
Sex-Specific Patterns of Diaphragm Phospholipid Content and Remodeling during Aging and in a Model of SELENON-Related Myopathy
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
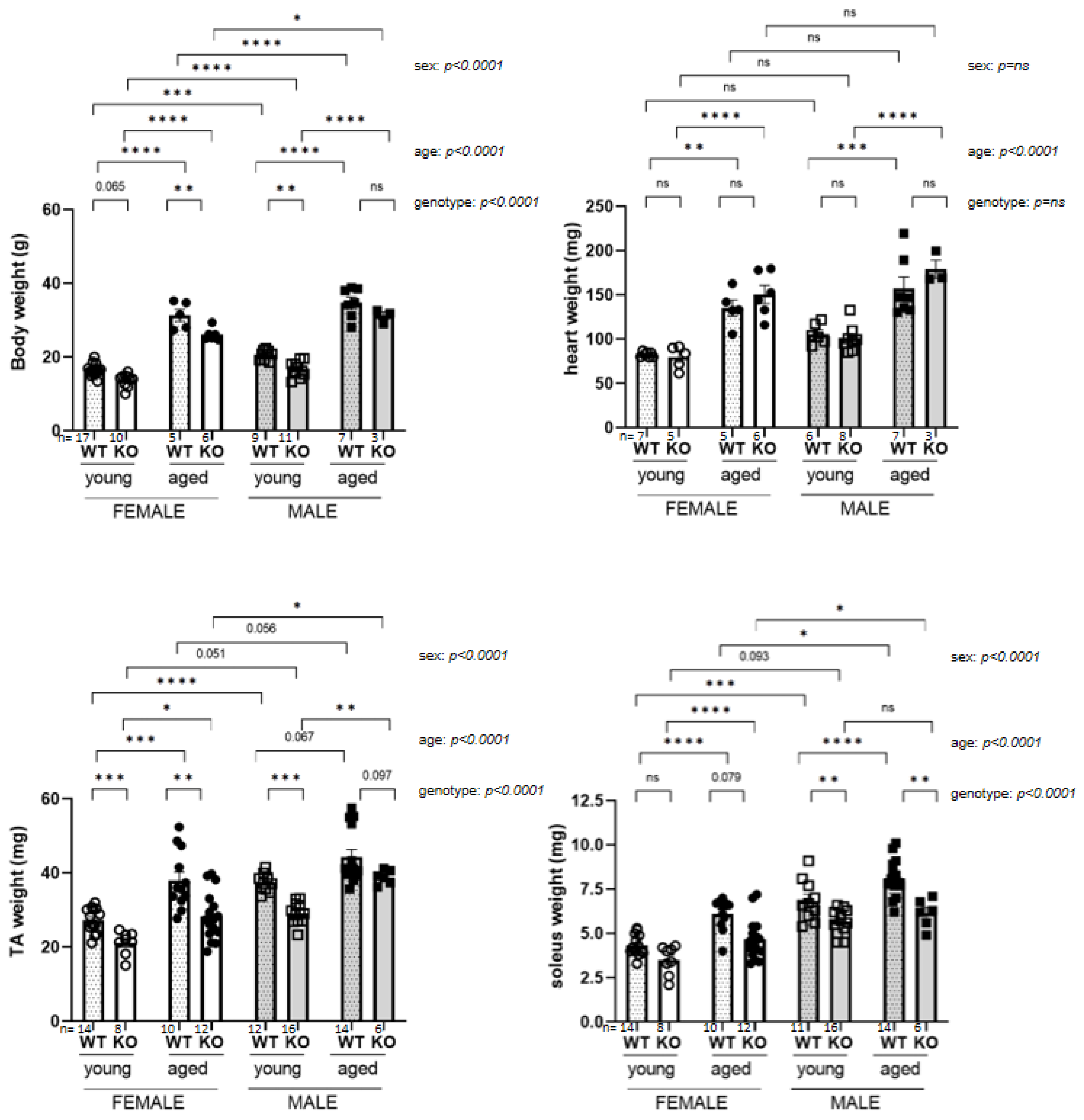
3.1. Anatomical Parameters Influenced by Age, Sex and Genotype
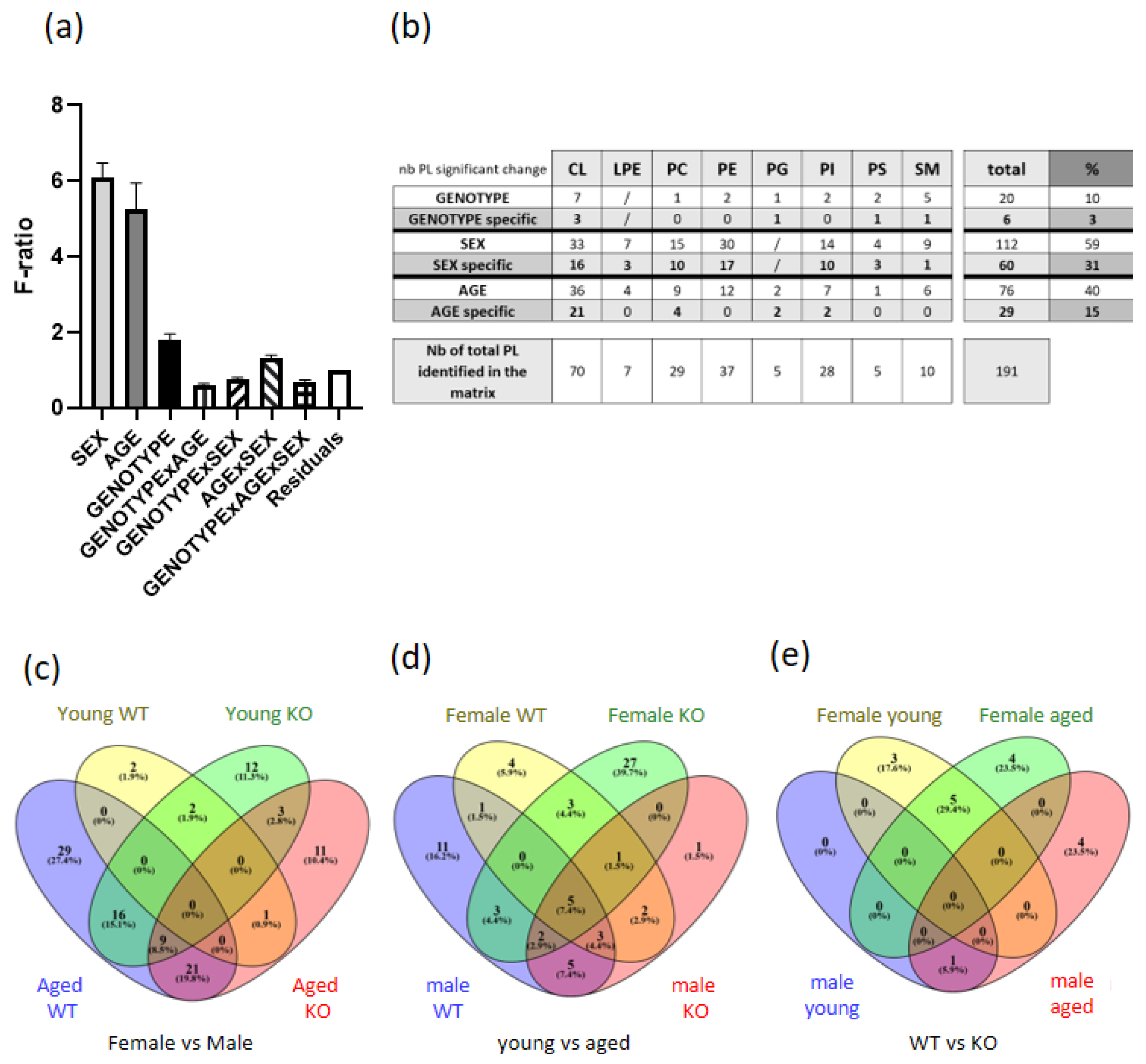
3.2. Phospholipidomic Analysis Revealed an Important Sex Difference in Diaphragm PL Content
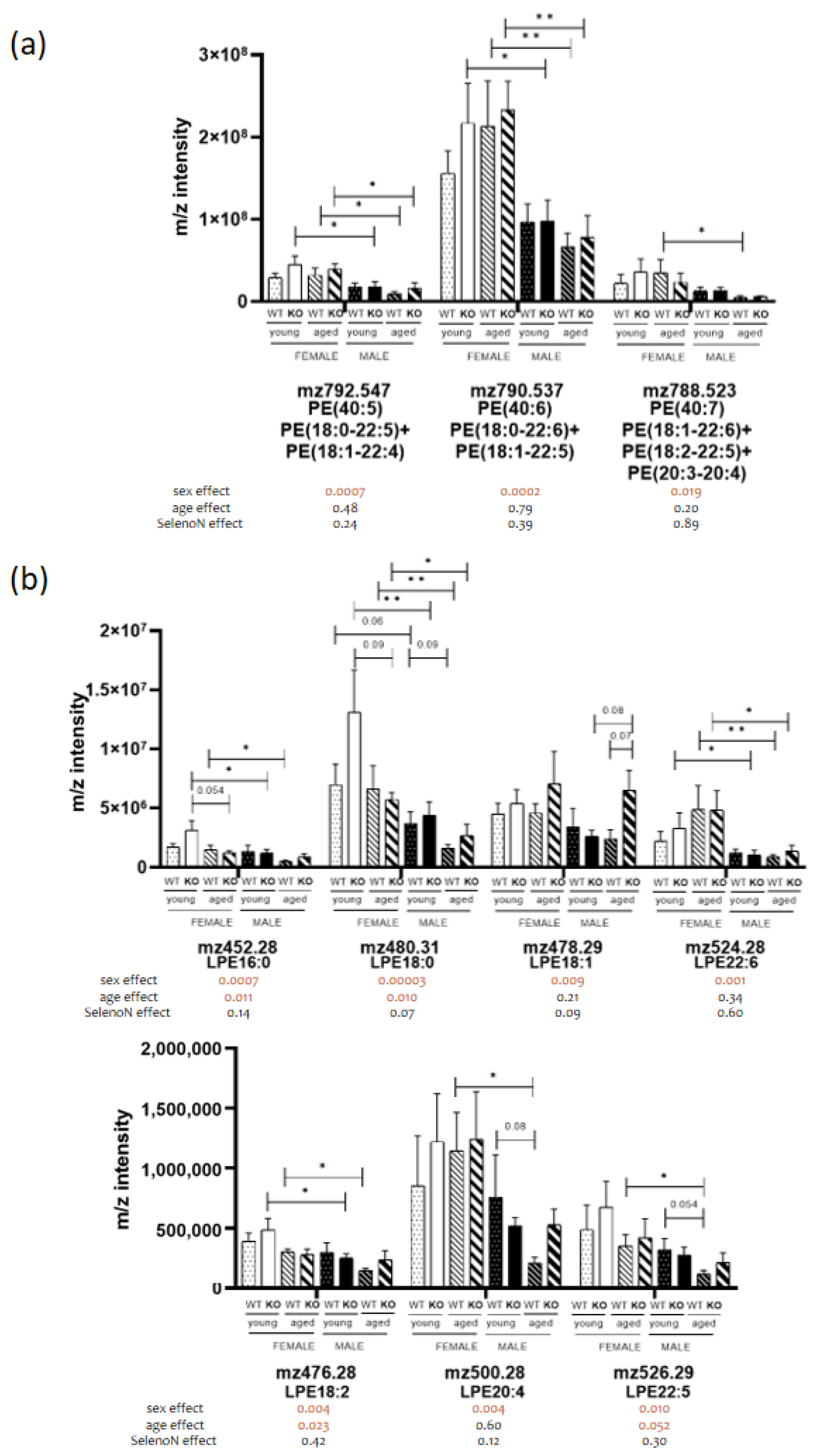
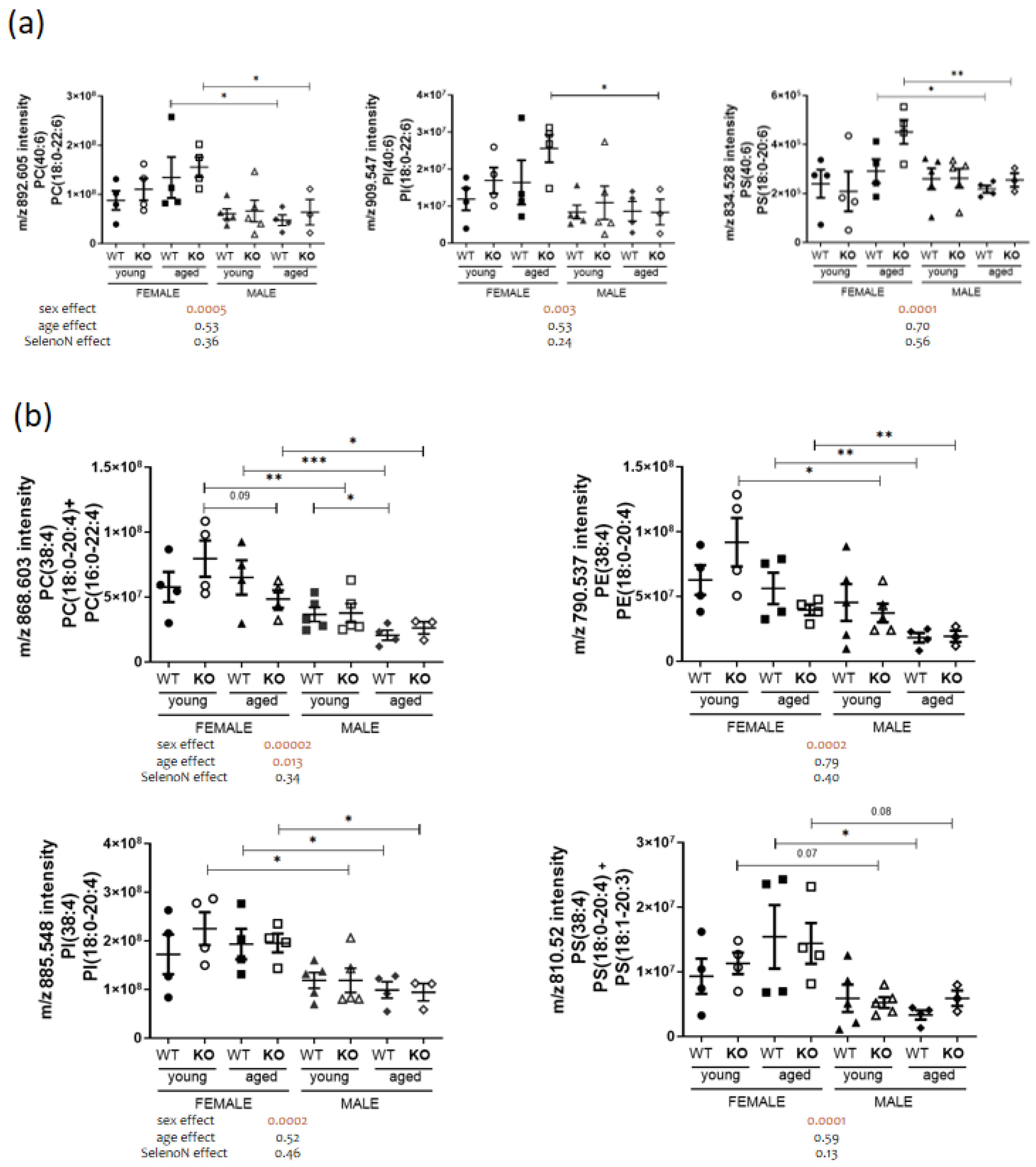
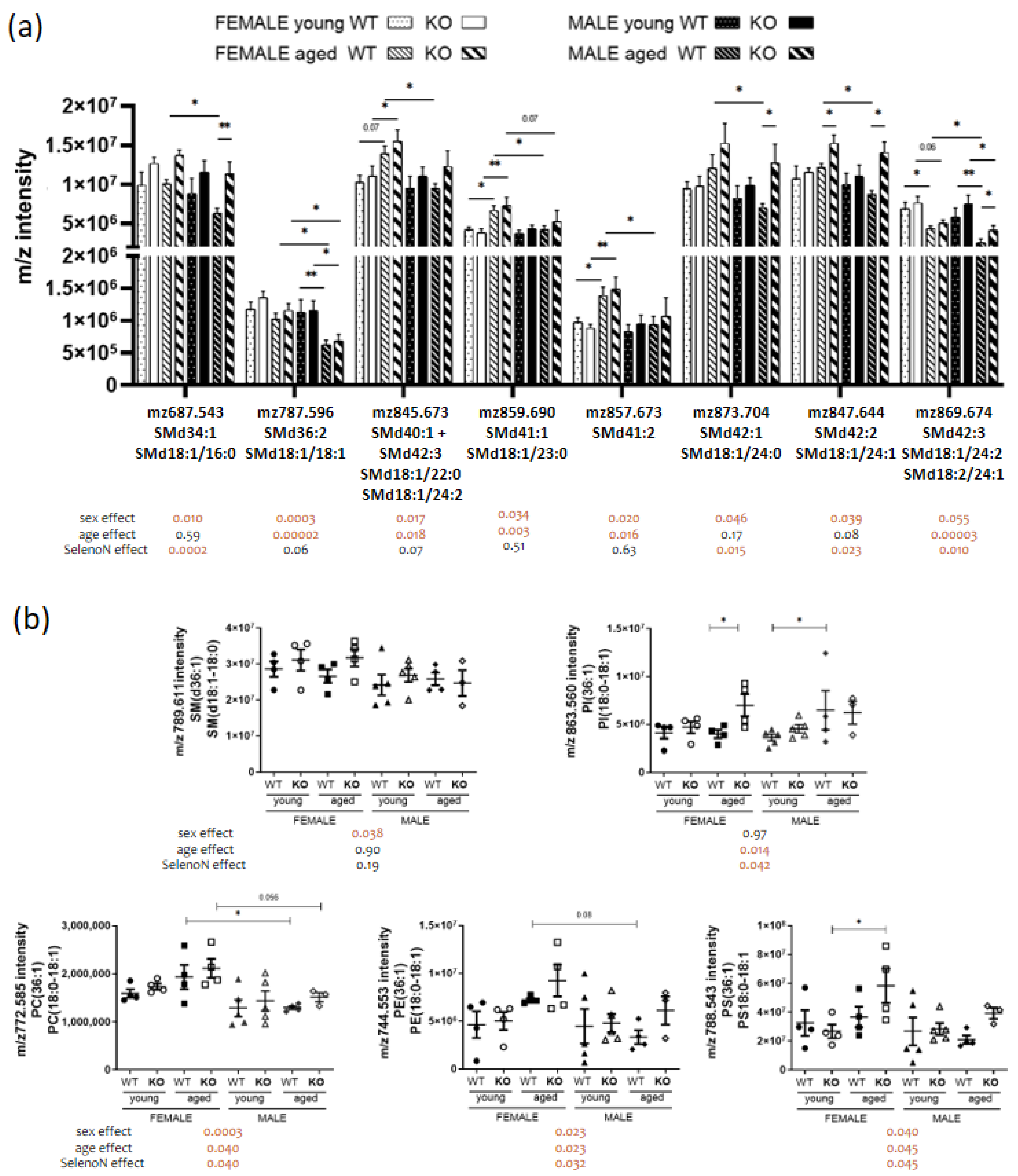
3.3. Sex-Specific PL Remodeling in More Than Half of PLs
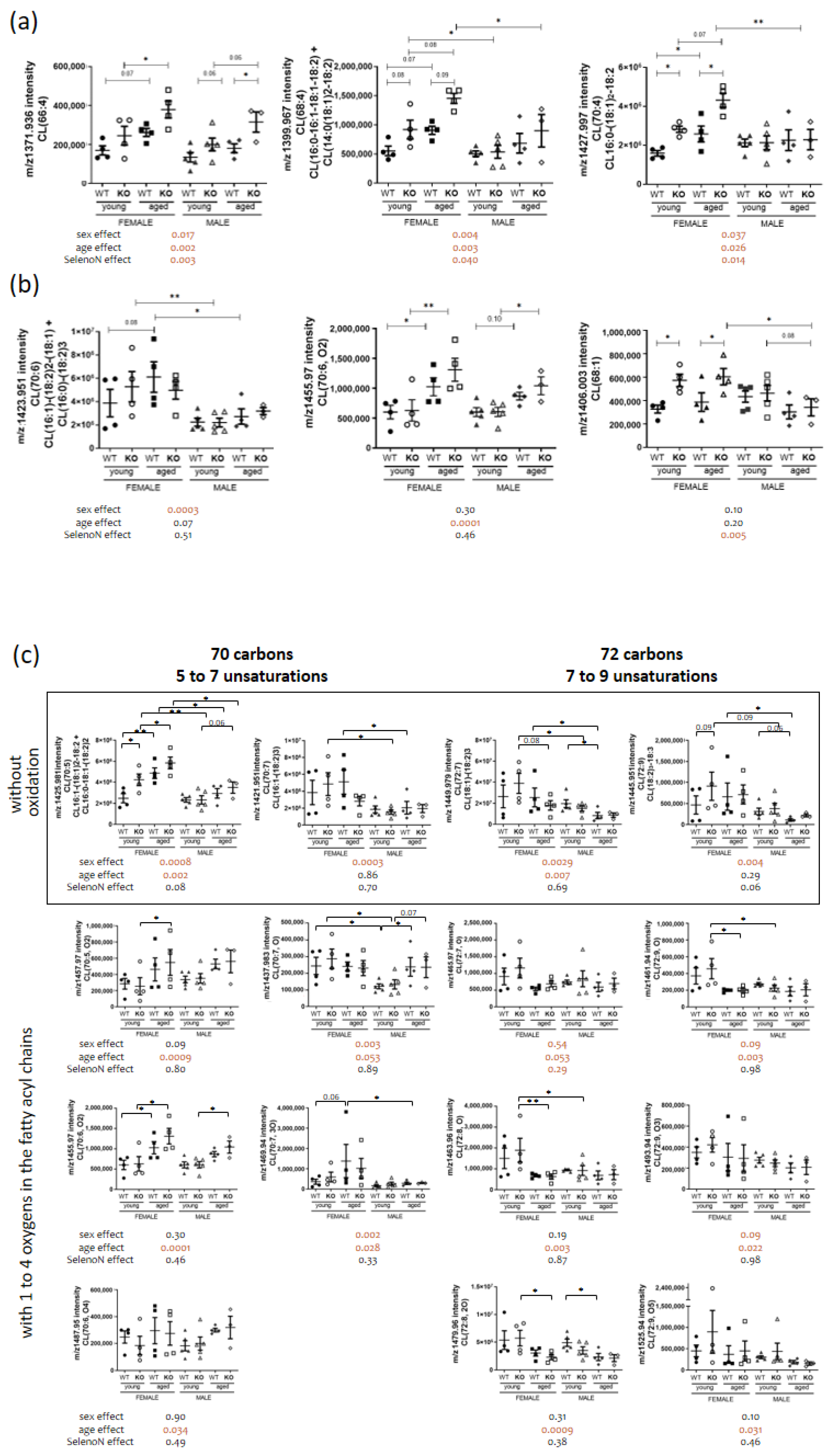
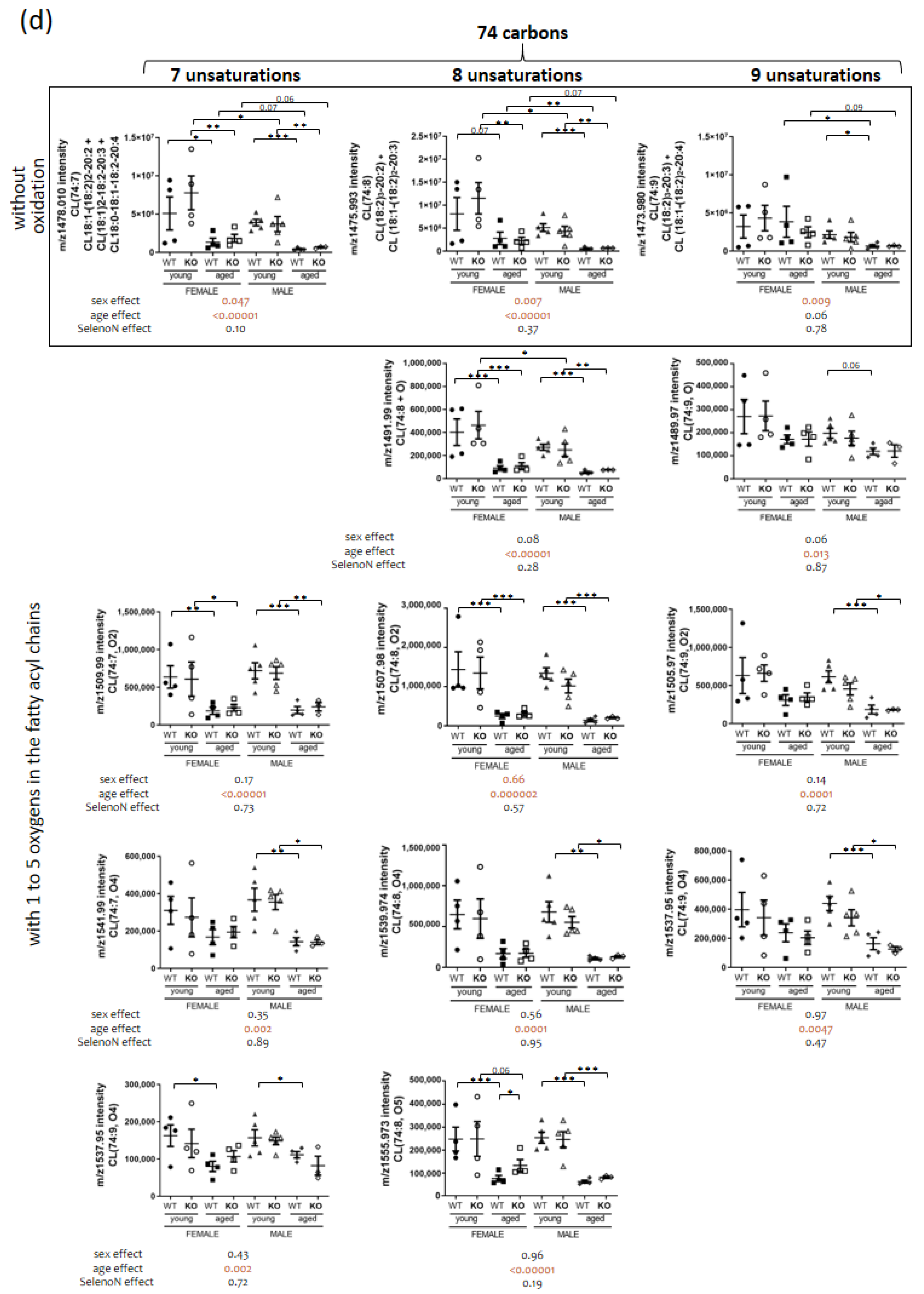
3.4. Sphingomyelin and Cardiolipin Changes in Diaphragm
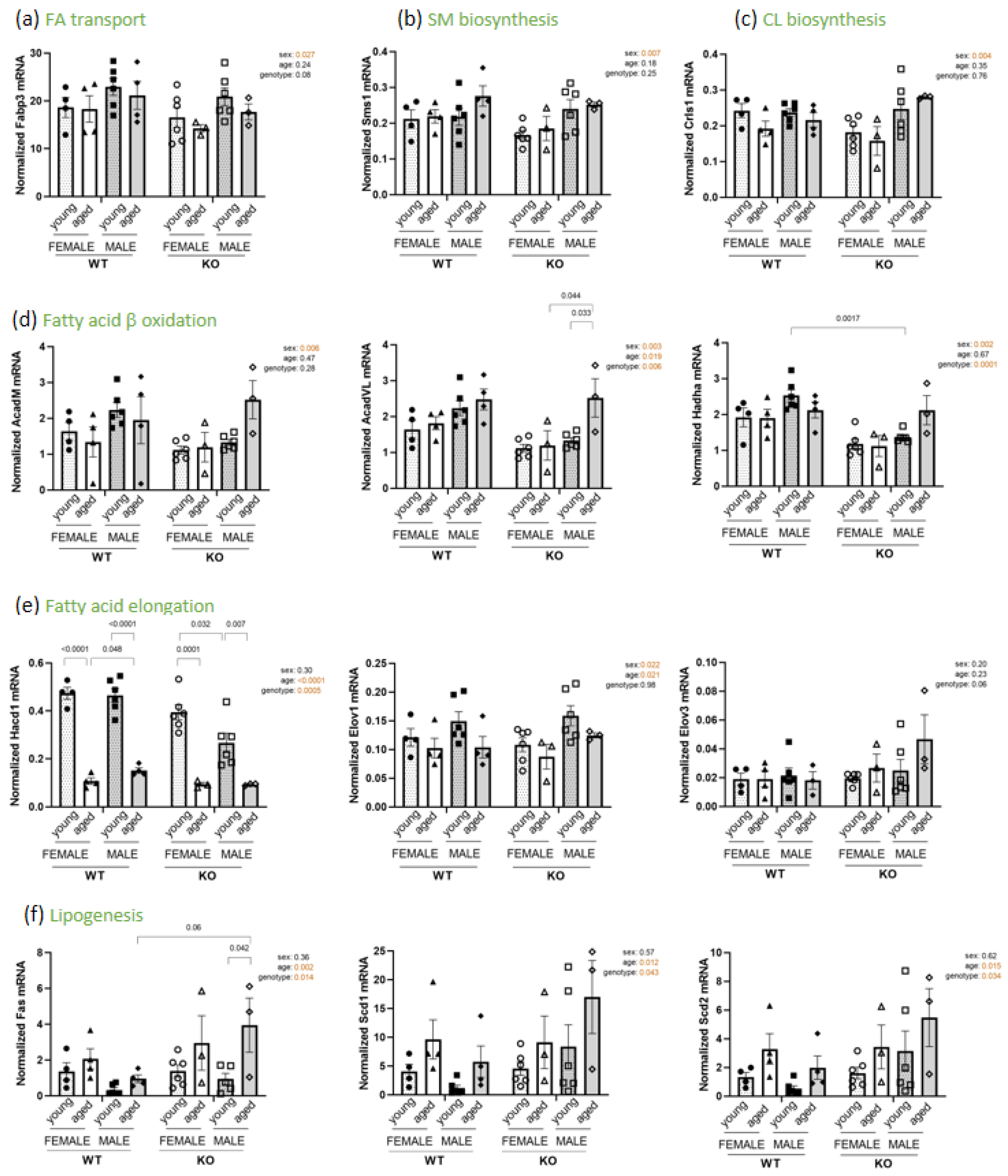
3.5. Modifications in PL and Fatty Acid Biosynthesis Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zumbuehl, A. Artificial phospholipids and their vesicles. Langmuir 2019, 35, 10223–10232. [Google Scholar] [CrossRef] [PubMed]
- Muallem, S.; Chung, W.Y.; Jha, A.; Ahuja, M. Lipids at membrane contact sites: Cell signaling and ion transport. EMBO Rep. 2017, 18, 1893–1904. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Jennings, W.; Epand, R.M. Roles of specific lipid species in the cell and their molecular mechanism. Prog. Lipid Res. 2016, 62, 75–92. [Google Scholar] [CrossRef]
