
Liver-Targeted Nanoparticles Facilitate the Bioavailability and Anti-HBV Efficacy of Baicalin In Vitro and In Vivo

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
2.3. Preparation of Liver-Targeted Baicalin Liposomes (BAA1)
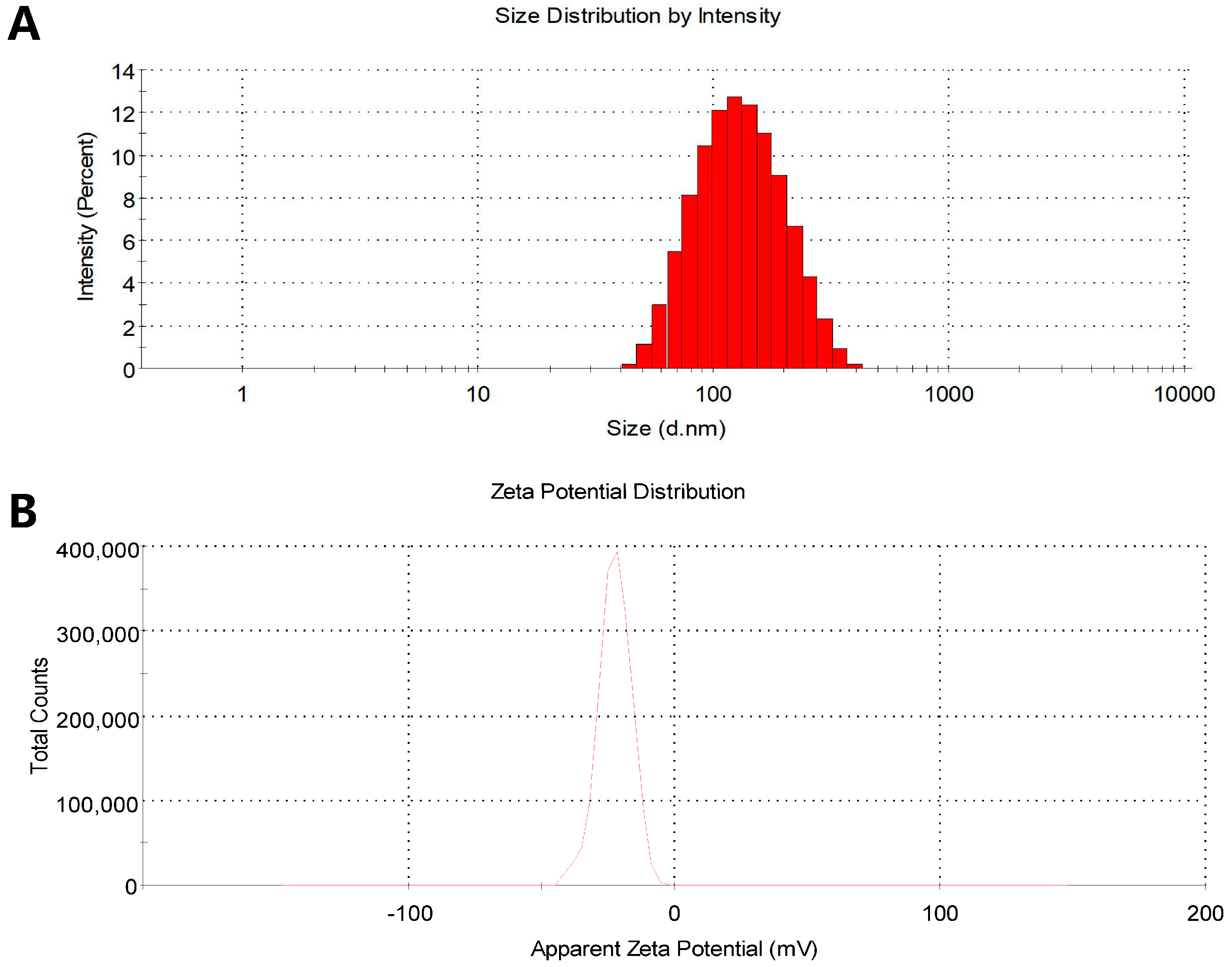
2.4. Physicochemical Characterization of BAA1
2.5. The Entrapment Efficiency of BAA1
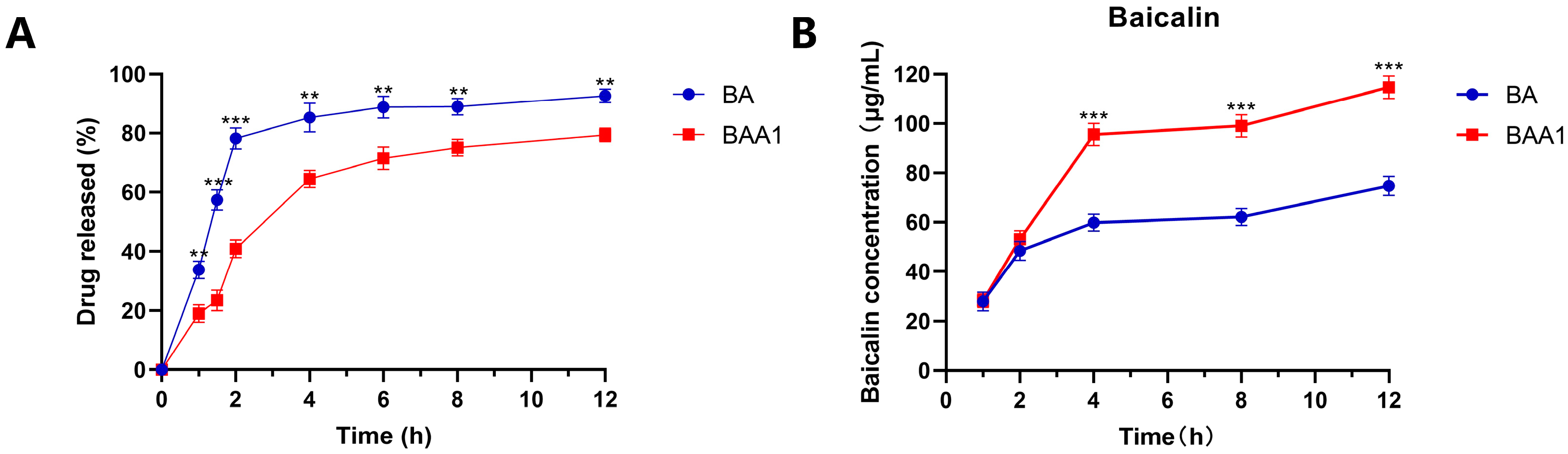
2.6. In Vitro Release Studies of Liposomes
2.7. Cell Culture and HBV Transfection
2.8. Cell Counting Kit-8 (CCK-8) Assay
2.9. HBsAg and HBeAg Assay In Vitro and In Vivo
2.10. Quantitative RT-PCR (qRT-PCR)
2.11. Western Blotting
2.12. Co-Immunoprecipitation (Co-IP)
2.13. Cellular Uptake of BA
2.14. HBV Infection and Mice Treatment
2.15. Pharmacokinetics and Tissue Distribution Studies
2.16. Transcriptomics Analysis
2.17. Statistical Analyses
3. Results
3.1. Characterization of BAA1
3.2. In Vitro Release and Cellular Uptake of BAA1
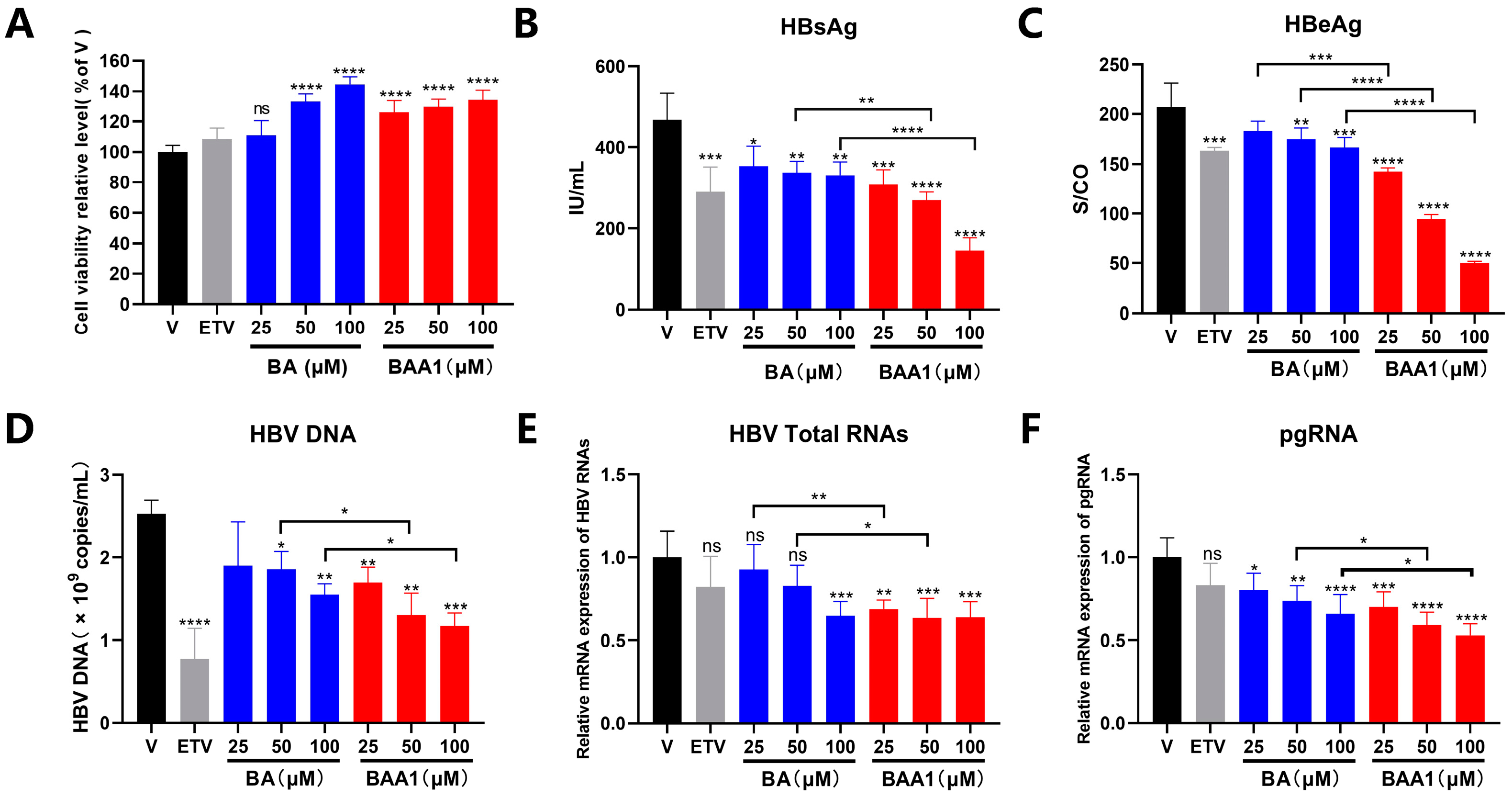
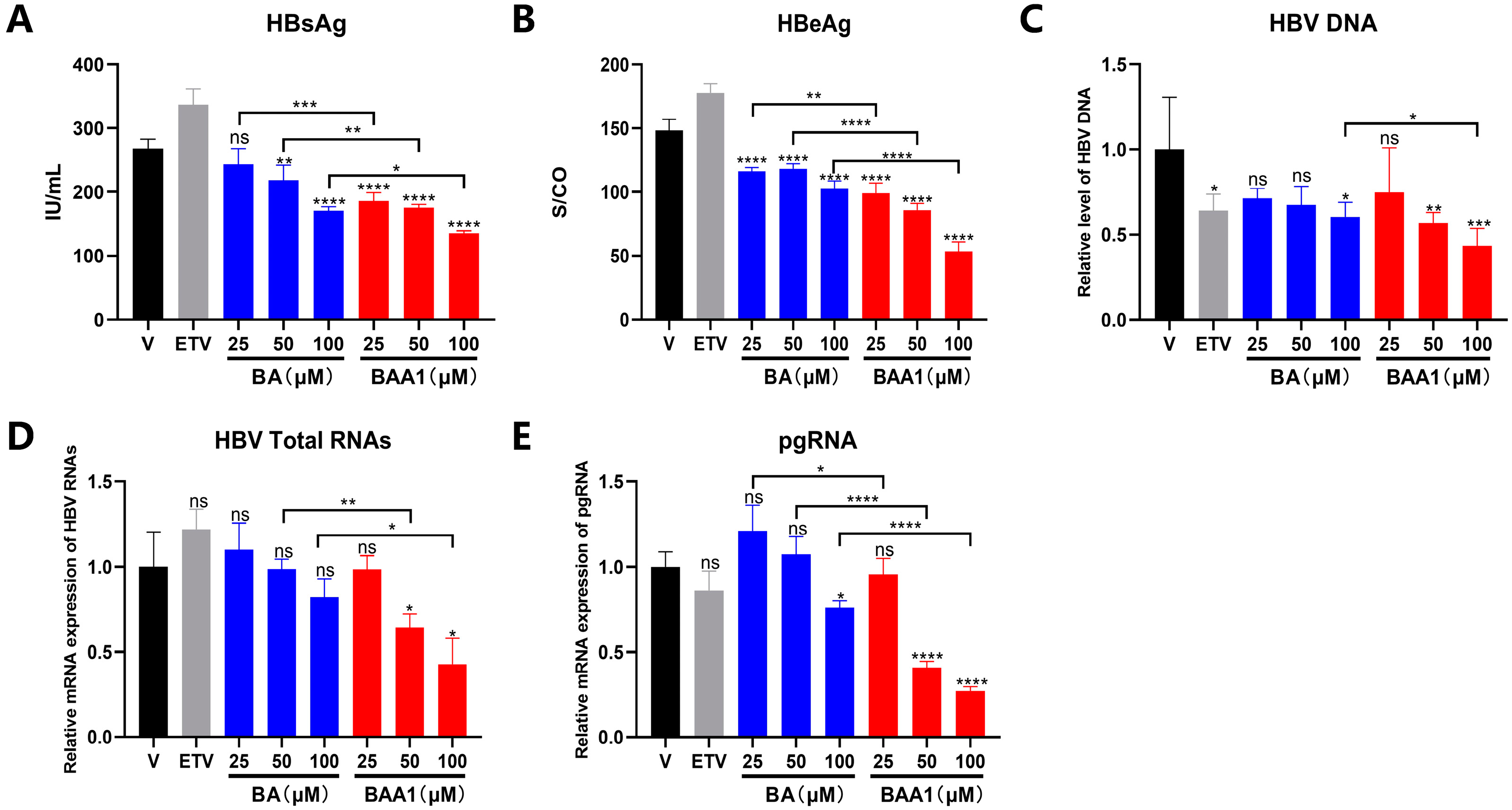
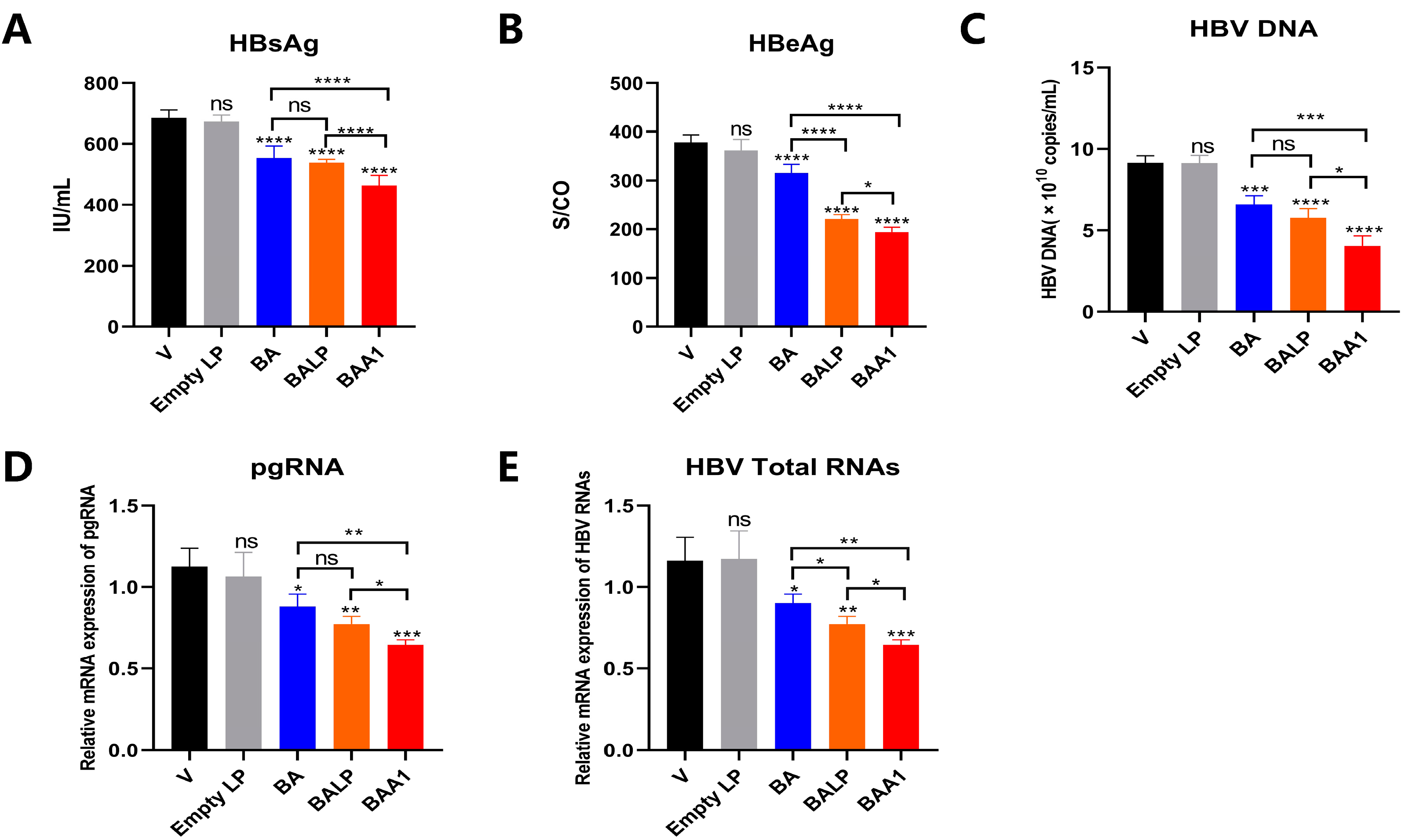
3.3. The Potent Inhibition of HBV Replication and Transcription by BAA1 in pHBV1.2 and rAAV-HBV-Transfected HepG2 Cells
3.4. Mediation of Enhanced Anti-HBV Activity of BA Liposomes by ApoA1 Modification
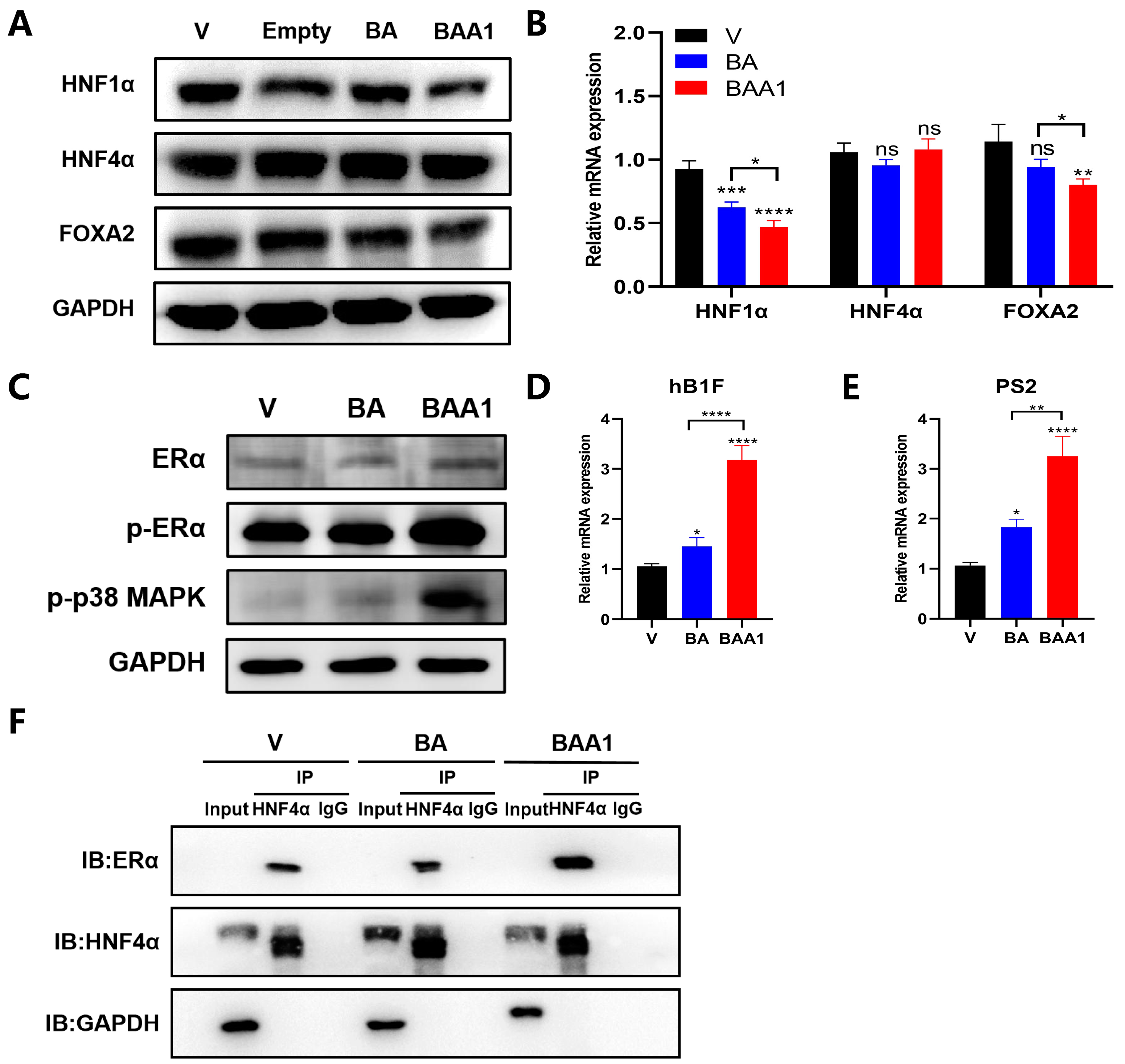
3.5. Augmentation of the ERα–HNF4α–HNF1α Signaling Axis by BAA1
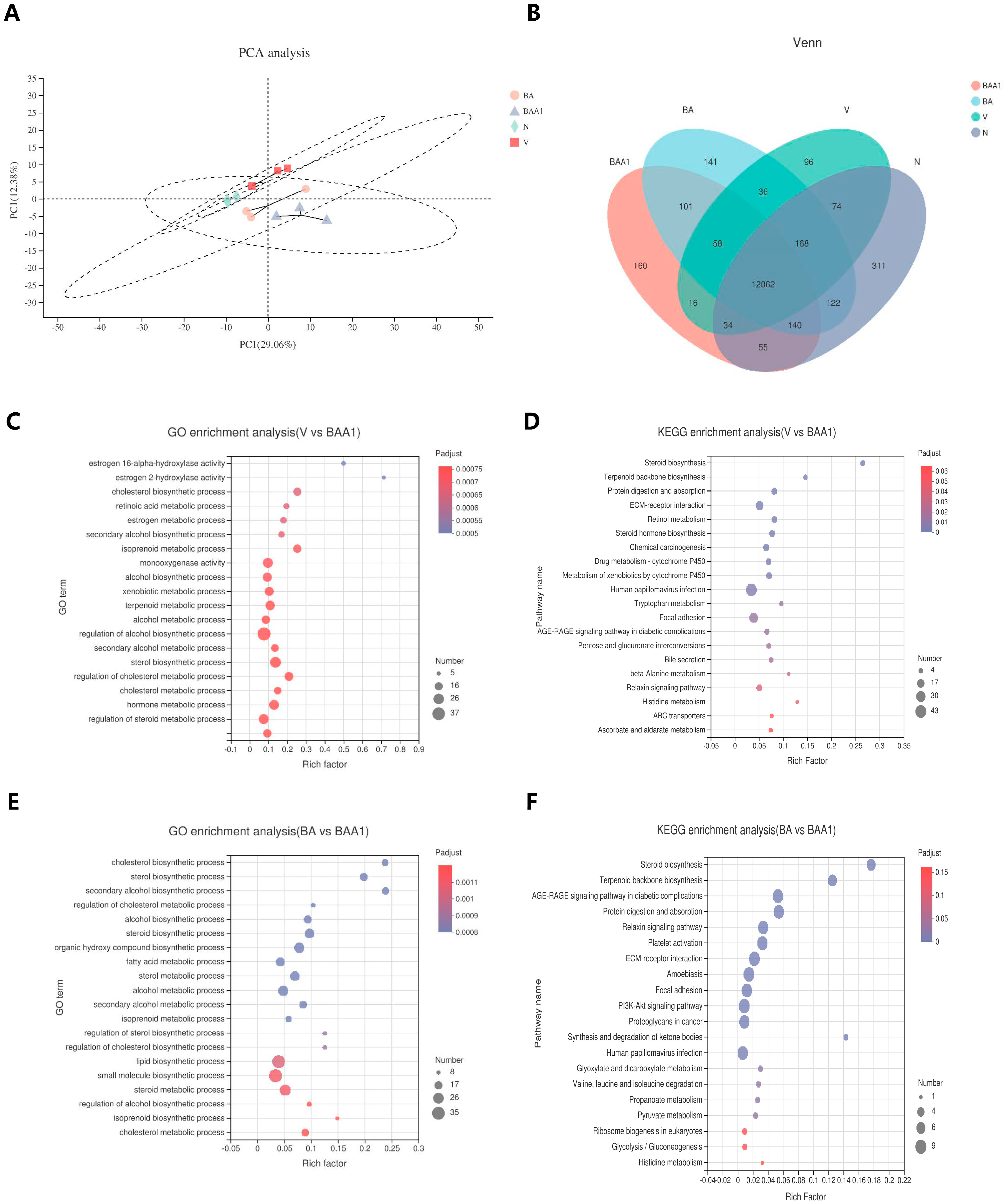
3.6. Transcriptomic Analysis
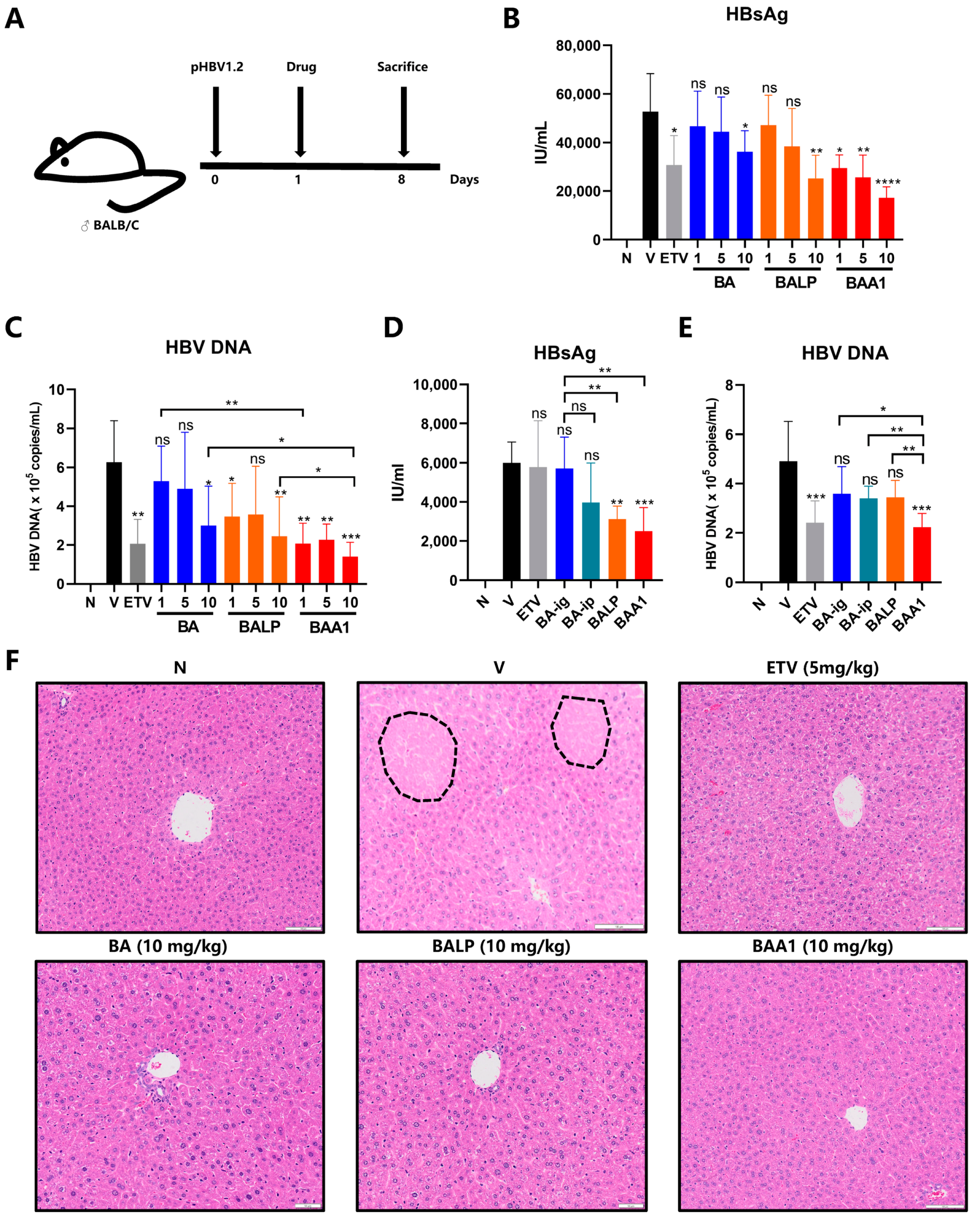
