Inactivated Platelet Lysate Supports the Proliferation and Immunomodulant Characteristics of Mesenchymal Stromal Cells in GMP Culture Conditions

 ,
, 
Abstract
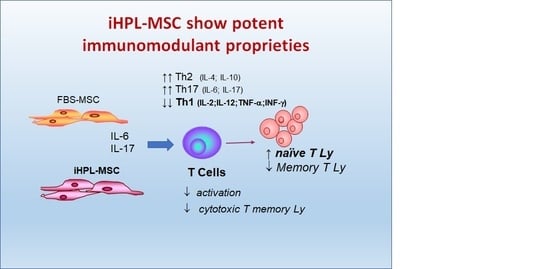
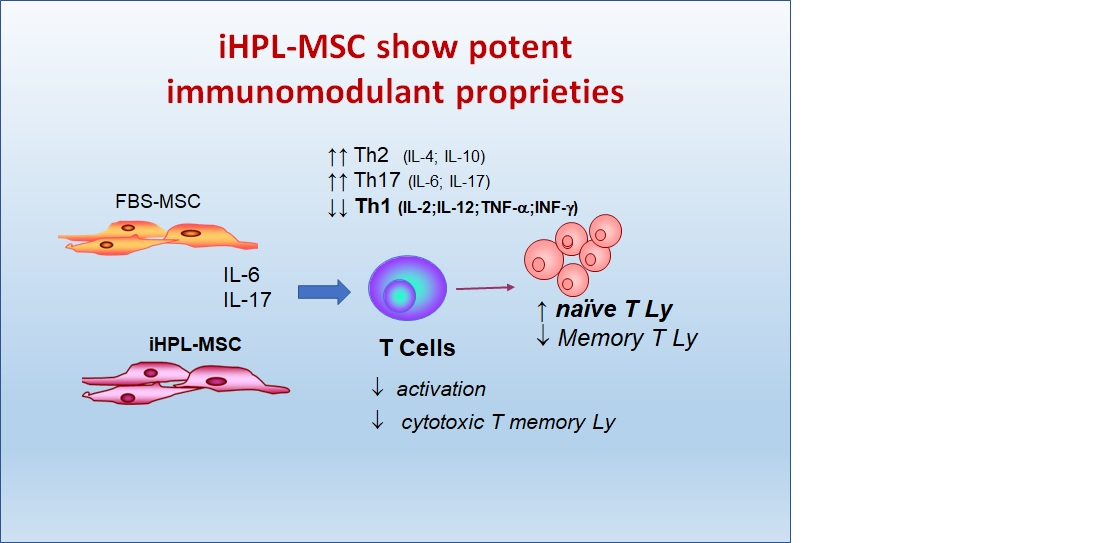
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
- the effect on the proliferation of the total Ly
- the effect on the proliferation of naïve T Ly subsets induced to Th1 and Th2 Ly differentiation
