Genistein Combined Polysaccharide (GCP) Can Inhibit Intracrine Androgen Synthesis in Prostate Cancer Cells

 , , and
, , and
Abstract
:1. Introduction
2. Experimental Procedures
2.1. Chemicals
2.2. GCP Extraction, Fractionation, and UPLC-MS Analysis
2.3. Cell Culture
2.4. Proliferation Assays
2.5. Immunoblot Analysis
2.6. Apoptosis Analysis
2.7. Reverse Transcription Quantitative Real-Time PCR
2.8. Sample Preparation and UPLC-MS/MS Analysis of Steroid Metabolites
2.9. NMR Analysis of Fraction 40
2.10. LC-MS Analysis of Fraction 40
2.11. Statistical Analyses
3. Results
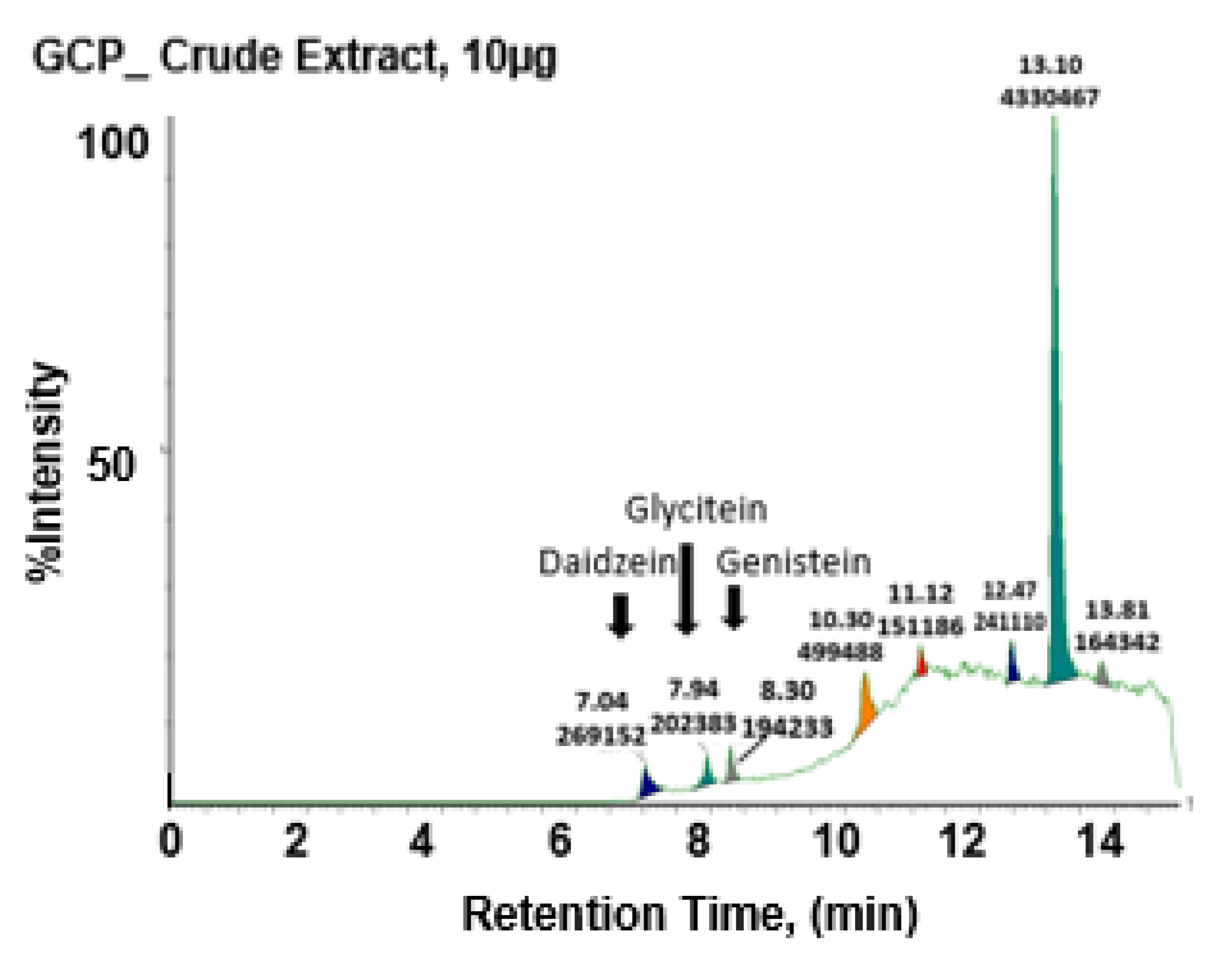
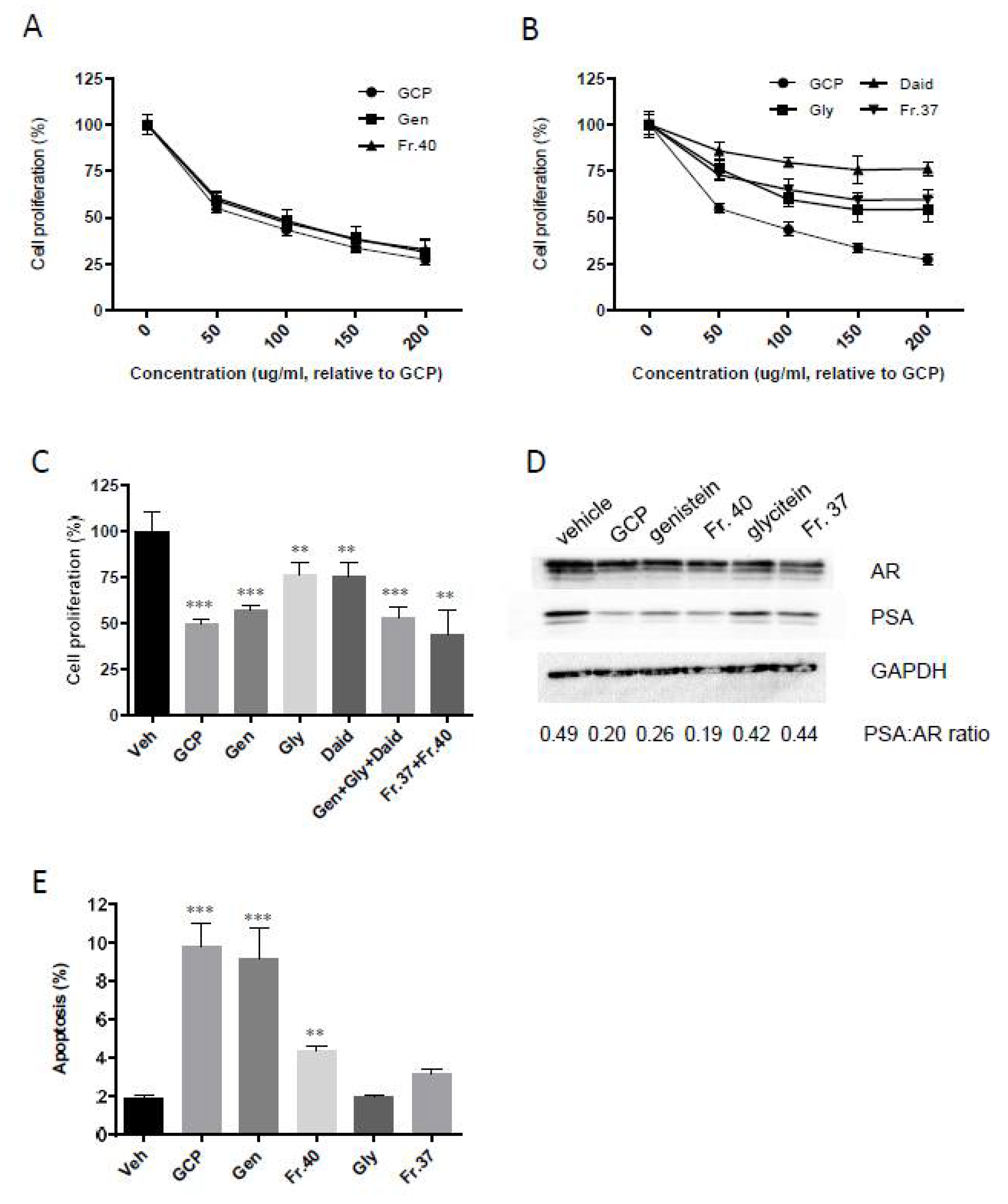
3.1. GCP Comprises High Levels of Genistein, Daidzein, and Glycitein as well as Other Unknown Compounds
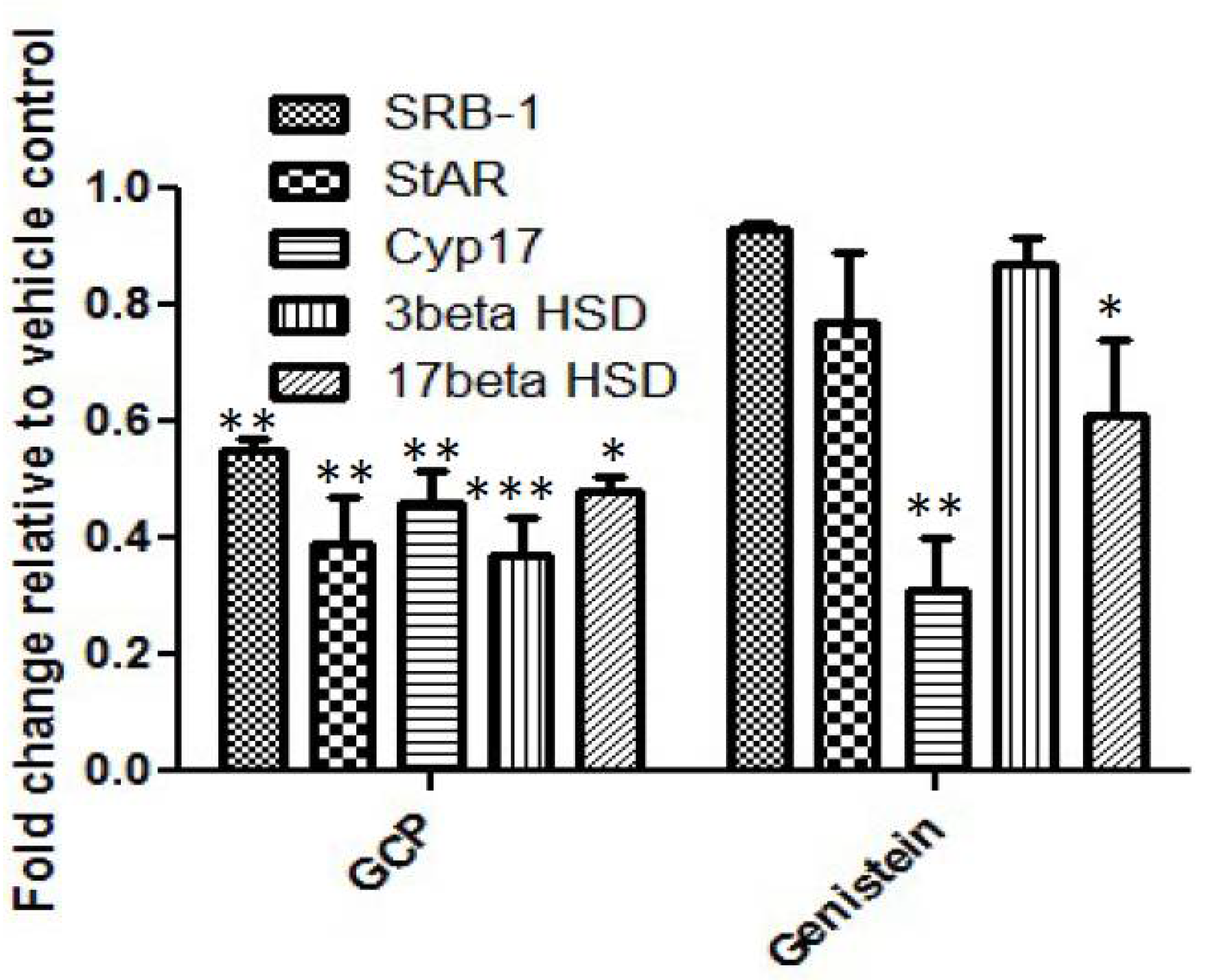
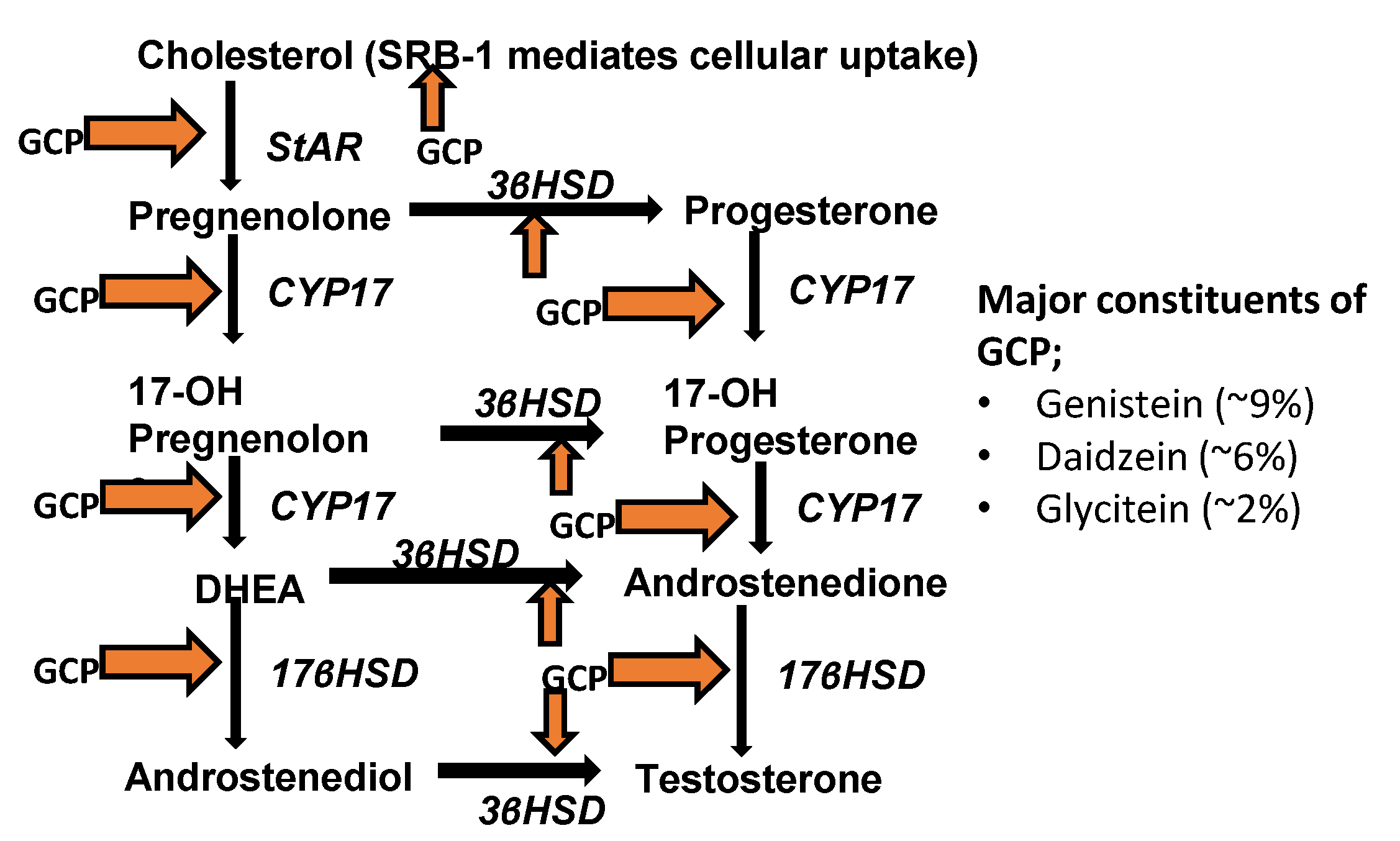
3.2. GCP Inhibits Intracrine Androgen Synthesis
3.3. GCP Fractionation and Cell Proliferation Assays show that GCP has Three Main Active Fractions
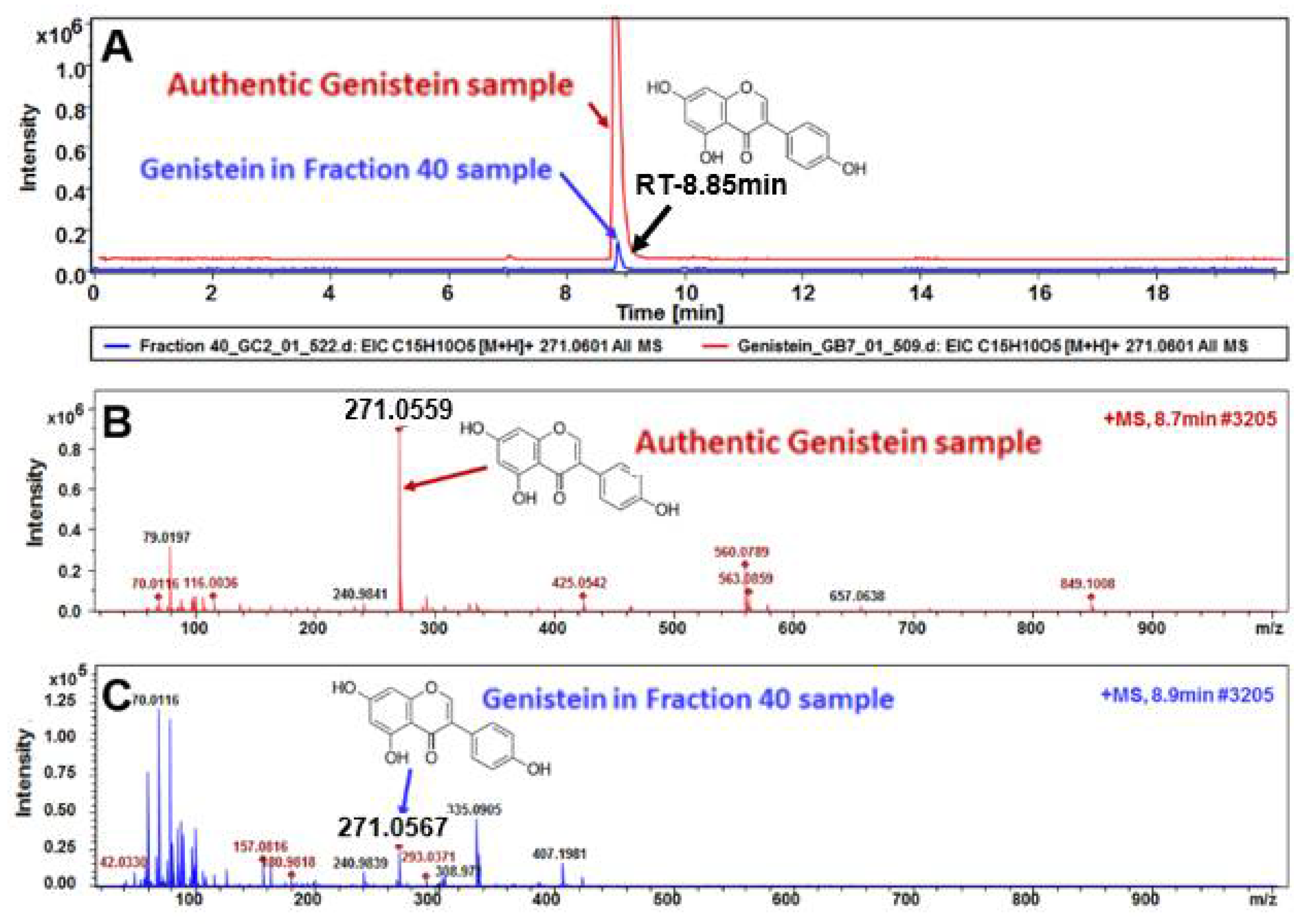
3.4. HPLC, LC-MS, and NMR Analysis Show the Primary Constituent of Fraction 40 is Genistein, and that the Primary Constituents of Fraction 37 is Daidzein
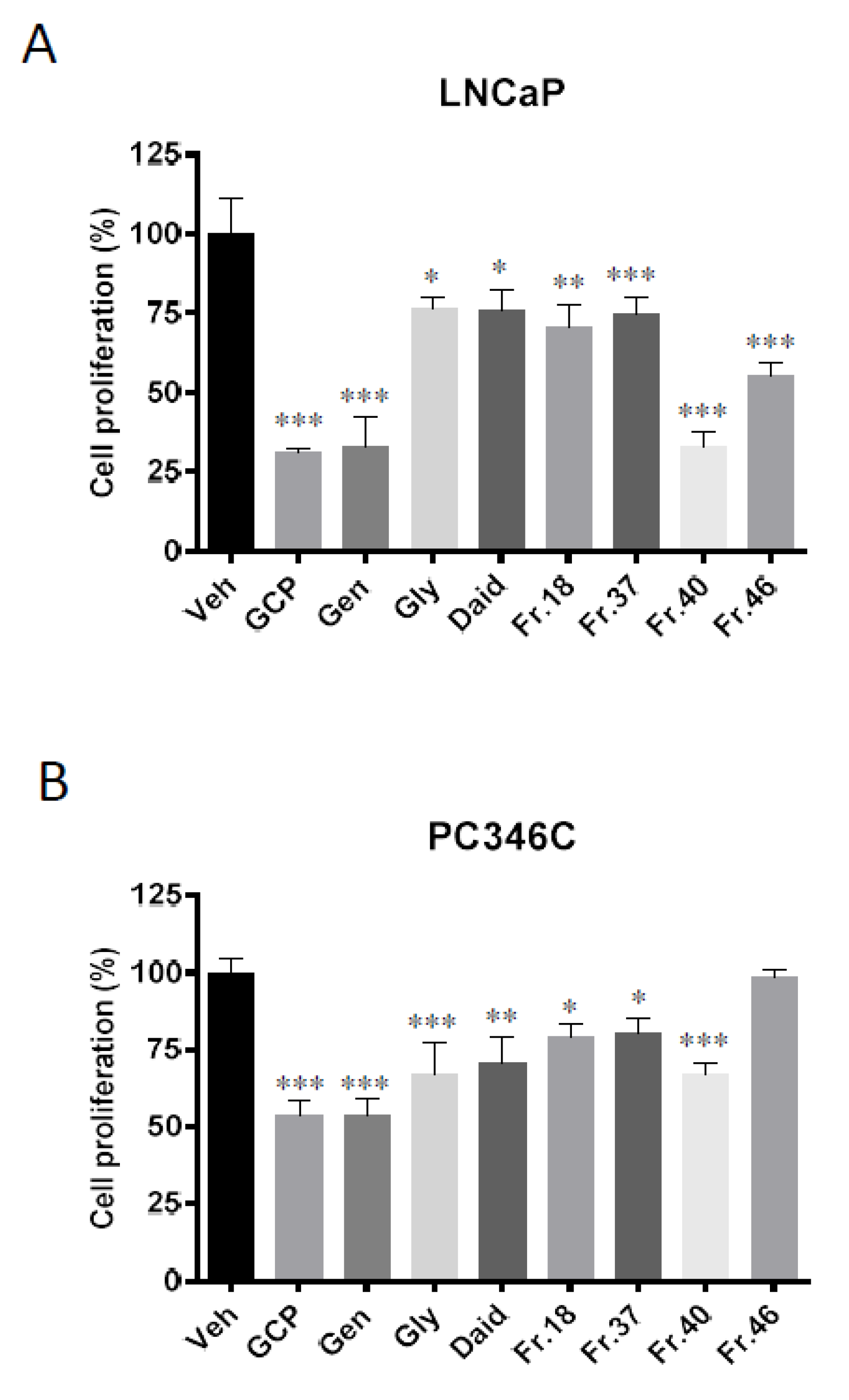
3.5. Genistein is Responsible for the Majority of GCP’s Anti-Proliferative, Anti-Androgen Receptor, and Pro-Apoptotic Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADT | Androgen deprivation therapy |
| AR | Androgen receptor |
| CAM | Complimentary alternative medicine |
| CaP | Prostate cancer |
| CRPC | Castration-resistant prostate cancer |
| CS | Cholesterol sulfate |
| GCP | Genistein combined polysaccharide |
| HPLC | High-Performance liquid chromatography |
| HSQC NMR | Heteronuclear multiple bond correlation nuclear magnetic resonance |
| NMR | Nuclear magnetic resonance |
| qPCR | Quantitative real-time polymerase chain reaction |
| TDCA | Taurodeoxycholicacid |
| UPLC-MS | Ultra-Performance liquid chromatography-mass spectrometry |
References