3.7. Improvement of the Anti-HBV Efficacy of BA Liposomes In Vivo by ApoA1 Modification
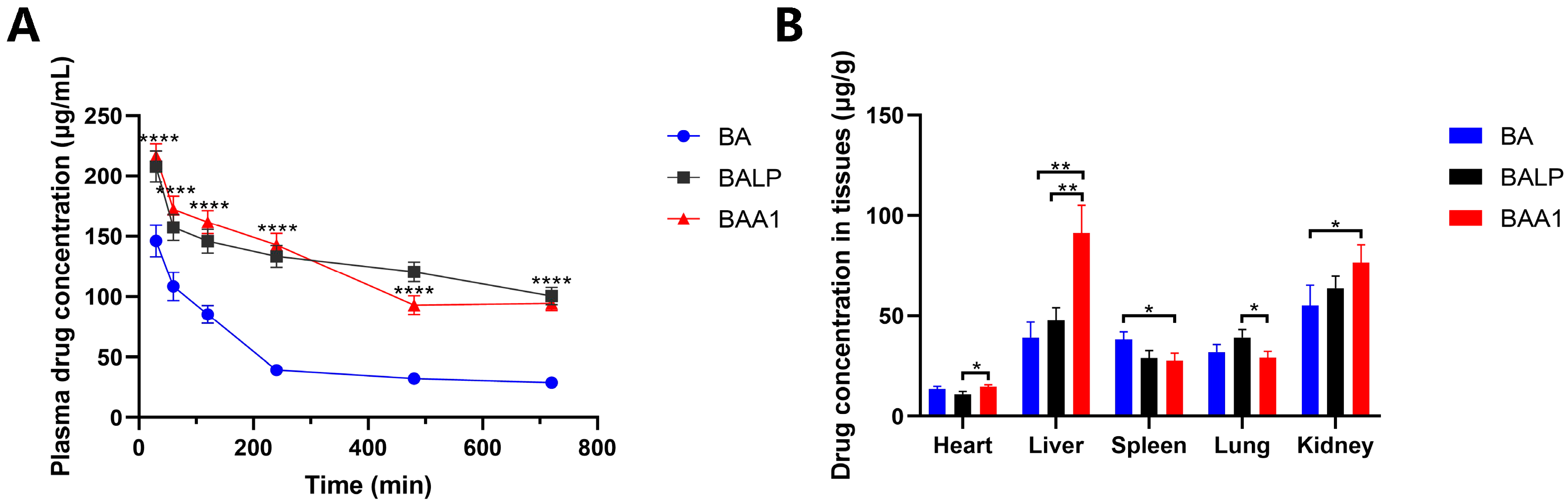
3.8. Pharmacokinetics and Tissue Distribution Study of BAA1 In Vivo
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Trépo, C.; Chan, H.L.Y.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- Razavi-Shearer, D.; Gamkrelidze, I.; Nguyen, M.H. Global prevalence, treatment, and prevention of hepatitis B virus infection in 2016: A modelling study. Lancet Gastroenterol. Hepatol. 2018, 3, 383–403. [Google Scholar] [CrossRef]
- Tu, T.; Douglas, M.W. Hepatitis B Virus Infection: From Diagnostics to Treatments. Viruses 2020, 12, 1366. [Google Scholar] [CrossRef]
- Gane, E.J. Future anti-HBV strategies. Liver Int. 2017, 37 (Suppl. 1), 40–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Chen, X.-Y.; Martin, C. Scutellaria baicalensis, the golden herb from the garden of Chinese medicinal plants. Sci. Bull. 2016, 61, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.Y.; Li, M.; Zhang, C.L.; Liu, D. Pharmacological properties of baicalin on liver diseases: A narrative review. Pharmacol. Rep. 2021, 73, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Tang, W.; Geng, P.; Zhu, H.; Zhou, W.; Huang, H.; Zhou, P.; Shi, X. Baicalin down-regulating hepatitis B virus transcription depends on the liver-specific HNF4alpha-HNF1alpha axis. Toxicol. Appl. Pharmacol. 2020, 403, 115131. [Google Scholar] [CrossRef]
