
Evidence-Based View of Safety and Effectiveness of Prokineticin Receptors Antagonists during Pregnancy

 , , ,
, , ,
Abstract
:1. Introduction
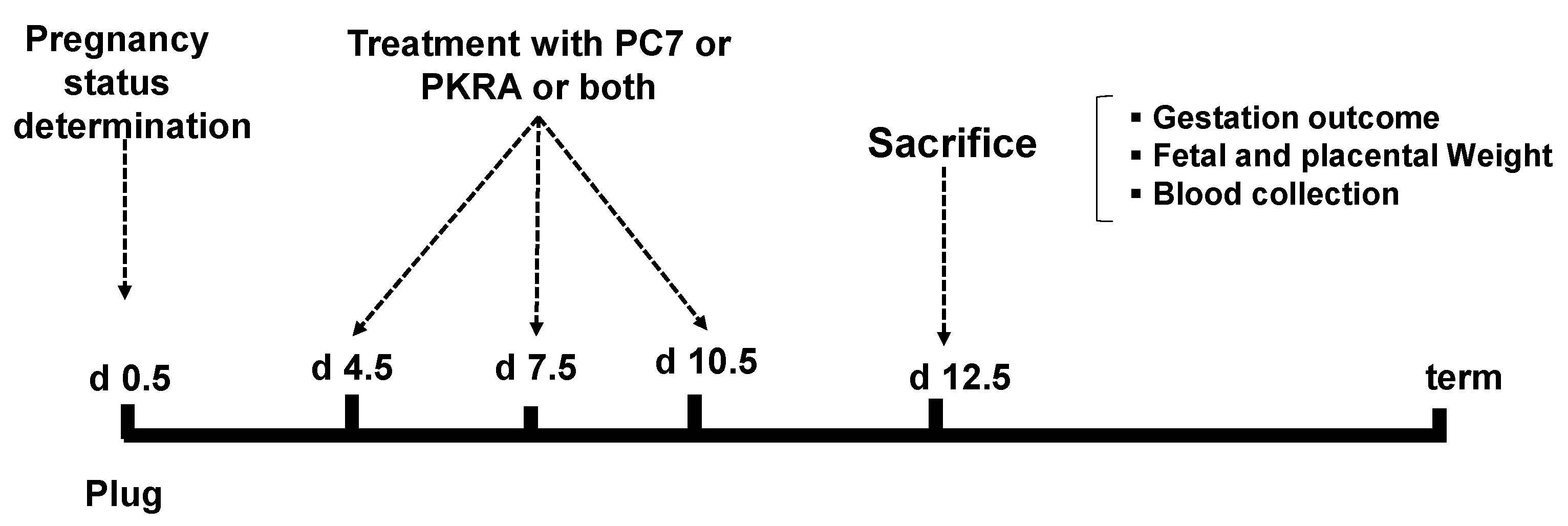
2. Materials & Methods
2.1. Animals
2.2. Real-Time RT-PCR Analysis of Placental Tissue
2.3. Placental Histology
2.4. Periodic Acid-Schiff Staining
2.5. Immunofluorescence
2.6. Western Blotting Analyses of Placental Tissues
2.7. RCHO-1 Cell Line Culture
2.8. Wound-Healing Assay
2.9. Matrigel Invasion Assay
2.10. Statistical Analysis
3. Results
3.1. Effects of Combined and Independent Treatments by PROKR1 (PC7) and PROKR2 (PKRA) Antagonists on the Litter Size, Feto-Placental Weights and Placental Efficiency
3.2. Effects of PC7 and PKRA Antagonists on Placental Growth and Vascularization
3.3. Effects of PC7 and PKRA Antagonists on Placental Structure
3.4. Effects of PC7 and PKRA Antagonists on Trophoblast Invasion
3.5. Effects of PC7 and PKRA Antagonists on Key Trophoblast Developmental Genes
3.6. Effects of PC7 and PKRA Antagonists on TGC Differentiation, Migration and Invasion in an In Vitro System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- LeCouter, J.; Kowalski, J.; Foster, J.; Hass, P.; Zhang, Z.; Dillard-Telm, L.; Frantz, G.; Rangell, L.; DeGuzman, L.; Keller, G.A.; et al. Identification of an angiogenic mitogen selective for endocrine gland endothelium. Nature 2001, 412, 877–884. [Google Scholar] [CrossRef]
- Lin, D.C.; Bullock, C.M.; Ehlert, F.J.; Chen, J.L.; Tian, H.; Zhou, Q.Y. Identification and molecular characterization of two closely related G protein-coupled receptors activated by prokineticins/endocrine gland vascular endothelial growth factor. J. Biol. Chem. 2002, 277, 19276–19280. [Google Scholar] [CrossRef] [Green Version]
- Robinson, C.J.; Stringer, S.E. The splice variants of vascular endothelial growth factor (VEGF) and their receptors. J. Cell Sci. 2001, 114, 853–865. [Google Scholar] [PubMed]
- Brouillet, S.; Hoffmann, P.; Feige, J.J.; Alfaidy, N. EG-VEGF: A key endocrine factor in placental development. Trends Endocrinol. Metab. 2012, 23, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Goryszewska, E.; Kaczynski, P.; Balboni, G.; Waclawik, A. Prokineticin 1-prokineticin receptor 1 signaling promotes angiogenesis in the porcine endometrium during pregnancydagger. Biol. Reprod. 2020, 103, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Traboulsi, W.; Brouillet, S.; Sergent, F.; Boufettal, H.; Samouh, N.; Aboussaouira, T.; Hoffmann, P.; Feige, J.J.; Benharouga, M.; Alfaidy, N. Prokineticins in central and peripheral control of human reproduction. Horm Mol. Biol. Clin. Investig. 2015, 24, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wu, J.; Wang, X.; Jia, H.; Chen, D.N.; Li, J.D. Prokineticins and their G protein-coupled receptors in health and disease. Prog. Mol. Biol. Transl. Sci. 2019, 161, 149–179. [Google Scholar] [CrossRef]