- Roehl, K.A.; Han, M.; Ramos, C.G.; Antenor, J.A.V.; Catalona, W.J. Cancer Progression and Survival Rates Following Anatomical Radical Retropubic Prostatectomy in 3478 Consecutive Patients: Long-Term Results. J. Urol. 2004, 172, 910–914. [Google Scholar] [CrossRef] [Green Version]
- Freedland, S.J.; Humphreys, E.B.; Mangold, L.A.; Eisenberger, M.; Dorey, F.J.; Walsh, P.C.; Partin, A.W. Risk of Prostate Cancer–Specific Mortality Following Biochemical Recurrence After Radical Prostatectomy. JAMA 2005, 294, 433–439. [Google Scholar] [CrossRef] [Green Version]
- Kupelian, P.; Mahadevan, A.; Reddy, C.A.; Reuther, A.M.; Klein, E. Use of different definitions of biochemical failure after external beam radiotherapy changes conclusions about relative treatment efficacy for localized prostate cancer. Urology 2006, 68, 593–598. [Google Scholar] [CrossRef]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [Green Version]
- Hotte, S.J.; Saad, F. Current management of castrate-resistant prostate cancer. Curr. Oncol. 2010, 17, S72–S79. [Google Scholar] [CrossRef]
- Kirby, M.; Hirst, C.; Crawford, E.D. Characterising the castration-resistant prostate cancer population: A systematic review. Int. J. Clin. Pr. 2011, 65, 1180–1192. [Google Scholar] [CrossRef]
- Kassem, L.; Shohdy, K.S.; Abdel-Rahman, O. Abiraterone acetate/androgen deprivation therapy combination versus docetaxel/androgen deprivation therapy combination in advanced hormone-sensitive prostate cancer: A network meta-analysis on safety and efficacy. Curr. Med. Res. Opin. 2018, 34, 903–910. [Google Scholar] [CrossRef]
- Harshman, L.C.; Taplin, M.-E. Abiraterone Acetate: Targeting Persistent Androgen Dependence in Castration-Resistant Prostate Cancer. Adv. Ther. 2013, 30, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Bemis, D.L.; Capodice, J.L.; Desai, M.; Buttyan, R.; Katz, A.E. A Concentrated Aglycone Isoflavone Preparation (GCP) That Demonstrates Potent Anti-Prostate Cancer Activity In vitro and In vivo. Clin. Cancer Res. 2004, 10, 5282–5292. [Google Scholar] [CrossRef] [Green Version]