- Xing, J.; Chen, X.; Zhong, D. Absorption and enterohepatic circulation of baicalin in rats. Life Sci. 2005, 78, 140–146. [Google Scholar] [CrossRef]
- Long, Y.; Xiang, Y.; Liu, S.; Zhang, Y.; Wan, J.; Yang, Q.; Cui, M.; Ci, Z.; Li, N.; Peng, W. Baicalin Liposome Alleviates Lipopolysaccharide-Induced Acute Lung Injury in Mice via Inhibiting TLR4/JNK/ERK/NF-kappaB Pathway. Mediat. Inflamm. 2020, 2020, 8414062. [Google Scholar] [CrossRef]
- Tang, H.-X.; Zhao, T.-W.; Zheng, T.; Sheng, Y.-J.; Zheng, H.-S.; Zhang, Y.-S. Liver-targeting liposome drug delivery system and its research progress in liver diseases. World Chin. J. Dig. 2016, 24, 4238–4246. [Google Scholar] [CrossRef]
- Asselah, T.; Loureiro, D.; Boyer, N.; Mansouri, A. Targets and future direct-acting antiviral approaches to achieve hepatitis B virus cure. Lancet Gastroenterol. Hepatol. 2019, 4, 883–892. [Google Scholar] [CrossRef]
- Wang, Y.; Hao, J.; Liu, X.; Wang, H.; Zeng, X.; Yang, J.; Li, L.; Kuang, X.; Zhang, T. The mechanism of apoliprotein A1 down-regulated by Hepatitis B virus. Lipids Health Dis. 2016, 15, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.I.; Shin, D.; Choi, T.H.; Lee, J.C.; Cheon, G.J.; Kim, K.Y.; Park, M.; Kim, M. Systemic and specific delivery of small interfering RNAs to the liver mediated by apolipoprotein A-I. Mol. Ther. 2007, 15, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Wang, W.; Wang, B.; Zhu, H.; Zhang, B.; Feng, M. Delivery of hydrophilic drug doxorubicin hydrochloride-targeted liver using apoAI as carrier. J. Drug Target. 