High-Fat Diet Affects Ceramide Content, Disturbs Mitochondrial Redox Balance, and Induces Apoptosis in the Submandibular Glands of Mice

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
- (1)
- Control group fed a control rodent diet ad libitum (Research Diets, New Brunswick, NJ, USA, D12450J).
- (2)
- Group of animals fed a high-fat diet (HFD) ad libitum (Research Diets, New Brunswick, NJ, USA D12492).
2.1. Concentration of Plasma Insulin and Blood Glucose, Calculation of HOMA-IR
2.2. Sphingolipids
2.3. Mitochondria Isolation
2.4. Mitochondrial Antioxidants
2.5. Mitochondrial Oxidative Stress
2.6. Mitochondrial ROS Production, Inflammation, and Apoptosis
2.7. Mitochondrial Activity
2.8. Mitochondrial Protein
2.9. Statistical Analysis
3. Results
3.1. General Characteristics
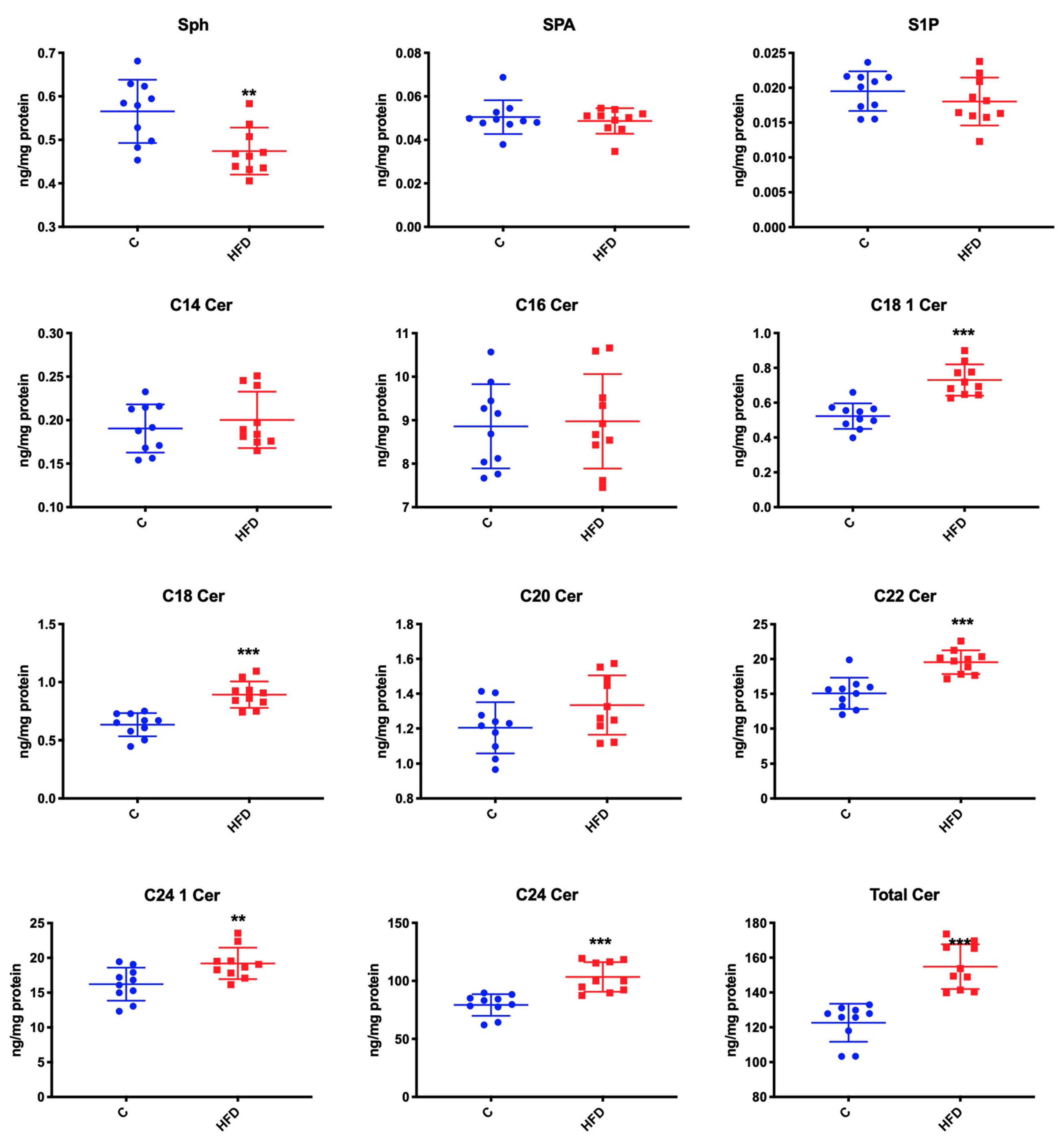
3.2. Effect of High-Fat Diet on Ceramide Contents as Well as S1P, Sphinganine, and Sphingosine Concentration
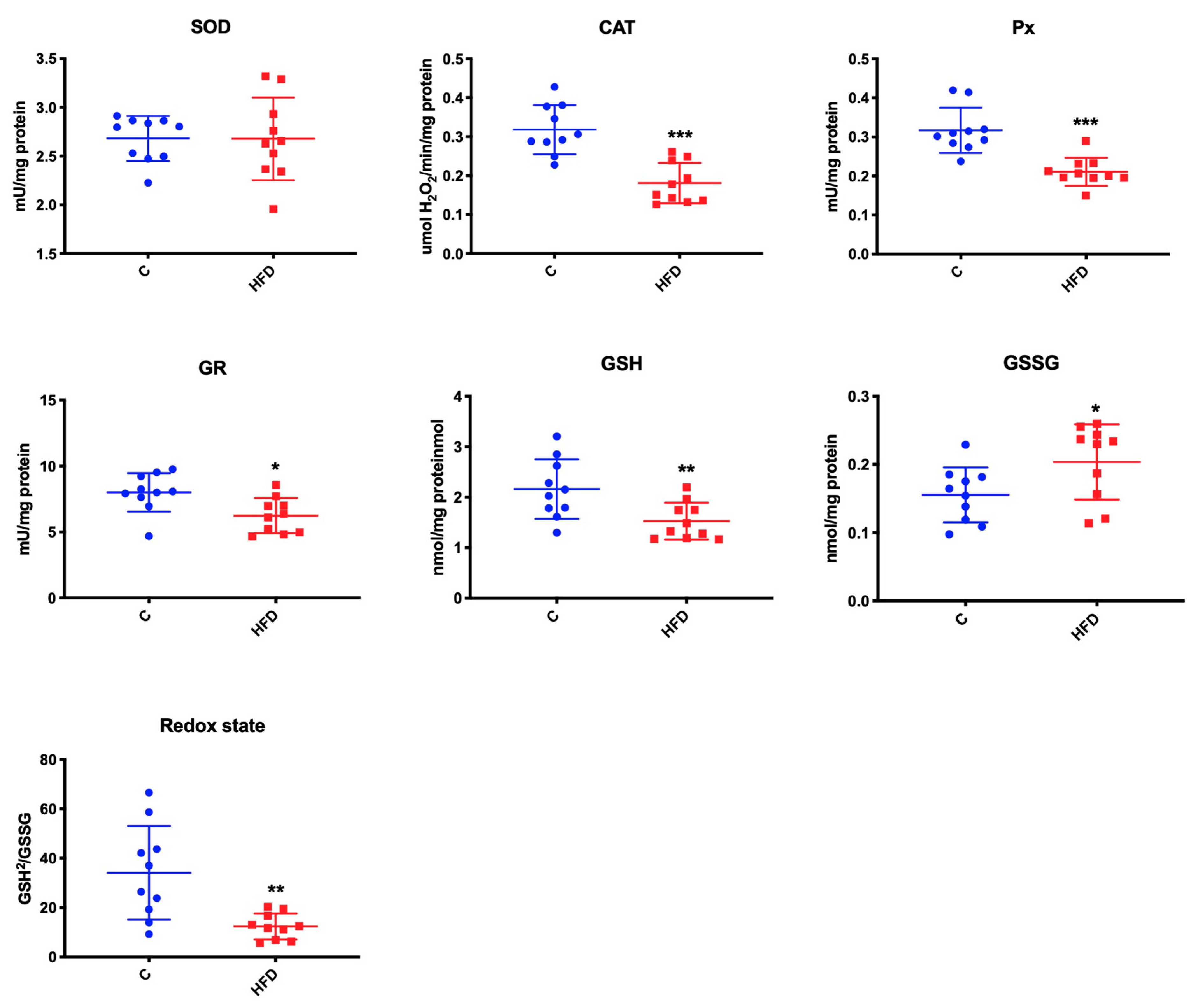
3.3. Effect of High-Fat Diet on Mitochondrial Antioxidants and Redox Ratio
