
Sex-Dependent End-of-Life Mental and Vascular Scenarios for Compensatory Mechanisms in Mice with Normal and AD-Neurodegenerative Aging


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Behavioral
2.3.1. Neuropsychiatric-Like Behaviors
2.3.2. Corner Test (CT) and Open-Field Test (OF)
2.3.3. Dark–Light Box Test (DLB)
2.3.4. Marble Burying Test (MB)
2.3.5. T-maze Test (TM)
2.3.6. Morris Water Maze Test (MWM)
2.4. Body Weight, Mouse Clinical Frailty Index Assessment, and Survival
2.5. Systolic Blood Pressure
2.6. MRI-ASL—Relative Cerebral Blood Flow
2.7. Angiogenesis
2.8. Aortic Function
2.9. HPA Axis Endocrine Status
2.10. Statistics
3. Results
3.1. Survival
3.2. HPA Axis Endocrine Status
3.3. Behavioral Assessment for Physical, Emotional, and Cognitive Phenotypes
3.3.1. Physical Phenotype
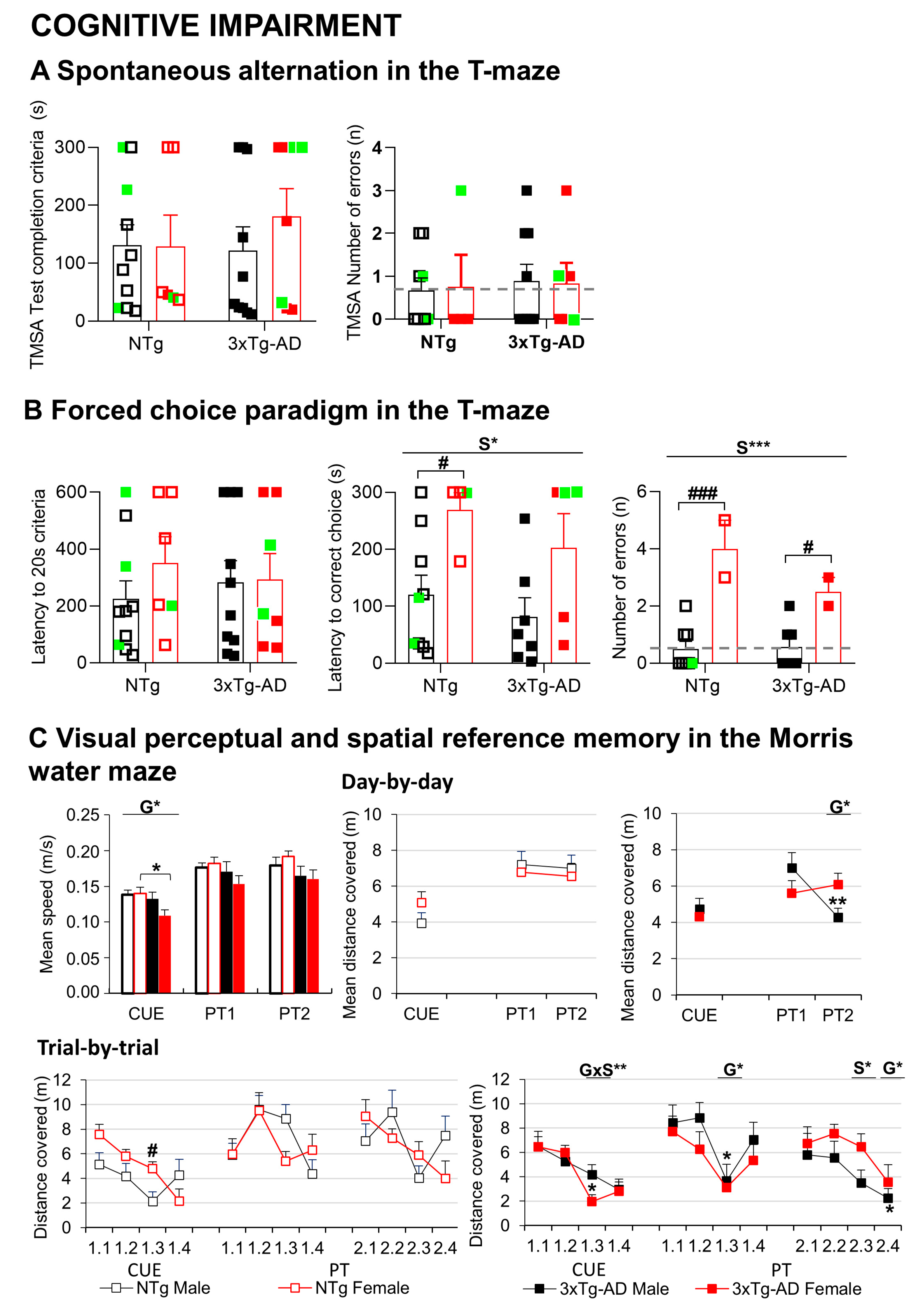
3.3.2. Neuropsychiatric symptoms (NPS)-like phenotype and cognitive impairment under different anxiogenic conditions
3.4. Systolic Blood Pressure
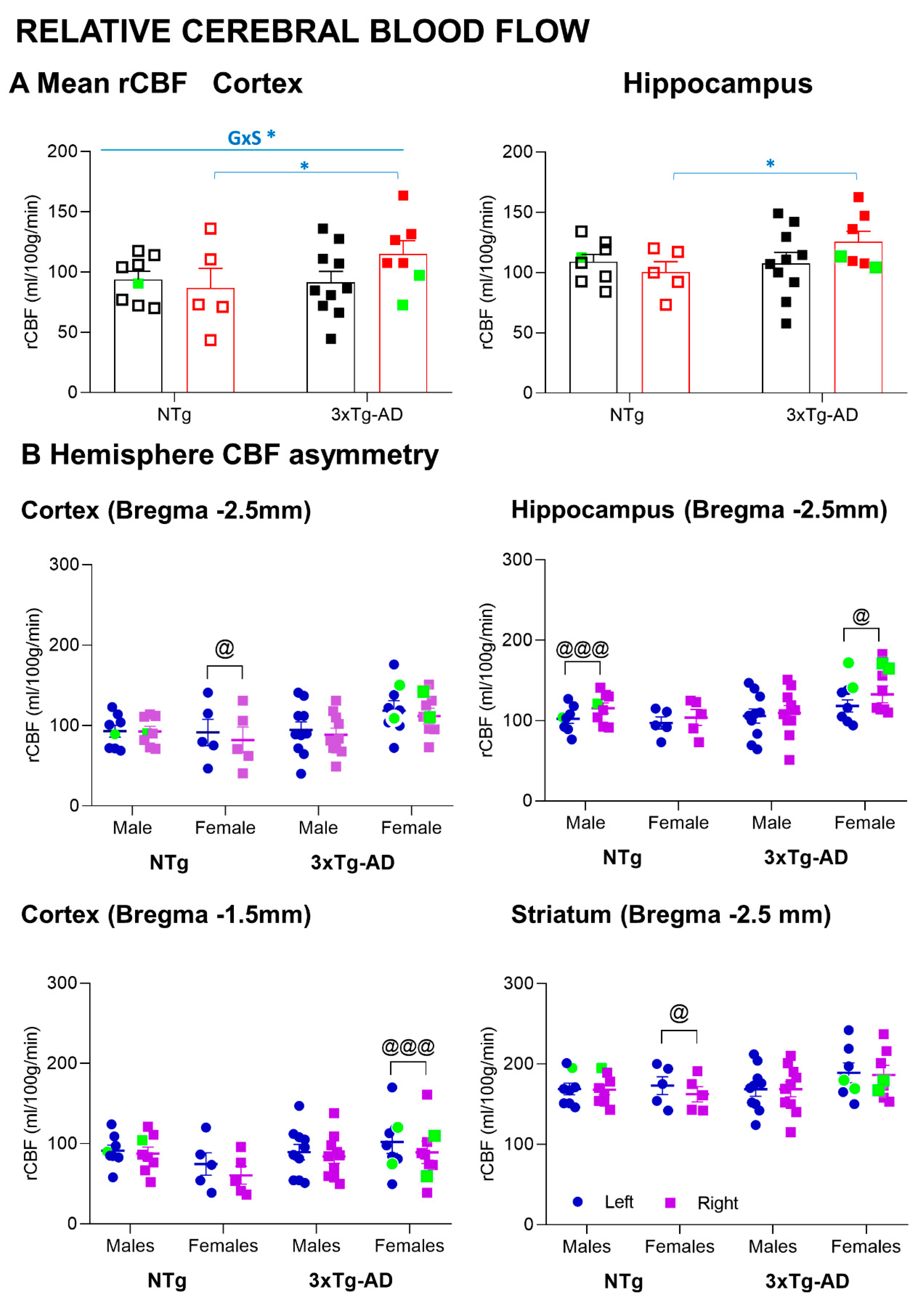
3.5. MRI Relative Cerebral Blood Flow
3.6. Angiogenesis
3.7. Arterial Properties
3.8. Mental Health and Cardiovascular Function Correlates
4. Discussion
4.1. Sex- and Genotype-Dependent Mortality/Morbidity Paradox
4.2. Down-Regulation of HPA Axis Endocrine Status in Female 3xTg-AD Mice
4.3. Different Behavioral Signatures for Physical, Emotional, and Cognitive Phenotypes
4.4. Increased MRI-ASL Regional Cerebral Blood Flow in 3xTg-AD Survivor Females
4.5. Sex- and Brain-Region-Dependent Asymmetry in the MRI-ASL Regional Cerebral Blood Flow
4.6. Improved Vascular Profile in 3xTg-AD Survivor Females
4.7. Behavioral Correlates Mental Health and Cardiovascular Measurements
4.8. Future Research Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 3xTg-AD | Triple transgenic mice |
| AD | Alzheimer’s disease |
| AI | Asymmetry index |
| ASL | Arterial spin labeling |
| AUC | Area under the curve |
| BLA | Basolateral amygdala |
| BPSD | Behavioral and psychological symptoms of dementia |
| CBF | Cerebral blood flow |
| CPu | Caudate putamen |
| CNR | Contrast-to-noise ratio |
| CT1 | Corner test—Day 1 |
| CT2 | Corner test—Day 2 |
| DALYs | Disability-adjusted life years |
| DLB | Dark–light box test |
| G | Genotype |
| HC | Hippocampus |
| HALE | Healthy life expectancy |
| HPA | Hypothalamic–pituitary–adrenal |
| MB | Marble test |
| MCA | Middle cerebral artery |
| MCFI | Mouse Clinical Frailty Index |
| MRA | Mesenteric resistance arteries |
| MRI | Magnetic resonance imaging |
| MWM | Morris water maze |
| NPS | Neuropsychiatric symptoms |
| NTg | Non transgenic mice |
| OF1 | Open-field test—Day 1 |
| OF1n | Open-field test—Day 1, minute n of the test |
| OF2 | Open-field test—Day 2 |
| OF2n | Open-field test—Day 2, minute n of the test |
| PT n n | Place task—Day, trial n of the test |
| RMA | Repeated measures ANOVA |
| ROI | Region of interest |
| S | Sex |
| SNR | Signal-to-noise ratio |
| T | Time |
| TM | T-maze |
| TMSA | T-maze spontaneous alternation |
References
- Kyu, H.H.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, Regional, and National Disability-Adjusted Life-Years (dalys) for 359 Diseases and Injuries and Healthy Life Expectancy (HALE) for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1859–1922. [Google Scholar] [CrossRef] [Green Version]
