
MicroRNAs Regulating Tumor and Immune Cell Interactions in the Prediction of Relapse in Early Stage Breast Cancer

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients’ Characteristics and Sample Collection
2.2. RNA Isolation
2.3. Quantitative Real-Time PCR Analysis of miRNA Expression
2.4. Statistical Analysis
3. Results
3.1. Patients’ Characteristics and Study Design
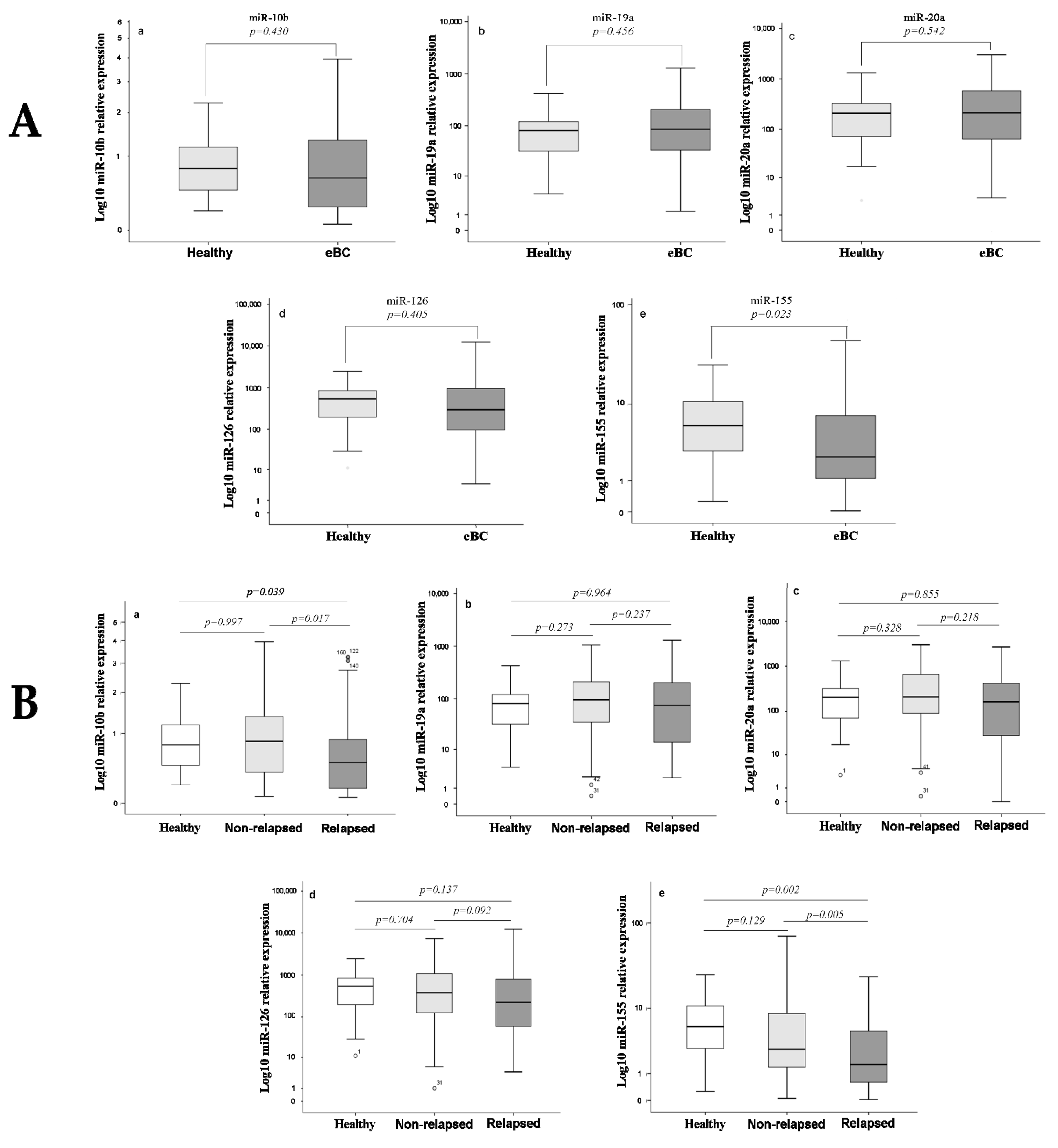
3.2. Differential Expression of miRNAs among Healthy Women and Breast Cancer Patients
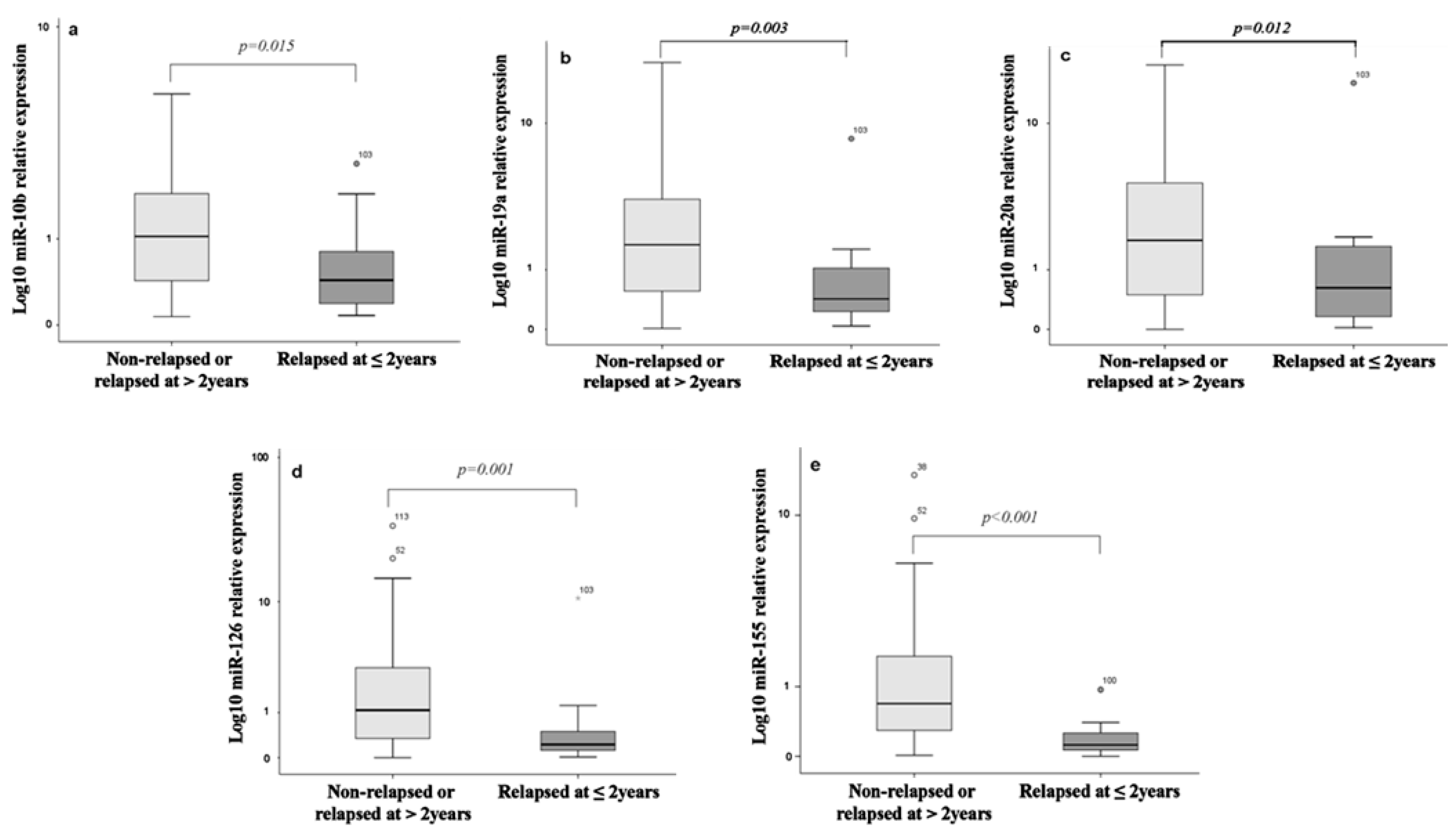
3.3. Differential Expression of miRNAs among Relapsed and Non-Relapsed Patients
3.4. Performance of miRNAs in Predictive Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.M.; Jemal, A. Cancer statistics, 2018. CA A Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Ghiso, J.A. Models, mechanisms and clinical evidence for cancer dormancy. Nat. Rev. Cancer 2007, 7, 834–846. [Google Scholar] [CrossRef] [Green Version]
- Dunn, G.P.; Bruce, A.T.; Ikeda, H.; Old, L.J.; Schreiber, R.D. Cancer immunoediting: From immunosurveillance to tumor escape. Nat. Immunol. 2002, 3, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Kumar, Y. Cancer stem cells and tumour immunoediting: Putting two and two together. Expert Rev. Clin. Immunol. 2016, 12, 605–607. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Nicoloso, M.S.; Spizzo, R.; Shimizu, M.; Rossi, S.; Calin, G.A. MicroRNAs—The micro steering wheel of tumour metastases. Nat. Rev. Cancer 2009, 9, 293–302. [Google Scholar] [CrossRef]
- Larrea, E.; Sole, C.; Lawrie, C.H.; Manterola, L.; Goicoechea, I.; Armesto, M.; Arestin, M.; Caffarel, M.M.; Araujo, A.M.; Araiz, M.; et al. New concepts in cancer biomarkers: Circulating miRNAs in liquid biopsies. Int. J. Mol. Sci. 2016, 17, 627. [Google Scholar] [CrossRef] [Green Version]
- Paladini, L.; Fabris, L.; Bottai, G.; Raschioni, C.; Calin, G.A.; Santarpia, L. Targeting microRNAs as key modulators of tumor immune response. J. Exp. Clin. Cancer Res. 2016, 35, 103–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschberger, S.; Hinske, L.C.; Kreth, S. MiRNAs: Dynamic regulators of immune cell functions in inflammation and cancer. Cancer Lett. 2018, 431, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Xu, L.; Jiao, Y.; Luo, S.; Li, A.; Wu, K. The role of cancer-derived microRNAs in cancer immune escape. J. Hematol. Oncol. 2020, 13, 251–314. [Google Scholar] [CrossRef] [Green Version]