- Burich, R.A.; Holland, W.S.; Vinall, R.L.; Tepper, C.; de Vere White, R.W.; Mack, P.C. Genistein combined polysaccharide enhances activity of docetaxel, bicalutamide and Src kinase inhibition in androgen-dependent and independent prostate cancer cell lines. BJU Int. 2008, 102. [Google Scholar] [CrossRef]
- de Vere White, R.W.; Tsodikov, A.; Stapp, E.C.; Soares, S.E.; Fujii, H.; Hackman, R.M. Effects of a High Dose, Aglycone-Rich Soy Extract on Prostate-Specific Antigen and Serum Isoflavone Concentrations in Men With Localized Prostate Cancer. Nutr. Cancer 2010, 62, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Ghafar, M.A.; Golliday, E.; Bingham, J.; Mansukhani, M.M.; Anastasiadis, A.G.; Katz, A.E. Regression of Prostate Cancer Following Administration of Genistein Combined Polysaccharide (GCP™), a Nutritional Supplement: A Case Report. J. Altern. Complement. Med. 2002, 8, 493–497. [Google Scholar] [CrossRef]
- Mooso, B.A.; Vinall, R.L.; Tepper, C.G.; Savoy, R.M.; Cheung, J.P.; Singh, S.; Siddiqui, S.; Wang, Y.; Bedolla, R.G.; Martinez, A.; et al. Enhancing the effectiveness of androgen deprivation in prostate cancer by inducing Filamin A nuclear localization. Endocr. Relat. Cancer 2012, 19, 759–777. [Google Scholar] [CrossRef] [Green Version]
- Tepper, C.G.; Vinall, R.L.; Wee, C.B.; Xue, L.; Shi, X.-B.; Burich, R.; Mack, P.C.; White, R.W.D. GCP-mediated growth inhibition and apoptosis of prostate cancer cells via androgen receptor-dependent and -independent mechanisms. Prostate 2007, 67, 521–535. [Google Scholar] [CrossRef]
- Vinall, R.L.; Hwa, K.; Ghosh, P.; Pan, C.-X.; Lara, P.N., Jr.; de Vere White, R.W. Combination Treatment of Prostate Cancer Cell Lines with Bioactive Soy Isoflavones and Perifosine Causes Increased Growth Arrest and/or Apoptosis. Clin. Cancer Res. 2007, 13, 6204–6216. [Google Scholar] [CrossRef] [Green Version]