- Mattiuzzi, C.; Lippi, G. Worldwide Disease Epidemiology in the Older Persons. Eur. Geriatr. Med. 2020, 11, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Alberts, S.C.; Archie, E.A.; Gesquiere, L.R.; Altmann, J.; Vaupel, J.W.; Christensen, K. The Male-Female Health-Survival Paradox: A Comparative Perspective on Sex Differences in Aging and Mortality. In Sociality, Hierarchy, Health: Comparative Biodemography: A Collection of Papers.; Maxine Weinstein, M.A.L., Ed.; National Academies Press (US): Washington, DC, USA, 2014; pp. 339–363. ISBN 978-0-309-30661-4. [Google Scholar]
- Mitchell, S.L.; Miller, S.C.; Teno, J.M.; Kiely, D.K.; Davis, R.B.; Shaffer, M.L. Prediction of 6-Month Survival of Nursing Home Residents with Advanced Dementia Using ADEPT vs Hospice Eligibility Guidelines. JAMA 2010, 304, 1929–1935. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.; Song, X.; Dong, J.; Mitnitski, A.; Liu, J.; Guo, Z.; Rockwood, K. Mortality in Relation to Frailty in Patients Admitted to a Specialized Geriatric Intensive Care Unit. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 70, 1586–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dijk, P.T.; Dippel, D.W.; Habbema, J.D.F. Survival of Patients with Dementia. J. Am. Geriatr. Soc. 1991, 39, 603–610. [Google Scholar] [CrossRef]
- Komarova, N.L.; Thalhauser, C.J. High Degree of Heterogeneity in Alzheimer’s Disease Progression Patterns. PLoS Comput. Biol. 2011, 7, e1002251. [Google Scholar] [CrossRef] [Green Version]
- Torres-Lista, V.; De la Fuente, M.; Giménez-Llort, L. Survival Curves and Behavioral Profiles of Female 3xTg-AD Mice Surviving to 18-Months of Age as Compared to Mice with Normal Aging. J. Alzheimer’s Dis. Rep. 2017, 1, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Oddo, S.; Caccamo, A.; Shepherd, J.D.; Murphy, M.P.; Golde, T.E.; Kayed, R.; Metherate, R.; Mattson, M.P.; Akbari, Y.; LaFerla, F.M. Triple-Transgenic Model of Alzheimer’s Disease with Plaques and Tangles: Intracellular Abeta and Synaptic Dysfunction. Neuron 2003, 39, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Giménez-Llort, L.; Blázquez, G.; Cañete, T.; Johansson, B.; Oddo, S.; Tobeña, A.; LaFerla, F.M.; Fernández-Teruel, A. Modeling Behavioral and Neuronal Symptoms of Alzheimer’s Disease in Mice: A Role for Intraneuronal Amyloid. Neurosci. Biobehav. Rev. 2007, 31, 125–147. [Google Scholar] [CrossRef]
- Blázquez, G.; Cañete, T.; Tobeña, A.; Giménez-Llort, L.; Fernández-Teruel, A. Cognitive and Emotional Profiles of Aged Alzheimer’s Disease (3×TgAD) Mice: Effects of Environmental Enrichment and Sexual Dimorphism. Behav. Brain Res. 2014, 268, 185–201. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Arranz, L.; Maté, I.; De La Fuente, M. Gender-Specific Neuroimmunoendocrine Aging in a Triple-Transgenic 3×Tg-AD Mouse Model for Alzheimer’s Disease and Its Relation with Longevity. Neuroimmunomodulation 2008, 15, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mesa, Y.; Colie, S.; Corpas, R.; Cristòfol, R.; Comellas, F.; Nebreda, A.R.; Giménez-Llort, L.; Sanfeliu, C. Oxidative Stress Is a Central Target for Physical Exercise Neuroprotection Against Pathological Brain Aging. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2015, 71, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Rae, E.A.; Brown, R.E. The Problem of Genotype and Sex Differences in Life Expectancy in Transgenic AD Mice. Neurosci. Biobehav. Rev. 2015, 57, 238–251. [Google Scholar] [CrossRef] [PubMed]