- the immunophenotype of different T cell subsets (naïve, memory, effector, and Th1 and Th2 lymphocytes)
2. Experimental Section
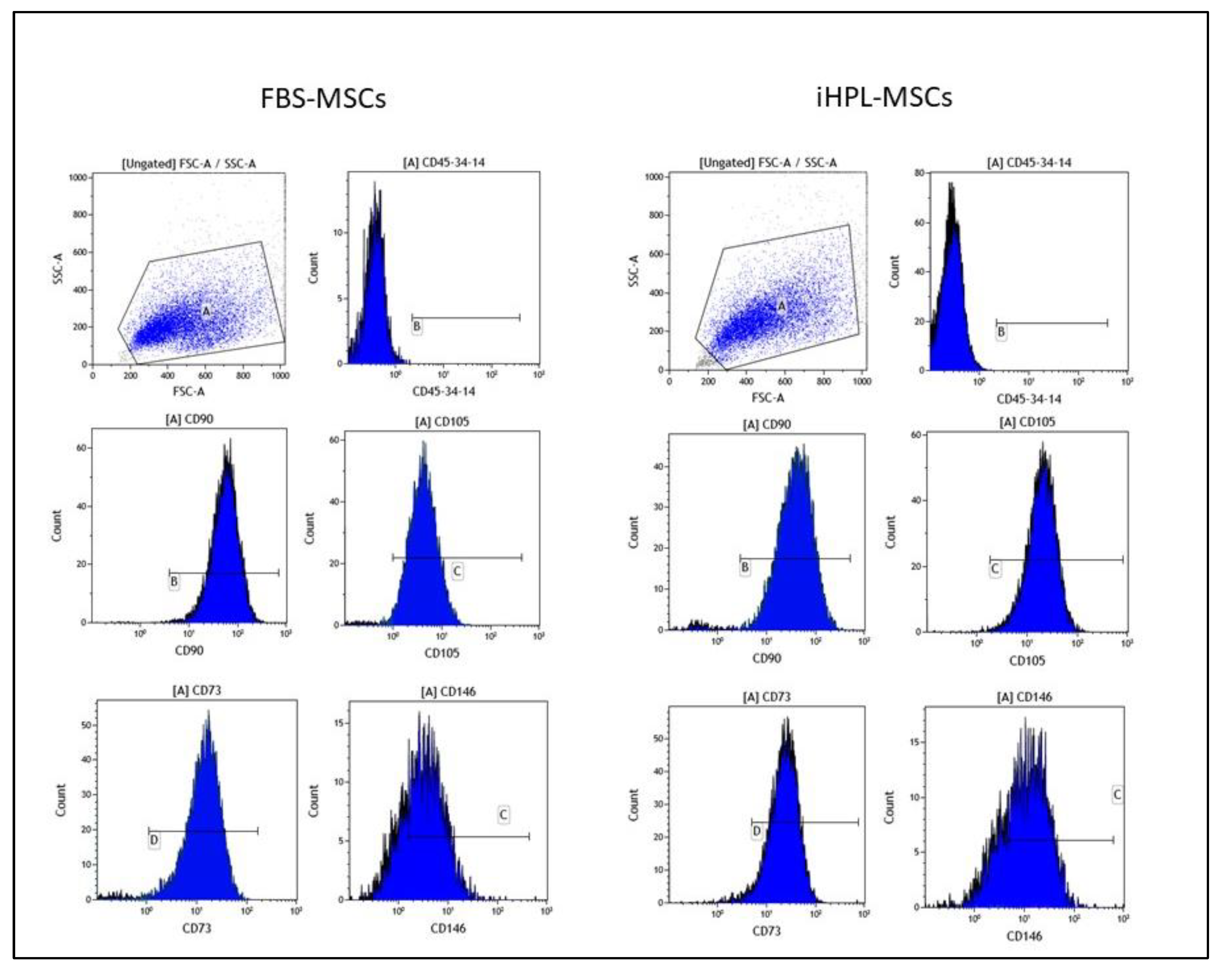
2.1. Human MSC Isolation, Expansion, Analysis, and Characterization
2.2. Preparation of Human Peripheral Blood Mononuclear Cells (PBMC)
2.3. Co-Culture MSCs/T Cells
2.4. Cytofluorimentric Analysis
2.5. Evaluation of Cytokine Release by ELISA