- Bargui, R.; Solgadi, A.; Prost, B.; Chester, M.; Ferreiro, A.; Piquereau, J.; Moulin, M. Phospholipids: Identification and implication in muscle pathophysiology. Int. J. Mol. Sci. 2021, 22, 8176. [Google Scholar] [CrossRef] [PubMed]
- Wenk, M.R. Lipidomics: New tools and applications. Cell 2010, 143, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Wortmann, S.B.; Espeel, M.; Almeida, L.; Reimer, A.; Bosboom, D.; Roels, F.; de Brouwer, A.P.; Wevers, R.A. Inborn errors of metabolism in the biosynthesis and remodelling of phospholipids. J. Inherit. Metab. Dis. 2015, 38, 99–110. [Google Scholar] [CrossRef]
- Garlid, A.O.; Schaffer, C.T.; Kim, J.; Bhatt, H.; Guevara-Gonzalez, V.; Ping, P. Taz encodes tafazzin, a transacylase essential for cardiolipin formation and central to the etiology of barth syndrome. Gene 2020, 726, 144148. [Google Scholar] [CrossRef]
- Janssen, I.; Heymsfield, S.B.; Wang, Z.M.; Ross, R. Skeletal muscle mass and distribution in 468 men and women aged 18-88 yr. J. Appl. Physiol. 2000, 89, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Alcazar, J.; Aagaard, P.; Haddock, B.; Kamper, R.S.; Hansen, S.K.; Prescott, E.; Alegre, L.M.; Frandsen, U.; Suetta, C. Age- and sex-specific changes in lower-limb muscle power throughout the lifespan. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2020, 75, 1369–1378. [Google Scholar] [CrossRef]
