Increased Expression of Adherens Junction Components in Mouse Liver following Bile Duct Ligation

 , , ,
, , ,  , ,
, , 
Abstract
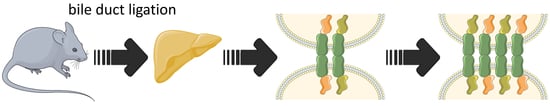
:
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Analysis of Serum Biochemical Parameters
2.3. Histological Examination of Liver Collagen Content
2.4. Reverse Transcription Quantitative Real-Time Polymerase Chain Reaction Analysis
2.5. Immunoblot Analysis
2.6. Immunohistochemistry Analysis
2.7. Statistical Analysis
3. Results
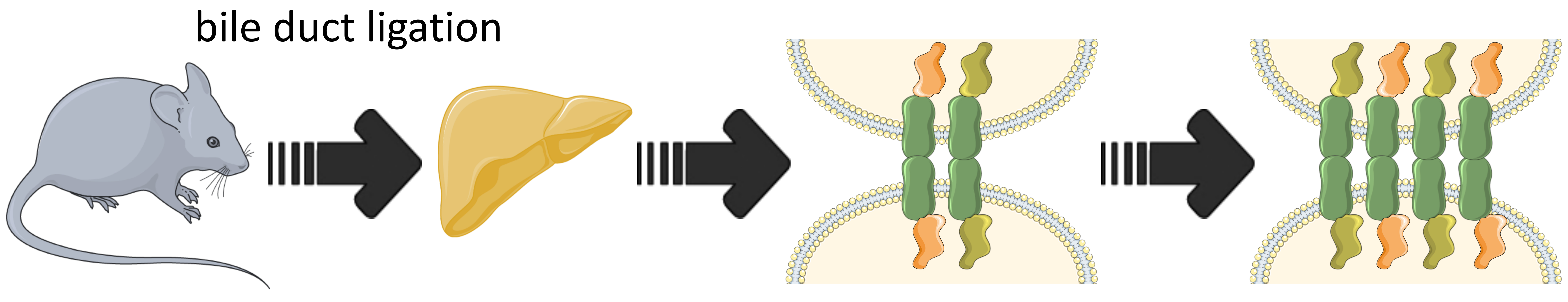
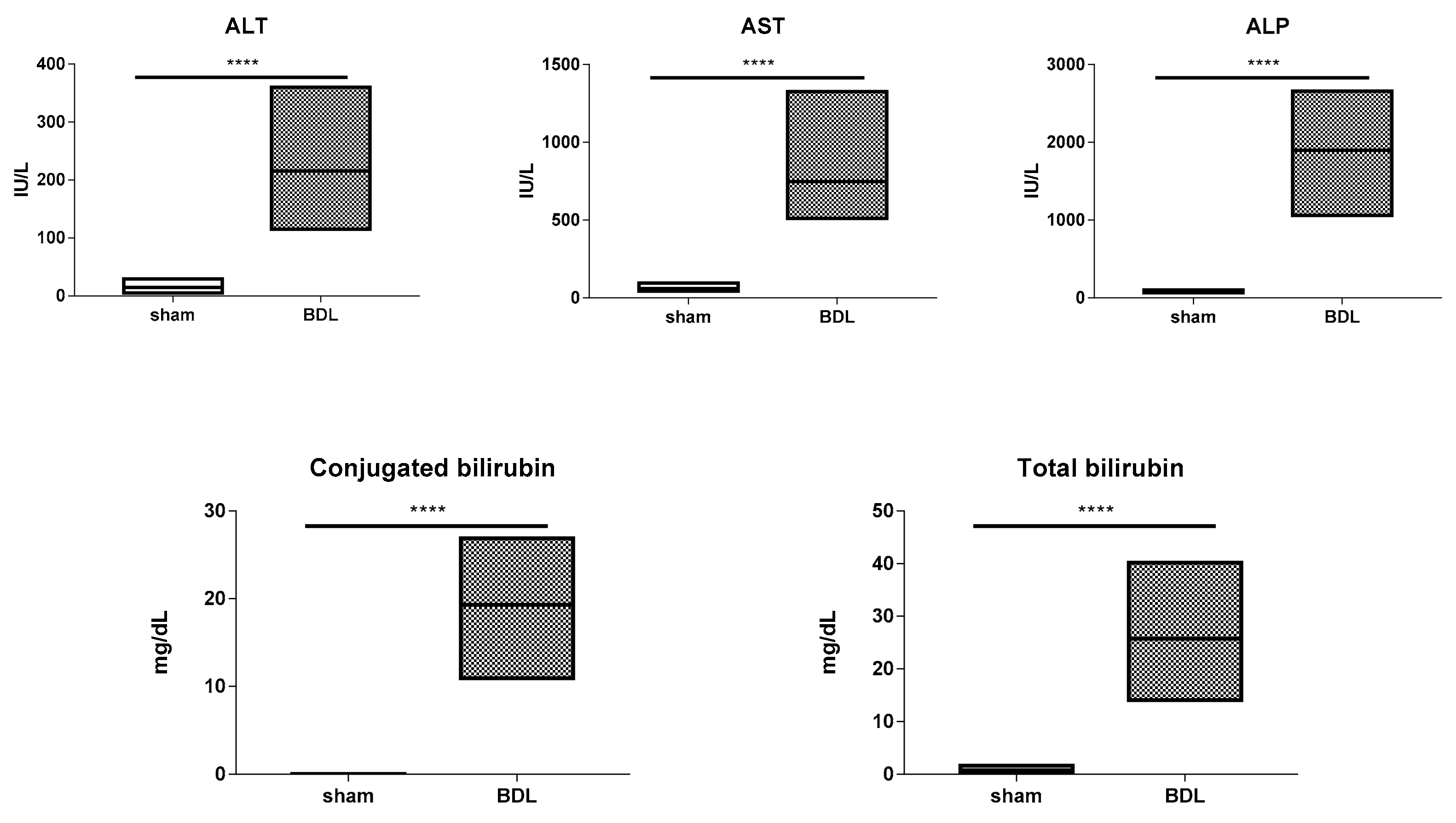
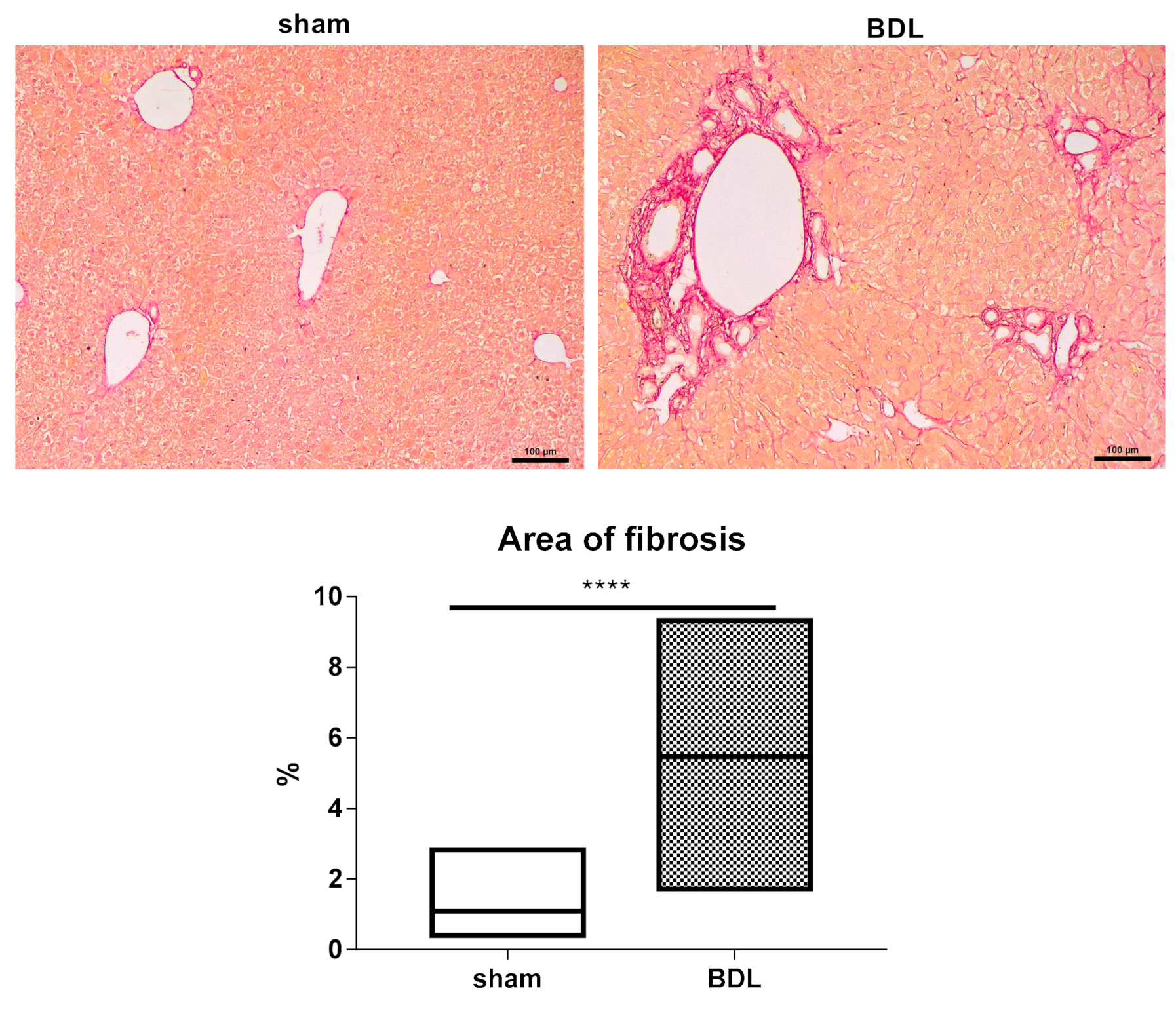
3.1. Characterization of the Bile Duct Ligation Model
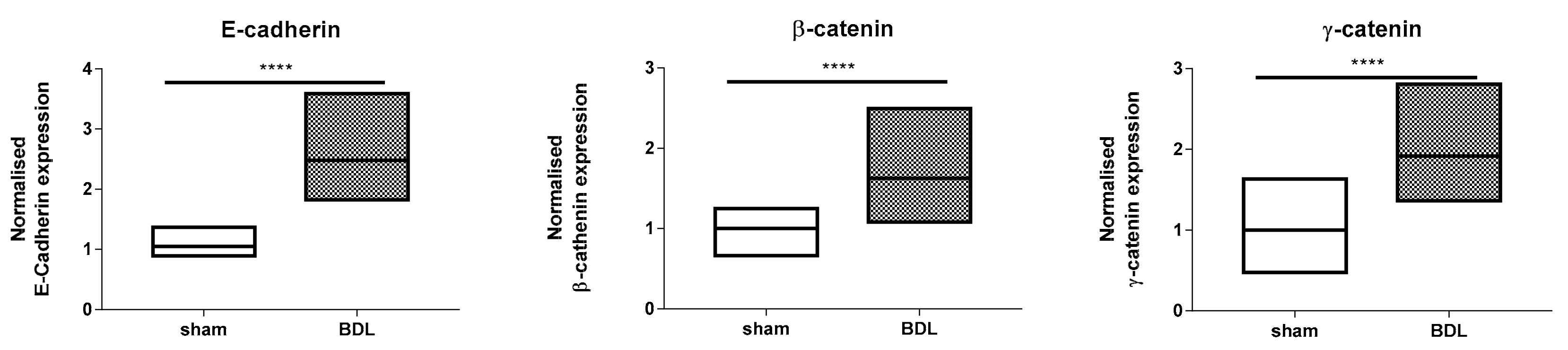
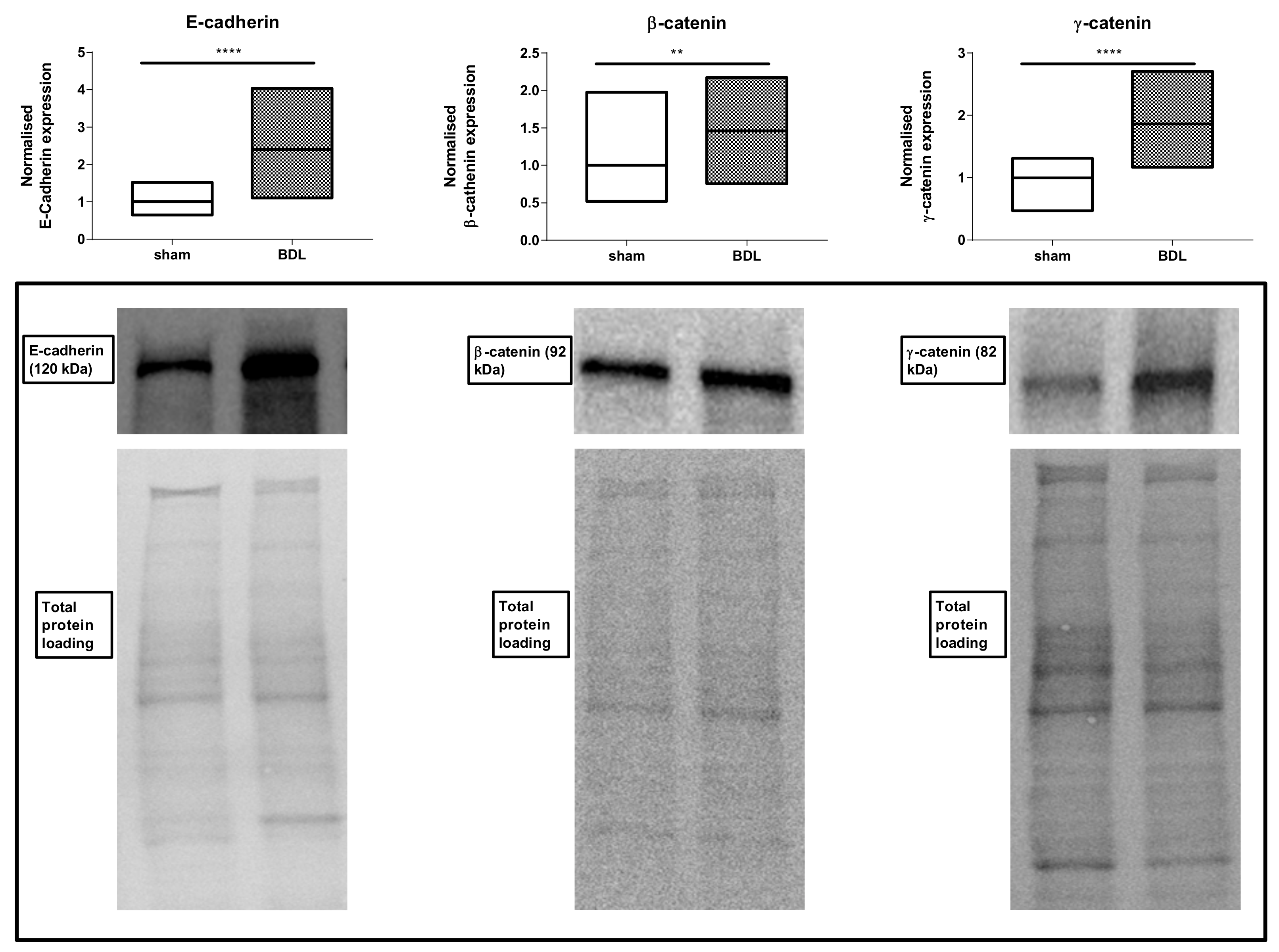
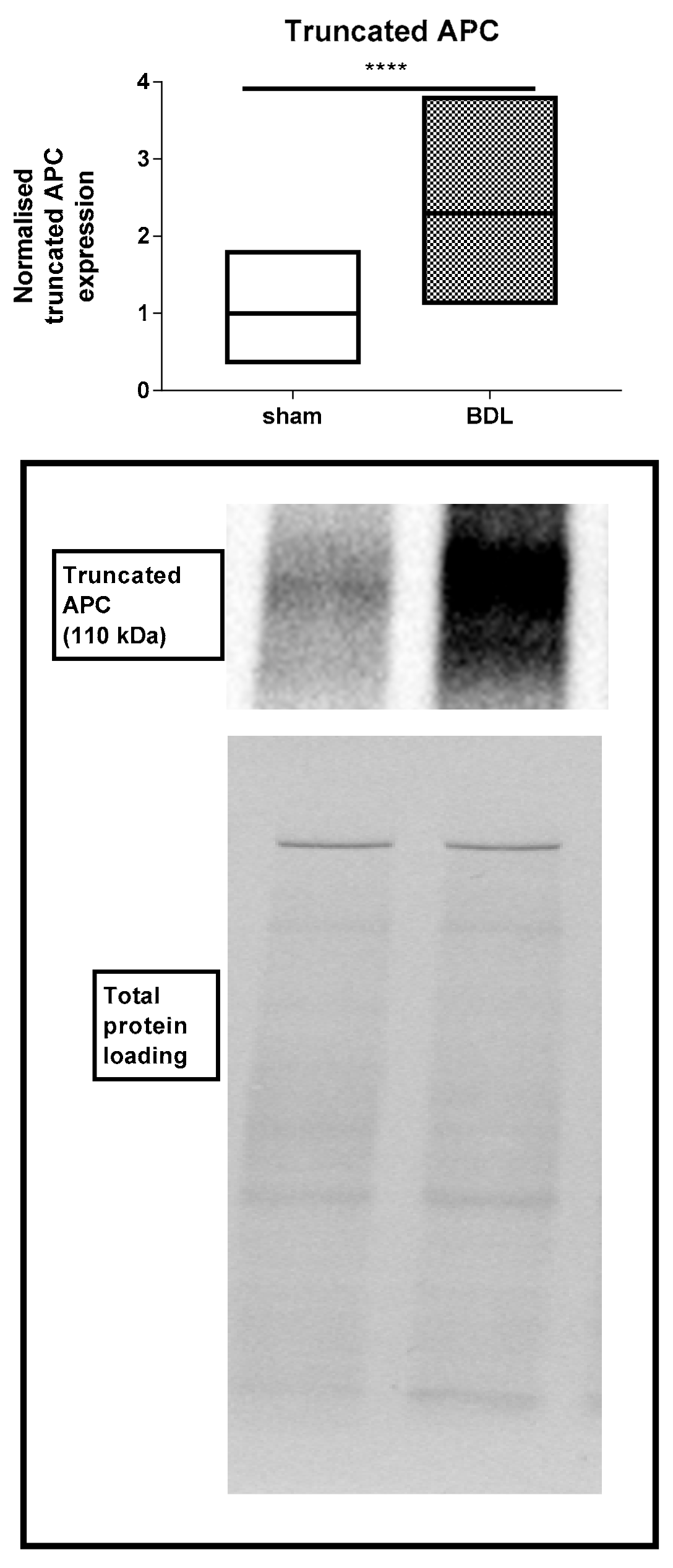
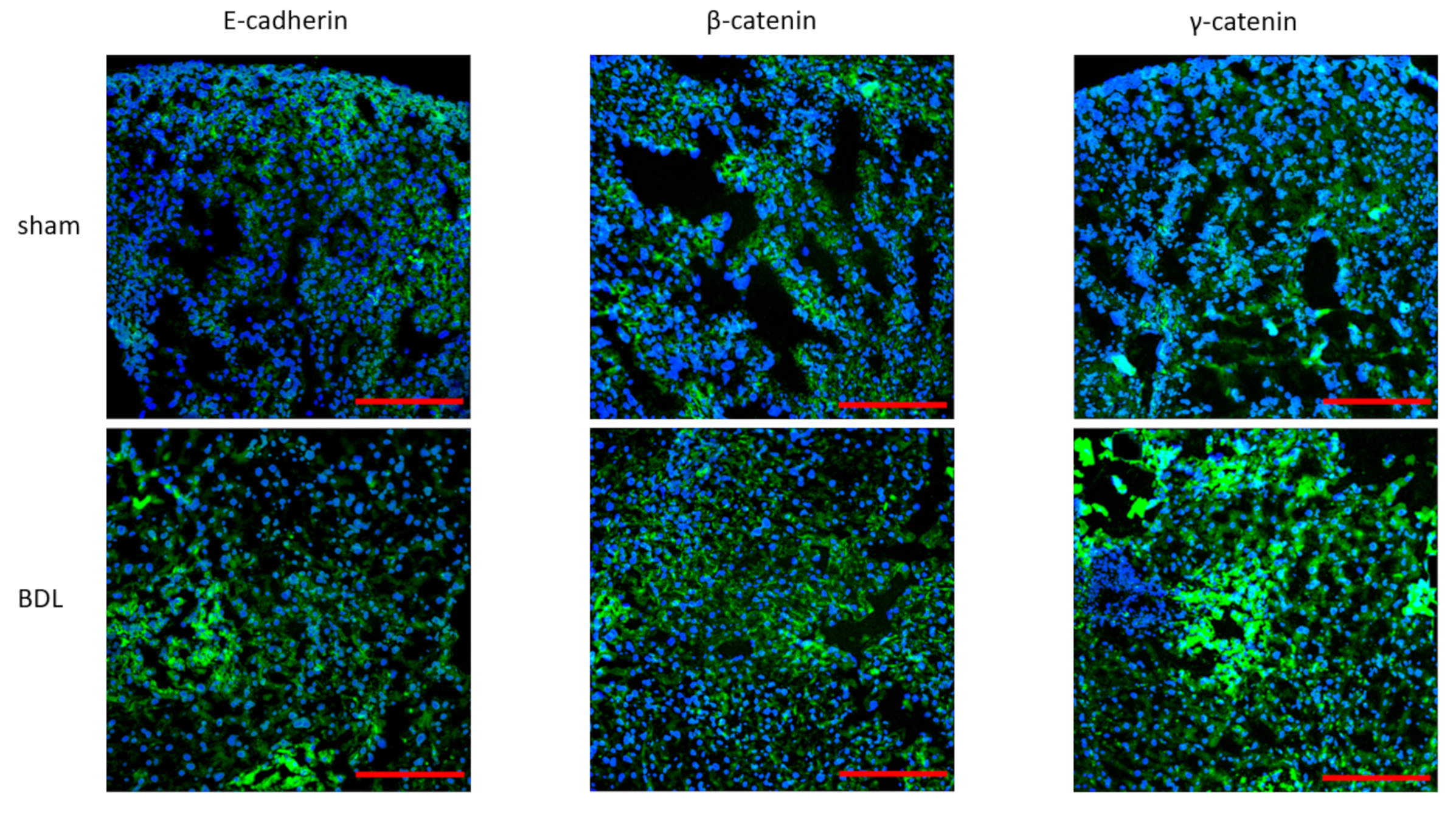
3.2. Effects of Bile Duct Ligation on Hepatic Adherens Junctions
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meng, W.; Takeichi, M. Adherens junction: Molecular architecture and regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a002899. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chung, J.Y.; Rai, U.; Esumi, N. Cadherins in the retinal pigment epithelium (RPE) revisited: P-cadherin is the highly dominant cadherin expressed in human and mouse RPE in vivo. PLoS ONE 2018, 13, e0191279. [Google Scholar] [CrossRef]
- Vinken, M.; Papeleu, P.; Snykers, S.; de Rop, E.; Henkens, T.; Chipman, J.K.; Rogiers, V.; Vanhaecke, T. Involvement of cell junctions in hepatocyte culture functionality. Crit. Rev. Toxicol. 2006, 36, 299–318. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, M.; Schmid, V.J.; Parrish, A.R. Selective disruption of cadherin/catenin complexes by oxidative stress in precision-cut mouse liver slices. Toxicol. Sci. 2001, 61, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, C.; Lawrence, N.; Arias, A.M. Wnt