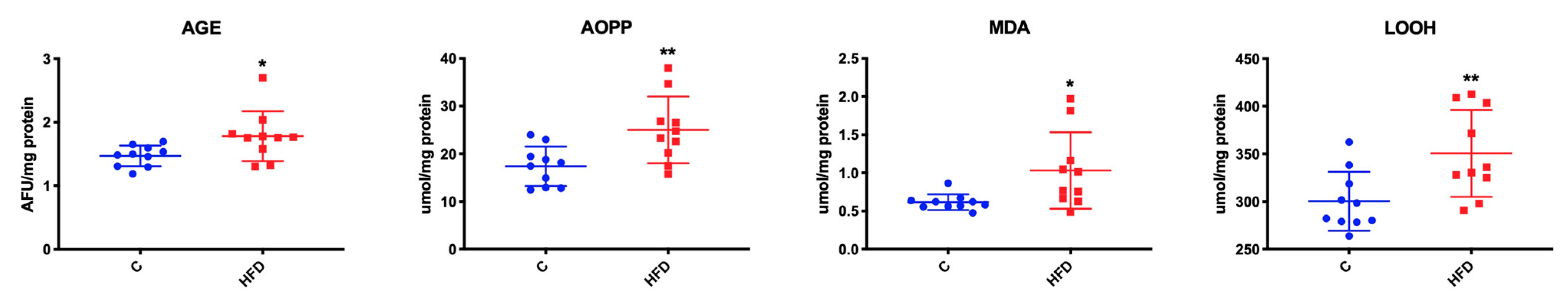
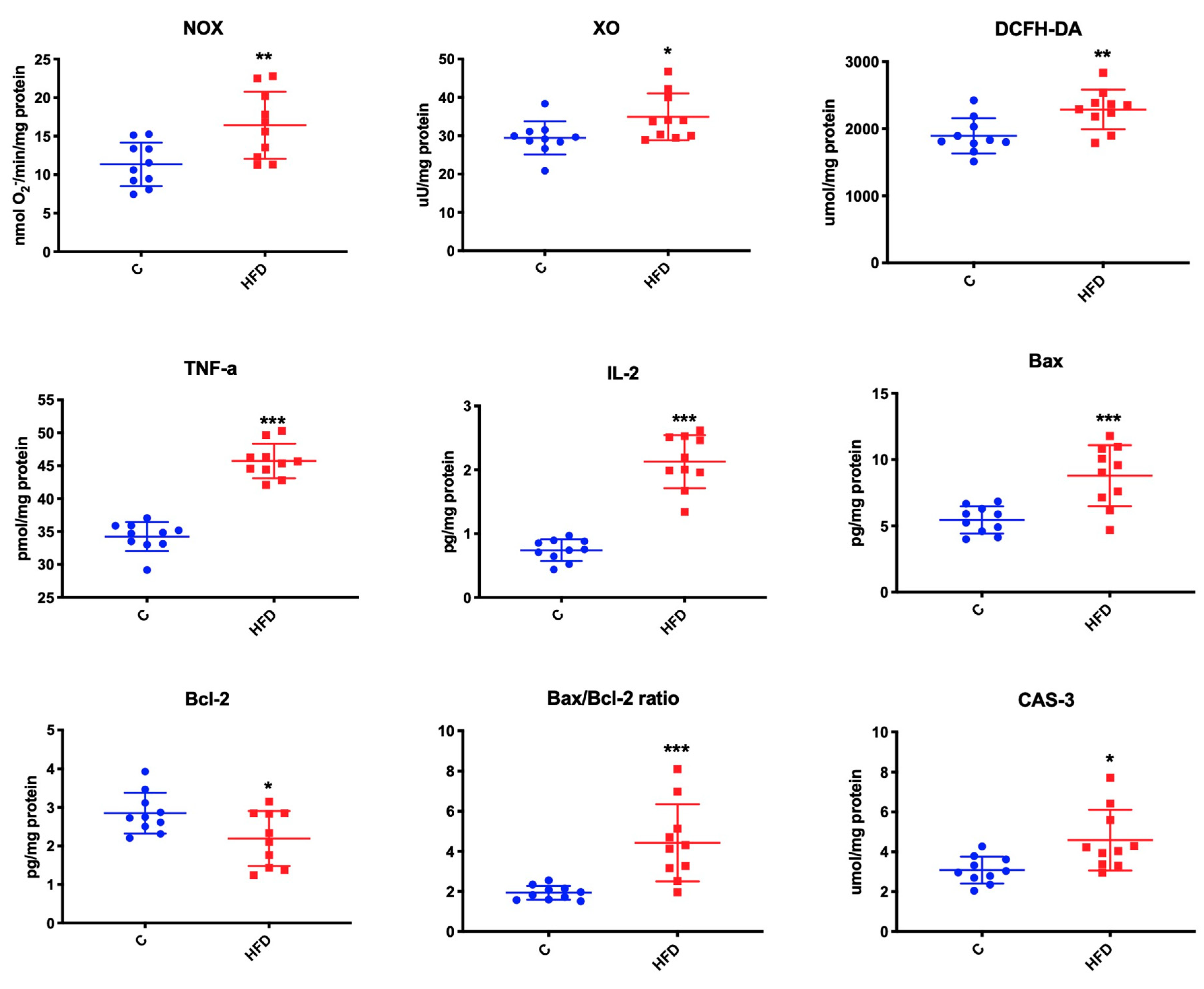
3.4. Effect of High-Fat Diet on Mitochondrial Oxidative Stress
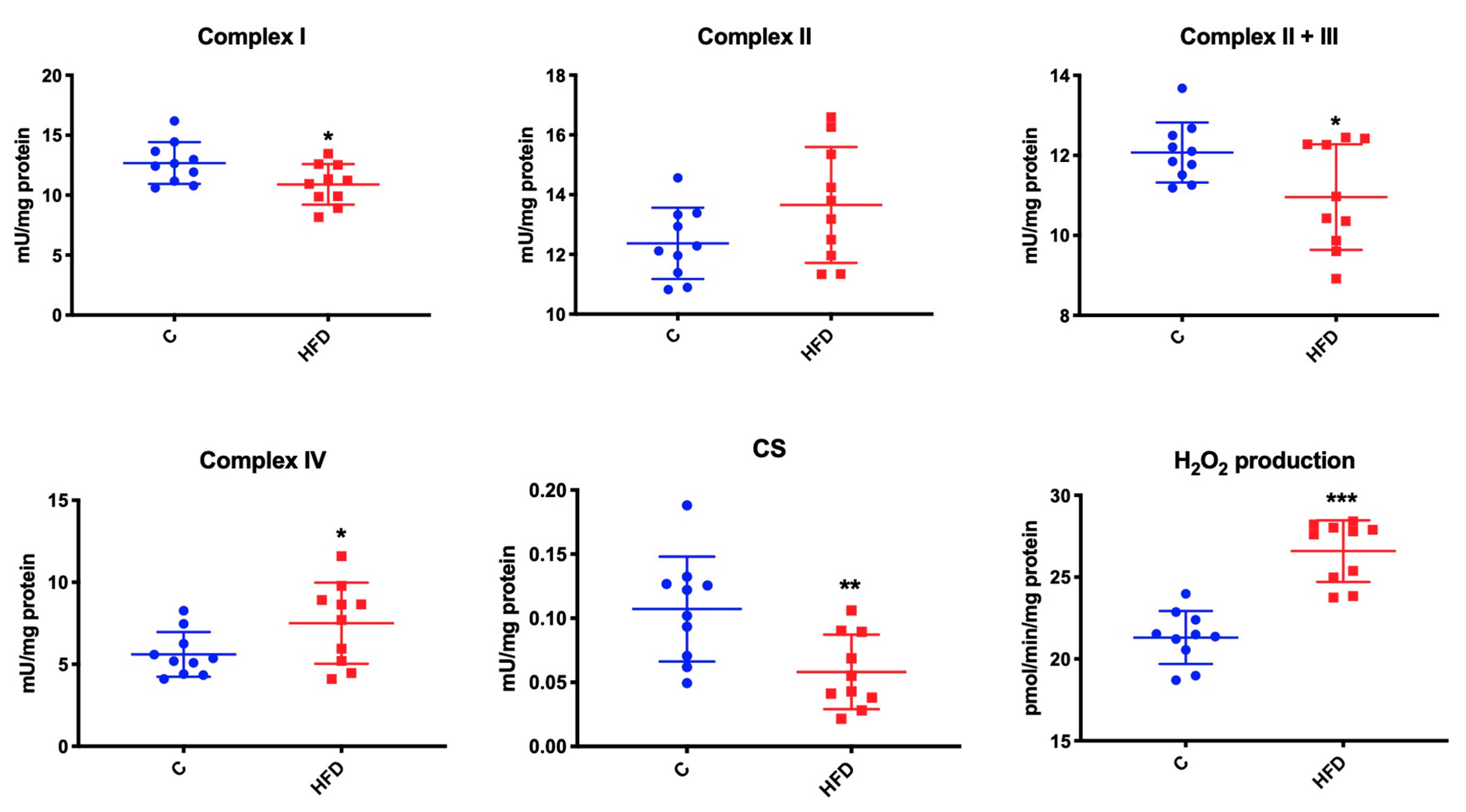
3.5. Effect of High-Fat Diet on Mitochondrial Respiratory Complexes and CS Activity as well as Mitochondrial H2O2 Production
3.6. Effect of High-Fat Diet on Mitochondrial ROS Production, Inflammation, and Apoptosis
3.7. Correlations
4. Discussion
5. Conclusions
- (1)
- A high-fat diet regimen increases the salivary gland ceramide composition.
- (2)
- A high-fat diet intensifies oxidative damage to proteins and lipids and results in inflammation and apoptosis of submandibular gland mitochondria in mice.
- (3)
- The accumulation of some ceramides appears to boost ROS production by affecting NOX activity and complexes I, II + III, and IV in the submandibular gland mitochondria of mice fed a high-fat diet.
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Global Database on Body Mass Index. Available online: http://www.Who.Int/bmi (accessed on 10 November 2019).
