Immunogenicity and Protective Effect of a Virus-Like Particle Containing the SAG1 Antigen of Toxoplasma gondii as a Potential Vaccine Candidate for Toxoplasmosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Preparation of Parasites and Cell Culture
2.4. Toxoplasma Gondii Antigen
2.5. The Expression of T. gondii-Specific Antigen and Influenza A virus Matrix
2.6. The Production and Preparation of Recombinant Baculovirus (rBV)
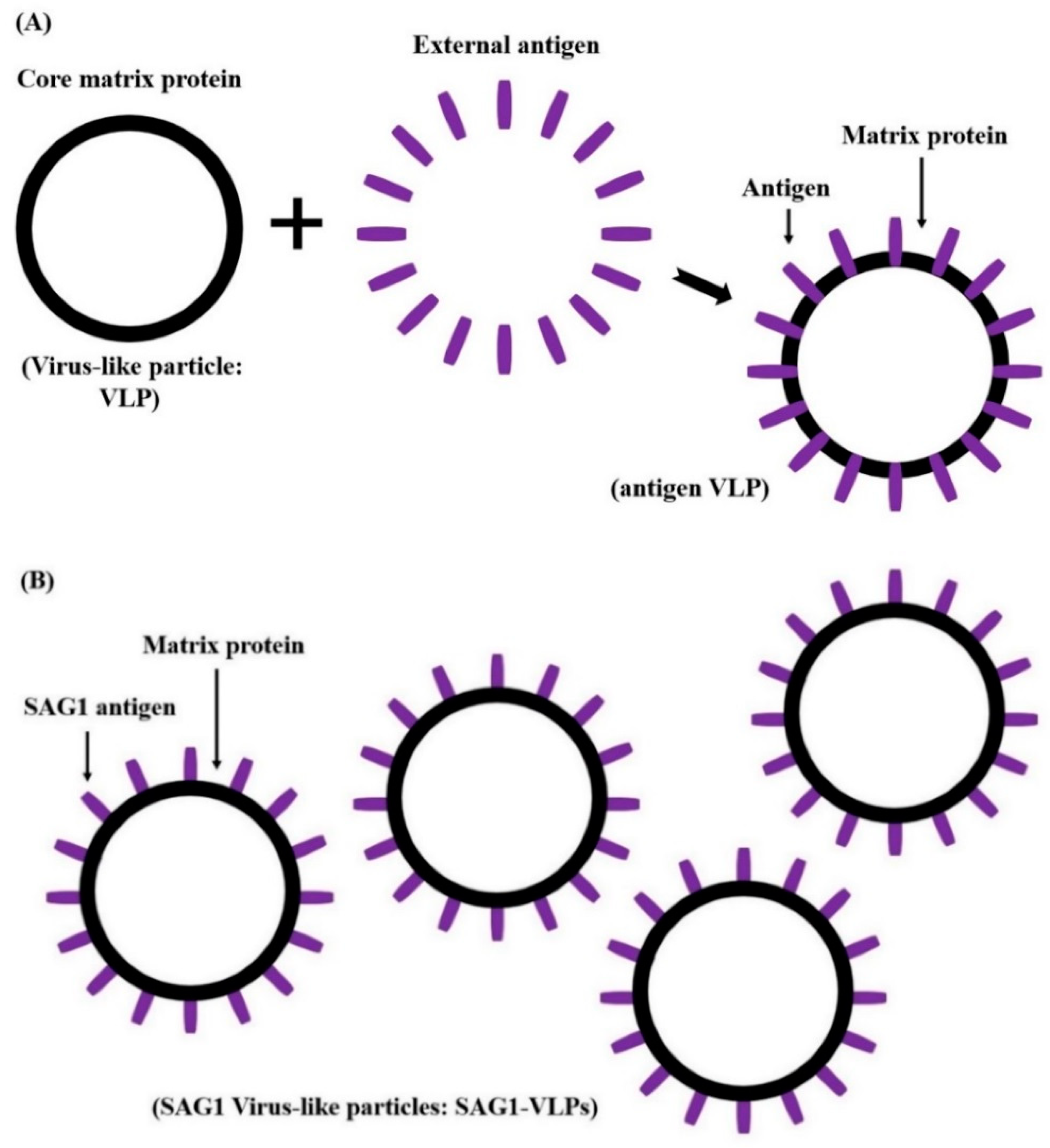
2.7. The Expression and Culture of Virus-Like Particles
2.8. The Evaluation of VLPs through Western Blot Analysis
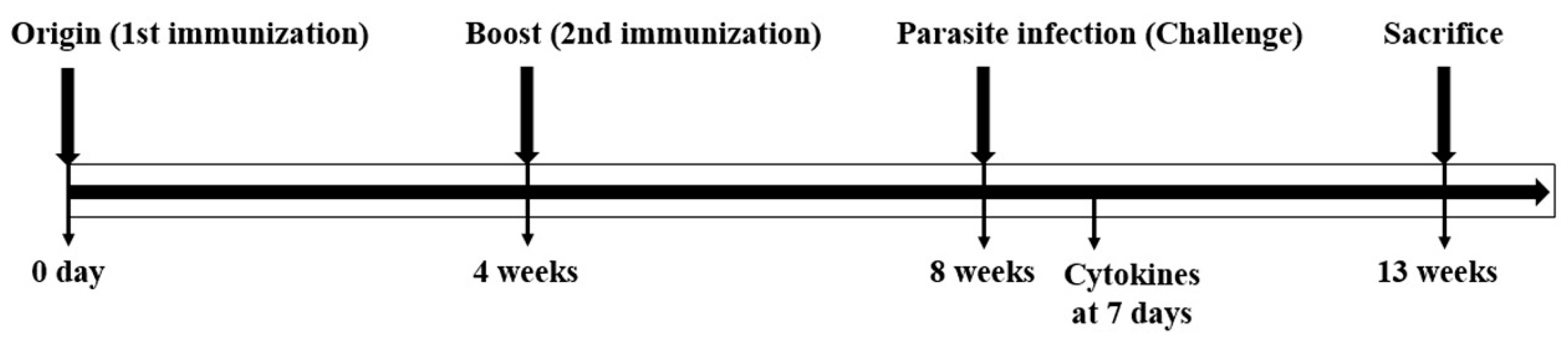
2.9. VLP Immunization and Parasite Infection
2.10. The Evaluation of Antibody Responses in Sera
2.11. Cytokine Analysis in the Spleen
2.12. Evaluation of the Proliferation of T. gondii through Immunized Sera
2.13. The Viability of Splenocytes after Immunization
2.14. Survival Rate of Mice through T. gondii Infection
2.15. Statistical Analysis
3. Results
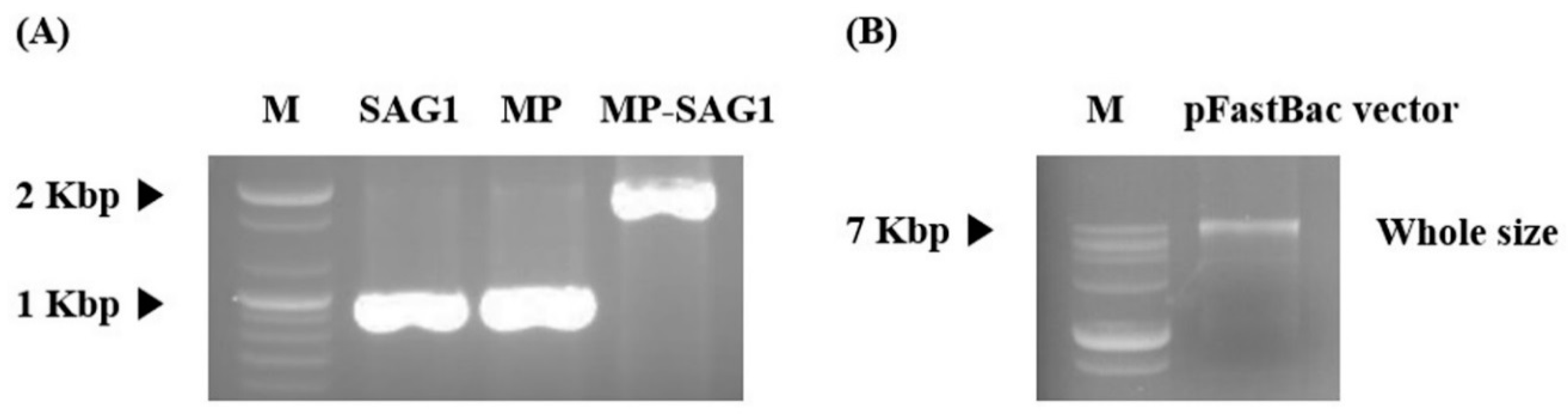
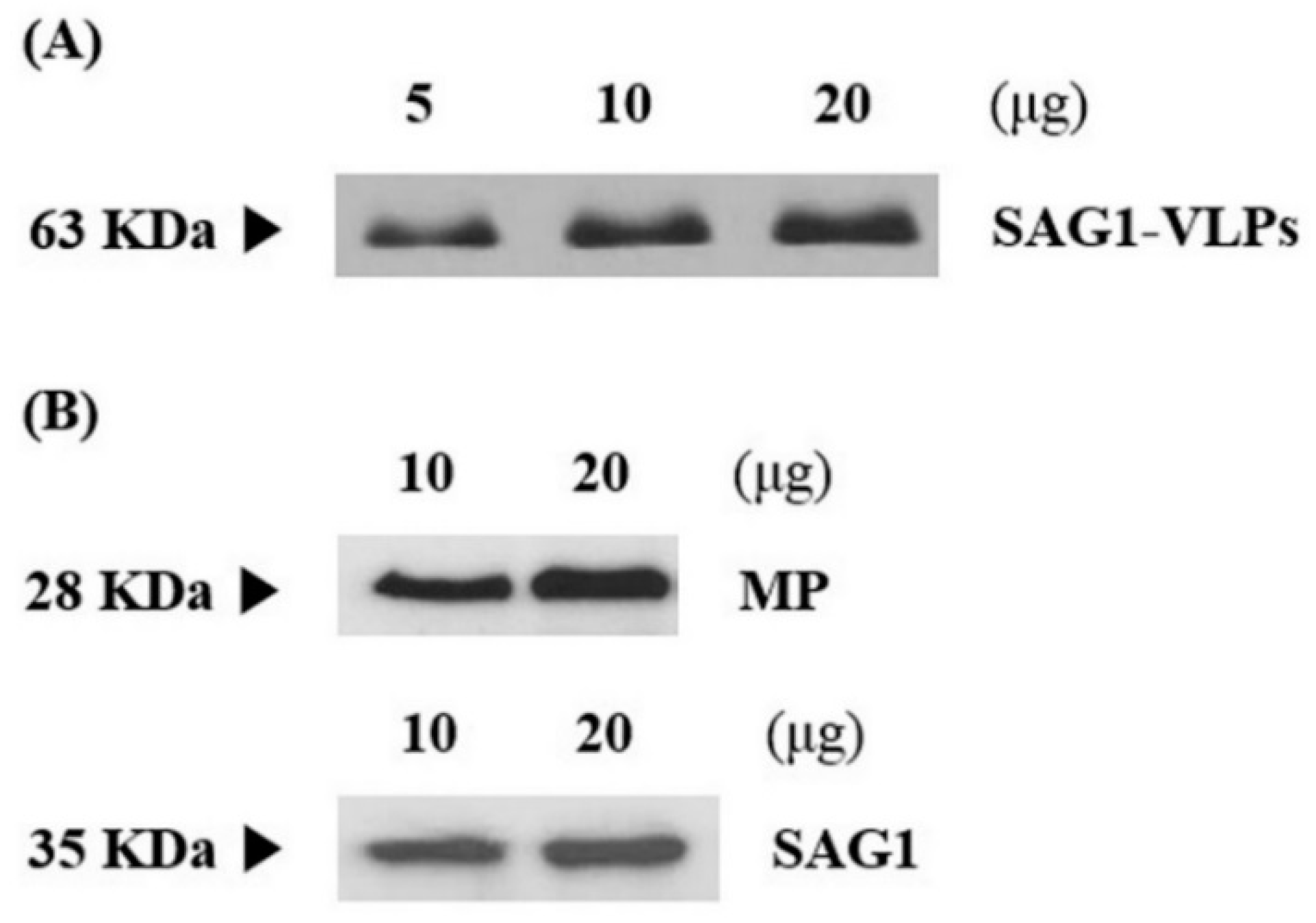
3.1. Production and Expression of T. gondii-Specific SAG1-VLPs
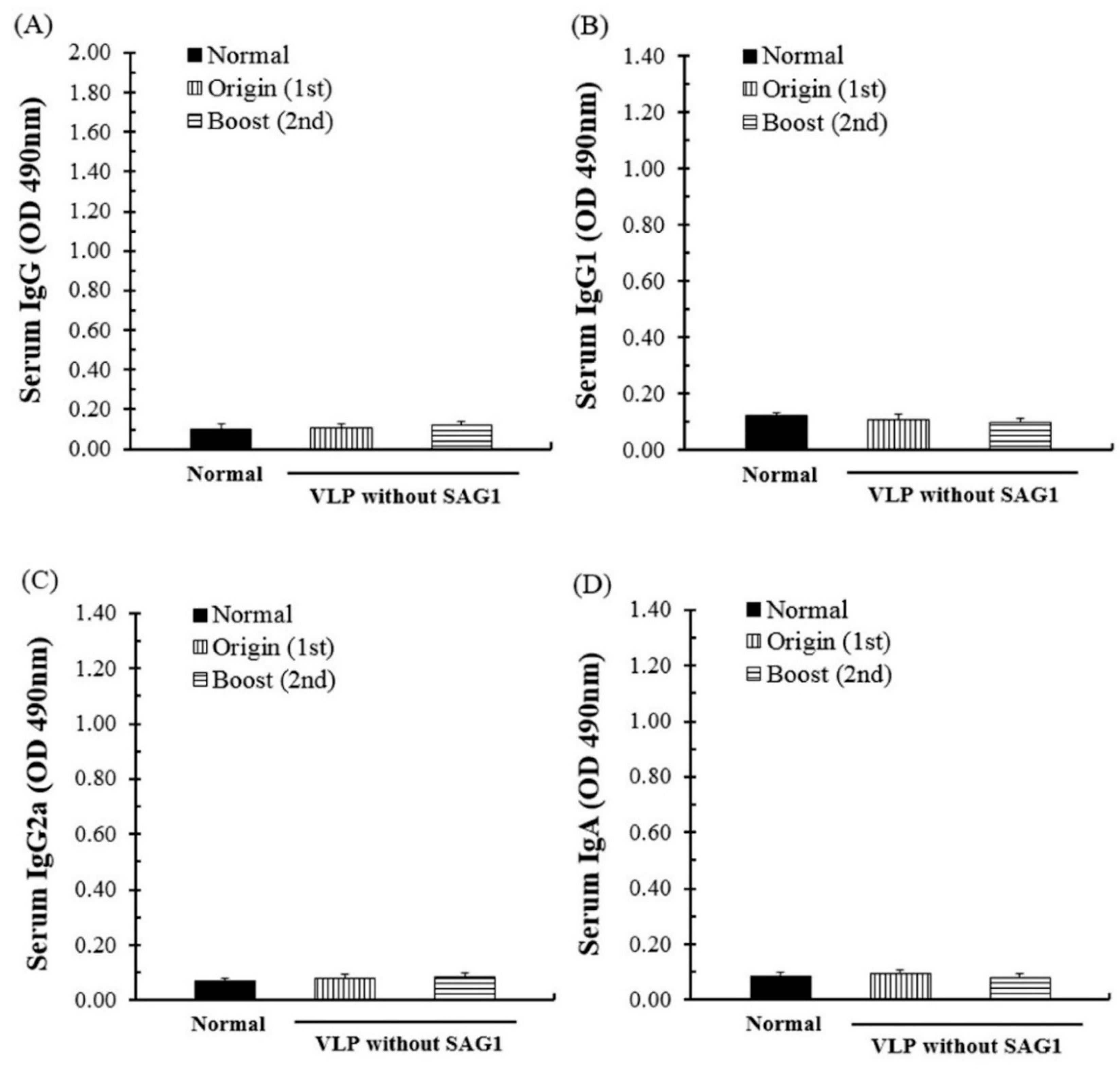
3.2. Evaluation of the Antibody Response through SAG1-VLP Immunization
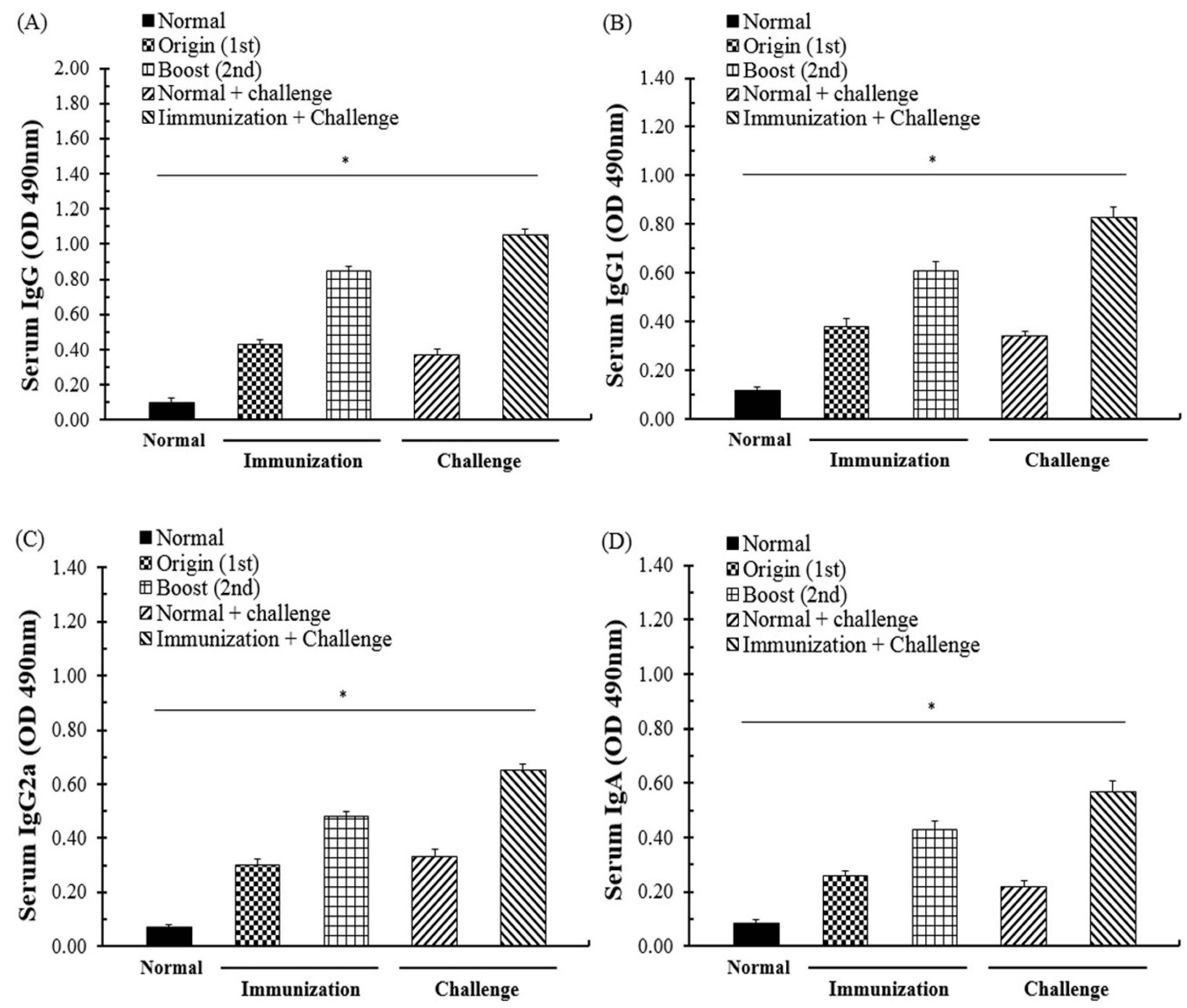
3.3. Specific Antibody Response through Immunization and T. gondii Infection
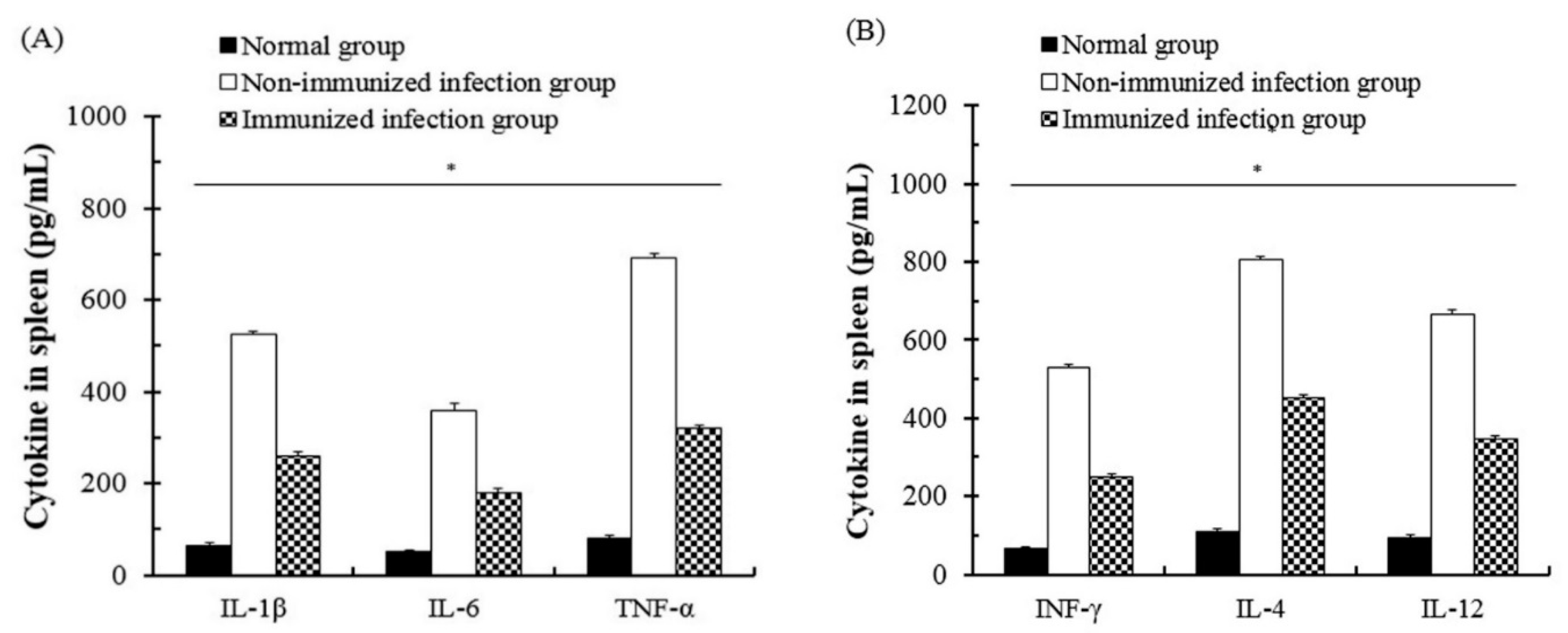
3.4. The Change of Cytokines of Immune Cells after T. gondii Infection
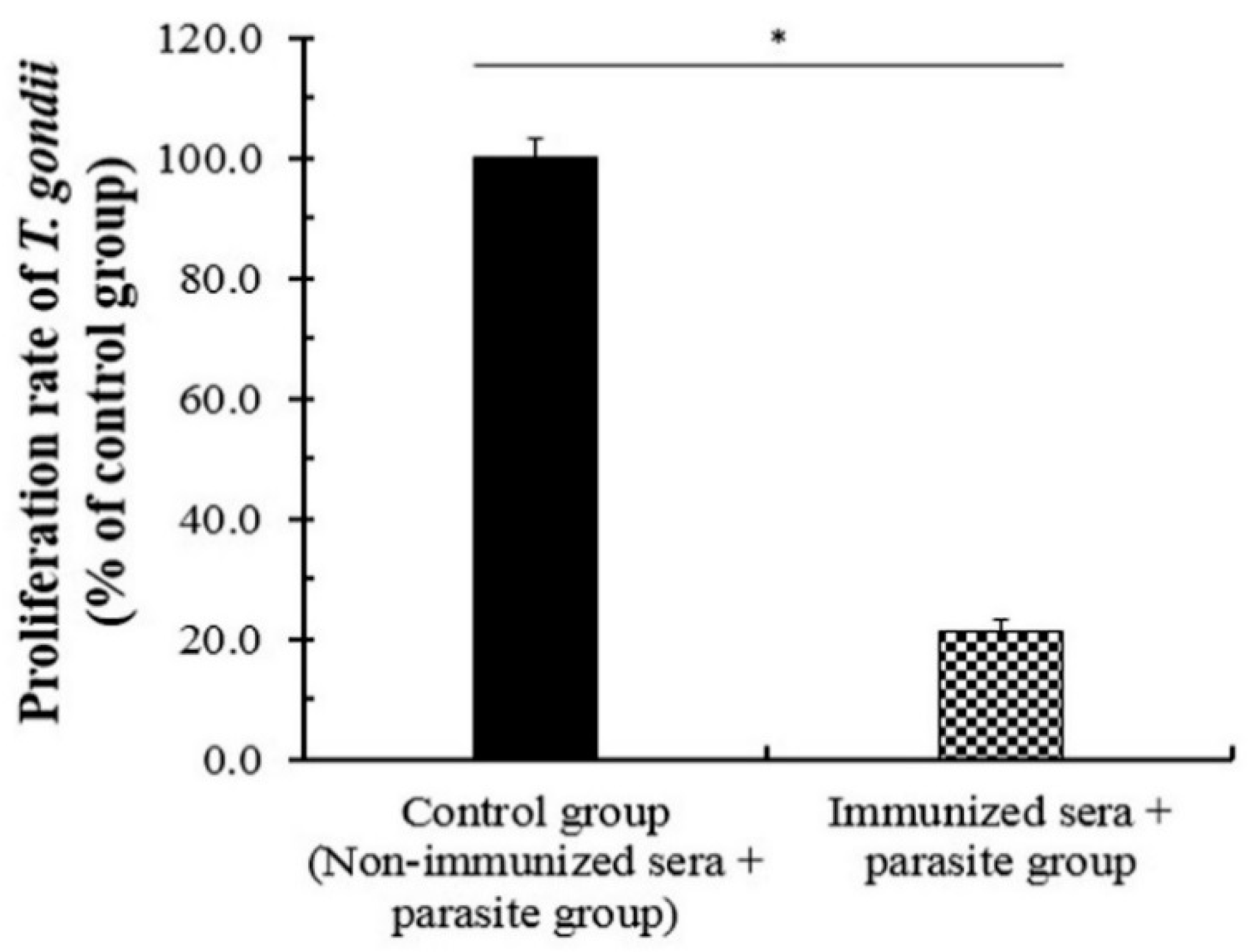
3.5. The Neutralizing Effect of the Immunized Sera against T. gondii
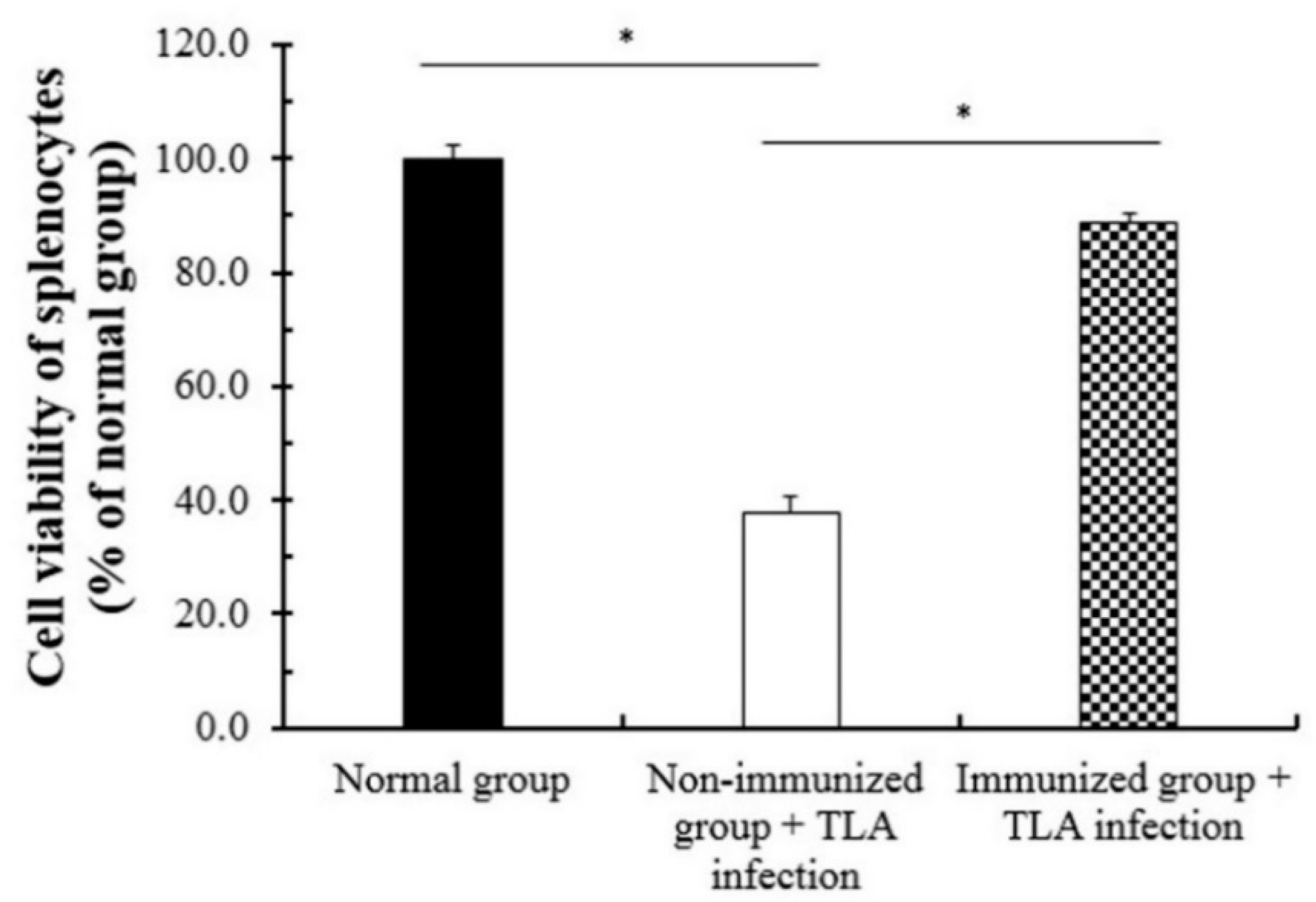
3.6. Cell Viability of Splenocytes after the Immunization of SAG1-VLPs
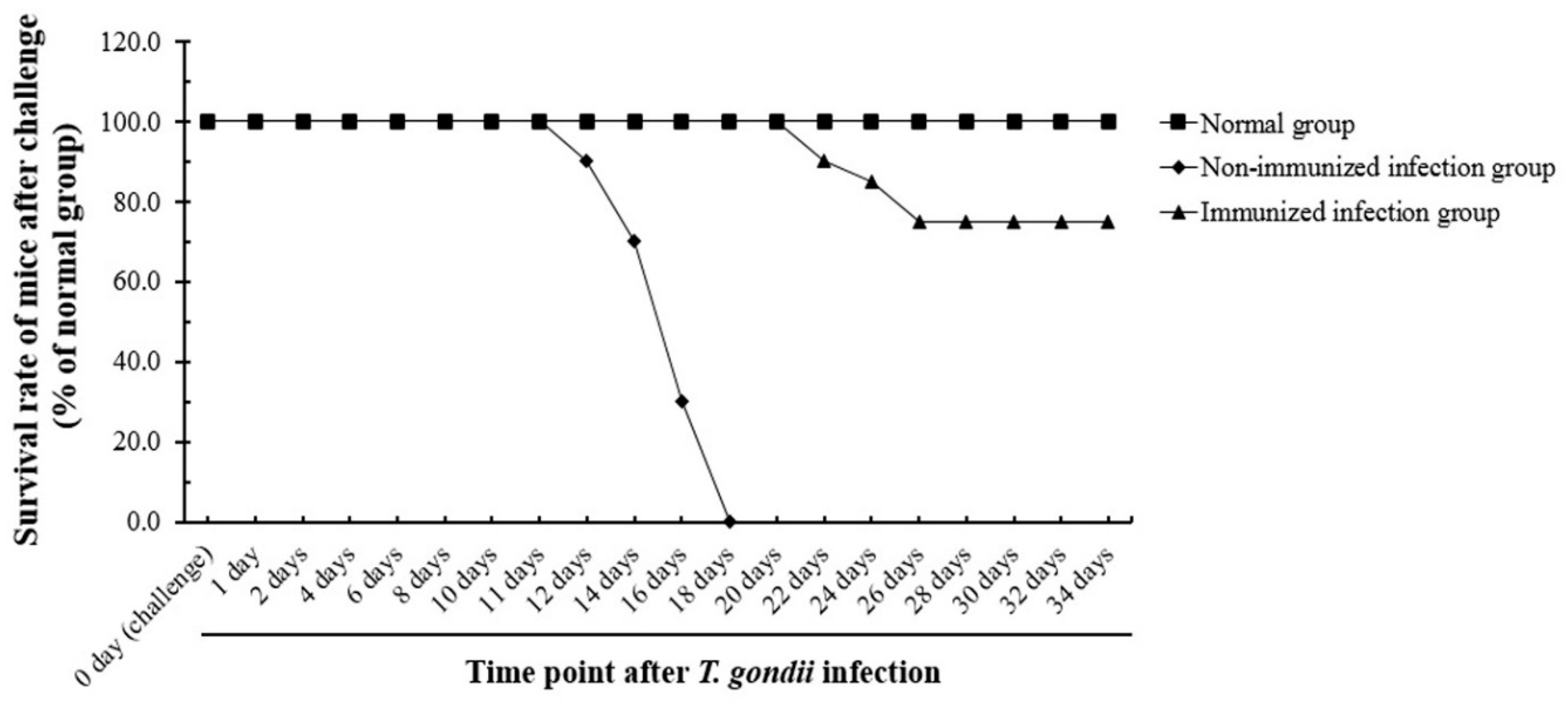