- Pavese, J.M.; Krishna, S.N.; Bergan, R.C. Genistein inhibits human prostate cancer cell detachment, invasion, and metastasis. Am. J. Clin. Nutr. 2014, 100, 431S–436S. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Yang, W.; Bosland, M.C. Soy isoflavones and prostate cancer: A review of molecular mechanisms. J. Steroid Biochem. Mol. Boil. 2014, 140, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Goetzl, M.A.; Vanveldhuizen, P.J.; Thrasher, J.B. Effects of soy phytoestrogens on the prostate. Prostate Cancer Prostatic Dis. 2007, 10, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Yuan, L.; Sun, B.; Fujii, H.; Yoshida, M.; Wakame, K.; Kosuna, K.-I. Isoflavone Aglycon Produced by Culture of Soybean Extracts with Basidiomycetes and Its Anti-angiogenic Activity. Biosci. Biotechnol. Biochem. 2002, 66, 2626–2631. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.H.E.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E.-L. Androgen receptor: Structure, role in prostate cancer and drug discovery. Acta Pharmacol. Sin. 2015, 36, 3–23. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Gong, S.; Roy-Burman, P.; Lee, P.; Culig, Z. Current mouse and cell models in prostate cancer research. Endocr. Relat. Cancer 2013, 20, R155–R170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veldscholte, I.; Ris-Stalpers, C.; Kuiper, G.; Jenster, G.; Berrevoets, C.; Claassen, E.; van Rooij, H.C.J.; Trapman, J.; Brinkmann, A.O.; Mulder, E. A mutation in the ligand binding domain of the androgen receptor of human LNCaP cells affects steroid binding characteristics and response to anti-androgens. Biochem. Biophys. Res. Commun. 1990, 173, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Marques, R.B.; Van Weerden, W.M.; Erkens-Schulze, S.; De Ridder, C.M.; Bangma, C.H.; Trapman, J.; Jenster, G. The Human PC346 Xenograft and Cell Line Panel: A Model System for Prostate Cancer Progression. Eur. Urol. 2006, 49, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Dillard, P.R.; Lin, M.-F.; Khan, S.A. Androgen-independent prostate cancer cells acquire the complete steroidogenic potential of synthesizing testosterone from cholesterol. Mol. Cell. Endocrinol. 2008, 295, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaikwad, N.W. Ultra performance liquid chromatography-tandem mass spectrometry method for profiling of steroid metabolome in human tissue. Anal. Chem. 2013, 85, 4951–4960. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, N.W. Methods for Comprehensive Profiling of Steroid Metabolome. WO2015161078 A1, 22 October 2015. [Google Scholar]
