Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal
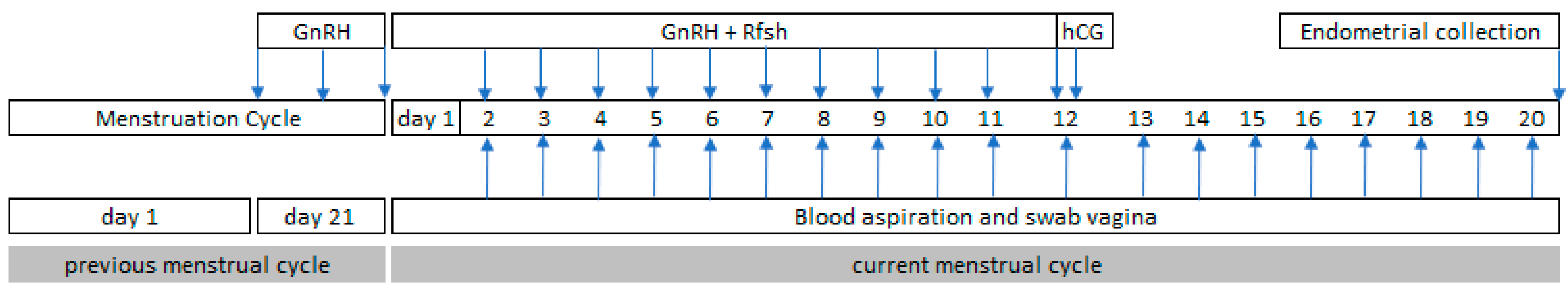
2.2. Controlled Ovarian Hyperstimulation Procedure
2.3. Endometrial Collection
2.4. Hormone Assay
2.5. Immunohistochemistry for HOXA10
2.6. Statistical Analysis
3. Result
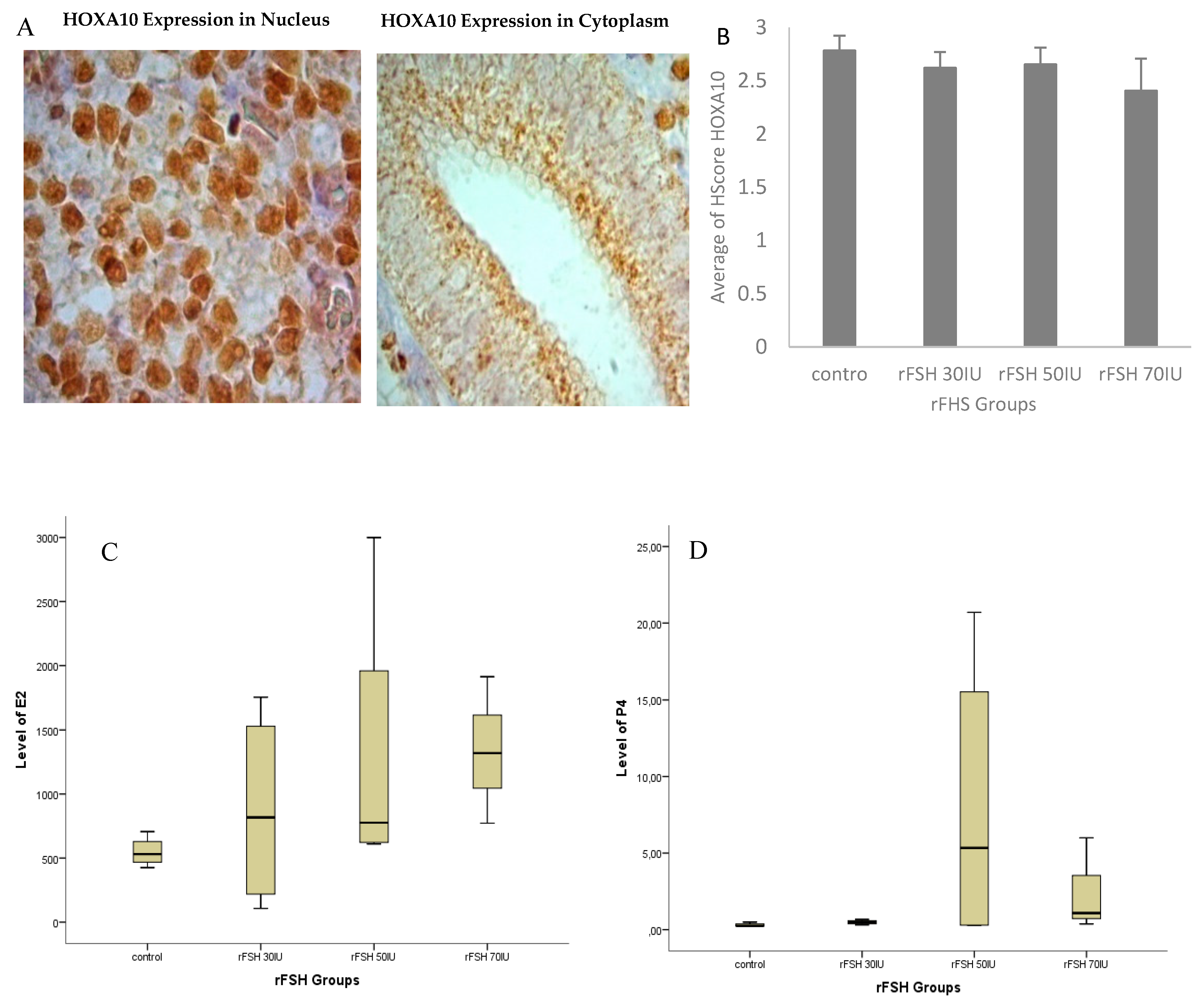
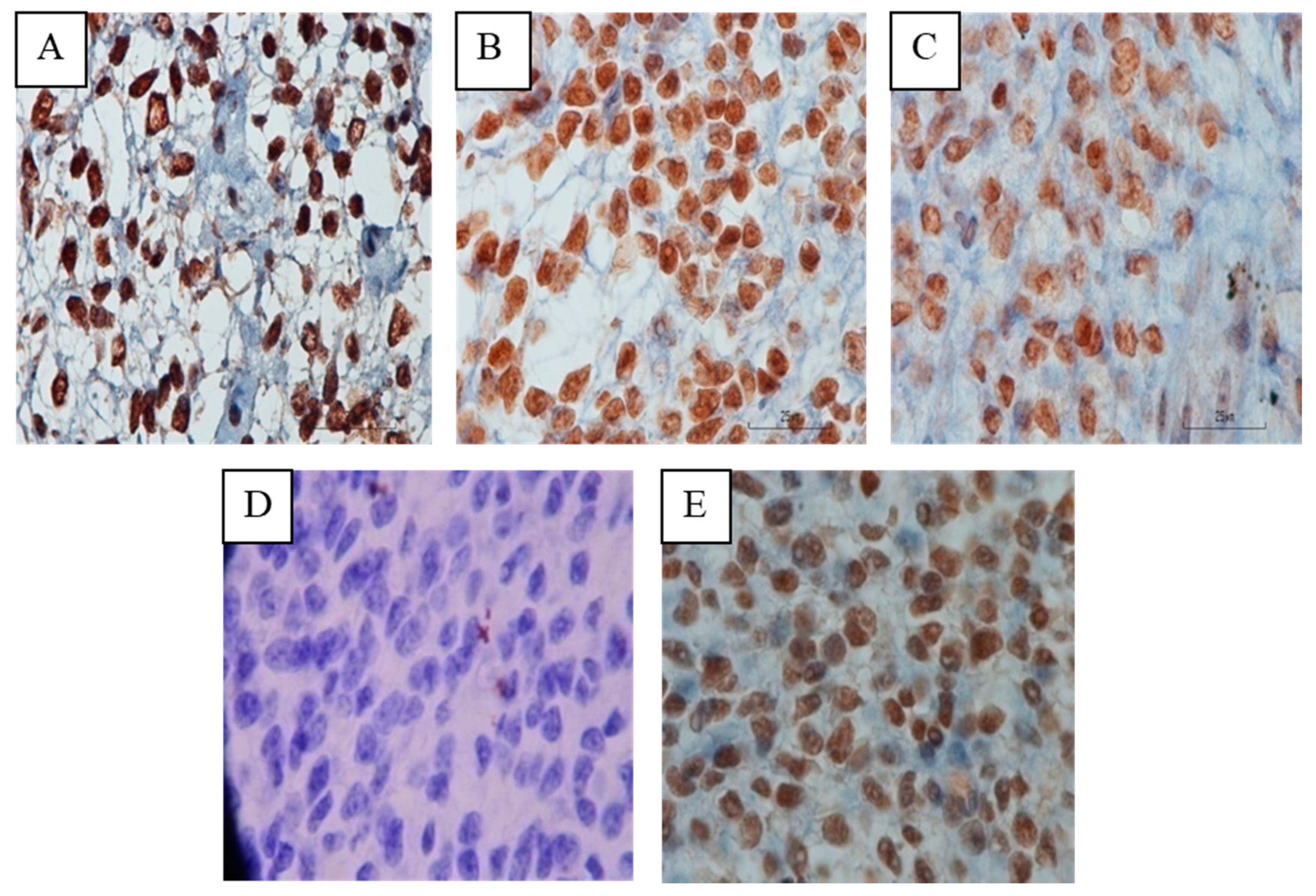
3.1. Location of HOXA10 Expression in Macaca nemestrina Endometrium
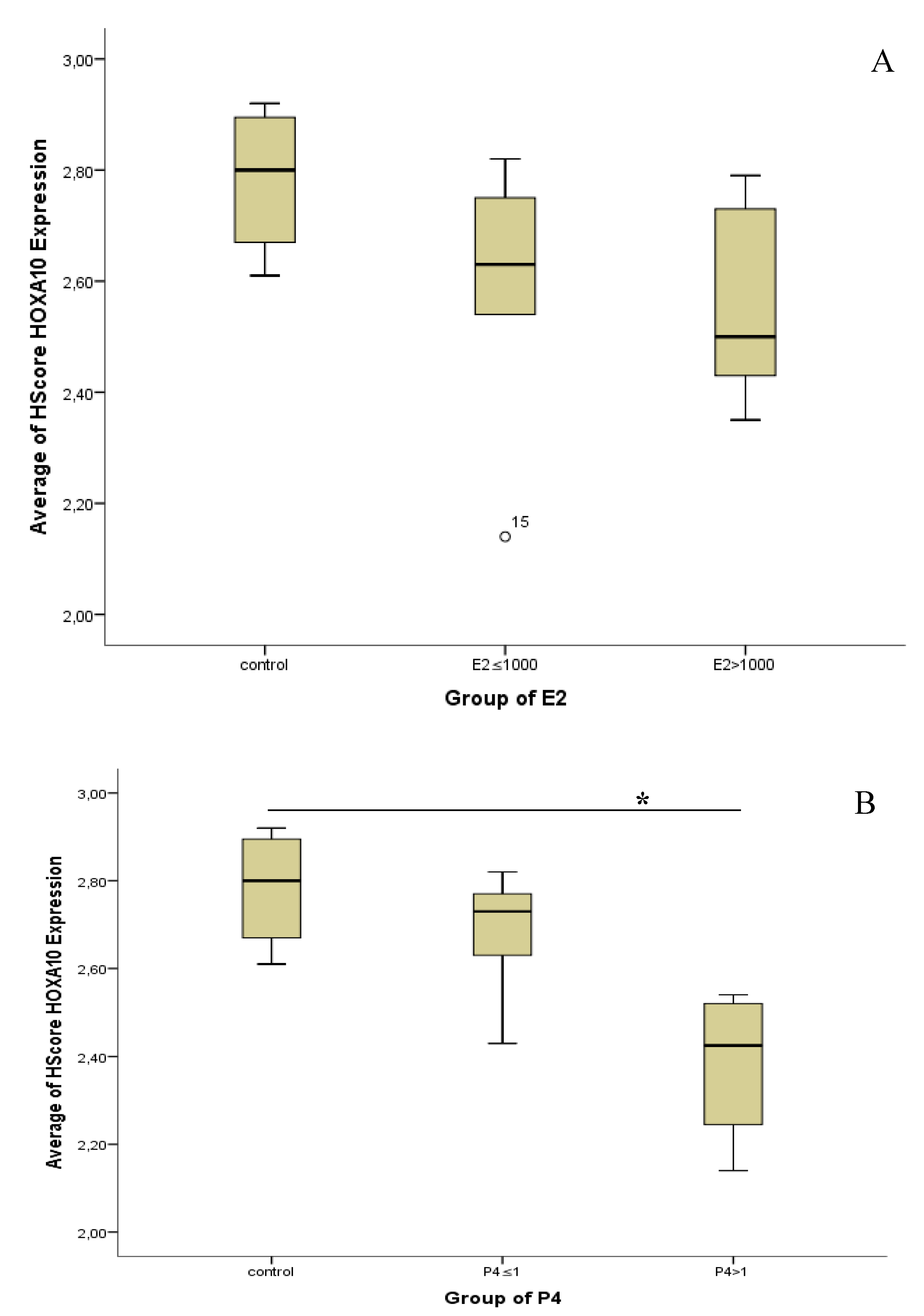
3.2. Hormone Serum Progesterone and HOXA10 Hscore Levels in Natural Cycles and Stimulated Cycles
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Al-Inany, H.G.; Abou-Setta, A.M.; Aboulghar, M. Gonadotrophin-releasing hormone antagonists for assisted conception: A Cochrane review. Reprod. Biomed. Online 2007, 14, 640–649. [Google Scholar] [CrossRef]
- Kolibianakis, E.; Bourgain, C.; Albano, C. Effect of ovarian stimulation with recombinant follicle-stimulating hormone, gonadotropin-releasing hormone antagonists, and humanchorionic gonadotropin on endometrial maturation on the day of oocyte pick-up. Fertil. Steril. 2002, 78, 1025–1029. [Google Scholar] [CrossRef]
- Bosch, E.; Valencia, I.E.; Escudero, E.; Crespo, J.; Simon, C.; Remohi, J.; Pellicer, A. Premature luteinization during gonadotropin-releasing hormone antagonist cycles and its relationship with in vitro fertilization outcome. Fertil. Steril. 2003, 80, 1444–1449. [Google Scholar] [CrossRef]
- Bosch, E.; Labarta, E.; Crespo, J.; Simon, C.; Remohi, J.; Jenkins, J. Circulating progesterone levels and ongoing pregnancy rates in controlled ovarian stimulation cycles for in vitro fertilization: Analysis of over 4000 cycles. Hum. Reprod. 2010, 25, 2092–2100. [Google Scholar] [CrossRef]