3.7. Evaluation of the Survival Rate of Immunized Mice after T. gondii Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Choi, W.H.; Lee, I.A. The Mechanism of Action of Ursolic Acid as a Potential Anti-Toxoplasmosis Agent, and Its Immunomodulatory Effects. Pathogens 2019, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, R.; Harrison, B.; Dangoudoubiyam, S.; Bandini, G.; Cheng, K.; Kosber, A.; Agop-Nersesian, C.; Howe, D.K.; Samuelson, J.; Ferguson, D.J.P.; et al. Differential Roles for Inner Membrane Complex Proteins across Toxoplasma gondii and Sarcocystis neurona Development. mSphere 2017, 2, e00409-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.R.; Meissner, M. The inner membrane complex throughdevelopment of Toxoplasma gondii and Plasmodium. Cell. Microbiol. 2014, 16, 632–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, J.R.; Rodriguez-Fernandez, I.A.; de Leon, J.C.; Huynh, M.H.; Carruthers, V.B.; Morrissette, N.S.; Bradley, P.J. A novel family of Toxoplasma IMCproteins displays a hierarchical organization and functions in coordinatingparasite division. PLoS Pathog. 2010, 6, e1001094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.H.; Lee, I.A. Evaluation of Anti-Toxoplasma gondii Effect of Ursolic Acid as a Novel Toxoplasmosis Inhibitor. Pharmaceuticals 2018, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Rome, M.E.; Beck, J.R.; Turetzky, J.M.; Webster, P.; Bradley, P.J. Intervacuolar Transport and Unique Topology of GRA14, a Novel Dense Granule Protein in Toxoplasma gondii. Infect. Immun. 2008, 76, 4865–4875. [Google Scholar] [CrossRef] [Green Version]
- Bonhomme, A.; Bouchot, A.; Pezzella, N.; Gomez, J.; Le, M.H.; Pinon, J.M. Signaling during the invasion of host cells by Toxoplasma gondii. FEMS Microbiol. Rev. 1999, 23, 551–561. [Google Scholar] [CrossRef]