- Fiandalo, M.V.; Wu, W.; Mohler, J.L. The role of intracrine androgen metabolism, androgen receptor and apoptosis in the survival and recurrence of prostate cancer during androgen deprivation therapy. Curr. Drug Targets 2013, 14, 420–440. [Google Scholar] [CrossRef]
- Vickman, R.E.; Crist, S.A.; Kerian, K.; Eberlin, L.; Cooks, R.G.; Burcham, G.N.; Buhman, K.K.; Hu, C.-D.; Mesecar, A.D.; Cheng, L.; et al. Cholesterol Sulfonation Enzyme, SULT2B1b, Modulates AR and Cell Growth Properties in Prostate Cancer. Mol. Cancer Res. 2016, 14, 776–786. [Google Scholar] [CrossRef] [Green Version]
- Basak, S.; Pookot, D.; Noonan, E.J.; Dahiya, R. Genistein down-regulates androgen receptor by modulating HDAC6-Hsp90 chaperone function. Mol. Cancer Ther. 2008, 7, 3195–3202. [Google Scholar] [CrossRef] [Green Version]
- Ajdžanović, V.; Medigović, I.; Živanović, J.; Mojić, M.; Milošević, V. Membrane Steroid Receptor-Mediated Action of Soy Isoflavones: Tip of the Iceberg. J. Membr. Boil. 2015, 248, 1–6. [Google Scholar] [CrossRef]
- Alex, A.B.; Pal, S.K.; Agarwal, N. CYP17 inhibitors in prostate cancer: Latest evidence and clinical potential. Ther. Adv. Med. Oncol. 2016, 8, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Hinoda, Y.; Shahryari, V.; Deng, G.; Tanaka, Y.; Tabatabai, Z.L.; Dahiya, R. Genistein downregulates onco-miR-1260b and upregulates sFRP1 and Smad4 via demethylation and histone modification in prostate cancer cells. Br. J. Cancer 2014, 110, 1645–1654. [Google Scholar] [CrossRef] [PubMed]