- Brouillet, S.; Hoffmann, P.; Chauvet, S.; Salomon, A.; Chamboredon, S.; Sergent, F.; Benharouga, M.; Feige, J.J.; Alfaidy, N. Revisiting the role of hCG: New regulation of the angiogenic factor EG-VEGF and its receptors. Cell Mol. Life Sci. 2012, 69, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N.; LeCouter, J.; Lin, R.; Peale, F. EG-VEGF and Bv8: A novel family of tissue-restricted angiogenic factors. Biochim. Biophys. Acta 2004, 1654, 69–78. [Google Scholar] [CrossRef]
- Zuena, A.R.; Casolini, P.; Lattanzi, R.; Maftei, D. Chemokines in Alzheimer’s Disease: New Insights Into Prokineticins, Chemokine-Like Proteins. Front. Pharmacol. 2019, 10, 622. [Google Scholar] [CrossRef] [Green Version]
- Alfaidy, N.; Hoffmann, P.; Gillois, P.; Gueniffey, A.; Lebayle, C.; Garcin, H.; Thomas-Cadi, C.; Bessonnat, J.; Coutton, C.; Villaret, L.; et al. PROK1 Level in the Follicular Microenvironment: A New Noninvasive Predictive Biomarker of Embryo Implantation. J. Clin. Endocrinol. Metab. 2016, 101, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, P.; Feige, J.J.; Alfaidy, N. Expression and oxygen regulation of endocrine gland-derived vascular endothelial growth factor/prokineticin-1 and its receptors in human placenta during early pregnancy. Endocrinology 2006, 147, 1675–1684. [Google Scholar] [CrossRef] [Green Version]
- Negri, L.; Maftei, D. Targeting the Prokineticin System to Control Chronic Pain and Inflammation. Curr. Med. Chem. 2018, 25, 3883–3894. [Google Scholar] [CrossRef]
- Yuill, E.A.; Hoyda, T.D.; Ferri, C.C.; Zhou, Q.Y.; Ferguson, A.V. Prokineticin 2 depolarizes paraventricular nucleus magnocellular and parvocellular neurons. Eur. J. Neurosci. 2007, 25, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.Y.; Lee, A.G.; Culbertson, C.; Sun, G.; Talati, R.K.; Manley, N.C.; Li, X.; Zhao, H.; Lyons, D.M.; Zhou, Q.Y.; et al. Prokineticin 2 is an endangering mediator of cerebral ischemic injury. Proc. Natl. Acad. Sci. USA 2012, 109, 5475–5480. [Google Scholar] [CrossRef] [Green Version]
- Maftei, D.; Marconi, V.; Florenzano, F.; Giancotti, L.A.; Castelli, M.; Moretti, S.; Borsani, E.; Rodella, L.F.; Balboni, G.; Luongo, L.; et al. Controlling the activation of the Bv8/prokineticin system reduces neuroinflammation and abolishes thermal and tactile hyperalgesia in neuropathic animals. Br. J. Pharmacol. 2014, 171, 4850–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severini, C.; Lattanzi, R.; Maftei, D.; Marconi, V.; Ciotti, M.T.; Petrocchi Passeri, P.; Florenzano, F.; Del Duca, E.; Caioli, S.; Zona, C.; et al. Bv8/prokineticin 2 is involved in Abeta-induced neurotoxicity. Sci. Rep. 2015, 5, 15301. [Google Scholar] [CrossRef] [Green Version]