2.6. Statistical Analysis
3. Results
3.1. MSC Characteristics
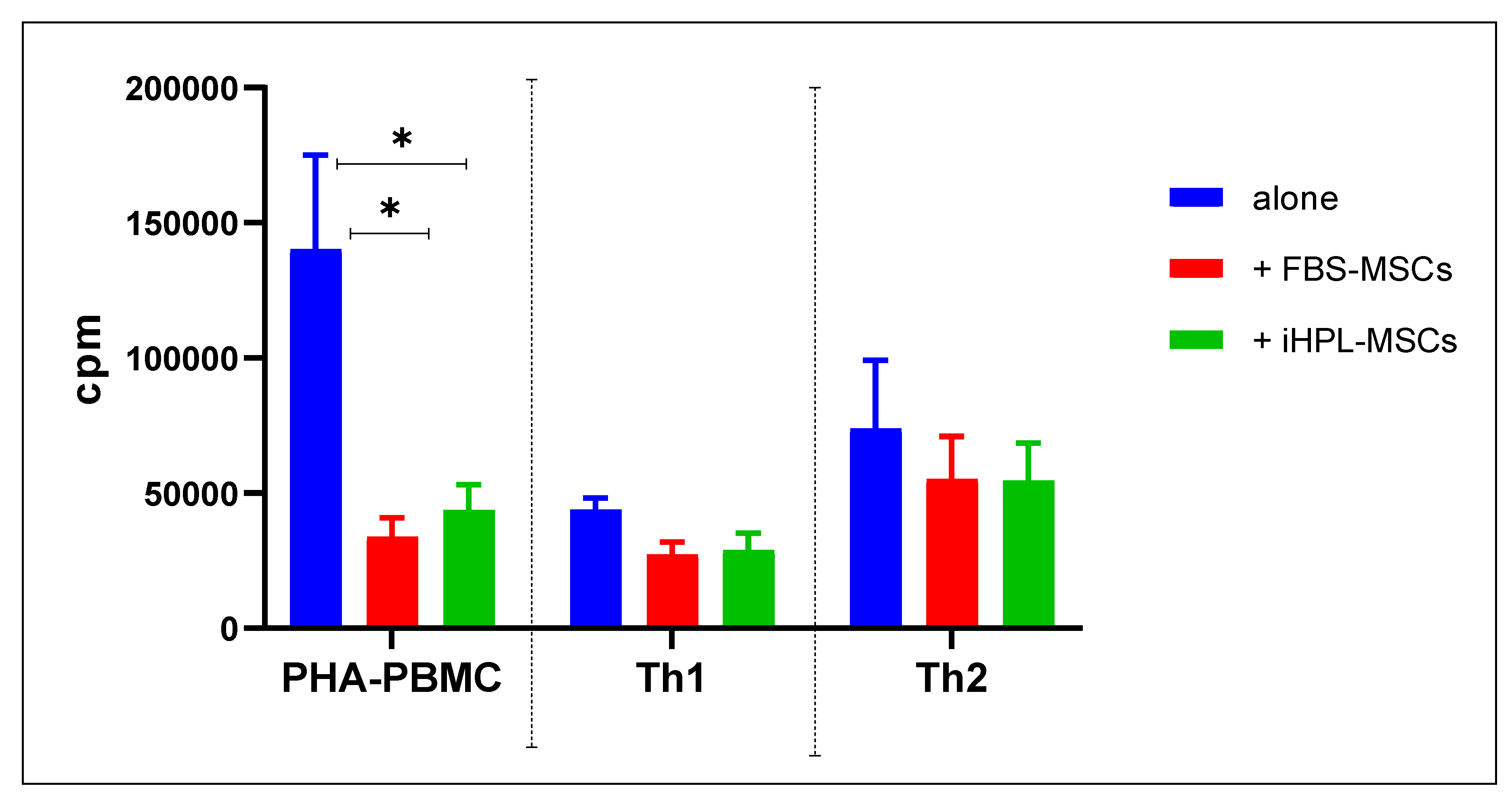
3.2. MSC/T Cell Interaction and Proliferative Assay
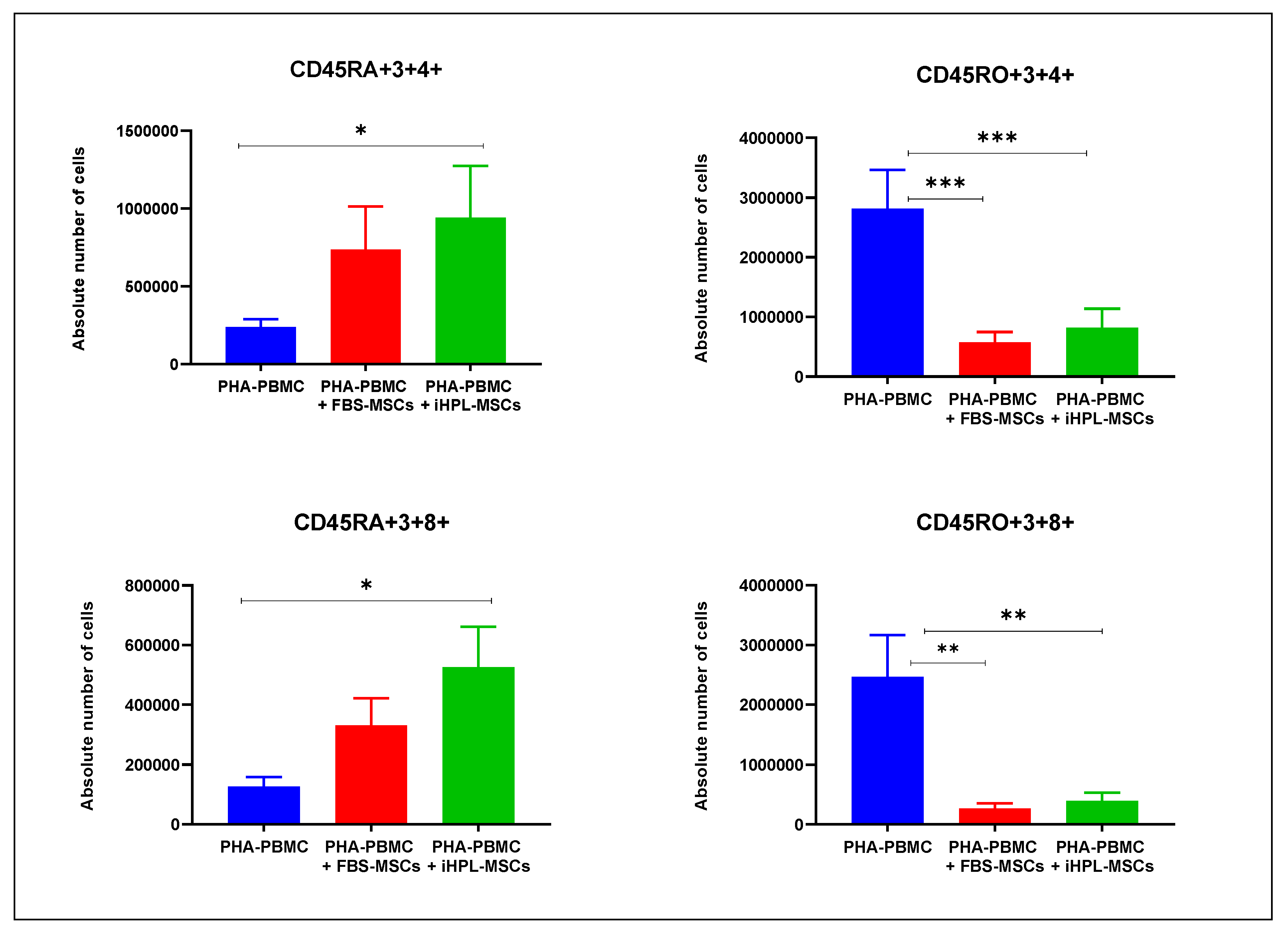
3.3. T Cell Subsets Determination
- Cytotoxic T naïve cells: CD45RA+/CD3+/CD8+
- Th naïve cells: CD45RA+/CD3+/CD4+
- Cytotoxic T memory cells CD45RO+/CD3+/CD8+
- Th memory cells: CD45RO+/CD3+/CD4+
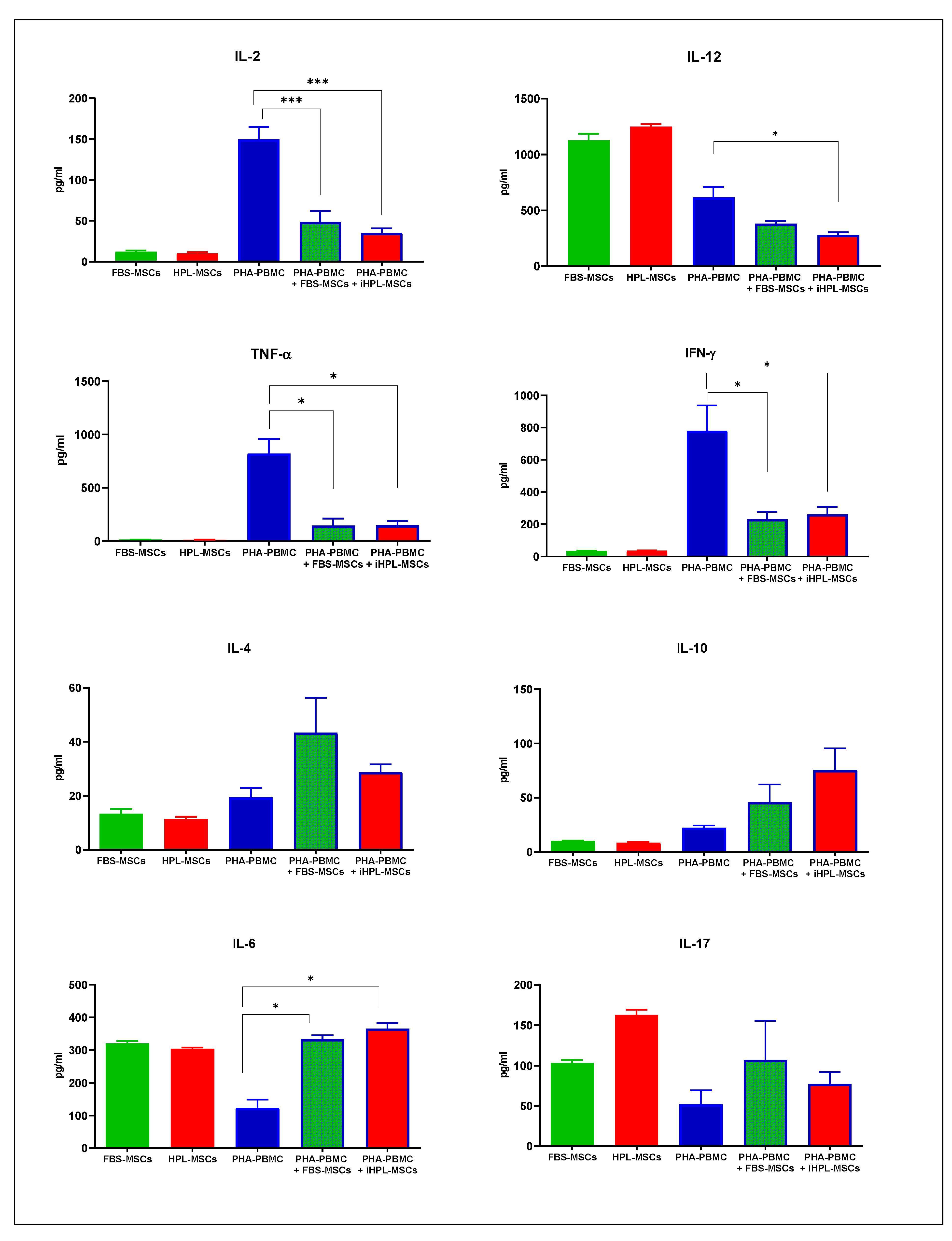
3.4. Th1/Th2/Th17 Cytokine Release
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Galipeau, J.; Sensébé, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, V.K.; Shih, H.-H.; Parveen, F.; Lenzen, D.; Ito, E.; Chan, T.-F.; Ke, L.Y. Identifying the Therapeutic Significance of Mesenchymal Stem Cells. Cells 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Daue, R. EudraLex—Volume 4—Good Manufacturing Practice (GMP) Guidelines. Public Health—European Commission, 2017. Available online: https://ec.europa.eu/health/documents/eudralex/vol-4_en (accessed on 25 March 2020).
- Anonymous. Guidelines Relevant for Advanced Therapy Medicinal Products. European Medicines Agency, 2018. Available online: https://www.ema.europa.eu/en/human-regulatory/research-development/advanced-therapies/guidelines-relevant-advanced-therapy-medicinal-products (accessed on 24 May 2020).