- Munera Lopez, J.; Ganuza, A.; Bogado, S.S.; Muñoz, D.; Ruiz, D.M.; Sullivan, W.J., Jr.; Vanagas, L.; Angel, S.O. Evaluation of ATM Kinase Inhibitor KU-55933 as Potential Anti-Toxoplasma gondii Agent. Front. Cell. Infect. Microbiol. 2019, 9, 26. [Google Scholar] [CrossRef] [Green Version]
- Moine, E.; Moire, N.; Dimier-Poisson, I.; Brunet, K.; Couet, W.; Colas, C.; Van Langendonck, N.; Enguehard-Gueiffier, C.; Gueiffier, A.; Heraut, B.; et al. Imidazo[1,2-b]pyridazines targeting Toxoplasma gondii calcium-dependent protein kinase 1 decrease the parasite burden in mice with acute toxoplasmosis. Int. J. Parasitol. 2018, 48, 561–568. [Google Scholar] [CrossRef]
- Si, H.; Xu, C.; Zhang, J.; Zhang, X.; Li, B.; Zhou, X.; Zhang, J. Licochalcone A: An effective and low-toxicity compound against Toxoplasma gondii in vitro and in vivo. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 238–245. [Google Scholar] [CrossRef]
- Silveira, G.R.; Campelo, K.A.; Lima, G.R.S.; Carvalho, L.P.; Samarao, S.S.; Vieira-da-Motta, O.; Mathias, L.; Matos, C.R.R.; Vieira, I.J.C.; Melo, E.J.T.; et al. In Vitro Anti-Toxoplasma gondii and Antimicrobial Activity of Amides Derived from Cinnamic Acid. Molecules 2018, 23, 774. [Google Scholar] [CrossRef] [Green Version]
- Alomar, M.L.; Rasse-Suriani, F.O.; Ganuza, A.; Coceres, V.M.; Cabrerizo, F.M.; Angel, S.O. In vitro evaluation of b-carboline alkaloids as potential anti-Toxoplasma agents. BMC Res. Notes 2013, 6, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavitha, N.; Noordin, R.; Chan, K.L.; Sasidharan, S. In vitro anti-Toxoplasma gondii activity of root extract/fractions of Eurycoma longifolia Jack. BMC Complement. Altern. Med. 2012, 12, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Z.Z.; Ho, Y.J.; Lu, J.W. Cephalotaxine inhibits Zika infection by impeding viral replication and stability. Biochem. Biophys. Res. Commun. 2020, 522, 1052–1058. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lee, E.M.; Wen, Z.; Cheng, Y.; Huang, W.K.; Qian, X.; Tcw, J.; Kouznetsova, J.; Ogden, S.C.; Hammack, C.; et al. Identification of small-molecule inhibitors of Zika virus infection and induced neural cell death via a drug repurposing screen. Nat. Med. 2016, 22, 1101–1107. [Google Scholar] [CrossRef]
- Tonk, M.; Pierrot, C.; Cabezas-Cruz, A.; Rahnamaeian, M.; Khalife, J.; Vilcinskas, A. The Drosophila melanogaster antimicrobial peptides Mtk-1 and Mtk-2 are active against the malarial parasite Plasmodium falciparum. Parasitol. Res. 2019, 118, 1993–1998. [Google Scholar] [CrossRef]
- Szamosvari, D.; Sylvester, K.; Schmid, P.; Lu, K.Y.; Derbyshire, E.R.; Bottcher, T. Close the ring to break the cycle: Tandem quinolone-alkyne-cyclisation gives access to tricyclic pyrrolo[1,2-a]quinolin-5-ones with potent anti-protozoal activity. Chem. Commun. 2019, 55, 7009–7012. [Google Scholar] [CrossRef]
- Choi, W.H. Novel pharmacological activity of artesunate and artemisinin: Their potential as anti-tubercular agents. J. Clin. Med. 2017, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Perez-Pertejo, Y.; Escudero-Martinez, J.M.; Reguera, R.M.; Balana-Fouce, R.; Garcia, P.A.; Jambrina, P.G.; San Feliciano, A.; Castro, M.A. Antileishmanial activity of terpenylquinones on Leishmania infantum and their effects on Leishmania topoisomerase IB. Int. J. Parasitol. Drugs Drug Resist. 2019, 11, 70–79. [Google Scholar] [CrossRef]
- Choi, W.H. Evaluation of anti-tubercular activity of linolenic acid and conjugated-linoleic acid as effective inhibitors against Mycobacterium tuberculosis. Asian Pac. J. Trop. Med. 2016, 9, 125–129. [Google Scholar] [CrossRef]