- Chiyomaru, T.; Yamamura, S.; Fukuhara, S.; Yoshino, H.; Kinoshita, T.; Majid, S.; Saini, S.; Chang, I.; Tanaka, Y.; Enokida, H.; et al. Genistein Inhibits Prostate Cancer Cell Growth by Targeting miR-34a and Oncogenic HOTAIR. PLoS ONE 2013, 8, e70372. [Google Scholar] [CrossRef] [PubMed]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic activity, biological effect and bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Nielsen, I.L.F.; Williamson, G. Review of the Factors Affecting Bioavailability of Soy Isoflavones in Humans. Nutr. Cancer 2007, 57, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ozasa, K.; Nakao, M.; Watanabe, Y.; Hayashi, K.; Miki, T.; Mikami, K.; Mori, M.; Sakauchi, F.; Washio, M.; Ito, Y.; et al. Serum phytoestrogens and prostate cancer risk in a nested case-control study among Japanese men. Cancer Sci. 2004, 95, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.-Y.; Yuan, Y.; Shi, X.; Che, Y.-Y. Improved drug delivery and anti-tumor efficacy of combinatorial liposomal formulation of genistein and plumbagin by targeting Glut1 and Akt3 proteins in mice bearing prostate tumor. Colloids Surf. B Biointerfaces 2020, 190, 110966. [Google Scholar] [CrossRef]
- Chanphai, P.; Tajmir-Riahi, H.-A. Encapsulation of micronutrients resveratrol, genistein, and curcumin by folic acid-PAMAM nanoparticles. Mol. Cell. Biochem. 2018, 449, 157–166. [Google Scholar] [CrossRef]
- Mei, L.; Zhang, H.; Zeng, X.; Huang, L.; Wang, Z.; Liu, G.; Wu, Y.; Yang, C. Fabrication of genistein-loaded biodegradable TPGS-b-PCL nanoparticles for improved therapeutic effects in cervical cancer cells. Int. J. Nanomed. 2015, 10, 2461–2473. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.; Sahu, P.K.; Beg, S.; Manohar Babu, S. Nanoparticles for Cancer Targeting: Current and Future Directions. Curr. Drug Deliv. 2016, 13, 1290–1302. [Google Scholar] [CrossRef]
- Soifer, H.S.; Souleimanian, N.; Wu, S.; Voskresenskiy, A.M.; Collak, F.K.; Cinar, B.; Stein, C.A. Direct Regulation of Androgen Receptor Activity by Potent CYP17 Inhibitors in Prostate Cancer Cells. J. Boil. Chem. 2012, 287, 3777–3787. [Google Scholar] [CrossRef] [Green Version]
- Vasaitis, T.; Belosay, A.; Schayowitz, A.; Khandelwal, A.; Chopra, P.; Gediya, L.K.; Guo, Z.; Fang, H.B.; Njar, V.C.; Brodie, A.M. Androgen receptor inactivation contributes to antitumor efficacy of 17{alpha}-hydroxylase/17,20-lyase inhibitor 3beta-hydroxy-17-(1H-benzimidazole-1-yl)androsta-5,16-diene in prostate cancer. Mol. Cancer Ther. 2008, 7, 2348–2357. [Google Scholar] [CrossRef] [Green Version]