signalling: A theme with nuclear variations. Bioessays 2001, 23, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Wu, R.; Schwartz, D.R.; Darrah, D.; Reed, H.; Kolligs, F.T.; Nieman, M.T.; Fearon, E.R.; Cho, K.R. Role of β-catenin/T-cell factor-regulated genes in ovarian endometrioid adenocarcinomas. Am. J. Pathol. 2002, 160, 1229–1238. [Google Scholar] [CrossRef]
- Akiyama, T. Wnt/β-catenin signaling. Cytokine Growth Factor Rev. 2000, 11, 273–282. [Google Scholar] [CrossRef]
- Davidson, G.; Shen, J.; Huang, Y.L.; Su, Y.; Karaulanov, E.; Bartscherer, K.; Hassler, C.; Stannek, P.; Boutros, M.; Niehrs, C. Cell cycle control of Wnt receptor activation. Dev. Cell 2009, 17, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Giles, R.H.; van Es, J.H.; Clevers, H. Caught up in a Wnt storm: Wnt signaling in cancer. Biochim. Biophys. Acta Rev. Cancer 2003, 1653, 1–24. [Google Scholar] [CrossRef]
- MacDonald, B.T.; Tamai, K.; He, X. Wnt/β-catenin signaling: Components, mechanisms, and diseases. Dev. Cell 2009, 17, 9–26. [Google Scholar] [CrossRef]
- Apte, U.; Singh, S.; Zeng, G.; Cieply, B.; Virji, M.A.; Wu, T.; Monga, S.P.S. Beta-catenin activation promotes liver regeneration after acetaminophen-induced injury. Am. J. Pathol. 2009, 175, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Hempel, M.; Schmitz, A.; Winkler, S.; Kucukoglu, O.; Brückner, S.; Niessen, C.; Christ, B. Pathological implications of cadherin zonation in mouse liver. Cell. Mol. Life Sci. 2015, 72, 2599–2612. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.S.; Wang, Y.J.; Wu, J.X.; Fan, J.G.; Chen, Y.W.; Zhu, L. β-catenin is overexpressed in hepatic fibrosis and blockage of Wnt/β-catenin signaling inhibits hepatic stellate cell activation. Mol. Med. Rep. 2014, 9, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Endo, K.; Ueda, T.; Ueyama, J.; Ohta, T.; Terada, T. Immunoreactive E-cadherin, alpha-catenin, beta-catenin, and gamma-catenin proteins in hepatocellular carcinoma: Relationships with tumor grade, clinicopathologic parameters, and patients’ survival. Hum. Pathol. 2000, 31, 558–565. [Google Scholar] [CrossRef]