- Zhang, W.; Bailey-Elkin, B.A.; Knaap, R.C.M.; Khare, B.; Dalebout, T.J.; Johnson, G.G.; van Kasteren, P.B.; McLeish, N.J.; Gu, J.; He, W.; et al. Potent and selective inhibition of pathogenic viruses by engineered ubiquitin variants. PLoS Pathog. 2017, 13, e1006372. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W.; Rosenbloom, R.; Petteruti, M.; Hilt, D.A.; McCall, A.W.; Williams, S.M. Avian coronavirus infectious bronchitis virus susceptibility to botanical oleoresins and essential oils in vitro and in vivo. Virus Res. 2010, 149, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Kudchodkar, S.B.; Reuschel, E.L.; Asija, K.; Borole, P.; Agarwal, S.; Van Gorder, L.; Reed, C.C.; Gulendran, G.; Ramos, S.; et al. Synthetic nucleic acid antibody prophylaxis confers rapid and durable protective immunity against Zika virus challenge. Hum. Vaccin. Immunother. 2019, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hagan, T.; Cortese, M.; Rouphael, N.; Boudreau, C.; Linde, C.; Maddur, M.S.; Das, J.; Wang, H.; Guthmiller, J.; Zheng, N.Y.; et al. Antibiotics-driven gut microbiome perturbation alters immunity to vaccines in humans. Cell 2019, 178, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Hook, L.M.; Pardi, N.; Wang, F.; Myles, A.; Cancro, M.P.; Cohen, G.H.; Weissman, D.; Friedman, H.M. Nucleoside-modified mRNA encoding HSV-2 glycoproteins C, D, and E prevents clinical and subclinical genital herpes. Sci. Immunol. 2019, 4, eaaw7083. [Google Scholar] [CrossRef] [PubMed]
- Mateo, M.; Reynard, S.; Carnec, X.; Journeaux, A.; Baillet, N.; Schaeffer, J.; Picard, C.; Legras-Lachuer, C.; Allan, R.; Perthame, E.; et al. Vaccines inducing immunity to Lassa virus glycoprotein and nucleoprotein protect macaques after a single shot. Sci. Transl. Med. 2019, 11, eaaw3163. [Google Scholar] [CrossRef]