- Tsekoura, M.; Kastrinis, A.; Katsoulaki, M.; Billis, E.; Gliatis, J. Sarcopenia and its impact on quality of life. Adv. Exp. Med. Biol. 2017, 987, 213–218. [Google Scholar]
- Fogarty, M.J.; Mantilla, C.B.; Sieck, G.C. Impact of sarcopenia on diaphragm muscle fatigue. Exp. Physiol. 2019, 104, 1090–1099. [Google Scholar] [CrossRef] [PubMed]
- Bordoni, B.; Morabito, B.; Simonelli, M. Ageing of the diaphragm muscle. Cureus 2020, 12, e6645. [Google Scholar] [CrossRef] [Green Version]
- O’Halloran, K.D. Diaphragm fatigue: Similarities and differences between sexes. J. Physiol. 2021, 599, 1023–1024. [Google Scholar] [CrossRef] [PubMed]
- Ricoy, J.; Rodriguez-Nunez, N.; Alvarez-Dobano, J.M.; Toubes, M.E.; Riveiro, V.; Valdes, L. Diaphragmatic dysfunction. Pulmonology 2019, 25, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Caggiano, S.; Khirani, S.; Dabaj, I.; Cavassa, E.; Amaddeo, A.; Arroyo, J.O.; Desguerre, I.; Richard, P.; Cutrera, R.; Ferreiro, A.; et al. Diaphragmatic dysfunction in sepn1-related myopathy. Neuromuscul. Disord. 