- Vignard, V.; Labbι, M.; Marec, N.; Andrι-Grιgoire, G.; Jouand, N.; Fonteneau, J.-F.; Labarriθre, N.; Fradin, D. MicroRNAs in tumor exosomes drive immune escape in melanoma. Cancer Immunol. Res. 2019, 8, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Stern-Ginossar, N.; Gur, C.; Biton, M.; Horwitz, E.; Elboim, M.; Stanietsky, N.; Mandelboim, M.; Mandelboim, O. Human microRNAs regulate stress-induced immune responses mediated by the receptor NKG2D. Nat. Immunol. 2008, 9, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Z.; Chen, C.; Liu, Y.; Si, Q.; Chuang, T.-H.; Li, N.; Gomez-Cabrero, A.; Reisfeld, R.A.; Xiang, R.; et al. MicroRNA-19a-3p inhibits breast cancer progression and metastasis by inducing macrophage polarization through downregulated expression of Fra-1 proto-oncogene. Oncogene 2014, 33, 3014–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agudo, J.; Ruzo, A.; Tung, N.; Salmon, H.; Leboeuf, M.; Hashimoto, D.; Becker, C.; Garrett-Sinha, L.-A.; Baccarini, A.; Merad, M.; et al. The miR-126–VEGFR2 axis controls the innate response to pathogen-associated nucleic acids. Nat. Immunol. 2013, 15, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Schambach, F.; DeJong, C.S.; Hammond, S.M.; Reiner, S.L. Micro-RNA-155 inhibits IFN-γ signaling in CD4+ T cells. Eur. J. Immunol. 2009, 40, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Trotta, R.; Chen, L.; Ciarlariello, D.; Josyula, S.; Mao, C.; Costinean, S.; Yu, L.; Butchar, J.P.; Tridandapani, S.; Croce, C.M.; et al. miR-155 regulates IFN-γ production in natural killer cells. Blood 2012, 119, 3478–3485. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Fioravanti, J.; Zhu, W.; Wang, H.; Wu, T.; Hu, J.; Lacey, N.E.; Gautam, S.; Le Gall, J.B.; Yang, X.; et al. miR-155 harnesses Phf19 to potentiate cancer immunotherapy through epigenetic reprogramming of CD8+ T cell fate. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Papadaki, C.; Stoupis, G.; Tsalikis, L.; Monastirioti, A.; Papadaki, M.; Maliotis, N.; Stratigos, M.; Mastrostamatis, G.; Mavroudis, D.; Agelaki, S. Circulating miRNAs as a marker of metastatic disease and prognostic factor in metastatic breast cancer. Oncotarget 2019, 10, 966–981. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jia, Y.; Wei, Y. Modulators of MicroRNA function in the immune system. Int. J. Mol. Sci. 2020, 21, 2357. [Google Scholar] [CrossRef]