- Holland, W.L.; Knotts, T.A.; Chavez, J.A.; Wang, L.P.; Hoehn, K.L.; Summers, S.A. Lipid mediators of insulin resistance. Nutr. Rev. 2007, 65, 39–46. [Google Scholar] [CrossRef]
- Kurek, K.; Miklosz, A.; Lukaszuk, B.; Chabowski, A.; Gorski, J.; Zendzian-Piotrowska, M. Inhibition of ceramide de novo synthesis ameliorates diet induced skeletal muscles insulin resistance. J. Diabetes Res. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, B.A.; Poynten, A.; Lowy, A.J.; Furler, S.M.; Chisholm, D.J.; Kraegen, E.W.; Cooney, G.J. Long-chain acyl-CoA esters as indicators of lipid metabolism and insulin sensitivity in rat and human muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E554–E560. [Google Scholar] [CrossRef] [PubMed]
- Berk, P.D.; Verna, E.C. Nonalcoholic fatty liver disease: Lipids and insulin resistance. Clin. Liver Dis. 2016, 20, 245–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matczuk, J.; Zalewska, A.; Łukaszuk, B.; Knaś, M.; Maciejczyk, M.; Grabowska, M.; Ziembicka, D.M.; Waszkiel, D.; Chabowski, A.; Żendzian-Piotrowska, M.; et al. Insulin resistance and obesity affect lipid profile in the salivary glands. J. Diabetes Res. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogot-Levin, A.; Saada, A. Ceramide and the mitochondrial respiratory chain. Biochimie 2014, 100, 88–94. [Google Scholar] [CrossRef]
- Zhang, L.; Keung, W.; Samokhvalov, V.; Wang, W.; Lopaschuk, G.D. Role of fatty acid uptake and fatty acid beta-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim. Biophys. Acta 2010, 1801, 1–22. [Google Scholar] [CrossRef]
- Kurek, K.; Wiesiołek-Kurek, P.; Piotrowska, D.M.; Łukaszuk, B.; Chabowski, A.; Żendzian-Piotrowska, M. Inhibition of ceramide de novo synthesis with myriocin affects lipid metabolism in the liver or rats with streptozotocin-induced type 1 diabetes. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef]
- Zhang, A.Y.; Teggatz, E.G.; Zou, A.P.; Campbell, W.B.; Li, P.L. Endostatin uncouples NO and Ca2+ response to bradykinin through enhanced O2−-production in the intact coronary endothelium. Am. J. Physiol. Heart Circ. Physiol. 2005, 288, H686–H694. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ruiz, C.; Colell, A.; Mari, M.; Morales, A.; Fernandez-Checa, J.C. Direct effect of ceramide on the mitochondrial electron transport chain leads to generation of reactive oxygen species. Role of mitochondrial glutathione. J. Biol. Chem. 1997, 272, 11369–11377. [Google Scholar] [CrossRef] [Green Version]
- Hatanaka, Y.; Fujii, J.; Fukutomi, T.; Watanabe, T.; Che, W.; Sanada, Y.; Igarashi, Y.; Taniguchi, N. Reactive oxygen species enhances the induction of inducible nitric oxide synthase by sphingomyelinase in RAW264.7 cells. Biochim. Biophys. Acta 1998, 1393, 203–210. [Google Scholar] [CrossRef]
- Voehringer, D.W.; McConkey, D.J.; McDonnell, T.J.; Brisbay, S.; Meyn, R.E. Bcl-2 expression causes redistribution of gluthatione to the nucleus. Proc. Natl. Acad. Sci. USA 1998, 95, 2956–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimabukuro, M.; Wang, M.Y.; Zhou, Y.T.; Newgard, C.B.; Unger, R.H. Protection against lipoapoptosis of beta cells through leptin-dependent maintenance of Bcl-2 expression. Proc. Natl. Acad. Sci. USA 1998, 95, 9558–9561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fejfer, K.P.B.; Niczyporuk, M.; Ładny, J.R.; Hady, R.H.; Knaś, M.; Waszkiel, D.; Klimiuk, A.; Zalewska, A.; Maciejczyk, M. Oxidative modicication of biomolecules in the nonstiumulated and stimulated saliva of patients with morbid obesity treated with bariatric surgery. BioMed Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kołodziej, U.; Maciejczyk, M.; Miąsko, A.; Matczuk, J.; Knaś, M.; Żukowski, P.; Żendzian-Piotrowska, M.; Borys, J.; Zalewska, A. Oxidative modification in the salivary glands of high fat-diet induced insulin resistant rats. Front. Physiol. 2017, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Narotzki, B.; Reznick, A.Z.; Mitki, T.; Aizenbud, D.; Levy, Y. Enhanced cardiovascular risk and altered oxidative status in elders with moderate excessive body fat. Rejuvenation Res. 2014, 17, 334–340. [Google Scholar] [CrossRef]