- Hirata-Fukae, C.; Li, H.F.; Hoe, H.S.; Gray, A.J.; Minami, S.S.; Hamada, K.; Niikura, T.; Hua, F.; Tsukagoshi-Nagai, H.; Horikoshi-Sakuraba, Y.; et al. Females Exhibit More Extensive Amyloid, but Not Tau, Pathology in an Alzheimer Transgenic Model. Brain Res. 2008, 1216, 92–103. [Google Scholar] [CrossRef]
- Jefferson, A.L. Cardiac Output as a Potential Risk Factor for Abnormal Brain Aging. J. Alzheimer’s Dis. 2010, 20, 813–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troncone, L.; Luciani, M.; Coggins, M.; Wilker, E.H.; Ho, C.Y.; Codispoti, K.E.; Frosch, M.P.; Kayed, R.; del Monte, F. Aβ Amyloid Pathology Affects the Hearts of Patients with Alzheimer’s Disease: Mind the Heart. J. Am. Coll. Cardiol. 2016, 68, 2395–2407. [Google Scholar] [CrossRef]
- Sena, C.M.; Pereira, A.M.; Carvalho, C.; Fernandes, R.; Seiça, R.M.; Oliveira, C.R.; Moreira, P.I. Type 2 Diabetes Aggravates Alzheimer’s Disease-Associated Vascular Alterations of the Aorta in Mice. J. Alzheimer’s Dis. 2015, 45, 127–138. [Google Scholar] [CrossRef]
- Navarro-Dorado, J.; Villalba, N.; Prieto, D.; Brera, B.; Martín-Moreno, A.M.; Tejerina, T.; de Ceballos, M.L. Vascular Dysfunction in a Transgenic Model of Alzheimer’s Disease: Effects of CB1R and CB2R Cannabinoid Agonists. Front. Neurosci. 2016, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Merlini, M.; Shi, Y.; Keller, S.; Savarese, G.; Akhmedov, A.; Derungs, R.; Spescha, R.D.; Kulic, L.; Nitsch, R.M.; Lüscher, T.F.; et al. Reduced Nitric Oxide Bioavailability Mediates Cerebroarterial Dysfunction Independent of Cerebral Amyloid Angiopathy in a Mouse Model of Alzheimer’s Disease. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H232–H238. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Altayó, F.; Sánchez-Ventura, J.; Vila, E.; Giménez-Llort, L. Crosstalk between Peripheral Small Vessel Properties and Anxious-like Profiles: Sex, Genotype, and Interaction Effects in Mice with Normal Aging and 3×Tg-AD Mice at Advanced Stages of Disease. J. Alzheimer’s Dis. 2018, 62, 1531–1538. [Google Scholar] [CrossRef]
- Vagnucci, A.H.; Li, W.W. Alzheimer’s Disease and Angiogenesis. Lancet 2003, 361, 605–608. [Google Scholar] [CrossRef]
- Jefferies, W.A.; Price, K.A.; Biron, K.E.; Fenninger, F.; Pfeifer, C.G.; Dickstein, D.L. Adjusting the Compass: New Insights into the Role of Angiogenesis in Alzheimer’s Disease. Alzheimer’s Res. Ther. 2013, 5, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biron, K.E.; Dickstein, D.L.; Gopaul, R.; Jefferies, W.A. Amyloid Triggers Extensive Cerebral Angiogenesis Causing Blood Brain Barrier Permeability and Hypervascularity in Alzheimer’s Disease. PLoS ONE 2011, 6, e23789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving Bioscience Research Reporting: The ARRIVE Guidelines for Reporting Animal Research. PLoS Biol. 2010, 8, e1000412. [Google Scholar] [CrossRef] [PubMed]