- Alfaidy, N. Prokineticin1 and pregnancy. Ann. Endocrinol. 2016, 77, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Benlahfid, M.; Traboulsi, W.; Sergent, F.; Benharouga, M.; Elhattabi, K.; Erguibi, D.; Karkouri, M.; Elattar, H.; Fadil, A.; Fahmi, Y.; et al. Endocrine gland-derived vascular endothelial growth factor (EG-VEGF) and its receptor PROKR2 are associated to human colorectal cancer progression and peritoneal carcinomatosis. Cancer Biomark. 2018, 21, 345–354. [Google Scholar] [CrossRef]
- Brouillet, S.; Hoffmann, P.; Benharouga, M.; Salomon, A.; Schaal, J.P.; Feige, J.J.; Alfaidy, N. Molecular characterization of EG-VEGF-mediated angiogenesis: Differential effects on microvascular and macrovascular endothelial cells. Mol. Biol. Cell 2010, 21, 2832–2843. [Google Scholar] [CrossRef] [Green Version]
- Chauvet, S.; Traboulsi, W.; Thevenon, L.; Kouadri, A.; Feige, J.J.; Camara, B.; Alfaidy, N.; Benharouga, M. EG-VEGF, BV8, and their receptor expression in human bronchi and their modification in cystic fibrosis: Impact of CFTR mutation (delF508). Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L314–L322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, P.; Feige, J.J.; Alfaidy, N. Placental expression of EG-VEGF and its receptors PKR1 (prokineticin receptor-1) and PKR2 throughout mouse gestation. Placenta 2007, 28, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Yun, E.J.; Gerber, H.P.; Ferrara, N.; Whitsett, J.A.; Vu, T.H. Epithelial-vascular cross talk mediated by VEGF-A and HGF signaling directs primary septae formation during distal lung morphogenesis. Dev. Biol. 2007, 308, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N. The role of vascular endothelial growth factor in pathological angiogenesis. Breast Cancer Res. Treat 1995, 36, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Garnier, V.; Traboulsi, W.; Salomon, A.; Brouillet, S.; Fournier, T.; Winkler, C.; Desvergne, B.; Hoffmann, P.; Zhou, Q.Y.; Congiu, C.; et al. PPARgamma controls pregnancy outcome through activation of EG-VEGF: New insights into the mechanism of placental development. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E357–E369. [Google Scholar] [CrossRef] [Green Version]
- Reynaud, D.; Sergent, F.; Abi Nahed, R.; Brouillet, S.; Benharouga, M.; Alfaidy, N. EG-VEGF Maintenance Over Early Gestation to Develop a Pregnancy-Induced Hypertensive Animal Model. Methods Mol. Biol. 2018, 1710, 317–324. [Google Scholar] [CrossRef]
- Sergent, F.; Hoffmann, P.; Brouillet, S.; Garnier, V.; Salomon, A.; Murthi, P.; Benharouga, M.; Feige, J.J.; Alfaidy, N. Sustained Endocrine Gland-Derived Vascular Endothelial Growth Factor Levels Beyond the First Trimester of Pregnancy Display Phenotypic and Functional Changes Associated With the Pathogenesis of Pregnancy-Induced Hypertension. Hypertension 2016, 68, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Traboulsi, W.; Sergent, F.; Boufettal, H.; Brouillet, S.; Slim, R.; Hoffmann, P.; Benlahfid, M.; Zhou, Q.Y.; Balboni, G.; Onnis, V.; et al. Antagonism of EG-VEGF Receptors as Targeted Therapy for Choriocarcinoma Progression In Vitro and In Vivo. Clin. Cancer Res. 2017, 23, 7130–7140. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, P.; Saoudi, Y.; Benharouga, M.; Graham, C.H.; Schaal, J.P.; Mazouni, C.; Feige, J.J.; Alfaidy, N. Role of EG-VEGF in human placentation: Physiological and pathological implications. J. Cell Mol. Med. 2009, 13, 2224–2235. [Google Scholar] [CrossRef] [Green Version]
- Brouillet, S.; Hoffmann, P.; Thomas-Cadi, C.; Bergues, U.; Feige, J.J.; Alfaidy, N.; Hennebicq, S. PROK1, prognostic marker of embryo implantation? Gynecol. Obstet Fertil 2013, 41, 562–565. [Google Scholar] [CrossRef]
- Brouillet, S.; Murthi, P.; Hoffmann, P.; Salomon, A.; Sergent, F.; De Mazancourt, P.; Dakouane-Giudicelli, M.; Dieudonne, M.N.; Rozenberg, P.; Vaiman, D.; et al. EG-VEGF controls placental growth and survival in normal and pathological pregnancies: Case of fetal growth restriction (FGR). Cell Mol. Life Sci. 2013, 70, 511–525. [Google Scholar] [CrossRef]
- Denison, F.C.; Battersby, S.; King, A.E.; Szuber, M.; Jabbour, H.N. Prokineticin-1: A novel mediator of the inflammatory response in third-trimester human placenta. Endocrinology 2008, 149, 3470–3477. [Google Scholar] [CrossRef]
- Balboni, G.; Lazzari, I.; Trapella, C.; Negri, L.; Lattanzi, R.; Giannini, E.; Nicotra, A.; Melchiorri, P.; Visentin, S.; Nuccio, C.D.; et al. Triazine compounds as antagonists at Bv8-prokineticin receptors. J. Med. Chem. 2008, 51, 7635–7639. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Noda, K.; Yoshida, K.; Otani, K.; Yoshiga, M.; Oto, Y.; Saito, S.; Kurosaka, D. Prokineticin 2 antagonist, PKRA7 suppresses arthritis in mice with collagen-induced arthritis. BMC Musculoskelet Disord. 2016, 17, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, O.; Weiss, I.D.; Niu, G.; Balboni, G.; Congiu, C.; Onnis, V.; Kiesewetter, D.O.; Lattanzi, R.; Salvadori, S.; Chen, X. Prokineticin receptor 1 antagonist PC-10 as a biomarker for imaging inflammatory pain. J. Nucl. Med. 2011, 52, 600–607. [Google Scholar] [CrossRef] [Green Version]
- Maftei, D.; Ratano, P.; Fusco, I.; Marconi, V.; Squillace, S.; Negri, L.; Severini, C.; Balboni, G.; Steardo, L.; Bronzuoli, M.R.; et al. The prokineticin receptor antagonist PC1 rescues memory impairment induced by beta amyloid administration through the modulation of prokineticin system. Neuropharmacology 2019, 158, 107739. [Google Scholar] [CrossRef]
- Moschetti, G.; Amodeo, G.; Paladini, M.S.; Molteni, R.; Balboni, G.; Panerai, A.; Sacerdote, P.; Franchi, S. Prokineticin 2 promotes and sustains neuroinflammation in vincristine treated mice: Focus on pain and emotional like behavior. Brain Behav. Immun. 2019, 82, 422–431. [Google Scholar] [CrossRef]