- Afroz, S.S.; Battu, S.; Matin, S.; Solouki, S.; Elmore, J.P.; Minhas, G.; Huang, W.; August, A.; Khan, N. Amino acid starvation enhances vaccine efficacy by augmenting neutralizing antibody production. Sci. Signal. 2019, 12, eaav4717. [Google Scholar] [CrossRef]
- Sok, D.; Burton, D.R. Recent progress in broadly neutralizing antibodies to HIV. Nat. Immunol. 2018, 19, 1179–1188. [Google Scholar] [CrossRef]
- Pegu, A.; Hessell, A.J.; Mascola, J.R.; Haigwood, N.L. Use of broadly neutralizing antibodies for HIV-1 prevention. Immunol. Rev. 2017, 275, 296–312. [Google Scholar] [CrossRef]
- Paquet, C.; Yudin, M.H. Society of Obstetricians and Gynaecologists of Canada. Toxoplasmosis in pregnancy: Prevention, screening, and treatment. J. Obstet. Gynaecol. Can. 2013, 35, 78–81. [Google Scholar] [CrossRef]
- Oz, H.S. Fetomaternal and Pediatric Toxoplasmosis. J. Pediatr. Infect. Dis. 2017, 12, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Hoshina, T.; Horino, T.; Saiki, E.; Aonuma, H.; Sawaki, K.; Miyajima, M.; Lee, K.; Nakaharai, K.; Shimizu, A.; Hosaka, Y.; et al. Seroprevalence and associated factors of Toxoplasma gondii among HIV-infected patients in Tokyo: A cross sectional study. J. Infect. Chemother. 2020, 26, 33–37. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Tang, Y.; Chen, L.; Cai, H.; Lai, D.H.; Liu, K.; Wan, L.; Gong, L.; Yu, L.; Luo, Q.; et al. Encephalitis is mediated by ROP18 of Toxoplasma gondii, a severe pathogen in AIDS patients. Proc. Natl. Acad. Sci. USA 2018, 115, E5344–E5352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blader, I.J.; Coleman, B.I.; Chen, C.T.; Gubbels, M.J. Lytic Cycle of Toxoplasma gondii: 15 Years Later. Annu. Rev. Microbiol. 