- Achsenkun, R.; Arzberger, A.; von Schonfeldt, V.; Gallwas, J.; Rogenhofer, N.; Crispin, A. Subtle progesterone rise on the day of human chorionic gonadotropin administration is associated with lower live birth rates in women undergoing assisted reproductive technology: A retrospective study with 2,555 fresh embryo transfers. Fertil. Steril. 2012, 98, 347–354. [Google Scholar] [CrossRef]
- Lahoud, R.; Kwik, M.; Ryan, J.; Al-Jefout, M.; Foley, J.; Illingworth, P. Elevated progesterone in GnRH agonist downregulated in vitro fertilization (IVFICSI) cycles reduces live birth rates but not embryo quality. Arch. Gynecol. Obstet. 2012, 285, 535–540. [Google Scholar] [CrossRef]
- Yding, A.C.; Bungum, L.; Nyboe, A.A.; Humaidan, P. Preovulatory progesterone concentration associates significantly to follicle number and LH concentration but not to pregnancy rate. Reprod. Biomed. Online 2011, 23, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Labarta, E.; Martinez-Conejoro, J.A.; Alama, P.; Harcojadas, J.A.; Pellicer, A.; Simon, C.; Bosch, E. Endometrial receptivity is affected in women with high circulating progesterone levels at the end of the follicular phase: A functional genomics analysis. Hum. Reprod. 2011, 26, 1813–1825. [Google Scholar] [CrossRef]
- Ashamita, J.; Vikas, S.; Swaiti, G. Impact of Progesterone Level on Day of hCG Injection in IVF Cycles on Clinical Pregnancy Rate. J. Hum. Reprod. Sci. 2017, 10, 265–270. [Google Scholar] [CrossRef]