- Shafiee, G.; Saidijam, M.; Tayebinia, H.; Khodadadi, I. Beneficial effects of genistein in suppression of proliferation, inhibition of metastasis, and induction of apoptosis in PC3 prostate cancer cells. Arch. Physiol. Biochem. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Grossebrummel, H.; Peter, T.; Mandelkow, R.; Weiss, M.; Muzzio, D.; Zimmermann, U.; Walther, R.; Jensen, F.; Knabbe, C.; Zygmunt, M.; et al. Cytochrome P450 17A1 inhibitor abiraterone attenuates cellular growth of prostate cancer cells independently from androgen receptor signaling by modulation of oncogenic and apoptotic pathways. Int. J. Oncol. 2016, 48, 793–800. [Google Scholar] [CrossRef] [Green Version]
- Bilir, B.; Sharma, N.V.; Lee, J.; Hammarstrom, B.; Svindland, A.; Kucuk, O.; Moreno, C.S. Effects of genistein supplementation on genome-wide DNA methylation and gene expression in patients with localized prostate cancer. Int. J. Oncol. 2017, 51, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Eberlin, L.S.; Dill, A.L.; Costa, A.; Ifa, D.R.; Cheng, L.; Masterson, T.; Koch, M.; Ratliff, T.L.; Cooks, R.G. Cholesterol Sulfate Imaging in Human Prostate Cancer Tissue by Desorption Electrospray Ionization Mass Spectrometry. Anal. Chem. 2010, 82, 3430–3434. [Google Scholar] [CrossRef] [Green Version]
- Toledo, A.; Yamaguchi, J.; Wang, J.Y.; Bass, B.L.; Turner, D.J.; Strauch, E.D. Taurodeoxycholate stimulates intestinal cell proliferation and protects against apoptotic cell death through activation of NF-kappaB. Dig. Dis. Sci. 2004, 49, 1664–1671. [Google Scholar] [CrossRef]
- Yamaguchi, J.; Toledo, A.; Bass, B.L.; A Celeste, F.; Rao, J.N.; Wang, J.-Y.; Strauch, E.D. Taurodeoxycholate increases intestinal epithelial cell proliferation through c-myc expression. Surgery 2004, 135, 215–221. [Google Scholar] [CrossRef]