2013, 21, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Guo, J.; Zheng, X.; Wu, J.; Zhou, Y.; Yu, Y.; Ye, Y.; Zhang, L.; Zhao, L. Preparation, pharmacokinetics and biodistribution of baicalin-loaded liposomes. Int. J. Nanomed. 2014, 9, 3623–3630. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Minh, L.V.; Liu, J.; Angelov, B.; Drechsler, M.; Garamus, V.M.; Willumeit-Römer, R.; Zou, A. Baicalin loaded in folate-PEG modified liposomes for enhanced stability and tumor targeting. Colloids Surf. B Biointerfaces 2016, 140, 74–82. [Google Scholar] [CrossRef]
- Huang, H.; Zhou, W.; Zhu, H.; Zhou, P.; Shi, X. Baicalin benefits the anti-HBV therapy via inhibiting HBV viral RNAs. Toxicol. Appl. Pharmacol. 2017, 323, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Yang, X.; Shang, X.; Gao, Z.; Li, Q.; Hong, Y.; Wu, J.; Meng, T.; Yuan, H.; Hu, F. Hepatocyte-targeting and microenvironmentally responsive glycolipid-like polymer micelles for gene therapy of hepatitis B. Mol. Ther. Nucleic Acids 2021, 24, 127–139. [Google Scholar] [CrossRef]
- Sun, C.; Lan, P.; Han, Q.; Huang, M.; Zhang, Z.; Xu, G.; Song, J.; Wang, J.; Wei, H.; Zhang, J.; et al. Oncofetal gene SALL4 reactivation by hepatitis B virus counteracts miR-200c in PD-L1-induced T cell exhaustion. Nat. Commun. 2018, 9, 1241. [Google Scholar] [CrossRef] [Green Version]