- Fekete, N.; Rojewski, M.T.; Fürst, D.; Kreja, L.; Ignatius, A.; Dausend, J.; Schrezenmeier, H. GMP-compliant isolation and large-scale expansion of bone marrow-derived MSC. PLoS ONE 2012, 7, e43255. [Google Scholar] [CrossRef] [PubMed]
- Capelli, C.; Domenghini, M.; Borleri, G.; Bellavita, P.; Poma, R.; Carobbio, A.; Micò, C.; Rambaldi, A.; Golay, J.; Introna, M. Human platelet lysate allows expansion and clinical grade production of mesenchymal stromal cells from small samples of bone marrow aspirates or marrow filter washouts. Bone Marrow Transpl. 2007, 40, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Schallmoser, K.; Rohde, E.; Bartmann, C.; Obenauf, A.C.; Reinisch, A.; Strunk, D. Platelet-derived growth factors for GMP-compliant propagation of mesenchymal stromal cells. Bio-Med. Mater. Eng. 2009, 19, 271–276. [Google Scholar] [CrossRef]
- Castiglia, S.; Mareschi, K.; Labanca, L.; Lucania, G.; Leone, M.; Sanavio, F.; Castello, L.; Rustichelli, D.; Signorino, E.; Gunetti, M. Inactivated human platelet lysate with psoralen: A new perspective for mesenchymal stromal cell production in Good Manufacturing Practice conditions. Cytotherapy 2014, 16, 750–763. [Google Scholar] [CrossRef]
- Horowitz, B.; Bonomo, R.; Prince, A.M.; Chin, S.N.; Brotman, B.; Shulman, R.W. Solvent/detergent-treated plasma: A virus-inactivated substitute for fresh frozen plasma. Blood 1992, 79, 826–831. [Google Scholar] [CrossRef] [Green Version]
- Prowse, C.V. Component pathogen inactivation: A critical review. Vox Sang 2013, 104, 183–199. [Google Scholar] [CrossRef]
- Tice, R.R.; Gatehouse, D.; Kirkland, D.; Speit, G. The pathogen reduction treatment of platelets with S-59 HCl (Amotosalen) plus ultraviolet A light: Genotoxicity profile and hazard assessment. Mutat. Res. 2007, 630, 50–68. [Google Scholar] [CrossRef]
- Blajchman, M.A. Protecting the blood supply from emerging pathogens: The role of pathogen inactivation. Transfus Clin. Biol. 2009, 16, 70–74. [Google Scholar] [CrossRef]
- Chen, M.-S.; Wang, T.-J.; Lin, H.-C.; Burnouf, T. Four types of human platelet lysate, including one virally inactivated by solvent-detergent, can be used to propagate Wharton jelly mesenchymal stromal cells. New Biotechnol. 2019, 49, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Viau, S.; Eap, S.; Chabrand, L.; Lagrange, A.; Delorme, B. Viral inactivation of human platelet lysate by gamma irradiation preserves its optimal efficiency in the expansion of human bone marrow mesenchymal stromal cells. Transfusion 2019, 59, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Li, C.; Wang, D. A novel ultraviolet illumination used in riboflavin photochemical method to inactivate drug-resistant bacteria in blood components. J. Photochem. Photobiol. B Biol. 2020, 204, 111782. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazik, H.; Spaggiari, G.M.; Chiossone, L.; Moretta, L. Mesenchymal stem cells expanded in human platelet lysate display a decreased inhibitory capacity on T- and NK-cell proliferation and function. Eur. J. Immunol. 2011, 41, 3281–3290. [Google Scholar] [CrossRef]
- Duffy, M.M.; Ritter, T.; Ceredig, R.; Griffin, M.D. Mesenchymal stem cell effects on T-cell effector pathways. Stem Cell Res. Ther. 2011, 2, 34. [Google Scholar] [CrossRef] [Green Version]
- Mareschi, K.; Castiglia, S.; Sanavio, F.; Rustichelli, D.; Muraro, M.; Defedele, D.; Bergallo, M.; Fagioli, F. Immunoregulatory effects on T lymphocytes by human mesenchymal stromal cells isolated from bone marrow, amniotic fluid, and placenta. Exp. Hematol. 2016, 44, 138–150.e1. [Google Scholar] [CrossRef] [Green Version]
- Consentius, C.; Reinke, P.; Volk, H.-D. Immunogenicity of allogeneic mesenchymal stromal cells: What has been seen in vitro and in vivo? Regen Med. 2015, 10, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, M.J.; Bae, E.-H.; Ryu, J.S.; Kaur, G.; Kim, H.J.; Kim, J.Y.; Barreda, H.; Jung, S.Y.; Choi, J.M.; et al. Comprehensive Molecular Profiles of Functionally Effective MSC-Derived Extracellular Vesicles in Immunomodulation. Mol. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz1, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Rustichelli, D.; Castiglia, S.; Gunetti, M.; Mareschi, K.; Signorino, E.; Muraro, M.; Castello, L.; Sanavio, F.; Leone, M.; Ferrero, I.; et al. Validation of analytical methods in compliance with good manufacturing practice: A practical approach. J. Transl. Med. 2013, 11, 197. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.-L.; Chiu, P.C.N.; Lam, K.K.W.; Siu, S.-O.; Chu, I.K.; Koistinen, R.; Koistinen, H.; Seppälä, M.; Lee, K.; Yeung, W.S.B. Differential actions of glycodelin-A on Th-1 and Th-2 cells: A paracrine mechanism that could produce the Th-2 dominant environment during pregnancy. Hum. Reprod. 2011, 26, 517–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Home-ClinicalTrials.gov n.d. Available online: https://clinicaltrials.gov/ (accessed on 24 March 2020).