2017, 27, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Villar-Quiles, R.N.; von der Hagen, M.; Metay, C.; Gonzalez, V.; Donkervoort, S.; Bertini, E.; Castiglioni, C.; Chaigne, D.; Colomer, J.; Cuadrado, M.L.; et al. The clinical, histologic, and genotypic spectrum of sepn1-related myopathy: A case series. Neurology 2020, 95, e1512–e1527. [Google Scholar] [CrossRef] [PubMed]
- Pozzer, D.; Invernizzi, R.W.; Blaauw, B.; Cantoni, O.; Zito, E. Ascorbic acid route to the endoplasmic reticulum: Function and role in disease. Antioxid. Redox Signal. 2021, 34, 845–855. [Google Scholar] [CrossRef]
- Gheller, B.J.; Riddle, E.S.; Lem, M.R.; Thalacker-Mercer, A.E. Understanding age-related changes in skeletal muscle metabolism: Differences between females and males. Annu. Rev. Nutr. 2016, 36, 129–156. [Google Scholar] [CrossRef]
- Filipe, A.; Chernorudskiy, A.; Arbogast, S.; Varone, E.; Villar-Quiles, R.N.; Pozzer, D.; Moulin, M.; Fumagalli, S.; Cabet, E.; Dudhal, S.; et al. Defective endoplasmic reticulum-mitochondria contacts and bioenergetics in sepn1-related myopathy. Cell Death Differ. 2021, 28, 123–138. [Google Scholar] [CrossRef]
- Moulin, M.; Solgadi, A.; Veksler, V.; Garnier, A.; Ventura-Clapier, R.; Chaminade, P. Sex-specific cardiac cardiolipin remodelling after doxorubicin treatment. Biol. Sex Differ. 2015, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 1 July 2019).