- Di Ciaula, A.; Wang, D.Q.-H.; Molina-Molina, E.; Baccetto, R.L.; Calamita, G.; Palmieri, V.O.; Portincasa, P. Bile Acids and Cancer: Direct and Environmental-Dependent Effects. Ann. Hepatol. 2017, 16, S87–S105. [Google Scholar] [CrossRef]
- Hu, Y.; Chau, T.; Liu, H.-X.; Liao, D.; Keane, R.; Nie, Y.; Yang, H.; Wan, Y.-J.Y. Bile acids regulate nuclear receptor (Nur77) expression and intracellular location to control proliferation and apoptosis. Mol. Cancer Res. 2015, 13, 281–292. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Vehicle (Pg/30 Million Cells (SD)) | GCP (Pg/30 Million Cells (SD)) | p-Value |
|---|---|---|---|
| Testosterone | 46 (4.1) | 14 (1.2) | p < 0.001 |
| Epitestosterone | 326 (7.2) | 0 (0) | p < 0.001 |
| Cholesterol | 440,339 (50.3) | 454,832 (90.9) | p < 0.001 |
| Cholesterol-3-SO4 | 431,845 (73.1) | 209,803 (29.9) | p < 0.001 |
| 7-Keto-Cholesterol | 126,973 (67.6) | 322,883 (87.9) | p < 0.001 |
| Taurodeoxycholic acid | 326 (15.8) | 23 (1.5) | p < 0.001 |
| Glycodeoxycholic acid | 102 (5.9) | 0 (0) | p < 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batra, N.; Sam, A.; Woldemariam, T.; Talbott, G.; de Vere White, R.W.; Ghosh, P.M.; Gaikwad, N.W.; Kotchoni, S.O.; Vinall, R.L. Genistein Combined Polysaccharide (GCP) Can Inhibit Intracrine Androgen Synthesis in Prostate Cancer Cells. Biomedicines 2020, 8, 282. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080282
Batra N, Sam A, Woldemariam T, Talbott G, de Vere White RW, Ghosh PM, Gaikwad NW, Kotchoni SO, Vinall RL. Genistein Combined Polysaccharide (GCP) Can Inhibit Intracrine Androgen Synthesis in Prostate Cancer Cells. Biomedicines. 2020; 8(8):282. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080282
Chicago/Turabian StyleBatra, Neelu, Anhao Sam, Tibebe Woldemariam, George Talbott, Ralph W. de Vere White, Paramita M. Ghosh, Nilesh W. Gaikwad, Simeon O. Kotchoni, and Ruth L. Vinall. 2020. "Genistein Combined Polysaccharide (GCP) Can Inhibit Intracrine Androgen Synthesis in Prostate Cancer Cells" Biomedicines 8, no. 8: 282. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8080282