- Bakopoulou, A.; Apatzidou, D.; Aggelidou, E.; Gousopoulou, E.; Leyhausen, G.; Volk, J.; Kritis, A.; Koidis, P.; Geurtsen, W. Isolation and prolonged expansion of oral mesenchymal stem cells under clinical-grade, GMP-compliant conditions differentially affects “stemness” properties. Stem Cell Res. Ther. 2017, 8, 247. [Google Scholar] [CrossRef] [PubMed]
- Czapla, J.; Matuszczak, S.; Kulik, K.; Wiśniewska, E.; Pilny, E.; Jarosz-Biej, M.; Smolarczyk, R.; Sirek, T.; Zembala, M.O.; Zembala, M.; et al. The effect of culture media on large-scale expansion and characteristic of adipose tissue-derived mesenchymal stromal cells. Stem Cell Res. Ther. 2019, 10, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mareschi, K.; Rustichelli, D.; Calabrese, R.; Gunetti, M.; Sanavio, F.; Castiglia, S.; Risso, A.; Ferrero, I.; Tarella, C.; Fagioli, F. Multipotent mesenchymal stromal stem cell expansion by plating whole bone marrow at a low cellular density: A more advantageous method for clinical use. Stem Cells Int. 2012, 2012, 920581. [Google Scholar] [CrossRef] [PubMed]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; Bron, D.; Toungouz, M.; Lagneaux, L. Mesenchymal stromal cells and immunomodulation: A gathering of regulatory immune cells. Cytotherapy 2016, 18, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Flemming, A.; Schallmoser, K.; Strunk, D.; Stolk, M.; Volk, H.-D.; Seifert, M. Immunomodulative efficacy of bone marrow-derived mesenchymal stem cells cultured in human platelet lysate. J. Clin. Immunol. 2011, 31, 1143–1156. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Avanzini, M.A.; Perotti, C.; Cometa, A.M.; Moretta, A.; Lenta, E.; del Fante, C.; Novara, F.; de Silvestri, A.; Amendola, G.; et al. Optimization of in vitro expansion of human multipotent mesenchymal stromal cells for cell-therapy approaches: Further insights in the search for a fetal calf serum substitute. J. Cell Physiol. 2007, 211, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Jonsdottir-Buch, S.M.; Sigurgrimsdottir, H.; Lieder, R.; Sigurjonsson, O.E. Expired and Pathogen-Inactivated Platelet Concentrates Support Differentiation and Immunomodulation of Mesenchymal Stromal Cells in Culture. Cell Transplant. 2015, 24, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, S.; Pittenger, M.F. Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 2005, 105, 1815–1822. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.É.S.; Sousa, M.R.R.; Alencar-Silva, T.; Carvalho, J.L.; Saldanha-Araujo, F. Mesenchymal stem cells immunomodulation: The road to IFN-γ licensing and the path ahead. Cytokine Growth Factor Rev. 2019, 47, 32–42. [Google Scholar] [CrossRef]
- Zhou, X.; Jin, N.; Wang, F.; Chen, B. Mesenchymal stem cells: A promising way in therapies of graft-versus-host disease. Cancer Cell Int. 2020, 20, 114. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yu, Q.; Hu, Y.; Shi, Y. Current Research and Use of Mesenchymal Stem Cells in the Therapy of Autoimmune Diseases. Curr. Stem Cell Res. Ther. 2019, 14, 579–582. [Google Scholar] [CrossRef]