- Tag, C.G.; Sauer-Lehnen, S.; Weiskirchen, S.; Borkham-Kamphorst, E.; Tolba, R.H.; Tacke, F.; Weiskirchen, R. Bile duct ligation in mice: induction of inflammatory liver injury and fibrosis by obstructive cholestasis. J. Vis. Exp. 2015, 96, e52438. [Google Scholar] [CrossRef]
- Yanguas, S.C.; da Silva, T.C.; Pereira, I.V.A.; Maes, M.; Willebrords, J.; Shestopalov, V.I.; Goes, B.M.; Nogueira, M.S.; de Castro, I.A.; Romualdo, G.R.; et al. Genetic ablation of pannexin1 counteracts liver fibrosis in a chemical, but not in a surgical mouse model. Arch. Toxicol. 2018, 92, 2607–2627. [Google Scholar] [CrossRef] [Green Version]
- Cogliati, B.; da silva, T.C.; Aloia, T.P.A.; Chaible, L.M.; Real-Lima, M.A.; Sanches, D.S.; Matsuzaki, P.; Hernandez-Blazquez, F.J.; Dagli, M.L.Z. Morphological and molecular pathology of CCl4-induced hepatic fibrosis in connexin43-deficient mice. Microsc. Res. Tech. 2011, 74, 421–429. [Google Scholar] [CrossRef]
- Schindelin, J.; Rueden, C.T.; Hiner, M.C.; Eliceiri, K.W. The ImageJ ecosystem: An open platform for biomedical image analysis. Mol. Reprod. Dev. 2015, 82, 518–529. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.; Mcgill, M.R.; Cristina, T.; Abels, C.; Weemhoff, J.L.; Tiburcio, T.; Veloso, I.; Pereira, A.; Paulo, S. Inhibition of pannexin1 channels alleviates acetaminophen-induced hepatotoxicity. Arch. Toxicol. 2017, 91, 2245–2261. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Eaton, S.L.; Roche, S.L.; Hurtado, M.L.; Oldknow, K.J.; Farquharson, C.; Gillingwater, T.H.; Wishart, T.M. Total protein analysis as a reliable loading control for quantitative fluorescent Western blotting. PLoS ONE 2013, 8, e72457. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Gutiérrez, B.; Anzola, A.; Martínez-Augustin, O.; de Medina, F.S. Stain-free detection as loading control alternative to Ponceau and housekeeping protein immunodetection in Western blotting. Anal. Biochem. 2014, 467, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.C.; Berkelman, T.; Yadav, G.; Hammond, M. A defined methodology for reliable quantification of western blot data. Mol. Biotechnol. 2013, 55, 217–226. [Google Scholar] [CrossRef]
- Taylor, S.C.; Posch, A. The design of a quantitative western blot experiment. Biomed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yanguas, S.C.; Cogliati, B.; Willebrords, J.; Maes, M.; Colle, I.; van den Bossche, B.; de Oliveira, C.P.M.S.; Andraus, W.; Alves, V.A.; Leclercq, I.; et al. Experimental models of liver fibrosis. Arch. Toxicol. 2016, 90, 1025–1048. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, P.; Jochum, W.; Heinrich, S.; Jang, J.H.; Nocito, A.; Dahm, F.; Clavien, P.A. Characterization of time-related changes after experimental bile duct ligation. Br. J. Surg. 2008, 95, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Maly, I.P.; Landmann, L. Bile duct ligation in the rat causes upregulation of ZO-2 and decreased colocalization of claudins with ZO−1 and occludin. Histochem. Cell Biol. 2008, 129, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Pradhan-Sundd, T.; Poddar, M.; Singh, S.; Kikuchi, A.; Stolz, D.B.; Shou, W.; Li, Z.; Nejak-Bowen, K.N.; Monga, S.P. Mice with hepatic loss of the desmosomal protein γ-catenin are prone to cholestatic injury and chemical carcinogenesis. Am. J. Pathol. 2015, 185, 3274–3289. [Google Scholar] [CrossRef] [PubMed]
- Theodorakis, N.G.; de Maio, A. Cx32 mRNA in rat liver: Effects of inflammation on poly(A) tail distribution and mRNA degradation. Am. J. Physiol. Integr. Comp. Physiol. 2017, 276, R1249–R1257. [Google Scholar] [CrossRef] [PubMed]