- Amin, K.V.; Nadkarni, P.; Singh, P.N.; Singh, P. Serum progesterone levels on the day of hCG trigger and ICSI outcome: A retrospective observational cohort study. Int. J. Reprod. Contracept. Obs. Gynecol. 2018, 7, 3194–3198. [Google Scholar] [CrossRef]
- Venetis, C.A.; Kolibianakis, E.M.; Papanikolaou, E.; Bontis, J.; Devroey, P.; Tarlatzis, B.C. Is progesterone elevation on the day of human chorionic gonadotrophin administration associated with the probability of pregnancy in vitro fertilization? A systematic review and meta-analysis. Hum. Reprod. Update 2007, 13, 343–355. [Google Scholar] [CrossRef]
- Kiliçdag, E.B.; Haydardedeoglu, B.; Cok, T.; Hacivelioglu, S.O.; Bagis, T. Premature progesterone elevation impairs implantation and live birth rates in GnRH-agonist IVF/ICSI cycles. Arch. Gynecol. Obs. 2010, 281, 747–752. [Google Scholar] [CrossRef]
- Taylor, H.S. The role of HOX genes in human implantation. Hum. Reprod. Update 2000, 6, 75–79. [Google Scholar] [CrossRef]
- Wu, Y.; Halverson, G.; Basir, Z. Aberrant methylation at HOXA10 may be responsible for its aberrant expression in the endometrium of patients with endometriosis. Am. J. Obs. Gynecol. 2005, 193, 371–380. [Google Scholar] [CrossRef]
- Bagot, C.N.; Troy, P.J.; Taylor, H.S. Alteration of maternal HOXA10 expression by in vivo gene transfection affects implantation. Gene 2000, 7, 1378–1384. [Google Scholar] [CrossRef]
- Daftary, G.S.; Troy, P.J.; Bagot, C.N.; Young, S.L.; Taylor, H.S. Direct regulation of β3-integrin subunit gene expression by HOXA10 in endometrial cells. Mol. Endocrinol. 2002, 16, 571–579. [Google Scholar] [CrossRef]