- Belzung, C.; Le Pape, G. Comparison of Different Behavioral Test Situations Used in Psychopharmacology for Measurement of Anxiety. Physiol. Behav. 1994, 56, 623–628. [Google Scholar] [CrossRef]
- Hall, C.; Ballachey, E.L. A Study of the Rat’s Behavior in a Field. A Contribution to Method in Comparative Psychology. Univ. Calif. Publ. Psychol. 1932, 6, 1–12. [Google Scholar]
- Torres-Lista, V.; Parrado-Fernández, C.; Alvarez-Montón, I.; Frontiñán-Rubio, J.; Durán-Prado, M.; Peinado, J.R.; Johansson, B.; Alcaín, F.J.; Giménez-Llort, L. Neophobia, NQO1 and SIRT1 as Premorbid and Prodromal Indicators of AD in 3xTg-AD Mice. Behav. Brain Res. 2014, 271, 140–146. [Google Scholar] [CrossRef]
- Douglas, R.J. Cues for Spontaneous Alternation. J. Comp. Physiol. Psychol. 1966, 62, 171–183. [Google Scholar] [CrossRef]
- Gulinello, M.; Gertner, M.; Mendoza, G.; Schoenfeld, B.P.; Oddo, S.; LaFerla, F.; Choi, C.H.; McBride, S.M.J.; Faber, D.S. Validation of a 2-Day Water Maze Protocol in Mice. Behav. Brain Res. 2009, 196, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, J.C.; Hildebrand, B.A.; Sun, M.; Rockwood, M.R.; Rose, R.A.; Rockwood, K.; Howlett, S.E. A Clinical Frailty Index in Aging Mice: Comparisons with Frailty Index Data in Humans. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2014, 69, 621–632. [Google Scholar] [CrossRef]
- Orejudo, M.; García-Redondo, A.B.; Rodrigues-Diez, R.R.; Rodrigues-Díez, R.; Santos-Sanchez, L.; Tejera-Muñoz, A.; Egido, J.; Selgas, R.; Salaices, M.; Briones, A.M.; et al. Interleukin-17A Induces Vascular Remodeling of Small Arteries and Blood Pressure Elevation. Clin. Sci. (Lond) 2020, 134, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, 3rd ed.; Elsevier Science: San Diego, CA, USA, 2008; ISBN 9780123742445. [Google Scholar]
- Vicente, D.; Hernández, B.; Segura, V.; Pascual, D.; Fornaciari, G.; Monto, F.; Mirabet, V.; Montesinos, M.C.; D’Ocon, P. Methodological Approach to Use Fresh and Cryopreserved Vessels as Tools to Analyze Pharmacological Modulation of the Angiogenic Growth. J. Cardiovasc. Pharmacol. 2016, 68, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Vila, E.; Solé, M.; Masip, N.; Puertas-Umbert, L.; Amaro, S.; Dantas, A.P.; Unzeta, M.; D’Ocon, P.; Planas, A.M.; Chamorro, Á.; et al. Uric Acid Treatment after Stroke Modulates the Krüppel-like Factor 2-VEGF-A Axis to Protect Brain Endothelial Cell Functions: Impact of Hypertension. Biochem. Pharmacol. 2019, 164, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Altayó, F.; Siegert, A.-M.; Bonorino, F.; Meirelles, T.; Barberà, L.; Dantas, A.P.; Vila, E.; Egea, G. Differences in the Thoracic Aorta by Region and Sex in a Murine Model of Marfan Syndrome. Front. Physiol. 2017, 8, 933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giménez-Llort, L.; García, Y.; Buccieri, K.; Revilla, S.; Suñol, C.; Cristofol, R.; Sanfeliu, C. Gender-Specific Neuroimmunoendocrine Response to Treadmill Exercise in 3xTg-AD Mice. Int. J. Alzheimers. Dis. 2010, 2010, 128354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Lista, V.; López-Pousa, S.; Giménez-Llort, L. Impact of Chronic Risperidone Use on Behavior and Survival of 3xTg-AD Mice Model of Alzheimer’s Disease and Mice with Normal Aging. Front. Pharmacol. 