2015, 69, 463–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naumov, A.; Kratzer, S.; Ting, L.M.; Kim, K.; Suvorova, E.S.; White, M.W. The Toxoplasma Centrocone Houses Cell Cycle Regulatory Factors. mBio 2017, 8, e00579-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhrif, Z.; Moreau, A.; Herault, B.; Di-Tommaso, A.; Juste, M.; Moire, N.; Dimier-Poisson, I.; Mevelec, M.N.; Aubrey, N. Targeted Delivery of Toxoplasma gondii Antigens to Dendritic Cells Promote Immunogenicity and Protective Efficiency against Toxoplasmosis. Front. Immunol. 2018, 9, 317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Li, Y.; Wang, M.; Xie, Q.; Li, P.; Zuo, S.; Kong, L.; Wang, C.; Wang, S. Immune Protection of Rhoptry Protein 21 (ROP21) of Toxoplasma gondii as a DNA Vaccine Against Toxoplasmosis. Front. Microbiol. 2018, 9, 909. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Liu, J.; Hao, P.; Wang, J.; Lei, T.; Shan, D.; Liu, Q. MIC3, a novel cross-protective antigen expressed in Toxoplasma gondii and Neospora caninum. Parasitol. Res. 2015, 114, 3791–3799. [Google Scholar] [CrossRef]
- Yang, L.; Liang, H.; Wang, B.; Ma, B.; Wang, J.; Zhang, W. Evaluation of the Potency of Two Pyolysin-Derived Recombinant Proteins as Vaccine Candidates of Trueperella Pyogenes in a Mouse Model: Pyolysin Oligomerization and Structural Change Affect the Efficacy of Pyolysin-Based Vaccines. Vaccines 2020, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Dziadek, B.; Gatkowska, J.; Grzybowski, M.; Dziadek, J.; Dzitko, K.; Dlugonska, H. Toxoplasma gondii: The vaccine potential of three trivalent antigen-cocktails composed of recombinant ROP2, ROP4, GRA4 and SAG1 proteins against chronic toxoplasmosis in BALB/c mice. Exp. Parasitol. 2012, 131, 133–138. [Google Scholar] [CrossRef]
- Gatkowska, J.; Dzitko, K.; Ferra, B.T.; Holec-Gąsior, L.; Kawka, M.; Dziadek, B. The Impact of the Antigenic Composition of Chimeric Proteins on Their Immunoprotective Activity against Chronic Toxoplasmosis in Mice. Vaccines 2019, 7, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, G.; Zhou, J.; Zhou, A.; Han, Y.; Guo, J.; Song, P.; Zhou, H.; Cong, H.; Hou, M.; Wang, L.; et al. SAG5B and SAG5C combined vaccine protects mice against Toxoplasma gondii infection. Parasitol. Int. 2017, 66, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Graille, M.; Stura, E.A.; Bossus, M.; Muller, B.H.; Letourneur, O.; Battail-Poirot, N.; Sibai, G.; Gauthier, M.; Rolland, D.; Le Du, M.H.; et al. Crystal structure of the complex between the monomeric form of Toxoplasma gondii surface antigen 1 (SAG1) and a monoclonal antibody that mimics the human immune response. J. Mol. Biol. 2005, 354, 447–458. [Google Scholar] [CrossRef]
- Chuang, S.C.; Ko, J.C.; Chen, C.P.; Du, J.T.; Yang, C.D. Encapsulation of chimeric protein rSAG1/2 into poly(lactide-co-glycolide) microparticles induces long-term protective immunity against Toxoplasma gondii in mice. Exp. Parasitol. 2013, 134, 430–437. [Google Scholar] [CrossRef] [PubMed]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, W.H.; Park, J.S. Immunogenicity and Protective Effect of a Virus-Like Particle Containing the SAG1 Antigen of Toxoplasma gondii as a Potential Vaccine Candidate for Toxoplasmosis. Biomedicines 2020, 8, 91. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8040091
Choi WH, Park JS. Immunogenicity and Protective Effect of a Virus-Like Particle Containing the SAG1 Antigen of Toxoplasma gondii as a Potential Vaccine Candidate for Toxoplasmosis. Biomedicines. 2020; 8(4):91. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8040091
Chicago/Turabian StyleChoi, Won Hyung, and Ji Sun Park. 2020. "Immunogenicity and Protective Effect of a Virus-Like Particle Containing the SAG1 Antigen of Toxoplasma gondii as a Potential Vaccine Candidate for Toxoplasmosis" Biomedicines 8, no. 4: 91. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8040091