- Holloway, A.C.; Salomon, A.; Soares, M.J.; Garnier, V.; Raha, S.; Sergent, F.; Nicholson, C.J.; Feige, J.J.; Benharouga, M.; Alfaidy, N. Characterization of the adverse effects of nicotine on placental development: In vivo and in vitro studies. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E443–E456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landucci, E.; Lattanzi, R.; Gerace, E.; Scartabelli, T.; Balboni, G.; Negri, L.; Pellegrini-Giampietro, D.E. Prokineticins are neuroprotective in models of cerebral ischemia and ischemic tolerance in vitro. Neuropharmacology 2016, 108, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Congiu, C.; Onnis, V.; Deplano, A.; Salvadori, S.; Marconi, V.; Maftei, D.; Negri, L.; Lattanzi, R.; Balboni, G. A new convenient synthetic method and preliminary pharmacological characterization of triazinediones as prokineticin receptor antagonists. Eur. J. Med. Chem. 2014, 81, 334–340. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Burton, K.J.; Neal, M.L.; Qiao, Y.; Kanthasamy, A.G.; Sun, Y.; Xu, X.; Ma, Y.; Li, X. Differential arousal regulation by prokineticin 2 signaling in the nocturnal mouse and the diurnal monkey. Molecular. Brain 2016, 9, 78. [Google Scholar] [CrossRef] [Green Version]
- Sahgal, N.; Canham, L.N.; Canham, B.; Soares, M.J. Rcho-1 trophoblast stem cells: A model system for studying trophoblast cell differentiation. Methods Mol. Med. 2006, 121, 159–178. [Google Scholar]
- Sahgal, N.; Canham, L.N.; Konno, T.; Wolfe, M.W.; Soares, M.J. Modulation of trophoblast stem cell and giant cell phenotypes: Analyses using the Rcho-1 cell model. Differentiation 2005, 73, 452–462. [Google Scholar] [CrossRef]
- Goryszewska, E.; Kaczynski, P.; Baryla, M.; Waclawik, A. Pleiotropic role of prokineticin 1 in the porcine endometrium during pregnancy establishment and embryo implantation dagger. Biol. Reprod. 2021, 104, 181–196. [Google Scholar] [CrossRef]
- Moschetti, G.; Kalpachidou, T.; Amodeo, G.; Lattanzi, R.; Sacerdote, P.; Kress, M.; Franchi, S. Prokineticin Receptor Inhibition With PC1 Protects Mouse Primary Sensory Neurons From Neurotoxic Effects of Chemotherapeutic Drugs in vitro. Front. Immunol. 2020, 11, 2119. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.C.; Baczyk, D.; Dobric, N.; Hemberger, M.; Hughes, M.; Simmons, D.G.; Yamamoto, H.; Kingdom, J.C. Genes, development and evolution of the placenta. Placenta 2003, 24, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Marsango, S.; Bonaccorsi di Patti, M.C.; Barra, D.; Miele, R. Evidence that prokineticin receptor 2 exists as a dimer in vivo. Cell Mol. Life Sci. 2011, 68, 2919–2929. [Google Scholar] [CrossRef] [PubMed]
- Ngan, E.S.; Lee, K.Y.; Sit, F.Y.; Poon, H.C.; Chan, J.K.; Sham, M.H.; Lui, V.C.; Tam, P.K. Prokineticin-1 modulates proliferation and differentiation of enteric neural crest cells. Biochim. Biophys. Acta 2007, 1773, 536–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slessareva, J.E.; Ma, H.; Depree, K.M.; Flood, L.A.; Bae, H.; Cabrera-Vera, T.M.; Hamm, H.E.; Graber, S.G. Closely related G-protein-coupled receptors use multiple and distinct domains on G-protein alpha-subunits for selective coupling. J. Biol. Chem. 2003, 278, 50530–50536. [Google Scholar] [CrossRef] [Green Version]