2019, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Maas, A.H.E.M.; Appelman, Y.E.A. Gender Differences in Coronary Heart Disease. Neth. Heart J. 2010, 18, 598–603. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Ratia, M.; Pérez, B.; Camps, P.; Muñoz-Torrero, D.; Badia, A.; Clos, M.V. AVCRI104P3, a Novel Multitarget Compound with Cognition-Enhancing and Anxiolytic Activities: Studies in Cognitively Poor Middle-Aged Mice. Behav. Brain Res. 2015, 286, 97–103. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Ramírez-Boix, P.; de la Fuente, M. Mortality of Septic Old and Adult Male Mice Correlates with Individual Differences in Premorbid Behavioral Phenotype and Acute-Phase Sickness Behavior. Exp. Gerontol. 2019, 127, 110717. [Google Scholar] [CrossRef]
- Gordon, E.H.; Peel, N.M.; Samanta, M.; Theou, O.; Howlett, S.E.; Hubbard, R.E. Sex Differences in Frailty: A Systematic Review and Meta-Analysis. Exp. Gerontol. 2017, 89, 30–40. [Google Scholar] [CrossRef]
- Trebbastoni, A.; Canevelli, M.; D’Antonio, F.; Imbriano, L.; Podda, L.; Rendace, L.; Campanelli, A.; Celano, V.; Bruno, G.; de Lena, C. The Impact of Frailty on the Risk of Conversion from Mild Cognitive Impairment to Alzheimer’s Disease: Evidences from a 5-Year Observational Study. Front. Med. 2017, 4, 178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, A.E.; Shin, S.; Wong, A.A.; Fertan, E.; Faustova, N.S.; Howlett, S.E.; Brown, R.E. Sex Differences in Healthspan Predict Lifespan in the 3xTg-AD Mouse Model of Alzheimer’s Disease. Front. Aging Neurosci. 2018, 10, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, A.; Veldhuis, J.D.; Deuschle, M.; Standhardt, H.; Heuser, I. Twenty-Four Hour Cortisol Release Profiles in Patients with Alzheimer’s and Parkinson’s Disease Compared to Normal Controls: Ultradian Secretory Pulsatility and Diurnal Variation. Neurobiol. Aging 1997, 18, 285–289. [Google Scholar] [CrossRef]
- Woiciechowsky, C.; Schöning, B.; Lanksch, W.R.; Volk, H.D.; Döcke, W.D. Mechanisms of Brain-Mediated Systemic Anti-Inflammatory Syndrome Causing Immunodepression. J. Mol. Med. 1999, 77, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Drude, S.; Geißler, A.; Olfe, J.; Starke, A.; Domanska, G.; Schuett, C.; Kiank-Nussbaum, C. Side Effects of Control Treatment Can Conceal Experimental Data When Studying Stress Responses to Injection and Psychological Stress in Mice. Lab Anim. (NY) 2011, 40, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Baeta-Corral, R.; Johansson, B.; Giménez-Llort, L. Long-Term Treatment with Low-Dose Caffeine Worsens BPSD-Like Profile in 3xTg-AD Mice Model of Alzheimer’S Disease and Affects Mice with Normal Aging. Front. Pharmacol. 2018, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- García-Mesa, Y.; López-Ramos, J.C.; Giménez-Llort, L.; Revilla, S.; Guerra, R.; Gruart, A.; Laferla, F.M.; Cristòfol, R.; Delgado-García, J.M.; Sanfeliu, C. Physical Exercise Protects against Alzheimer’s Disease in 3xTg-AD Mice. J. Alzheimer’s Dis. 2011, 24, 421–454. [Google Scholar] [CrossRef] [Green Version]