- Zalewska, A.; Ziembicka, D.; Zendzian-Piotrowska, M.; Maciejczyk, M. The impact of high-fat diet on mitochondrial function, free radical production, and nitrosative stress in the salivary glands of Wistar rats. Oxid. Med. Cell. Longev. 2019. [Google Scholar] [CrossRef] [Green Version]
- Zalewska, A.; Knaś, M.; Żendzian-Piotrowska, M.; Waszkiewicz, N.; Szulimowska, J.; Prokopiuk, S.; Waszkiel, D.; Car, H. Antioxidant profile of salivary glands in high fat diet- induced insulin resistance rats. Oral. Dis. 2014, 20, 560–566. [Google Scholar] [CrossRef]
- Żebrowska-Gamdzyk, M.; Maciejczyk, M.; Zalewska, A.; Guzińska-Ustymowicz, K.; Tokajuk, A.; Car, H. Whey Protein Concentrate WPC-80 Intensifies Glycoconjugate Catabolism and Induces Oxidative Stress in the Liver of Rats. Nutrients 2018, 10, 1178. [Google Scholar] [CrossRef] [Green Version]
- Blachnio-Zabielska, A.U.; Persson, X.M.; Koutsari, C.Z.; Zabielski, P.; Jensen, M.D. A liquid chromatography/tandem mass spectrometry method for measuring the in vivo incorporation of plasma free fatty acids into intramyocellular ceramides in humans. Rapid Commun. Mass Spectrom. 2012, 26, 1134–1140. [Google Scholar] [CrossRef] [Green Version]
- Maciejczyk, M.; Kossakowska, A.; Szulimowska, J.; Klimiuk, A.; Knas, M.; Car, H.; Niklinska, W.; Ladny, J.R.; Chabowski, A.; Zalewska, A. Lysosomal exoglycosidase profile and secretory function in the salivary glands of rats with streptozotocin-induced diabetes. J. Diabetes Res. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borys, J.; Maciejczyk, M.; Antonowicz, B.; Kretowski, A.; Sidun, J.; Domel, E.; Dabrowski, J.R.; Ladny, J.R.; Morawska, K.; Zalewska, A. Glutathione metabolism, mitochondria activity, and nitrosative stress in patients treated for mandible fractures. J. Clin. Med. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Mansson-Rahemtulla, B.; Baldone, D.C.; Pruitt, K.M.; Rahemtulla, F. Specific assays for peroxidases in human saliva. Arch. Oral Biol. 1986, 31, 661–668. [Google Scholar] [CrossRef]
- Mize, C.E.; Langdon, R.G. Hepatic glutathione reductase I. Purification and general kinetic properties. J. Biol. Chem. 1962, 237, 1589–1595. [Google Scholar] [PubMed]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Kalousová, M.; Skrha, J.; Zima, T. Advanced glycation end-products and advanced oxidation protein products in patients with diabetes mellitus. Physiol. Res. 2002, 51, 597–604. [Google Scholar]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar]
- Nourooz-Zadeh, J.; Tajaddini-Sarmadi, J.; Wolff, S.P.; Birlouez-Aragon, I. Measurement of hydroperoxides in edible oils using the ferrous oxidation in xylenol orange assay. J. Agric. Food Chem. 1995, 43, 17–21. [Google Scholar] [CrossRef]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prajda, N.; Weber, G. Malignant transformation-linked imbalance: Decreased xanthine oxidase activity in hepatomas. FEBS Lett. 1975, 59, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Bondy, S.C.; Guo, S.X. Effect of ethanol treatment on indices of cumulative oxidative stress. Eur. J. Pharmacol. 1994, 270, 349–355. [Google Scholar] [CrossRef]
- Borys, J.; Maciejczyk, M.; Antonowicz, B.; Sidun, J.; Świderska, M.; Zalewska, A. Free radical production, inflammation and apoptosis in patients treated with titanium mandibular fixations—An observational study. Front. Immunol. 2019. [Google Scholar] [CrossRef]
- Meki, A.R.M.; Esmail, E.E.D.F.; Hussein, A.A.; Hassanein, H.M. Caspase-3 and heat shock protein-70 in rat liver treated with aflatoxin B1: Effect of melatonin. Toxicon 2004, 43, 93–100. [Google Scholar] [CrossRef]
- Janssen, A.J.M.; Trijbels, F.J.M.; Sengers, R.C.A.; Smeitink, J.A.M.; Van Den Heuvel, L.P.; Wintjes, L.T.M.; Stoltenborg-Hogenkamp, B.J.M.; Rodenburg, R.J.T. Spectrophotometric assay for complex I of the respiratory chain in tissue samples and cultured fibroblasts. Clin. Chem. 2007, 53, 729–734. [Google Scholar] [CrossRef]
- Rustin, P.; Chretien, D.; Bourgeron, T.; Gérard, B.; Rötig, A.; Saudubray, J.M.; Munnich, A. Biochemical and molecular investigations in respiratory chain deficiencies. Clin. Chim. Acta. 1994. [Google Scholar] [CrossRef]
