Akassoglou, K.; Insel, P.A. Inhibition of Apoptosis by P2Y2 Receptor Activation: Novel Pathways for Neuronal Survival. J. Neurosci. 2006, 26, 3798–3804. [Google Scholar] [CrossRef]
- Agca, Y.; Qian, S.; Agca, C.; Seye, C.I. Direct Evidence for P2Y2 Receptor Involvement in Vascular Response to Injury. J. Vasc. Res. 2016, 53, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Hutchinson, D.S.; Halls, M.L.; Furness, S.G.; Bengtsson, T.; Evans, B.A.; Summers, R.J. Interaction with Caveolin-1 Modulates G Protein Coupling of Mouse β3-Adrenoceptor. J. Biol. Chem. 2012, 287, 20674–20688. [Google Scholar] [CrossRef]
- Ketteler, J.; Klein, D. Caveolin-1, cancer and therapy resistance. Int. J. Cancer 2018, 143, 2092–2104. [Google Scholar] [CrossRef]
- Quest, A.F.; Lobos-González, L.; Nuñez, S.; Sanhueza, C.; Fernández, J.G.; Aguirre, A.; Rodríguez, D.; Leyton, L.; Torres, V. The caveolin-1 connection to cell death and survival. Curr. Mol. Med. 2013, 13, 266–281. [Google Scholar] [CrossRef]
- Pani, B.; Singh, B.B. Lipid rafts/caveolae as microdomains of calcium signaling. Cell Calcium 2009, 45, 625–633. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Lee, S.; An, C.; Xu, J.; Joshi, B.; Nabi, I.R.; Jin, Y. Caveolin-1 mediates Fas–BID signaling in hyperoxia-induced apoptosis. Free Radic. Biol. Med. 2011, 50, 1252–1262. [Google Scholar] [CrossRef]
- Xu, L.; Wang, L.; Wen, Z.; Wu, L.; Jiang, Y.; Yang, L.; Liu, X. Caveolin-1 is a checkpoint regulator in hypoxia-induced astrocyte apoptosis via Ras/Raf/ERK pathway. Am. J. Physiol. -Cell Physiol. 2016, 310. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Ziegler, N.; Reiner, S.; Krasel, C.; Lohse, M.J. Agonist-selective, Receptor-specific Interaction of Human P2YReceptors with β-Arrestin-1 and -2. J. Biol. Chem. 2008, 283, 30933–30941. [Google Scholar] [CrossRef] [PubMed]
- Khalafalla, F.G.; Greene, S.; Khan, H.; Ilves, K.; Monsanto, M.M.; Alvarez, R.; Sussman, M.A. P2Y2 Nucleotide Receptor Prompts Human Cardiac Progenitor Cell Activation by Modulating Hippo Signaling. Circ. Res. 2017, 121, 1224–1236. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAV-1-EXPRESSING | CAV1 KD | |||||
|---|---|---|---|---|---|---|
| SIGNALING | WT-1321N1 | hHAP2Y2R 1321N1 | p-Value | CAV-1 KD 1321N1 | CAV-1 KD hHAP2Y2R 1321N1 | p-Value |
| pERK1/2 | 2.33 (+/−0.48) | 20.31 (+/−2.24) | *** | 2.47 (+/−0.65) | 8.41 (+/−0.95) | * |
| pAkt | 5.50 (+/−1.61) | 37.23 (+/−2.81) | *** | 8.59 (+/−1.54) | 17.93 (+/−1.50) | * |
| Net P2Y2R-mediated pERK1/2 signaling | 17.98 (+/−2.284) | 5.94 (+/−0.90) | ** | |||
| Net P2Y2R-mediated pAkt signaling | 31.73 (+/−3.51) | 9.35 (+/−1.13) | *** | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez, M.; Martínez, N.A.; Miranda, J.D.; Maldonado, H.M.; Silva Ortiz, W.I. Caveolin-1 Regulates P2Y2 Receptor Signaling during Mechanical Injury in Human 1321N1 Astrocytoma. Biomolecules 2019, 9, 622. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100622
Martínez M, Martínez NA, Miranda JD, Maldonado HM, Silva Ortiz WI. Caveolin-1 Regulates P2Y2 Receptor Signaling during Mechanical Injury in Human 1321N1 Astrocytoma. Biomolecules. 2019; 9(10):622. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100622
Chicago/Turabian StyleMartínez, Magdiel, Namyr A. Martínez, Jorge D. Miranda, Héctor M. Maldonado, and Walter I. Silva Ortiz. 2019. "Caveolin-1 Regulates P2Y2 Receptor Signaling during Mechanical Injury in Human 1321N1 Astrocytoma" Biomolecules 9, no. 10: 622. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100622