- Xiang, C.; Du, Y.; Meng, G.; Soon Yi, L.; Sun, S.; Song, N.; Zhang, X.; Xiao, Y.; Wang, J.; Yi, Z.; et al. Long-term functional maintenance of primary human hepatocytes in vitro. Science 2019, 364, 399–402. [Google Scholar] [CrossRef]
- Shi, X.; Zhou, W.; Huang, H.; Zhu, H.; Zhou, P.; Zhu, H.; Ju, D. Inhibition of the inflammatory cytokine tumor necrosis factor-alpha with etanercept provides protection against lethal H1N1 influenza infection in mice. Crit. Care 2013, 17, R301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Liu, X.; Yang, H.; Zhao, R.; Liu, C.; Zhang, R.; Zhang, Q. Comparative pharmacokinetic study on phenolic acids and flavonoids in spinal cord injury rats plasma by UPLC-MS/MS after single and combined oral administration of danshen and huangqin extract. J. Pharm. Biomed. Anal. 2019, 172, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; An, R.; Du, G.; Liang, K.; Li, G. Validation of an LC-MS/MS method for simultaneous detection of diverse components of Qinxing Qingre Zhike Granule in rat plasma and its application to pharmacokinetic study after oral administration to rats. Biomed. Chromatogr. 2019, 33, e4524. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Xiao, J.; Xu, R.; Wang, Q.; Wang, X. Simultaneous determination of baicalin, baicalein, wogonoside, wogonin, scutellarin, berberine, coptisine, ginsenoside Rb1 and ginsenoside Re of Banxia xiexin decoction in rat plasma by LC-MS/MS and its application to a pharmacokinetic study. Biomed. Chromatogr. 