- Monga, S.P. β-catenin signaling and roles in liver homeostasis, injury, and tumorigenesis. Gastroenterology 2015, 148, 1294–1310. [Google Scholar] [CrossRef]
- Matsui, T.; Kinoshita, T.; Morikawa, Y.; Tohya, K.; Katsuki, M.; Ito, Y.; Kamiya, A.; Miyajima, A. K-Ras mediates cytokine-induced formation of E-cadherin-based adherens junctions during liver development. EMBO J. 2008, 21, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.S.; Diehl, A.M. Epithelial-to-mesenchymal transitions in the liver. Hepatology 2009, 50, 2007–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, G.; Diehl, A.M. Evidence for and against epithelial-to-mesenchymal transition in the liver. Am. J. Physiol. Liver Physiol. 2013, 305, G881–G890. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.F.; Wu, X.M.; Liu, M. Hepatic regeneration and the epithelial to mesenchymal transition. World J. Gastroenterol. 2013, 19, 1380–1386. [Google Scholar] [CrossRef] [Green Version]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2011, 119, 1420–1428. [Google Scholar] [CrossRef]
- Wells, R.G. The epithelial-to-mesenchymal transition in liver fibrosis: Here today, gone tomorrow? Hepatology 2010, 51, 737–740. [Google Scholar] [CrossRef]
- Xia, J.L.; Dai, C.; Michalopoulos, G.K.; Liu, Y. Hepatocyte growth factor attenuates liver fibrosis induced by bile duct ligation. Am. J. Pathol. 2006, 168, 1500–1512. [Google Scholar] [CrossRef]
- Zeisberg, M.; Yang, C.; Martino, M.; Duncan, M.B.; Rieder, F.; Tanjore, H.; Kalluri, R. Fibroblasts derive from hepatocytes in liver fibrosis via epithelial to mesenchymal transition. J. Biol. Chem. 2007, 282, 23337–23347. [Google Scholar] [CrossRef]
- Thiery, J.P.; Sleeman, J.P. Complex networks orchestrate epithelial-mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 2006, 7, 131–142. [Google Scholar] [CrossRef]