- Benton, H.P.; Want, E.J.; Ebbels, T.M. Correction of mass calibration gaps in liquid chromatography-mass spectrometry metabolomics data. Bioinformatics 2010, 26, 2488–2489. [Google Scholar] [CrossRef] [PubMed]
- Moulin, M.; Piquereau, J.; Mateo, P.; Fortin, D.; Rucker-Martin, C.; Gressette, M.; Lefebvre, F.; Gresikova, M.; Solgadi, A.; Veksler, V.; et al. Sexual dimorphism of doxorubicin-mediated cardiotoxicity: Potential role of energy metabolism remodeling. Circulation 2015, 8, 98–108. [Google Scholar] [CrossRef] [Green Version]
- Cho, W.; Stahelin, R.V. Membrane-protein interactions in cell signaling and membrane trafficking. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 119–151. [Google Scholar] [CrossRef] [PubMed]
- Regitz-Zagrosek, V. Sex and gender differences in health. Science & society series on sex and science. EMBO Rep. 2012, 13, 596–603. [Google Scholar] [PubMed] [Green Version]
- Group, E.U.C.C.S.; Regitz-Zagrosek, V.; Oertelt-Prigione, S.; Prescott, E.; Franconi, F.; Gerdts, E.; Foryst-Ludwig, A.; Maas, A.H.; Kautzky-Willer, A.; Knappe-Wegner, D.; et al. Gender in cardiovascular diseases: Impact on clinical manifestations, management, and outcomes. Eur. Heart J. 2016, 37, 24–34. [Google Scholar]
- Baroni, I.; Caruso, R.; Dellafiore, F.; Ausili, D.; Barello, S.; Vangone, I.; Russo, S.; Magon, A.; Conte, G.; Guardamagna, L.; et al. Self-care and type 2 diabetes mellitus (t2dm): A literature review in sex-related differences. Acta Bio-Med. Atenei Parm. 2022, 93, e2022277. [Google Scholar]
- Zhu, D.; Montagne, A.; Zhao, Z. Alzheimer’s pathogenic mechanisms and underlying sex difference. Cell. Mol. Life Sci. 2021, 78, 4907–4920. [Google Scholar] [CrossRef]
- Rosa-Caldwell, M.E.; Greene, N.P. Muscle metabolism and atrophy: Let’s talk about sex. Biol. Sex Differ. 2019, 10, 43. [Google Scholar] [CrossRef]
- Seifarth, J.E.; McGowan, C.L.; Milne, K.J. Sex and life expectancy. Gend. Med. 2012, 9, 390–401. [Google Scholar] [CrossRef]
- Zucker, I.; Beery, A.K. Males still dominate animal studies. Nature 2010, 465, 690. [Google Scholar] [CrossRef]
- Muralidharan, S.; Shimobayashi, M.; Ji, S.; Burla, B.; Hall, M.N.; Wenk, M.R.; Torta, F. A reference map of sphingolipids in murine tissues. Cell Rep. 2021, 35, 109250. [Google Scholar] [CrossRef] [PubMed]
- Janovska, A.; Hatzinikolas, G.; Mano, M.; Wittert, G.A. The effect of dietary fat content on phospholipid fatty acid profile is muscle fiber type dependent. Am. J. Physiol. 2010, 298, E779–E786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela, R.; Barrera, C.; Espinosa, A.; Llanos, P.; Orellana, P.; Videla, L.A. Reduction in the desaturation capacity of the liver in mice subjected to high fat diet: Relation to lcpufa depletion in liver and extrahepatic tissues. Prostaglandins Leukot. Essent. Fat. Acids 2015, 98, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Albouery, M.; Buteau, B.; Gregoire, S.; Martine, L.; Gambert, S.; Bron, A.M.; Acar, N.; Chassaing, B.; Bringer, M.A. Impact of a high-fat diet on the fatty acid composition of the retina. Exp. Eye Res. 2020, 196, 108059. [Google Scholar] [CrossRef]
- Mesmin, B. Mitochondrial lipid transport and biosynthesis: A complex balance. J. Cell Biol. 2016, 214, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A central target for sex differences in pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef]
- Herrera-Cruz, M.S.; Simmen, T. Over six decades of discovery and characterization of the architecture at mitochondria-associated membranes (mams). Adv. Exp. Med. Biol. 2017, 997, 13–31. [Google Scholar]
- Vance, J.E. Phospholipid synthesis in a membrane fraction associated with mitochondria. J. Biol. Chem. 1990, 265, 7248–7256. [Google Scholar] [CrossRef]
- Moltedo, O.; Remondelli, P.; Amodio, G. The mitochondria-endoplasmic reticulum contacts and their critical role in aging and age-associated diseases. Front. Cell Dev. Biol. 2019, 7, 172. [Google Scholar] [CrossRef]
- Flowers, M.T.; Ntambi, J.M. Role of stearoyl-coenzyme a desaturase in regulating lipid metabolism. Curr. Opin. Lipidol. 2008, 19, 248–256. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Tang, H.; Pang, S. The crucial roles of phospholipids in aging and lifespan regulation. Front. Physiol. 2021, 12, 775648. [Google Scholar] [CrossRef] [PubMed]
- Prola, A.; Blondelle, J.; Vandestienne, A.; Piquereau, J.; Denis, R.G.P.; Guyot, S.; Chauvin, H.; Mourier, A.; Maurer, M.; Henry, C.; et al. Cardiolipin content controls mitochondrial coupling and energetic efficiency in muscle. Sci. Adv. 2021, 7, eabd6322. [Google Scholar] [CrossRef] [PubMed]
- Ikon, N.; Ryan, R.O. Barth syndrome: Connecting cardiolipin to cardiomyopathy. Lipids 2017, 52, 99–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlame, M.; Ren, M. Barth syndrome, a human disorder of cardiolipin metabolism. FEBS Lett. 2006, 580, 5450–5455. [Google Scholar] [CrossRef]
- Pomatto, L.C.D.; Davies, K.J.A. Adaptive homeostasis and the free radical theory of ageing. Free. Radic. Biol. Med. 2018, 124, 420–430. [Google Scholar] [CrossRef]
- Mutlu, A.S.; Duffy, J.; Wang, M.C. Lipid metabolism and lipid signals in aging and longevity. Dev. Cell 2021, 56, 1394–1407. [Google Scholar] [CrossRef]