- Ma, R.; Jiang, T.; Kang, X. Circulating microRNAs in cancer: Origin, function and application. J. Exp. Clin. Cancer Res. 2012, 31, 38. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Donninger, H.; Eaton, J.; Yaddanapudi, K. Regulatory role of immune cell-derived extracellular vesicles in cancer: The message is in the envelope. Front. Immunol. 2020, 11, 1525. [Google Scholar] [CrossRef]
- de Candia, P.; De Rosa, V.; Casiraghi, M.; Matarese, G. Extracellular RNAs: A secret arm of immune system regulation. J. Biol. Chem. 2016, 291, 7221–7228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtale, G. MiRNAs at the Crossroads between Innate Immunity and Cancer: Focus on Macrophages. Cells 2018, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Testa, U.; Pelosi, E.; Castelli, G.; Labbaye, C. miR-146 and miR-155: Two key modulators of immune response and tumor development. Non-Coding RNA 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Yin, Y.; Li, N.; Zhu, D.; Zhang, J.; Zhang, C.-Y.; Zen, K. Re-polarization of tumor-associated macrophages to pro-inflammatory M1 macrophages by microRNA-155. J. Mol. Cell Biol. 2012, 4, 341–343. [Google Scholar] [CrossRef]
- Hsin, J.-P.; Lu, Y.; Loeb, G.B.; Leslie, C.S.; Rudensky, A.Y. The effect of cellular context on miR-155-mediated gene regulation in four major immune cell types. Nat. Immunol. 2018, 19, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Zonari, E.; Pucci, F.; Saini, M.; Mazzieri, R.; Politi, L.S.; Gentner, B.; Naldini, L. A role for miR-155 in enabling tumor-infiltrating innate immune cells to mount effective antitumor responses in mice. Blood 2013, 122, 243–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffaker, T.B.; Hu, R.; Runtsch, M.C.; Bake, E.; Chen, X.; Zhao, J.; Round, J.L.; Baltimore, D.; O’Connell, R.M. Epistasis between MicroRNAs 155 and 146a during T cell-mediated antitumor immunity. Cell Rep. 2012, 2, 1697–1709. [Google Scholar] [CrossRef] [Green Version]
- Dudda, J.C.; Salaun, B.; Ji, Y.; Palmer, D.C.; Monnot, G.C.; Merck, E.; Boudousquie, C.; Utzschneider, D.T.; Escobar, T.M.; Perret, R.; et al. MicroRNA-155 is required for effector CD8+ T cell responses to virus infection and cancer. Immunity 2013, 38, 742–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, L.; Fan, J.; Donye, D.; Dominguez, D.; Zhang, Y.; Curiel, T.J.; Fang, D.; Kuzel, T.M.; Zhang, B. Host miR155 promotes tumor growth through a myeloid-derived suppressor cell—dependent mechanism. Cancer Res. 2015, 75, 519–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorio, M.V.; Ferracin, M.; Mιnard, S.; Palazzo, J.P.; Rosenberg, A.; Musiani, P.; Volinia, S.; Nenci, I.; Calin, G.A.; Querzoli, P.; et al. MicroRNA gene expression deregulation in human breast cancer. Cancer Res. 2005, 65, 7065–7070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Zhang, H.