- Wharton, D.C.; Tzagoloff, A. Cytochrome oxidase from beef heart mitochondria. Methods Enzymol. 1967. [Google Scholar] [CrossRef]
- Srere, P.A. Citrate synthase: [ec 4.1.3.7 citrate oxaloacetate-lyase (coa-acetylating)]. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1969; Volume 13, pp. 3–11. [Google Scholar] [CrossRef]
- Muller, F.L.; Liu, Y.; Van Remmen, H. Complex III releases superoxide to both sides of the inner mitochondrial membrane. J. Biol. Chem. 2004. [Google Scholar] [CrossRef] [Green Version]
- Ebertz, C.E.; Bonfleur, M.L.; Bertasso, I.M.; Mendes, M.C.; Lubaczeuski, C.; Araujo, A.C.; Paes, A.M.; Amorim, E.M.P.; Balbo, S.L. Duodenal jejunal bypass attenuates non-alcoholic fatty liver disease in western diet-obese rats. Acta Cir. Bras. 2014, 29, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Żukowski, P.M.M.; Matczuk, J.; Kurek, K.; Waszkiel, D.; Żendzian-Piotrowska, M.; Zalewska, A. Effect of N-acetylcysteine on antioxidant defense, oxidative modification, and salivary gland function in a rat model of insulin resistance. Oxid. Med. Cell. Longev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Veret, J.; Bellini, L.; Giussani, P.; Ng, C.; Magnan, C.; Le Stunff, H. Roles of sphingolipid metabolism in pancreatic beta cell dysfunction induced by lipotoxicity. J. Clin. Med. 2014, 3, 646–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veret, J.; Coant, N.; Berdyshev, E.V.; Skobeleva, A.; Therville, N.; Bailbe, D.; Gorshkova, I.; Natarajan, V.; Portha, B.; Le Stunff, H. Ceramide synthase 4 and de novo production of ceramides with specific N-acyl chain lengths are involved in glucolipotoxicity-induced apoptosis of INS-1 β-cells. Biochem. J. 2011, 438, 177–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: Evidence and mechanisms. Lipids Health Dis. 2013, 12, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhunia, A.K.; Han, H.; Snowden, A.; Chatterjee, S. Redox-regulated signaling by lactosylceramide in the proliferation of human aortic smooth muscle cells. J. Biol. Chem. 1997, 272, 15642–15649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Block, K.; Gorin, Y.; Abboud, H.E. Subcellular localization of NOX4 and regulation in diabetes. Proc. Natl. Acad. Sci. USA 2009, 106, 14385–14390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takac, I.; Schroder, K.; Zhang, L.; Lardy, B.; Anilkumar, N.; Lambeth, J.D.; Shah, A.M.; Morel, F.; Brandes, R.P. The e-loop is involved in hydrogen peroxide formation by the NADPH oxidase NOX 4. J. Biol. Chem. 2011, 286, 13304–13313. [Google Scholar] [CrossRef] [Green Version]
- de la Cal, C.; Lomniczi, A.; Mohn, C.E.; De Laurentiis, A.; Casal, M.; Chiarenza, A.; Paz, D.; McCann, S.M.; Rettori, V.; Elverdin, J.C. Decrease in salivary secretion by radiation mediated by nitric oxide and prostaglandins. Neuroimmunomodulation 2006, 13, 19–27. [Google Scholar] [CrossRef]
- Gudz, T.I.; Tsering, K.Y.; Hoppel, C.L. Direct inhibition of mitochondrial respiratory chain complex III by cell-permeable ceramide. J. Biol. Chem. 1997, 272, 24154–24158. [Google Scholar] [CrossRef] [Green Version]
- Di Paola, M.; Cocco, T.; Lorusso, M. Ceramide interaction with the respiratory chain of heart mitochondria. Biochemistry 2000, 39, 6660–6668. [Google Scholar] [CrossRef]
- Zigdon, H.; Kogot-Levin, A.; Park, J.W.; Goldschmidt, R.; Kelly, S.; Merrill, A.H., Jr.; Scherz, A.; Pewzner-Jung, Y.; Saada, A.; Futerman, A.H. Ablation of ceramide synthase 2 causes chronic oxidative stress due to disruption of the mitochondrial respiratory chain. J. Biol. Chem. 2013, 288, 4947–4956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Mileykovskaya, E.; Dowhan, W. Gluing the respiratory chain together. Cardiolipin is required for supercomplex formation in the inner mitochondrial membrane. J. Biol. Chem. 2002, 277, 43553–43556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kota, V.; Szulc, Z.M.; Hama, H. Identification of C6-ceramide-interacting proteins in D6P2T Schwannoma cells. Proteomics 2012, 12, 