- Muntsant, A.; Giménez-Llort, L. Impact of Social Isolation on the Behavioral, Functional Profiles, and Hippocampal Atrophy Asymmetry in Dementia in Times of Coronavirus Pandemic (COVID-19): A Translational Neuroscience Approach. Front. Psychiatry 2020, 11, 572583. [Google Scholar] [CrossRef]
- Cañete, T.; Blázquez, G.; Tobeña, A.; Giménez-Llort, L.; Fernández-Teruel, A. Cognitive and Emotional Alterations in Young Alzheimer’s Disease (3xTgAD) Mice: Effects of Neonatal Handling Stimulation and Sexual Dimorphism. Behav. Brain Res. 2015, 281, 156–171. [Google Scholar] [CrossRef]
- Torres-Lista, V.; López-Pousa, S.; Giménez-Llort, L. Marble-Burying Is Enhanced in 3xTg-AD Mice, Can Be Reversed by Risperidone and It Is Modulable by Handling. Behav. Process. 2015, 116, 69–74. [Google Scholar] [CrossRef]
- Guayerbas, N.; Puerto, M.; Ferrández, M.D.; De La Fuente, M. A Diet Supplemented with Thiolic Anti-Oxidants Improves Leucocyte Function in Two Strains of Prematurely Ageing Mice. Clin. Exp. Pharmacol. Physiol. 2002, 29, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- D’Hooge, R.; De Deyn, P.P. Applications of the Morris Water Maze in the Study of Learning and Memory. Brain Res. Rev. 2001, 36, 60–90. [Google Scholar] [CrossRef]
- España, J.; Giménez-Llort, L.; Valero, J.; Miñano, A.; Rábano, A.; Rodriguez-Alvarez, J.; LaFerla, F.M.; Saura, C.A. Intraneuronal β-Amyloid Accumulation in the Amygdala Enhances Fear and Anxiety in Alzheimer’s Disease Transgenic Mice. Biol. Psychiatry 2010, 67, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Llort, L.; Ratia, M.; Pérez, B.; Camps, P.; Muñoz-Torrero, D.; Badia, A.; Clos, M.V. Behavioural Effects of Novel Multitarget Anticholinesterasic Derivatives in Alzheimer’s Disease. Behav. Pharmacol. 2017, 28, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Sierra-Marcos, A. Regional Cerebral Blood Flow in Mild Cognitive Impairment and Alzheimer’s Disease Measured with Arterial Spin Labeling Magnetic Resonance Imaging. Int. J. Alzheimers Dis. 2017, 2017, 5479597. [Google Scholar] [CrossRef]
- Frontiñán-Rubio, J.; Sancho-Bielsa, F.J.; Peinado, J.R.; LaFerla, F.M.; Giménez-Llort, L.; Durán-Prado, M.; Alcain, F.J. Sex-Dependent Co-Occurrence of Hypoxia and β-Amyloid Plaques in Hippocampus and Entorhinal Cortex Is Reversed by Long-Term Treatment with Ubiquinol and Ascorbic Acid in the 3xTg-AD Mouse Model of Alzheimer’s Disease. Mol. Cell. Neurosci. 2018, 92, 67–81. [Google Scholar] [CrossRef]
- Guo, Y.; Li, X.; Zhang, M.; Chen, N.; Wu, S.; Lei, J.; Wang, Z.; Wang, R.; Wang, J.; Liu, H. Age- and Brain Region-associated Alterations of Cerebral Blood Flow in Early Alzheimer’s Disease Assessed in AβPP SWE /PS1 ΔE9 Transgenic Mice Using Arterial Spin Labeling. Mol. Med. Rep. 2019, 19, 3045–3052. [Google Scholar] [CrossRef] [Green Version]
- Fazlollahi, A.; Calamante, F.; Liang, X.; Bourgeat, P.; Raniga, P.; Dore, V.; Fripp, J.; Ames, D.; Masters, C.L.; Rowe, C.C.; et al. Increased Cerebral Blood Flow with Increased Amyloid Burden in the Preclinical Phase of Alzheimer’s Disease. J. Magn. Reson. Imaging 2020, 51, 505–513. [Google Scholar] [CrossRef]
- Bracko, O.; Njiru, B.N.; Swallow, M.; Ali, M.; Haft-Javaherian, M.; Schaffer, C.B. Increasing Cerebral Blood Flow Improves Cognition into Late Stages in Alzheimer’s Disease Mice. J. Cereb. Blood Flow Metab. 2020, 40, 1441–1452. [Google Scholar] [CrossRef]