-W.; Lu, M.-H.; He, X.-H.; Li, Y.; Gu, H.; Liu, M.-F.; Wang, E.-D. MicroRNA-155 functions as an oncomiR in breast cancer by targeting the suppressor of cytokine signaling 1 gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nariman-Saleh-Fam, Z.; Saadatian, Z.; Daraei, A.; Mansoori, Y.; Bastami, M.; Tavakkoli-Bazzaz, J. The intricate role of miR-155 in carcinogenesis: Potential implications for esophageal cancer research. Biomarkers Med. 2019, 13, 147–159. [Google Scholar] [CrossRef]
- Xiang, X.; Zhuang, X.; Ju, S.; Zhang, S.; Jiang, H.; Mu, J.; Zhang, L.; Miller, D.; Grizzle, W.E.; Zhang, H.-G. miR-155 promotes macroscopic tumor formation yet inhibits tumor dissemination from mammary fat pads to the lung by preventing EMT. Oncogene 2011, 30, 3440–3453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Jiang, W.; Xu, X.; Zheng, X. Serum microRNA-155 in early diagnosis and prognosis of breast cancer. Int. J. Clin. Exp. Med. 2016, 9, 10289–10296. [Google Scholar]
- Swellam, M.; Ramadan, A.; El-Hussieny, E.A.; Bakr, N.M.; Hassan, N.M.; Sobeih, M.E.; Ezzelarab, L.R. Clinical significance of blood-based miRNAs as diagnostic and prognostic nucleic acid markers in breast cancer: Comparative to conventional tumor markers. J. Cell. Biochem. 2019, 120, 12321–12330. [Google Scholar] [CrossRef]
- Bašová, P.; Peta, M.; Sochor, M.; Stopka, T. Prediction Potential of Serum miR-155 and miR-24 for Relapsing Early Breast Cancer. Int. J. Mol. Sci. 2017, 18, 2116. [Google Scholar] [CrossRef] [Green Version]
- Farazi, T.A.; Horlings, H.M.; Kreike, B.; Sie, D.; Hovestadt, V.; Wessels, L.F.A.; Van De Vijver, M.J.; Tuschl, T.; Hoeve, J.J.T.; Mihailovic, A.; et al. MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Cancer Res. 2011, 71, 4443–4453. [Google Scholar] [CrossRef] [Green Version]
- Olive, V.; Bennett, M.J.; Walker, J.C.; Ma, C.; Jiang, I.; Cordon-Cardo, C.; Li, Q.-J.; Lowe, S.W.; Hannon, G.J.; He, L. miR-19 is a key oncogenic component of mir-17-92. Genes Dev. 2009, 23, 2839–2849. [Google Scholar] [CrossRef] [Green Version]