- Rackow, B.W.; Jorgensen, E.; Taylor, H.S. Endometrial polyps affect uterine receptivity. Fertil. Steril. 2011, 95, 2690–2692. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, S.; Canis, M.; Darcha, C.; Pouly, J.L.; Mage, G. HOXA-10 expression in the mid-secretory endometrium of infertile patients with either endometriosis, uterine fibromas or unexplained infertility. Hum. Reprod. 2009, 24, 3180–3187. [Google Scholar] [CrossRef]
- Browne, H.; Taylor, H.S. HOXA10 expression in ectopic endometrial tissue. Fertil. Steril. 2006, 85, 1386–1390. [Google Scholar] [CrossRef]
- Paria, B.C.; Reese, J.; Das, S.K.; Dey, S.K. Deciphering the cross-talk of implantation: Advances and challenges. Science 2002, 296, 2185–2188. [Google Scholar] [CrossRef]
- Modi, D.; Godbole, M. HOXA10 signals on the highway through pregnancy. J. Reprod. Immunol. 2009, 83, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Elter, K.; Kavak, Z.N.; Gokasian, H. Antral follicle assessment after down-regulation may be a useful tool for predicting pregnancy loss in vitro fertilization pregnancies. Gynecol. Endocrinol. 2005, 21, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Lunenfeld, B. What’s new in ovarian stimulation? Life 2006, 14, 25. [Google Scholar]
- Taylor, H.S.; Daftary, G.S.; Selam, B. Endometrial HOXA10 expression after controlled ovarian hyperstimulation with the recombinant follicle-stimulating hormone. Fertil. Steril. 2003, 8, 839–843. [Google Scholar] [CrossRef]
- Miller, P.B.; Parnell, B.A.; Bushnell, G.; Tallman, N.; Forstein, D.A.; Higdon, H.L.; Lessey, B.A. Endometrial receptivity defects during IVF cycles with and without letrozole. Hum. Reprod. 2012, 27, 881–888. [Google Scholar] [CrossRef] [Green Version]