- Ardekani, B.A.; Hadid, S.A.; Blessing, E.; Bachman, A.H. Sexual Dimorphism and Hemispheric Asymmetry of Hippocampal Volumetric Integrity in Normal Aging and Alzheimer Disease. Am. J. Neuroradiol. 2019, 40, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Wachinger, C.; Salat, D.H.; Weiner, M.; Reuter, M. Whole-Brain Analysis Reveals Increased Neuroanatomical Asymmetries in Dementia for Hippocampus and Amygdala. Brain 2016, 139, 3253–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, K.; Kurashima, R.; Gokan, H.; Inoue, N.; Ito, I.; Watanabe, S. Left-right Asymmetry Defect in the Hippocampal Circuitry Impairs Spatial Learning and Working Memory in IV Mice. PLoS ONE 2010, 5, e15468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimbo, A.; Kosaki, Y.; Ito, I.; Watanabe, S. Mice Lacking Hippocampal Left-Right Asymmetry Show Non-Spatial Learning Deficits. Behav. Brain Res. 2018, 336, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Tadic, M.; Cuspidi, C.; Hering, D. Hypertension and Cognitive Dysfunction in Elderly: Blood Pressure Management for This Global Burden. BMC Cardiovasc. Disord. 2016, 16, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoog, I.; Gustafson, D. Update on Hypertension and Alzheimer’s Disease. Neurol. Res. 2006, 28, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Gilsanz, P.; Mayeda, E.R.; Glymour, M.M.; Quesenberry, C.P.; Mungas, D.M.; DeCarli, C.; Dean, A.; Whitmer, R.A. Female Sex, Early-Onset Hypertension, and Risk of Dementia. Neurology 2017, 89, 1886–1893. [Google Scholar] [CrossRef]
- Barsha, G.; Denton, K.M.; Mirabito Colafella, K.M. Sex- and Age-Related Differences in Arterial Pressure and Albuminuria in Mice. Biol. Sex Differ. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wiesmann, M.; Zerbi, V.; Jansen, D.; Lütjohann, D.; Veltien, A.; Heerschap, A.; Kiliaan, A.J. Hypertension, Cerebrovascular Impairment, and Cognitive Decline in Aged AβPP/PS1 Mice. Theranostics 2017, 7, 1277–1289. [Google Scholar] [CrossRef]
- Dede, D.S.; Yavuz, B.; Yavuz, B.B.; Cankurtaran, M.; Halil, M.; Ulger, Z.; Cankurtaran, E.S.; Aytemir, K.; Kabakci, G.; Ariogul, S. Assessment of Endothelial Function in Alzheimer’s Disease: Is Alzheimer’s Disease a Vascular Disease? J. Am. Geriatr. Soc. 2007, 55, 1613–1617. [Google Scholar] [CrossRef]
- Dickstein, D.L.; Walsh, J.; Brautigam, H.; Stockton, S.D.; Gandy, S.; Hof, P.R. Role of Vascular Risk Factors and Vascular Dysfunction in Alzheimer’s Disease. Mt. Sinai J. Med. 2010, 77, 82–102. [Google Scholar] [CrossRef] [Green Version]
- Desai, B.S.; Schneider, J.A.; Li, J.L.; Carvey, P.M.; Hendey, B. Evidence of Angiogenic Vessels in Alzheimer’s Disease. J. Neural. Transm. 2009, 116, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ethell, D.W. An Amyloid-Notch Hypothesis for Alzheimer’s Disease. Neuroscientist 2010, 16, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.J.; Galvin, C.; Alkam, T.; Sidhu, H.; Ellison, J.; Luna, S.; Ethell, D.W. Alzheimer’s-Related Peptide Amyloid-β Plays a Conserved Role in Angiogenesis. PLoS ONE 2012, 7, e39598. [Google Scholar] [CrossRef] [PubMed]