- Heck, D.; Wortmann, S.; Kraus, L.; Ronchi, C.L.; Sinnott, R.O.; Fassnacht, M.; Sbiera, S. Role of Endocrine Gland-Derived Vascular Endothelial Growth Factor (EG-VEGF) and Its Receptors in Adrenocortical Tumors. Horm Cancer 2015, 6, 225–236. [Google Scholar] [CrossRef] [Green Version]
- Lozneanu, L.; Avadanei, R.; Cimpean, A.M.; Giusca, S.E.; Amalinei, C.; Caruntu, I.D. Relationship between the Proangiogenic Role of Eg-Vegf, Clinicopathological Characteristics and Survival in Tumoral Ovary. Rev. Med. Chir. Soc. Med. Nat. Iasi 2015, 119, 461–465. [Google Scholar]
- Murthi, P.; Brouillet, S.; Pratt, A.; Borg, A.; Kalionis, B.; Goffin, F.; Tsatsaris, V.; Munaut, C.; Feige, J.; Benharouga, M.; et al. An EG-VEGF-dependent decrease in homeobox gene NKX3.1 contributes to cytotrophoblast dysfunction: A possible mechanism in human fetal growth restriction. Mol. Med. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bardin, N.; Murthi, P.; Alfaidy, N. Normal and pathological placental angiogenesis. BioMed Res. Int. 2015, 2015, 354359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado-Perez, D.; Evans, J.; Denison, F.; Millar, R.P.; Jabbour, H.N. Potential roles of the prokineticins in reproduction. Trends Endocrinol. Metab. 2007, 18, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.L.; Denison, F.C.; Evans, J.; Durno, K.; Williams, A.R.; Entrican, G.; Critchley, H.O.; Jabbour, H.N.; Horne, A.W. Evidence of prokineticin dysregulation in fallopian tube from women with ectopic pregnancy. Fertil Steril 2010, 94, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Li, J.J.; Bay, B.H.; Yung, L.Y. Investigating the antiproliferative activity of high affinity DNA aptamer on cancer cells. PLoS ONE 2013, 8, e50964. [Google Scholar] [CrossRef] [Green Version]
- Kaur, H.; Yung, L.Y. Probing high affinity sequences of DNA aptamer against VEGF165. PLoS ONE 2012, 7, e31196. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.P.; Zhang, C.; Li, J.D.; Luo, Z.D.; Amadesi, S.; Bunnett, N.; Zhou, Q.Y. Impaired pain sensation in mice lacking prokineticin 2. Mol. Pain 2006, 2, 35. [Google Scholar] [CrossRef] [Green Version]
- Maftei, D.; Vellani, V.; Artico, M.; Giacomoni, C.; Severini, C.; Lattanzi, R. Abnormal Pain Sensation in Mice Lacking the Prokineticin Receptor PKR2: Interaction of PKR2 with Transient Receptor Potential TRPV1 and TRPA1. Neuroscience 2020, 427, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Negri, L.; Lattanzi, R.; Giannini, E.; Colucci, M.; Margheriti, F.; Melchiorri, P.; Vellani, V.; Tian, H.; De Felice, M.; Porreca, F. Impaired nociception and inflammatory pain sensation in mice lacking the prokineticin receptor PKR1: Focus on interaction between PKR1 and the capsaicin receptor TRPV1 in pain behavior. J. Neurosci. 