- Zeisberg, M.; Neilson, E.G. Review series personal perspective biomarkers for epithelial-mesenchymal transitions. J. Clin. Investig. 2009, 119, 1429–1437. [Google Scholar] [CrossRef]
- Zhao, Y.L.; Zhu, R.T.; Sun, Y.L. Epithelial-mesenchymal transition in liver fibrosis (review). Biomed. Rep. 2016, 4, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Rygiel, K.A.; Robertson, H.; Marshall, H.L.; Pekalski, M.; Zhao, L.; Booth, T.A.; Jones, D.E.; Burt, A.D.; Kirby, K.J. Epithelial-mesenchymal transition contributes to portal tract fibrogenesis during human chronic liver disease. Lab. Investig. 2008, 88, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Aktary, Z.; Alaee, M.; Pasdar, M. Beyond cell-cell adhesion: Plakoglobin and the regulation of tumorigenesis and metastasis. Oncotarget 2017, 8, 32270–32291. [Google Scholar] [CrossRef] [PubMed]
- Maeda, O.; Usami, N.; Kondo, M.; Takahashi, M.; Goto, H.; Shimokata, K.; Kusugami, K.; Sekido, Y. Plakoglobin (γ-catenin) has TCF/LEF family-dependent transcriptional activity in β-catenin-deficient cell line. Oncogene 2004, 23, 964–972. [Google Scholar] [CrossRef]
- Wickline, E.D.; Awuah, P.K.; Behari, J.; Ross, M.; Stolz, D.B.; Monga, S.P.S. Hepatocyte γ-catenin compensates for conditionally deleted β-catenin at adherens junctions. J. Hepatol. 2011, 55, 1256–1262. [Google Scholar] [CrossRef]
- Kolligs, F.T.; Kolligs, B.; Hajra, K.M.; Gang, H.; Tani, M.; Kathleen, R.C.; Fearon, E.R. γ-Catenin is regulated by the APC tumor suppressor and its oncogenic activity is distinct from that of β-catenin. Genes Dev. 2000, 14, 1319–1331. [Google Scholar]
- Barth, A.I.M.; Näthke, I.S.; Nelson, W.J. Cadherins, catenins and APC protein: Interplay between cytoskeletal complexes and signaling pathways. Curr. Opin. Cell Biol. 1997, 9, 683–690. [Google Scholar] [CrossRef]