- Moulin, M.; Ferreiro, A. Muscle redox disturbances and oxidative stress as pathomechanisms and therapeutic targets in early-onset myopathies. Semin. Cell Dev. Biol. 2016, 64, 213–223. [Google Scholar] [CrossRef]
- Dabaj, I.; Ferey, J.; Marguet, F.; Gilard, V.; Basset, C.; Bahri, Y.; Brehin, A.C.; Vanhulle, C.; Leturcq, F.; Marret, S.; et al. Muscle metabolic remodelling patterns in duchenne muscular dystrophy revealed by ultra-high-resolution mass spectrometry imaging. Sci. Rep. 2021, 11, 1906. [Google Scholar] [CrossRef]
- Valentine, W.J.; Mostafa, S.A.; Tokuoka, S.M.; Hamano, F.; Inagaki, N.F.; Nordin, J.Z.; Motohashi, N.; Kita, Y.; Aoki, Y.; Shimizu, T.; et al. Lipidomic analyses reveal specific alterations of phosphatidylcholine in dystrophic mdx muscle. Front. Physiol. 2021, 12, 698166. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Rpl19 Hprt Fabp3 Sms1 Crls1 AcadM AcadVL Hadha Elov1 Elov3 Fas Scd1 Scd2 | GGGCAGGCATATGGGCATA AGCTACTGTAATGATCAGTCAACG TTCTGGAAGCTAGTGGACAG TCCATCACAGGCTCGCACA TGACCTATGCAGATCTTATTCCA CCGTTCCCTCTCATCAAAAG TGGGCCTCTCTAATACCCAGT GCTTGGGAGGAGGACTTGA GGCCCTGATCCCTTTGAA GCACCATCTTTGGCATACTG GCAAGTGCAGCCTGAGGGAC CTACATGACCAGCGCTCTG GTAATCAGCGCCCTGGGCAT | GGCGGTCAATCTTCTTGGATT AGAGGTCCTTTTCACCAGCA TGATGGTAGTAGGCTTGGTCAT ACGCTGAGGAGCCAGCAAA CCTTGCTGATGAATGTTGGTT ACACCCATACGCCAACTCTT TCCCAGGGTAACGCTAACAC AGCAACACTTCAGGGACACC CCATCCTGGCTAAGGACTCA TTGTGGGTGTGGCATCCTTTC ACAGCCTGGGGTCATCTTTGC CGTACACGTCATTCTGGAACG CATACACGTCATTCTGGAACGC |
| Detailed Identification | Global Identification | m/z | p_SEX | p_AGE | p_SELENON |
|---|---|---|---|---|---|
| ND | CL(66:4) | 1371.90 | 0.0168 | 0.0016 | 0.0026 |
| ND | CL(66:5) | 1369.91 | 0.0148 | 0.74 | 0.18 |
| 16:0-16:1-18:1-18:2 + 14:0-(18:1)2-18:2 | CL(68:4) | 1399.97 | 0.0027 | 0.0035 | 0.0403 |
| 14:0-18:1-(18:2)2 + (16:1)2-18:1-18:2 | CL(68:5) | 1397.95 | 0.0025 | 0.053 | 0.08 |
| ND | CL(68:6) | 1395.93 | 0.0026 | 0.07 | 0.62 |
| 16:0-(18:1)3 | CL(70:3) | 1430.00 | 0.054 | 0.06 | 0.0199 |
| 16:0-(18:1)2-18:2 | CL(70:4) | 1427.99 | 0.0369 | 0.0259 | 0.0136 |
| 16:1-(18:1)2-18:2 + 16:0-18:1-(18:2)2 | CL(70:5) | 1425.98 | 0.0008 | 0.0016 | 0.08 |
| 16:1-(18:2)2-18:1 + 16:0-(18:2)3 | CL(70:6) | 1423.97 | 0.0003 | 0.07 | 0.51 |
| 16:1-(18:2)3 | CL(70:7) | 1421.95 | 0.0003 | 0.86 | 0.70 |
| CL(70:7. +1O) | 1437.95 | 0.0029 | 0.053 | 0.89 | |
| CL(70:7. +3O) | 1469.94 | 0.0020 | 0.0289 | 0.33 | |
| (18:1)3-18:2 + 18:0-18:1-(18:2)2 | CL(72:5) | 1454.00 | 0.0110 | 0.0374 | 0.11 |
| (18:1)2-(18:2)2 | CL(72:6) | 1451.99 | 0.0065 | 0.25 | 0.31 |
| CL(72:6. +3O) | 1499.99 | 0.0089 | 0.26 | 0.20 | |
| 18:1-(18:2)3 | CL(72:7) | 1449.98 | 0.0029 | 0.0072 | 0.69 |
| CL(72:7. +3O) | 1497.97 | 0.0044 | 0.71 | 0.32 | |
| CL(72:7. +6O) | 1545.95 | 0.0411 | 0.32 | 0.76 | |
| (18:2)4 | CL(72:8) | 1447.96 | 0.0018 | 0.0008 | 0.78 |
| CL(72:8. +3O) | 1495.95 | 0.0014 | 0.15 | 0.81 | |
| CL(72:8. +5O) | 1527.94 | 0.0447 | 0.09 | 0.85 | |
| CL(72:8. +6O) | 1543.93 | 0.0151 | 0.86 | 0.68 | |
| CL(72:8. +9O) | 1591.93 | 0.0182 | 0.12 | 0.94 | |
| (18:2)3-18:3 | CL(72:9) | 1445.95 | 0.0039 | 0.29 | 0.06 |
| ND | CL(74:10) | 1471.96 | 0.0046 | 0.27 | 0.26 |
| 18:1-(18:2)2-20:2 + (18:1)2-18:2-20:3 + 18:0-18:1-18:2-20:4 | CL(74:7) | 1478.01 | 0.0475 | 0.000002 | 0.10 |
| (18:2)3-20:2 + 18:1-(18:2)2-20:3 | CL(74:8) | 1475.99 | 0.0066 | 0.000002 | 0.37 |
| CL(74:8. +3O) | 1523.99 | 0.0132 | 0.0006 | 0.26 | |
| (18:2)3-20:3 + 18:1-(18:2)2-20:4 | CL(74:9) | 1473.98 | 0.0086 | 0.06 | 0.78 |
| CL(74:9. +3O) | 1521.97 | 0.0047 | 0.0446 | 0.64 | |
| ND | CL(76:8) | 1504.02 | 0.0143 | 0.00002 | 0.17 |
| ND | CL(76:9) | 1502.00 | 0.0249 | 0.16 | 0.27 |
| 16:0 | LPE(16:0) | 452.28 | 0.0007 | 0.0112 | 0.14 |
| 18:0 | LPE(18:0) | 480.31 | 0.00003 | 0.0097 | 0.07 |
| 18:1 | LPE(18:1) | 478.29 | 0.0090 | 0.21 | 0.08 |
| 18:2 | LPE(18:2) | 476.28 | 0.0043 | 0.0232 | 0.42 |
| 20:4 | LPE(20:4) | 500.28 | 0.0035 | 0.60 | 0.12 |
| 22:6 | LPE(22/6) | 524.28 | 0.0010 | 0.34 | 0.60 |
| 22:5 | LPE(22:5) | 526.29 | 0.0097 | 0.0523 | 0.30 |
| 16:0-14:0 | PC(30:0) | 764.54 | 0.0236 | 0.0035 | 0.09 |
| 18:1-18:0 + 20:1-16:0 | PC(36:1) | 846.62 | 0.0002 | 0.07 | 0.0389 |
| 18:0-18:1 | PC(36:1) | 772.58 | 0.00003 | 0.0401 | 0.0401 |
| 18:2-18:0 + 18:1-18:1 + 16:0-20:2 | PC(36:2) | 844.61 | 0.0092 | 0.0006 | 0.50 |
| 16:0-20:4 + 18:2-18:2 | PC(36:4) | 840.57 | 0.0013 | 0.0081 | 0.80 |
| 20:4-18:0 + 22:4-16:0 | PC(38:4) | 868.60 | 0.00002 | 0.0130 | 0.34 |
| 22:5-16:0 + 20:4-18:1 | PC(38:5) | 866.59 | 0.0103 | 0.16 | 0.37 |
| 22:6-16:0 | PC(38:6) | 864.57 | 0.0040 | 0.87 | 0.69 |
| ND | PC(39:3(OH)) | 876.59 | 0.0272 | 0.31 | 0.46 |
| ND | PC(39:4(OH)) | 874.58 | 0.0100 | 0.52 | 0.33 |
| 18:0-22:5 | PC(40:5) | 894.61 | 0.0017 | 0.52 | 0.12 |
| 18:0-22:6 | PC(40:6) | 892.61 | 0.0005 | 0.53 | 0.36 |
| 22:6-18:1 + 22:5-18:2 + 20:3-20:4 | PC(40:7) | 890.59 | 0.0256 | 0.89 | 0.59 |
| ND | PC(44:8) | 870.59 | 0.0000 | 0.11 | 0.18 |
| ND | PC(46:11(OH)) | 908.58 | 0.0210 | 0.21 | 0.14 |
| ND | PE(32:0) | 690.51 | 0.0232 | 0.51 | 0.21 |
| 16:0-18:1 + 16:1-18:0 | PE(34:1) | 716.52 | 0.0013 | 0.77 | 0.10 |
| 16:0-18:2 + 16:1-18:1 | PE(34:2) | 714.51 | 0.0387 | 0.0063 | 0.0494 |
| 18:0-18:1 | PE(36:1) | 744.55 | 0.0226 | 0.0233 | 0.0320 |
| 18:0-18:2 + 18:1-18:1 | PE(36:2) | 742.54 | 0.0008 | 0.0092 | 0.46 |