2006, 26, 6716–6727. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.C.; Li, X.M.; Wang, X.J.; Liu, Y.Q.; Qiu, F.; Wu, D.; Gan, Y.B.; Wang, B.H.; Hu, W.P. Prokineticin 2 suppresses GABA-activated current in rat primary sensory neurons. Neuropharmacology 2010, 59, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Lattanzi, R.; Maftei, D.; Marconi, V.; Florenzano, F.; Franchi, S.; Borsani, E.; Rodella, L.F.; Balboni, G.; Salvadori, S.; Sacerdote, P.; et al. Prokineticin 2 upregulation in the peripheral nervous system has a major role in triggering and maintaining neuropathic pain in the chronic constriction injury model. BioMed Res. Int. 2015, 2015, 301292. [Google Scholar] [CrossRef] [PubMed]
- Collet, C.; Abi Nahed, R.; Gemy, K.; Traboulsi, W.; Lemaitre, N.; Bolze, P.-A.; Hoffmann, P.; Benharouga, M.; Alfaidy, N. Multifaceted Functions of Prokineticins in Reproductive Cancers and Proposed Associated Therapies. J. Cancer Sci. Clin. Ther. 2020, 4, 408–423. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward | Reverse | Amplicon Size | Tm (°C) |
|---|---|---|---|---|
| mGapdh | AACGACCCCTTCATTGAC | TCCACGACATACTCAGCA | 191 | 57 |
| mMash 2 | GGTGACTCCTGGTGGACCTA | TCCGGAAGATGGAAGATGTC | 151 | 56 |
| mPl1 | TGTCATACTGCTTCCATCCATAC | CCAGGTGTTTCAGAGGTTCTT | 125 | 60 |
| mPl2 | ACGCCCATGATCTTGCTTCA | TGGCAGGGGCTTAACATCAG | 114 | 60 |
| mProliferin | TGTGTGCAATGAGGAATGGT | TAGTGTGTGAGCCTGGCTTG | 223 | 58 |
| mGcm1 | TTTTTCCAGTCCAAAGGCGAG | TGACTCGGGATTTCAGCAGG | 164 | 60 |
| mCd31 | GCATCGGCAAAGTGGTCAA | GTTCCATTTTCGGACTGG C | 145 | 60 |
| mHand1 | AAGCAAGCGGAAAAGGAGT | GGCCTGGTCTCACTGGTTTA | 159 | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reynaud, D.; Sergent, F.; Abi Nahed, R.; Traboulsi, W.; Collet, C.; Marquette, C.; Hoffmann, P.; Balboni, G.; Zhou, Q.-Y.; Murthi, P.; et al. Evidence-Based View of Safety and Effectiveness of Prokineticin Receptors Antagonists during Pregnancy. Biomedicines 2021, 9, 309. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030309
Reynaud D, Sergent F, Abi Nahed R, Traboulsi W, Collet C, Marquette C, Hoffmann P, Balboni G, Zhou Q-Y, Murthi P, et al. Evidence-Based View of Safety and Effectiveness of Prokineticin Receptors Antagonists during Pregnancy. Biomedicines. 2021; 9(3):309. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030309
Chicago/Turabian StyleReynaud, Deborah, Frederic Sergent, Roland Abi Nahed, Wael Traboulsi, Constance Collet, Christel Marquette, Pascale Hoffmann, Gianfranco Balboni, Qun-Yong Zhou, Padma Murthi, and et al. 2021. "Evidence-Based View of Safety and Effectiveness of Prokineticin Receptors Antagonists during Pregnancy" Biomedicines 9, no. 3: 309. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9030309