- Okabe, H.; Yang, J.; Sylakowski, K.; Yovchev, M.; Miyagawa, Y.; Nagarajan, S.; Chikina, M.; Thompson, M.; Oertel, M.; Baba, H.; et al. Wnt signaling regulates hepatobiliary repair following cholestatic liver injury in mice. Hepatology 2016, 64, 1652–1666. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.D.; Monga, S.P.S. WNT/β-catenin signaling in liver health and disease. Hepatology 2007, 45, 1298–1305. [Google Scholar] [CrossRef]
- Jirouskova, M.; Nepomucka, K.; Oyman-Eyrilmez, G.; Kalendova, A.; Havelkova, H.; Sarnova, L.; Chalupsky, K.; Schuster, B.; Benada, O.; Miksatkova, P.; et al. Plectin controls biliary tree architecture and stability in cholestasis. J. Hepatol. 2018, 68, 1006–1017. [Google Scholar] [CrossRef]
- Pradhan-sundd, T.; Monga, S.P. Review blood—Bile barrier: Morphology, regulation, and pathophysiology. Gene Expr. J. Liver Res. 2019, 19, 69–87. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, X.; Zhang, F.; Ma, J.; Xu, C. Expression patterns of the cell junction-associated genes during rat liver regeneration. J. Genet. Genom. 2007, 34, 892–908. [Google Scholar] [CrossRef]
- Glück, U.; Fernández, J.R.; Pankov, R.; Ben-Ze’ev, A. Regulation of adherens junction protein expression in growth-activated 3T3 cells and in regenerating liver. Exp. Cell Res. 1992, 202, 477–486. [Google Scholar] [CrossRef]
- Herr, K.J.; Tsang, Y.H.N.; Ong, J.W.E.; Li, Q.; Yap, L.L.; Yu, W.; Yin, H.; Bogorad, R.L.; Dahlman, J.E.; Chan, Y.G.; et al. Loss of α-catenin elicits a cholestatic response and impairs liver regeneration. Sci. Rep. 2014, 4, 6835. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Assay ID | Accession Number | Assay Location | Amplicon Size (Base Pairs) | Exon Boundary |
|---|---|---|---|---|---|
| Actb | Mm00607939_s1 | NM_007393.3 | 1233 | 115 | 6-6 |
| B2m | Mm00437762_m1 | NM_009735.3 | 111 | 77 | 1-2 |
| Cdh1 | Mm01247357_m1 | NM_009864.2 | 1452 | 71 | 9-10 |
| Ctnnb1 | Mm00483039_m1 | NM_007614.3 | 2366 | 77 | 13-14 |