- Mu, P.; Han, Y.-C.; Betel, D.; Yao, E.; Squatrito, M.; Ogrodowski, P.; De Stanchina, E.; D’Andrea, A.; Sander, C.; Ventura, A. Genetic dissection of the miR-17 92 cluster of microRNAs in Myc-induced B-cell lymphomas. Genes Dev. 2009, 23, 2806–2811. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Liu, M.; Ma, F.; Luo, Y.; Cai, R.; Wang, L.; Xu, N.; Xu, B. Circulating miR-19a and miR-205 in serum may predict the sensitivity of luminal a subtype of breast cancer patients to neoadjuvant chemotherapy with epirubicin plus paclitaxel. PLoS ONE 2014, 9, e104870. [Google Scholar] [CrossRef] [Green Version]
- Alunni-Fabbroni, M.; on behalf of the SUCCESS Study Group; Majunke, L.; Trapp, E.K.; Tzschaschel, M.; Mahner, S.; Fasching, P.A.; Fehm, T.; Schneeweiss, A.; Beck, T.; et al. Whole blood microRNAs as potential biomarkers in post-operative early breast cancer patients. BMC Cancer 2018, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Sochor, M.; Basova, P.; Pesta, M.; Dusilkova, N.; Bartos, J.; Burda, P.; Pospisil, V.; Stopka, T. Oncogenic MicroRNAs: miR-155, miR-19a, miR-181b, and miR-24 enable monitoring of early breast cancer in serum. BMC Cancer 2014, 14, 448. [Google Scholar] [CrossRef] [Green Version]
- Tsukerman, P.; Stern-Ginossar, N.; Lankry, D.; Mandelboim, O.; Gur, C.; Glasner, A.; Nachmani, D.; Bauman, Y.; Yamin, R.; Vitenshtein, A.; et al. MiR-10b Downregulates the stress-induced cell surface molecule MICB, a critical ligand for cancer cell recognition by natural killer cells. Cancer Res. 2012, 72, 5463–5472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nat. Cell Biol. 2007, 449, 682–688. [Google Scholar] [CrossRef] [PubMed]
- Gee, H.E.; Buffa, F.M.; Camps, C.; Ramachandran, A.; Leek, R.; Taylor, M.; Patil, M.; Sheldon, H.; Betts, G.; Homer, J.; et al. The small-nucleolar RNAs commonly used for microRNA normalisation correlate with tumour pathology and prognosis. Br. J. Cancer 2011, 104, 1168–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gee, H.E.; Camps, C.; Buffa, F.M.; Colella, S.; Sheldon, H.; Gleadle, J.M.; Ragoussis, J.; Harris, A.L. MicroRNA-10b and breast cancer metastasis. Nat. Cell Biol. 2008, 455, E8–E9. [Google Scholar] [CrossRef]
- Khalighfard, S.; Alizadeh, A.M.; Irani, S.; Omranipour, R. Plasma miR-21, miR-155, miR-10b, and Let-7a as the potential biomarkers for the monitoring of breast cancer patients. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jurkovicova, D.; Smolkova, B.; Magyerkova, M.; Sestakova, Z.; Kajabova, V.H.; Kulcsar, L.; Zmetakova, I.; Kalinkova, L.; Krivulcik, T.; Karaba, M.; et al. Down-regulation of traditional oncomiRs in plasma of breast cancer patients. Oncotarget 2017, 8, 77369–77384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Zheng, Z.; Guo, J.; Ding, X. Correlation and quantitation of microRNA aberrant expression in tissues and sera from patients with breast tumor. Gynecol. Oncol. 2010, 119, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Tavazoie, S.F.; Alarcσn, C.; Oskarsson, T.; Padua, D.; Wang, Q.; Bos, P.D.; Gerald, W.L.; Massaguι, J. Endogenous human microRNAs that suppress breast cancer metastasis. Nat. Cell Biol. 2008, 451, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Liu, Q.; Guo, F.; Zhang, Z.; Yang, R.; Mi, S.; Liang, X.; Zhang, Z.; Su, X.; Liu, J.; et al. Both miR-17-5p and miR-20a Alleviate suppressive potential of myeloid-derived suppressor cells by modulating STAT3 expression. J. Immunol. 2011, 186, 4716–4724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Liu, M.; Li, Y.; Nie, Y.; Mi, Q.; Zhao, S. Ovarian tumor-associated microRNA-20a decreases natural killer cell cytotoxicity by downregulating MICA/B expression. Cell. Mol. Immunol. 2014, 11, 495–502. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, P.; Sun, T.; Li, D.; Xu, X.; Rui, Y.; Li, C.; Chong, M.; Ibrahim, T.; Mercatali, L.; et al. miR-126 and miR-126* repress recruitment of mesenchymal stem cells and inflammatory monocytes to inhibit breast cancer metastasis. Nat. Cell Biol. 2013, 15, 284–294. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; O’Hara, S.P.; Chen, X.-M. MicroRNA regulation of innate immune responses in epithelial cells. Cell. Mol. Immunol. 2011, 8, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Rueda, O.M.; Sammut, S.-J.; Seoane, J.A.; Chin, S.-F.; Caswell-Jin, J.L.; Callari, M.; Batra, R.; Pereira, B.; Bruna, A.; Ali, H.R.; et al. Dynamics of breast-cancer relapse reveal late-recurring ER-positive genomic subgroups. Nature 2019, 567, 399–404. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| All Patients | Non Relapse | Relapse | ||
|---|---|---|---|---|
| Characteristic | n (%) | n (%) | n (%) | p-Value |
| Number of patients | 140 | 94 (67.2) | 46 (32.8) | |
| Age, median (range) | 55 (27–82) | 54 (35–79) | 56 (27–82) | ns * |
| Menopausal status | ns * | |||
| Premenopausal | 53 (37.9) | 38 (40.4) | 15 (32.6) | |
| Postmenopausal | 87 (62.1) | 56 (59.6) | 31 (67.4) | |
| Tumor size (cm) | 0.031 * | |||
| T1 | 62 (44.3) | 44 (46.8) | 18 (39.1) | |
| T2 | 70 (50.0) | 48 (51.1) | 22 (47.9) | |