| 20:3-16:0 + 18:2-18:1 | PE(36:3) | 740.52 | 0.0065 | 0.0207 | 0.59 |
| 20:4-16:0 + 18:2-18:2 | PE(36:4) | 738.51 | 0.0009 | 0.0395 | 0.54 |
| ND | PE(38:4(OH)) + PE(P-36:4) | 782.53 | 0.0058 | 0.0036 | 0.33 |
| 20:4-18:0 | PE(38:4) | 766.54 | 0.0003 | 0.0033 | 0.62 |
| 22:5-16:0 | PE(38:5) | 764.52 | 0.0021 | 0.0462 | 0.31 |
| ND | PE(38:6(OH)) | 778.50 | 0.0009 | 0.54 | 0.51 |
| ND | PE(38:6(OH)) + PE (41:7) | 838.52 | 0.0041 | 0.17 | 0.28 |
| 22:6-16:0 | PE(38:6) | 762.51 | 0.0003 | 0.76 | 0.32 |
| 22:5-18:0 + 22:4-18:1 | PE(40:5) | 792.55 | 0.0007 | 0.48 | 0.24 |
| ND | PE(40:6(OH)) | 806.53 | 0.0002 | 0.22 | 0.19 |
| 22:6-18:0 + 22:5-18:1 | PE(40:6) | 790.54 | 0.0002 | 0.79 | 0.40 |
| 22:6-18:1 + 22:5-18:2 | PE(40:7) | 788.52 | 0.0187 | 0.20 | 0.89 |
| ND | PE(O-32:1) + PE(P-32:0) | 674.51 | 0.0005 | 0.0244 | 0.68 |
| ND | PE(O-34:1) + PE(P-34:0) | 702.54 | 0.0011 | 0.86 | 0.35 |
| PE(P-16:0-18:1) | PE(O-34:2) + PE(P-34:1) | 700.53 | 0.0034 | 0.99 | 0.47 |
| ND | PE(O-34:3) + PE(P-34:2) | 698.51 | 0.0010 | 0.0166 | 0.73 |
| PE(P-18:0-18:1) | PE(O-36:2) + PE(P-36:1) | 728.56 | 0.0268 | 0.0122 | 0.38 |
| PE(P-18:1-18:1) | PE(O-36:3) + PE(P-36:2) | 726.54 | 0.0295 | 0.0160 | 0.51 |
| PE(P-16:0-20:4) | PE(O-36:5) + PE(P-36:4) | 722.51 | 0.0009 | 0.20 | 0.15 |
| ND | PE(O-38:4) + PE(P-38:3) | 752.56 | 0.0011 | 0.52 | 0.13 |
| PE(P-18:0-20:4) + PE(P-16:0-22:4) | PE(O-38:5) + PE(P-38:4) | 750.54 | 0.0007 | 0.28 | 0.19 |
| PE(P-18:0-22:5) | PE(P-18:0-22:5) | 776.56 | 0.0010 | 0.68 | 0.27 |
| ND | PE(P-32:1) | 672.50 | 0.0022 | 0.14 | 0.35 |
| PE(P-16:0-22:6) | PE(P-38:6) | 746.51 | 0.0004 | 0.91 | 0.77 |
| PE(P-18:0-22:6) | PE(P-40:6) | 774.54 | 0.0002 | 0.21 | 0.47 |
| (16:0)2 | PI(32:0) | 809.52 | 0.0048 | 0.0040 | 0.0212 |
| ND | PI(32:2) | 865.51 | 0.0007 | 0.18 | 0.97 |
| ND | PI(34:3) | 891.52 | 0.0021 | 0.62 | 0.89 |
| ND | PI(36:3(OH)) | 875.53 | 0.0013 | 0.39 | 0.19 |
| 20:4-16:0 | PI(36:4) | 857.52 | 0.0011 | 0.0358 | 0.14 |
| ND | PI(36:5) | 915.52 | 0.0313 | 0.15 | 0.90 |
| 18:0-20:3 | PI(38:3) | 887.56 | 0.0012 | 0.68 | 0.60 |
| 18:0-20:4 | PI(38:4) | 885.55 | 0.0002 | 0.52 | 0.46 |
| 18:1-20:4 + 18:0-20:5 + 16:0-22:5 | PI(38:5) | 883.53 | 0.0031 | 0.32 | 0.24 |
| ND | PI(38:6) | 881.52 | 0.0013 | 0.07 | 0.18 |
| 18:0-22:4 | PI(40:4) | 913.57 | 0.0180 | 0.0069 | 0.13 |
| 18:0-22:5 | PI(40:5) | 911.56 | 0.0292 | 0.0115 | 0.18 |
| 18:0-22:6 | PI(40:6) | 909.55 | 0.0031 | 0.53 | 0.24 |
| ND | PI(P-37:2) | 895.54 | 0.0005 | 0.48 | 0.68 |
| 18:1-18:0 | PS(36:1) | 788.54 | 0.0403 | 0.0450 | 0.0453 |
| 20:3-18:0 | PS(38:3) | 812.54 | 0.0013 | 0.12 | 0.07 |
| 18:0-20:4 + 18:1-20:3 | PS(38:4) | 810.53 | 0.0001 | 0.59 | 0.13 |
| 22:6-18:0 | PS(40:6) | 834.53 | 0.0001 | 0.70 | 0.56 |
| d18:1/16:0 | SM(d34:1) | 761.58 | 0.0344 | 0.71 | 0.00 |
| d18:1/18:0 | SM(d36:1) | 765.57 | 0.0143 | 0.40 | 0.43 |
| d18:1/18:1 | SM(d36:2) | 787.60 | 0.0021 | 0.0003 | 0.11 |
| d18:1/22:0 + d18:1/24:2 | SM(d40:1) + SM(d42:3) | 845.67 | 0.0315 | 0.0338 | 0.10 |
| d18:1/23:0 | SM(d41:1) | 859.69 | 0.0278 | 0.0023 | 0.48 |
| ND | SM(d41:2) | 857.67 | 0.0231 | 0.0195 | 0.64 |
| d18:1/24:0 | SM(d42:1) | 873.70 | 0.0388 | 0.08 | 0.0234 |
| d18:1/24:1 | SM(d42:2) | 871.69 | 0.0464 | 0.17 | 0.0147 |
| Detailed Identification | Global Identification | m/z | p_SEX | p_AGE | p_SELENON |
|---|---|---|---|---|---|
| ND | CL(66:4) | 1371.90 | 0.0168 | 0.0016 | 0.0026 |
| 16:0-16:1-18:1-18:2 + 14:0-(18:1)2-18:2 | CL(68:4) | 1399.97 | 0.0027 | 0.0035 | 0.0403 |
| 14:0-18:1-(18:2)2 + (16:1)2-18:1-18:2 | CL(68:5) | 1397.95 | 0.0025 | 0.0529 | 0.08 |
| 16:0-(18:1)2-18:2 | CL(70:4) | 1427.99 | 0.0369 | 0.0259 | 0.0136 |
| 16:1-(18:1)2-18:2 + 16:0-18:1-(18:2)2 | CL(70:5) | 1425.98 | 0.0008 | 0.0016 | 0.08 |
| CL(70:5, +2O) | 1457.97 | 0.09 | 0.0009 | 0.80 | |
| CL(70:6, +2O) | 1455.97 | 0.30 | 0.0001 | 0.46 | |
| CL(70:6, +4O) | 1487.95 | 0.90 | 0.0338 | 0.49 | |
| CL(70:7, +1O) | 1437.95 | 0.0029 | 0.0531 | 0.89 | |
| CL(70:7, +3O) | 1469.94 | 0.0020 | 0.0289 | 0.33 | |
| (18:1)3-18:2 + 18:0-18:1-(18:2)2 | CL(72:5) | 1454.00 | 0.0110 | 0.0374 | 0.11 |
| 18:1-(18:2)3 | CL(72:7) | 1449.98 | 0.0029 | 0.0072 | 0.69 |
| CL(72:7, +1O) | 1465.97 | 0.54 | 0.0527 | 0.29 | |
| (18:2)4 | CL(72:8) | 1447.96 | 0.0018 | 0.0008 | 0.78 |
| CL(72:8, +1O) | 1463.96 | 0.19 | 0.0029 | 0.87 | |
| CL(72:8, +2O) | 1479.96 | 0.31 | 0.0009 | 0.38 | |
| CL(72:9, +1O) | 1461.94 | 0.09 | 0.0031 | 0.98 | |
| CL(72:9, +3O) | 1493.93 | 0.09 | 0.0220 | 0.98 | |
| CL(72:9, +5O) | 1525.94 | 0.10 | 0.0305 | 0.46 | |
| 18:1-(18:2)2-20:2 + (18:1)2-18:2-20:3 + 18:0-18:1-18:2-20:4 | CL(74:7) | 1478.01 | 0.0475 | 0.000002 | 0.10 |
| ND | CL(74:7, +2O) | 1510.00 | 0.17 | 0.00002 | 0.73 |
| ND | CL(74:7, +4O) | 1541.98 | 0.35 | 0.0015 | 0.89 |
| ND | CL(74:7, +5O) | 1557.95 | 0.43 | 0.0019 | 0.72 |
| (18:2)3-20:2 + 18:1-(18:2)2-20:3 | CL(74:8) | 1475.99 | 0.0066 | 0.000002 | 0.37 |
| ND | CL(74:8, +1O) | 1491.99 | 0.08 | 0.0000003 | 0.28 |
| ND | CL(74:8, +2O) | 1507.98 | 0.66 | 0.0000002 | 0.57 |
| ND | CL(74:8, +3O) | 1523.99 | 0.0132 | 0.0006 | 0.26 |
| ND | CL(74:8, +4O) | 1539.97 | 0.56 | 0.00001 | 0.95 |
| ND | CL(74:8, +5O) | 1555.97 | 0.96 | 0.000001 | 0.19 |
| ND | CL(74:8, +6O) | 1571.97 | 0.10 | 0.0001 | 0.82 |
| ND | CL(74:9, +1O) | 1489.97 | 0.06 | 0.0128 | 0.87 |