2179–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, C.; Morales, A.; Colell, A.; Rodes, J.; Yi, J.R.; Kaplowitz, N.; Fernandez-Checa, J.C. Evidence that the rat hepatic mitochondrial carrier is distinct from the sinusoidal and canalicular transporters for reduced glutathione. Expression studies in xenopus laevis oocytes. J. Biol. Chem. 1995, 270, 15946–15949. [Google Scholar] [CrossRef] [Green Version]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Żendzian-Piotrowska, M.; Łukaszuk, B.; Maciejczyk, M.; Ostrowska, L.; Zalewska, A.; Chabowski, A.; Kurek, K. High-fat, high-protein, and high-carbohydrate diets affect sphingolipid profile in pancreatic steatosis in Wistar rats. Nutrition 2019. [Google Scholar] [CrossRef]
- Hannun, Y.A. Functions of ceramide in coordinating cellular responses to stress. Science 1996, 274, 1855–1859. [Google Scholar] [CrossRef]
- Joseph, C.K.; Wright, S.D.; Bornmann, W.G.; Randolph, J.T.; Kumar, E.R.; Bittman, R.; Liu, J.; Kolesnick, R.N. Bacterial lipopolysaccharide has structural similarity to ceramide and stimulates ceramide-activated protein kinase in myeloid cells. J. Biol. Chem. 1994, 269, 17606–17610. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Control Diet (g) | HFD (g) | |
|---|---|---|---|
| Protein | Casein, lactic, 30 mesh | 200.00 | 200.00 |
| Cystine, L | 3.00 | 3.00 | |
| Carbohydrate | Starch, corn | 506.20 | 0.00 |
| Lodex 10 | 125.00 | 125.00 | |
| Sucrose, fine granulated | 72.80 | 72.80 | |
| Fiber | Solka floc, FCC200 | 50.00 | 50.00 |
| Fat | Soybean oil, USP | 25.00 | 25.00 |
| Lard | 20.00 | 245.00 | |
| Mineral | S10026B | 50.00 | 50.00 |
| Vitamin | Choline bitartrate | 2.00 | 2.00 |
| V10001C | 1.00 | 1.00 |
| Control Diet (mg/g) | HFD (mg/g) | |
|---|---|---|
| C14:0, Myristic | 2.20 ± 0.20 | 4.43 ± 0.24 * |
| C16:0, Palmitic | 9.21 ± 1.56 | 58.25 ± 1.10 * |
| C16:1, Palmitoleic | 0.55 ± 0.12 | 4.84 ± 0.09 * |
| C18:0, Stearic | 3.55 ± 0.60 | 27.73 ± 0.43 * |
| C18:1, Oleic | 13.50 ± 2.24 | 90.83 ± 5.04 * |
| C18:2, Linoleic | 12.42 ± 2.06 | 50.36 ± 1.04 * |
| C18:3, Linolenic | 0.52 ± 0.09 | 2.91 ± 0.06 * |
| C20:0, Arachidic | 0.11 ± 0.02 | 0.41 ± 0.01 * |
| C20:4, Arachidonic | 0.05 ± 0.00 | 0.43 ± 0.01 * |
| C20:5, EPA | 0.00 ± 0.00 | 0.03 ± 0.01 * |
| C22:0, Behenic | 0.03 ± 0.00 | 0.07 ± 0.01 * |
| C22:6, DHA | 0.01 ± 0.01 | 0.06 ± 0.01 * |
| C24:0, Lignoceric | 0.04 ± 0.00 | 0.05 ± 0.00 |
| C24:1 Selacholeic | 0.00 ± 0.00 | 0.03 ± 0.00 * |
| Total | 42.19 ± 6.85 | 240.43 ± 7.93 * |
| C (n = 10) | HFD (n = 10) | p | |
|---|---|---|---|
| Body weight (g) | 28.75 ± 0.51 | 37.88 ± 0.63 | <0.001 |
| Left salivary gland weight (g) | 0.092 ± 0.01 | 0.097 ± 0.01 | NS |
| Right salivary gland weight (g) | 0.091 ± 0.002 | 0.093 ± 0.003 | NS |
| Left salivary gland index (%) | 0.32 ± 0.03 | 0.25 ± 0.03 | NS |
| Right salivary gland index (%) | 0.32 ± 0.03 | 0.26 ± 0.03 | NS |
| Glucose (mg/dL) | 101.5 ± 3.79 | 173.1 ± 8.77 | <0.001 |
| Insulin (ng/mL) | 0.83 ± 0.02 | 1.6 ± 0.04 | <0.001 |
| HOMA-IR | 3.34 ± 0.17 | 10.93 ± 0.57 | <0.001 |
| Total protein (μg/mL) | 4004.5 ± 229 | 3671 ± 483 | 0.04 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalewska, A.; Maciejczyk, M.; Szulimowska, J.; Imierska, M.; Błachnio-Zabielska, A. High-Fat Diet Affects Ceramide Content, Disturbs Mitochondrial Redox Balance, and Induces Apoptosis in the Submandibular Glands of Mice. Biomolecules 2019, 9, 877. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120877
Zalewska A, Maciejczyk M, Szulimowska J, Imierska M, Błachnio-Zabielska A. High-Fat Diet Affects Ceramide Content, Disturbs Mitochondrial Redox Balance, and Induces Apoptosis in the Submandibular Glands of Mice. Biomolecules. 2019; 9(12):877. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120877
Chicago/Turabian StyleZalewska, Anna, Mateusz Maciejczyk, Julita Szulimowska, Monika Imierska, and Agnieszka Błachnio-Zabielska. 2019. "High-Fat Diet Affects Ceramide Content, Disturbs Mitochondrial Redox Balance, and Induces Apoptosis in the Submandibular Glands of Mice" Biomolecules 9, no. 12: 877. https://0-doi-org.brum.beds.ac.uk/10.3390/biom9120877