- Elnashar, A.M. Progesterone rise on the day of HCG administration (premature luteinization) in IVF; an overdue update. J. Assist. Reprod. Genet. 2010, 27, 149–155. [Google Scholar] [CrossRef]
- Kyrou, D.; Al-Azemi, M.; Papanikolaou, E.G.; Donoso, P.; Tziomalos, K.; Devroey, P. The The relationship of premature progesterone rise with serum estradiol levels and number of follicles in GnRH antagonist/recombinant FSH-stimulated cycles. Eur. J. Obstet. Gynecol. Reprod. Biol. 2012, 162, 165–168. [Google Scholar] [CrossRef]
- Papanikolaou, E.G.; Pados, G.; Grimbizis, G.; Bili, E.; Kyriazi, L.; Polyzos, N.P. GnRH-agonist versus GnRH-antagonist IVF cycles: Is the reproductive outcome affected by the incidence of progesterone elevation on the day of HCG triggering? A randomized prospective study. Hum. Reprod. 2012, 27, 1822–1828. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahar, N.; Mujihartini, N.; Pudjianto, D.A.; Pradhita, A.D.; Thuffi, R.; Kusmardi, K. Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity. Biomedicines 2019, 7, 83. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines7040083
Sahar N, Mujihartini N, Pudjianto DA, Pradhita AD, Thuffi R, Kusmardi K. Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity. Biomedicines. 2019; 7(4):83. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines7040083
Chicago/Turabian StyleSahar, Nurhuda, Ninik Mujihartini, Dwi Ari Pudjianto, Adhea Debby Pradhita, Rosalina Thuffi, and Kusmardi Kusmardi. 2019. "Increased Progesterone on the Day of Administration of hCG in Controlled Ovarian Hyperstimulation Affects the Expression of HOXA10 in Primates’ Endometrial Receptivity" Biomedicines 7, no. 4: 83. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines7040083