2018, 32, e4083. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, H.T.; Tsai, H.F.; Liao, H.J.; Lin, Y.J.; Chen, L.; Chen, P.J.; Hsu, P.N. PD-1 blockage reverses immune dysfunction and hepatitis B viral persistence in a mouse animal model. PLoS ONE 2012, 7, e39179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Yuan, Y.; Gong, X.; Zhang, L.; Zhou, Q.; Wu, S.; Zhang, X.; Hu, J.; Kuang, G.; Yin, X.; et al. Baicalin and its nanoliposomes ameliorates nonalcoholic fatty liver disease via suppression of TLR4 signaling cascade in mice. Int. Immunopharmacol. 2020, 80, 106208. [Google Scholar] [CrossRef]
- Lin, C.L.; Kao, J.H. Natural history of acute and chronic hepatitis B: The role of HBV genotypes and mutants. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 249–255. [Google Scholar] [CrossRef]
- Huang, T.; Liu, Y.; Zhang, C. Pharmacokinetics and Bioavailability Enhancement of Baicalin: A Review. Eur. J. Drug Metab. Pharmacokinet. 2019, 44, 159–168. [Google Scholar] [CrossRef]
- Li, K.; Liang, Y.; Cheng, A.; Wang, Q.; Li, Y.; Wei, H.; Zhou, C.; Wan, X. Antiviral Properties of Baicalin: A Concise Review. Rev. Bras. Farmacogn. 2021, 31, 408–419. [Google Scholar] [CrossRef]
- Oo, A.; Teoh, B.T.; Sam, S.S.; Bakar, S.A.; Zandi, K. Baicalein and baicalin as Zika virus inhibitors. Arch. Virol. 2019, 164, 585–593. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Spagnuolo, C.; Tedesco, I.; Russo, G.L. Roles of flavonoids against coronavirus infection. Chem. Biol. Interact. 