| T3 | 8 (5.7) | 2 (2.1) | 6 (13.0) | |
| Grade | 0.034 * | |||
| I | 5 (3.6) | 5 (5.3) | ||
| II | 56 (40.0) | 42 (44.7) | 14 (30.4) | |
| III | 67 (47.9) | 39 (41.5) | 28 (60.9) | |
| Lobular | 8 (5.7) | 4 (4.3) | 4 (8.7) | |
| Unknown | 4 (2.9) | 4 (4.3) | ||
| Infiltrated lymph nodes | <0.001 * | |||
| 0 | 60 (42.9) | 49 (52.1) | 11 (23.9) | |
| 1–3 | 50 (35.7) | 34(36.2) | 16 (34.8) | |
| ≥4 | 30 (21.4) | 11 (11.7) | 19 (41.3) | |
| ER status | ns * | |||
| Positive | 88 (62.9) | 58 (61.7) | 30 (65.2) | |
| Negative | 52 (37.1) | 36(38.3) | 16 (34.8) | |
| PR status | ns * | |||
| Positive | 88 (62.9) | 60(63.8) | 28 (60.9) | |
| Negative | 52 (37.1) | 34 (36.2) | 18 (39.1) | |
| Her2 status | ns * | |||
| Positive | 19 (13.6) | 11(11.7) | 8 (17.4) | |
| Negative | 121 (86.4) | 83(88.3) | 38 (82.6) | |
| Adjuvant chemotherapy | ||||
| Anthracyclines-based | 10 (7.1) | 7 (7.4) | 3 (6.5) | |
| Taxanes + Antracyclines | 95 (67.9) | 59 (62.9) | 36 (78.3) | |
| Taxanes-based | 26 (18.6) | 20(21.3) | 6 (13) | |
| Other | 3 (2.1) | 3 (3.3) | ||
| None | 6 (4.3) | 5 (5.2) | 1 (2.2) |
| Early Relapse | Late Relapse | p-Value | |
|---|---|---|---|
| Characteristic | n (%) | n | |
| Number of patients | 16 (11.4) | 17 | |
| Age, median (range) | 56 (27–82) | 55 (41–74) | ns * |
| Menopausal status | ns * | ||
| Premenopausal | 7 (43.8) | 5 (29.4) | |
| Postmenopausal | 9 (56.2) | 12 (70.2) | |
| Tumor size (cm) | ns * | ||
| T1 | 5 (31.3) | 8 (47.1) | |
| T2 | 9 (56.3) | 7 (41.2) | |
| T3 | 2 (12.5) | 2 (11.8) | |
| Grade | ns * | ||
| I | |||
| II | 6 (37.5) | 3 (17.6) | |
| III | 10 (62.5) | 14 (82.4) | |
| Infiltrated lymph nodes | ns * | ||
| 0 | 6 (37.5) | 1 (5.9) | |
| 1–3 | 6 (37.5) | 7 (41.2) | |
| ≥4 | 4 (25) | 9 (52.9) | |
| ER status | 0.002 * | ||
| Positive | 7 (43.8) | 16 (94.1) | |
| Negative | 9 (56.2) | 1 (5.9) | |
| PR status | 0.009 * | ||
| Positive | 7 (43.8) | 15 (88.2) | |
| Negative | 9 (56.3) | 2 (11.8) | |
| Her2 status | ns * | ||
| Positive | 3 (18.8) | 4 (23.5) | |
| Negative | 13 (81.2) | 13 (76.5) | |
| Adjuvant chemotherapy | ns * | ||
| Anthracyclines-based | 1 (6.3) | 1 (5.9) | |
| Taxanes+Antracyclines | 11 (68.7) | 15 (88.2) | |
| Taxanes-based | 3 (18.7) | 1 (5.9) | |
| Others | 1 (6.3) |
| Potential Predictors | Cutoff | Sensitivity (%) | Specificity (%) | AUC (95% CI) | p-Value | PPV (%) | NPV (%) | Mean AUC | Mean p-Value |
|---|---|---|---|---|---|---|---|---|---|