| ND | CL(74:9, +2O) | 1505.97 | 0.14 | 0.0001 | 0.72 |
| ND | CL(74:9, +3O) | 1521.97 | 0.0047 | 0.0446 | 0.64 |
| ND | CL(74:9, +4O) | 1537.96 | 0.97 | 0.0047 | 0.47 |
| ND | CL(76:8) | 1504.02 | 0.0143 | 0.00002 | 0.17 |
| 16:0 | LPE(16:0) | 452.28 | 0.0007 | 0.0112 | 0.14 |
| 18:0 | LPE(18:0) | 480.31 | 0.00003 | 0.0097 | 0.07 |
| 18:2 | LPE(18:2) | 476.28 | 0.0043 | 0.0232 | 0.42 |
| 22:5 | LPE(22:5) | 526.29 | 0.0097 | 0.052 | 0.30 |
| 16:0-14:0 | PC(30:0) | 764.54 | 0.0236 | 0.0035 | 0.08 |
| 16:0-16:0 | PC(32:0) | 768.53 | 0.29 | 0.0010 | 0.19 |
| 18:2-16:0 + 18:1-16:1 | PC(34:2) | 816.57 | 0.98 | 0.00002 | 0.47 |
| ND | PC(34:3) | 814.56 | 0.94 | 0.0069 | 0.99 |
| 18:0-18:1 | PC(36:1) | 772.58 | 0.00001 | 0.0401 | 0.0401 |
| 18:2-18:0 + 18:1-18:1 + 16:0-20:2 | PC(36:2) | 844.61 | 0.0092 | 0.0006 | 0.50 |
| 18:1-18:2 + 16:0-20:3 | PC(36:3) | 842.59 | 0.43 | 0.0137 | 0.41 |
| 16:0-20:4 + 18:2-18:2 | PC(36:4) | 840.57 | 0.0013 | 0.0081 | 0.80 |
| 20:4-18:0 + 22:4-16:0 | PC(38:4) | 868.60 | 0.0000 | 0.0130 | 0.34 |
| 16:0-18:2 + 16:1-18:1 | PE(34:2) | 714.51 | 0.0387 | 0.0063 | 0.0494 |
| 18:0-18:1 | PE(36:1) | 744.55 | 0.0226 | 0.0233 | 0.0320 |
| 18:0-18:2 + 18:1-18:1 | PE(36:2) | 742.54 | 0.0008 | 0.0092 | 0.46 |
| 20:3-16:0 + 18:2-18:1 | PE(36:3) | 740.52 | 0.0065 | 0.0207 | 0.59 |
| 20:4-16:0 + 18:2-18:2 | PE(36:4) | 738.51 | 0.0009 | 0.0395 | 0.54 |
| ND | PE(38:4(OH)) + PE(P-36:4) | 782.53 | 0.0058 | 0.0036 | 0.33 |
| 20:4-18:0 | PE(38:4) | 766.54 | 0.0003 | 0.0033 | 0.62 |
| 22:5-16:0 | PE(38:5) | 764.52 | 0.0021 | 0.0462 | 0.31 |
| ND | PE(O-32:1) + PE(P-32:0) | 674.51 | 0.0005 | 0.0244 | 0.68 |
| ND | PE(O-34:3) + PE(P-34:2) | 698.51 | 0.0010 | 0.0166 | 0.73 |
| PE(P-18:0-18:1) | PE(O-36:2) + PE(P-36:1) | 728.56 | 0.0268 | 0.0122 | 0.38 |
| PE(P-18:1-18:1) | PE(O-36:3) + PE(P-36:2) | 726.54 | 0.0295 | 0.0160 | 0.51 |
| 18:2-16:0 + 18:1-16:1 | PG(34:2) | 745.50 | 0.06 | 0.0514 | 0.19 |
| 16:0-19:1 + 17:0-18:1 | PG(35:1) | 761.53 | 0.29 | 0.0453 | 0.09 |
| (16:0)2 | PI(32:0) | 809.52 | 0.0048 | 0.0040 | 0.0212 |
| 18:2-16:0 + 18:1-16:1 + 18:0-16:2 | PI(34:2) | 833.52 | 0.47 | 0.0060 | 0.36 |
| 18:0-18:1 | PI(36:1) | 863.56 | 0.97 | 0.0139 | 0.0422 |
| 20:4-16:0 | PI(36:4) | 857.52 | 0.0011 | 0.0358 | 0.14 |
| ND | PI(38:6) | 941.54 | 0.17 | 0.0351 | 0.42 |
| 18:0-22:4 | PI(40:4) | 913.57 | 0.0180 | 0.0069 | 0.13 |
| 18:0-22:5 | PI(40:5) | 911.56 | 0.0292 | 0.0115 | 0.18 |
| 18:1-18:0 | PS(36:1) | 788.54 | 0.0403 | 0.0450 | 0.0453 |
| d18:1/18:1 | SM(d36:2) | 787.60 | 0.0021 | 0.0003 | 0.11 |
| d18:1/22:0 + d18:1-24:2 | SM(d40:1) + SM(d42:3) | 845.67 | 0.0315 | 0.0338 | 0.10 |
| d18:1/23:0 | SM(d41:1) | 859.69 | 0.0278 | 0.0023 | 0.48 |
| ND | SM(d41:2) | 857.67 | 0.0231 | 0.0195 | 0.64 |
| d18:1/24:2 + d18:2/24:1 | SM(d42:3) | 869.67 | 0.055 | 0.00003 | 0.0094 |
| Detailed Identification | Global Identification | m/z | p_SEX | p_AGE | p_SELENON |
|---|---|---|---|---|---|
| ND | CL(66:4) | 1371.90 | 0.0168 | 0.0016 | 0.0026 |
| ND | CL(68:1) | 1406.00 | 0.10 | 0.20 | 0.0047 |
| (16:0)2-(18:1)2 | CL(68:2) | 1404.00 | 0.34 | 0.51 | 0.0031 |
| 16:0-16:1-(18:2)2 + (16:0)2-18:1-18:2 | CL(68:3) | 1401.98 | 0.13 | 0.94 | 0.0039 |
| 16:0-16:1-18:1-18:2 + 14:0-(18:1)2-18:2 | CL(68:4) | 1399.97 | 0.0027 | 0.0035 | 0.0403 |
| 16:0-(18:1)3 | CL(70:3) | 1430.00 | 0.054 | 0.058 | 0.0199 |
| 16:0-(18:1)2-18:2 | CL(70:4) | 1427.99 | 0.0369 | 0.0259 | 0.0136 |
| 18:0-18:1 | PC(36:1) | 772.58 | 0.00003 | 0.0401 | 0.0401 |
| 16:0-18:2 + 16:1-18:1 | PE(34:2) | 714.51 | 0.0387 | 0.0063 | 0.0494 |
| 18:0-18:1 | PE(36:1) | 744.55 | 0.0226 | 0.0233 | 0.0320 |
| 14:0-16:0 | PG(30:0) | 693.47 | 0.08 | 0.70 | 0.0156 |
| (16:0)2 | PI(32:0) | 809.52 | 0.0048 | 0.0040 | 0.0212 |
| 18:0-18:1 | PI(36:1) | 863.56 | 0.97 | 0.0139 | 0.0422 |
| 18:1-18:0 | PS(36:1) | 788.54 | 0.0403 | 0.0450 | 0.0453 |
| 18:1-18:1 + 18:0-18:2 | PS(36:2) | 786.53 | 0.08 | 0.47 | 0.0232 |
| d18:1/16:0 | SM(d34:1) | 761.58 | 0.0344 | 0.71 | 0.0006 |
| ND | SM(d34:2) | 759.57 | 0.16 | 0.07 | 0.0034 |
| d18:1/24:0 | SM(d42:1) | 873.70 | 0.0388 | 0.08 | 0.0234 |
| d18:1/24:1 | SM(d42:2) | 871.69 | 0.0464 | 0.17 | 0.0147 |
| d18:1/24:2 + d18:2/24:1 | SM(d42:3) | 869.67 | 0.055 | 0.00003 | 0.0094 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bargui, R.; Solgadi, A.; Dumont, F.; Prost, B.; Vadrot, N.; Filipe, A.; Ho, A.T.V.; Ferreiro, A.; Moulin, M. Sex-Specific Patterns of Diaphragm Phospholipid Content and Remodeling during Aging and in a Model of SELENON-Related Myopathy. Biomedicines 2023, 11, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11020234
Bargui R, Solgadi A, Dumont F, Prost B, Vadrot N, Filipe A, Ho ATV, Ferreiro A, Moulin M. Sex-Specific Patterns of Diaphragm Phospholipid Content and Remodeling during Aging and in a Model of SELENON-Related Myopathy. Biomedicines. 2023; 11(2):234. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11020234
Chicago/Turabian StyleBargui, Rezlène, Audrey Solgadi, Florent Dumont, Bastien Prost, Nathalie Vadrot, Anne Filipe, Andrew T. V. Ho, Ana Ferreiro, and Maryline Moulin. 2023. "Sex-Specific Patterns of Diaphragm Phospholipid Content and Remodeling during Aging and in a Model of SELENON-Related Myopathy" Biomedicines 11, no. 2: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines11020234