2020, 328, 109211. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liang, J.; Zheng, X.; Pi, C.; Liu, H.; Yang, H.; Zou, Y.; Ye, Y.; Zhao, L. Lung-targeting drug delivery system of baicalin-loaded nanoliposomes: Development, biodistribution in rabbits, and pharmacodynamics in nude mice bearing orthotopic human lung cancer. Int. J. Nanomed. 2017, 12, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Zhou, X.; Jia, H.; Chen, C.; Zhao, W.; Zhang, J.; Zhang, J.; Tong, S. Stronger enhancer II/core promoter activities of hepatitis B virus isolates of B2 subgenotype than those of C2 subgenotype. Sci. Rep. 2016, 6, 30374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Zhao, C.; Zhu, H.; She, W.; Zhang, J.; Liu, J.; Li, L.; Zheng, S.; Wen, Y.; Xie, Y. Hepatitis B virus (HBV) surface antigen interacts with and promotes cyclophilin a secretion: Possible link to pathogenesis of HBV infection. J. Virol. 2010, 84, 3373–3381. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.H.; Yeh, S.H.; Lin, W.H.; Yeh, K.H.; Yuan, Q.; Xia, N.S.; Chen, D.S.; Chen, P.J. Estrogen receptor α represses transcription of HBV genes via interaction with hepatocyte nuclear factor 4α. Gastroenterology 2012, 142, 989–998. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Units | BA | BALP | BAA1 |
|---|---|---|---|---|
| AUC0-t | mg/L*min | 32,446.896 ± 2833.222 | 87,550.161 ± 4794.225 | 84,249.757 ± 3453.486 |