- Zhukareva, V.; Obrocka, M.; Houle, J.D.; Fischer, I.; Neuhuber, B. Secretion profile of human bone marrow stromal cells: Donor variability and response to inflammatory stimuli. Cytokine 2010, 50, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Kyurkchiev, D.; Bochev, I.; Ivanova-Todorova, E.; Mourdjeva, M.; Oreshkova, T.; Belemezova, K.; Kyurkchiev, S. Secretion of immunoregulatory cytokines by mesenchymal stem cells. World J. Stem Cells 2014, 6, 552–570. [Google Scholar] [CrossRef]
- Laranjeira, P.; Pedrosa, M.; Pedreiro, S.; Gomes, J.; Martinho, A.; Antunes, B.; Ribeiro, T.; Santos, F.; Trindade, H.; Paiva, A. Effect of human bone marrow mesenchymal stromal cells on cytokine production by peripheral blood naive, memory, and effector T cells. Stem Cell Res. Ther. 2015, 6, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida-Porada, G.; Atala, A.J.; Porada, C.D. Therapeutic Mesenchymal Stromal Cells for Immunotherapy and for Gene and Drug Delivery. Mol. Ther. Methods Clin. Dev. 2020, 16, 204–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Fu, X.; Dai, G.; Wang, X.; Zhang, Z.; Cheng, H.; Zheng, P.; An, Y. Comparative analysis of curative effect of bone marrow mesenchymal stem cell and bone marrow mononuclear cell transplantation for spastic cerebral palsy. J. Transl. Med. 2017, 15. [Google Scholar] [CrossRef] [Green Version]
- Lange, C.; Cakiroglu, F.; Spiess, A.-N.; Cappallo-Obermann, H.; Dierlamm, J.; Zander, A.R. Accelerated and safe expansion of human mesenchymal stromal cells in animal serum-free medium for transplantation and regenerative medicine. J. Cell. Physiol. 2007, 213, 18–26. [Google Scholar] [CrossRef]
- Müller, I.; Kordowich, S.; Holzwarth, C.; Spano, C.; Isensee, G.; Staiber, A.; Viebahn, S.; Gieseke, F.; Langer, H.; Gawaz, M.P.; et al. Animal serum-free culture conditions for isolation and expansion of multipotent mesenchymal stromal cells from human BM. Cytotherapy 2006, 8, 437–444. [Google Scholar] [CrossRef]
- Grass, J.A.; Hei, D.J.; Metchette, K.; Cimino, G.D.; Wiesehahn, G.P.; Corash, L.; Lin, L. Inactivation of leukocytes in platelet concentrates by photochemical treatment with psoralen plus UVA. Blood 1998, 91, 2180–2188. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mareschi, K.; Castiglia, S.; Adamini, A.; Rustichelli, D.; Marini, E.; Banche Niclot, A.G.S.; Bergallo, M.; Labanca, L.; Ferrero, I.; Fagioli, F. Inactivated Platelet Lysate Supports the Proliferation and Immunomodulant Characteristics of Mesenchymal Stromal Cells in GMP Culture Conditions. Biomedicines 2020, 8, 220. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8070220
Mareschi K, Castiglia S, Adamini A, Rustichelli D, Marini E, Banche Niclot AGS, Bergallo M, Labanca L, Ferrero I, Fagioli F. Inactivated Platelet Lysate Supports the Proliferation and Immunomodulant Characteristics of Mesenchymal Stromal Cells in GMP Culture Conditions. Biomedicines. 2020; 8(7):220. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8070220
Chicago/Turabian StyleMareschi, Katia, Sara Castiglia, Aloe Adamini, Deborah Rustichelli, Elena Marini, Alessia Giovanna Santa Banche Niclot, Massimiliano Bergallo, Luciana Labanca, Ivana Ferrero, and Franca Fagioli. 2020. "Inactivated Platelet Lysate Supports the Proliferation and Immunomodulant Characteristics of Mesenchymal Stromal Cells in GMP Culture Conditions" Biomedicines 8, no. 7: 220. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8070220