| Gapdh | Mm99999915_g1 | NM_008084.3 | 265 | 107 | 2-3 |
| Hmbs | Mm01143545_m1 | NM_013551.2 | 473 | 81 | 6-7 |
| Jup | Mm00550256_m1 | NM_034723.1 | 1657 | 66 | 8-9 |
| Ubc | Mm02525934_g1 | NM_019639.4 | 370 | 176 | 2-2 |
| Antigen | Supplier | Cat. No. | Species | Type | Dilution |
|---|---|---|---|---|---|
| E-cadherin | BD Biosciences | 610181 | Mouse | Monoclonal | 1/5000 (IB) |
| Cell Signaling Technology | CST 3195S | Rabbit | Monoclonal | 1/200 (IHC) | |
| β-catenin | BD Biosciences | 610153 | Mouse | Monoclonal | 1/1000 (IB) |
| Abcam | Ab2365 | Rabbit | Polyclonal | 1/200 (IHC) | |
| γ-catenin | BD Biosciences | 610253 | Mouse | Monoclonal | 1/1000 (IB) |
| Cell Signaling Technology | CST 2309S | Rabbit | Polyclonal | 1/200 (IHC) | |
| APC | Santa Cruz | sc-9998 | Mouse | Monoclonal | 1/1000 (IB) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Campenhout, R.; Crespo Yanguas, S.; Cooreman, A.; Gijbels, E.; Leroy, K.; Vilas-Boas, V.; Devoogdt, N.; Muyldermans, S.; Cogliati, B.; Vinken, M. Increased Expression of Adherens Junction Components in Mouse Liver following Bile Duct Ligation. Biomolecules 2019, 9, 636. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100636
Van Campenhout R, Crespo Yanguas S, Cooreman A, Gijbels E, Leroy K, Vilas-Boas V, Devoogdt N, Muyldermans S, Cogliati B, Vinken M. Increased Expression of Adherens Junction Components in Mouse Liver following Bile Duct Ligation. Biomolecules. 2019; 9(10):636. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100636
Chicago/Turabian StyleVan Campenhout, Raf, Sara Crespo Yanguas, Axelle Cooreman, Eva Gijbels, Kaat Leroy, Vânia Vilas-Boas, Nick Devoogdt, Serge Muyldermans, Bruno Cogliati, and Mathieu Vinken. 2019. "Increased Expression of Adherens Junction Components in Mouse Liver following Bile Duct Ligation" Biomolecules 9, no. 10: 636. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9100636