| AUC0-∞ | mg/L*min | 45,990.555 ± 13,235.327 | 246,086.047 ± 17,232.716 | 157,887.132 ± 33,642.481 |
| MRT0-t | min | 283.610 ± 2.9314 | 347.976 ± 2.102 | 325.323 ± 2.277 |
| MRT0-∞ | min | 689.113 ± 211.102 | 1606.579 ± 85.404 | 999.485 ± 103.403 |
| t1/2z | min | 411.193 ± 242.653 | 1093.433 ± 52.416 | 601.197 ± 152.636 |
| Vz/F | L/kg | 1.196 ± 0.360 | 0.643 ± 0.033 | 0.543 ± 0.032 |
| CLz/F | L/min/kg | 0.002 | 0.001 | 0.001 |
| Cmax | mg/L | 108.451 ± 9.493 | 157.420 ± 8.945 | 173.132 ± 8.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Niu, Y.; Ai, X.; Xia, C.; Geng, P.; Zhu, H.; Zhou, W.; Huang, H.; Shi, X. Liver-Targeted Nanoparticles Facilitate the Bioavailability and Anti-HBV Efficacy of Baicalin In Vitro and In Vivo. Biomedicines 2022, 10, 900. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040900
Xu W, Niu Y, Ai X, Xia C, Geng P, Zhu H, Zhou W, Huang H, Shi X. Liver-Targeted Nanoparticles Facilitate the Bioavailability and Anti-HBV Efficacy of Baicalin In Vitro and In Vivo. Biomedicines. 2022; 10(4):900. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040900
Chicago/Turabian StyleXu, Weiming, Yijun Niu, Xin Ai, Chengjie Xia, Ping Geng, Haiyan Zhu, Wei Zhou, Hai Huang, and Xunlong Shi. 2022. "Liver-Targeted Nanoparticles Facilitate the Bioavailability and Anti-HBV Efficacy of Baicalin In Vitro and In Vivo" Biomedicines 10, no. 4: 900. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040900