- Koike, M.A.; Lin, A.J.; Pham, J.; Nguyen, E.; Yeh, J.J.; Rahimian, R.; Tromberg, B.J.; Choi, B.; Green, K.N.; LaFerla, F.M. APP Knockout Mice Experience Acute Mortality as the Result of Ischemia. PLoS ONE 2012, 7, e42665. [Google Scholar] [CrossRef] [PubMed]
- Gaelle, D.; Nadia, H.; Thomas, P.; Vincent, D.; Jean-Louis, G.; Catherine, B.; Nicole, M.; Daniel, B. Sustained Corticosterone Rise in the Prefrontal Cortex Is a Key Factor for Chronic Stress-Induced Working Memory Deficits in Mice. Neurobiol. Stress 2019, 10, 100161. [Google Scholar] [CrossRef]
- Thifault, S.; Lalonde, R.; Joyal, C.C.; Hamet, P. Neurobehavioral Evaluation of High Blood Pressure and Low Blood Pressure Mice. Psychobiology 1999, 27, 415–425. [Google Scholar] [CrossRef]
- Giménez-Llort, L.; Martínez, E.; Ferré, S. Different Effects of Dopamine Antagonists on Spontaneous and NMDA-Induced Motor Activity in Mice. Pharmacol. Biochem. Behav. 1997, 56, 549–553. [Google Scholar] [CrossRef]
- Colorado, R.A.; Shumake, J.; Conejo, N.M.; Gonzalez-Pardo, H.; Gonzalez-Lima, F. Effects of Maternal Separation, Early Handling, and Standard Facility Rearing on Orienting and Impulsive Behavior of Adolescent Rats. Behav. Process. 2006, 71, 51–58. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Domains | Tests | Effect | Between Groups Differences | Figure |
|---|---|---|---|---|
| Physical Condition | ||||
| Survival | Survival curve | Sex ** | 3xTg-AD females vs. males | Figure 1A |
| Frailty | Frailty index | Genotype *** | 3xTg-AD males vs. NTg males | Figure 1D |
| Weight | Body weight | Genotype * | 3xTg-AD females vs. NTg females | Figure 1B |
| HPA axis | Corticosterone | Genotype*; Sex** | Females vs. males | Figure 1C |
| Behavioral Phenotype | ||||
| Neuropsychiatric-Like Domain | ||||
| Neophobia | CT,OF | Sex * | Figure 2A,B | |
| Hyperactivity | OF | Genotype * | 3xTg-AD males vs. NTg males | Figure 2B |
| Disinhibition | DLB | Genotype * | Figure 2C | |
| Cognitive Domain | ||||
| Long-term memory | OF2 | Genotype * | Figure 2B | |
| Working memory | TM | Sex *** | Females vs. males | Figure 3B |
| Swimming speed | MWM | Genotype * | 3xTg-AD females vs. NTg females | Figure 3C |
| Paradoxical performance | MWM | Genotype * | 3xTg-AD males vs. NTg males | Figure 3C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muntsant, A.; Jiménez-Altayó, F.; Puertas-Umbert, L.; Jiménez-Xarrie, E.; Vila, E.; Giménez-Llort, L. Sex-Dependent End-of-Life Mental and Vascular Scenarios for Compensatory Mechanisms in Mice with Normal and AD-Neurodegenerative Aging. Biomedicines 2021, 9, 111. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020111
Muntsant A, Jiménez-Altayó F, Puertas-Umbert L, Jiménez-Xarrie E, Vila E, Giménez-Llort L. Sex-Dependent End-of-Life Mental and Vascular Scenarios for Compensatory Mechanisms in Mice with Normal and AD-Neurodegenerative Aging. Biomedicines. 2021; 9(2):111. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020111
Chicago/Turabian StyleMuntsant, Aida, Francesc Jiménez-Altayó, Lidia Puertas-Umbert, Elena Jiménez-Xarrie, Elisabet Vila, and Lydia Giménez-Llort. 2021. "Sex-Dependent End-of-Life Mental and Vascular Scenarios for Compensatory Mechanisms in Mice with Normal and AD-Neurodegenerative Aging" Biomedicines 9, no. 2: 111. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020111