| (10-Fold Cross Validation) | (10-Fold Cross Validation) | ||||||||
| Relapse | |||||||||
| miR-10b | 0.920 | 69.6 | 56.4 | 0.619 (0.517–0.721) | 0.022 | 43.84 | 79.1 | 0.605 | 0.01891 |
| miR-155 | 0.405 | 56.5 | 69.1 | 0.647 (0.550–0.745) | 0.005 | 46.3 | 75.6 | 0.684 | 0.00643 |
| LN and grade | 0.416 | 85.4 | 50.0 | 0.726 (0.631–0.821) | <0.001 | 72.7 | 70.0 | 0.699 | 0.00121 |
| miR-155 and LN and grade | 0.292 | 85.4 | 80.0 | 0.861 (0.788–0.935) | <0.001 | 80.6 | 82.2 | 0.767 | 0.00329 |
| Early relapse (≤2 years) | |||||||||
| miR-10b | 0.855 | 81.3 | 54.8 | 0.688 (0.556–0.819) | 0.015 | 18.8 | 95.8 | 0.704 | 0.01813 |
| miR-19a | 1.560 | 93.8 | 53.2 | 0.729 (0.608–0.850) | 0.003 | 19.4 | 97.1 | 0.725 | 0.00410 |
| miR-20a | 1.940 | 93.8 | 49.2 | 0.694 (0.564–0.823) | 0.012 | 19.2 | 98.4 | 0.689 | 0.01011 |
| miR-126 | 0.530 | 81.0 | 65.3 | 0.745 (0.620–0.829) | 0.001 | 22.4 | 96.3 | 0.739 | 0.00082 |
| miR-155 | 0.445 | 93.8 | 64.5 | 0.855 (0.722–0.939) | <0.001 | 27.3 | 98.8 | 0.860 | 0.00078 |
| miR-19a, miR-20a | 0.265 | 93.8 | 64.5 | 0.816 (0.732–0.900) | <0.001 | 50.0 | 89.1 | 0.834 | 0.00071 |
| Late relapse (≥5 years) | |||||||||
| miR-10b | 1.205 | 88.2 | 51.1 | 0.642 (0.516–0.768) | 0.063 | 22.6 | 93.9 | 0.628 | 0.07835 |
| LN and grade | 0.115 | 82.4 | 88.8 | 0.898 (0.806–0.990) | <0.001 | 69.2 | 90.5 | 0.865 | 0.00089 |
| miR-10b and LN and grade | 0.467 | 88.2 | 98.8 | 0.971(0.923–1.000) | <0.001 | 93.7 | 97.5 | 0.931 | 0.00066 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, C.; Thomopoulou, K.; Monastirioti, A.; Koronakis, G.; Papadaki, M.A.; Rounis, K.; Vamvakas, L.; Nikolaou, C.; Mavroudis, D.; Agelaki, S. MicroRNAs Regulating Tumor and Immune Cell Interactions in the Prediction of Relapse in Early Stage Breast Cancer. Biomedicines 2021, 9, 421. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040421
Papadaki C, Thomopoulou K, Monastirioti A, Koronakis G, Papadaki MA, Rounis K, Vamvakas L, Nikolaou C, Mavroudis D, Agelaki S. MicroRNAs Regulating Tumor and Immune Cell Interactions in the Prediction of Relapse in Early Stage Breast Cancer. Biomedicines. 2021; 9(4):421. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040421
Chicago/Turabian StylePapadaki, Chara, Konstantina Thomopoulou, Alexia Monastirioti, George Koronakis, Maria A. Papadaki, Konstantinos Rounis, Lambros Vamvakas, Christoforos Nikolaou, Dimitrios Mavroudis, and Sofia Agelaki. 2021. "MicroRNAs Regulating Tumor and Immune Cell Interactions in the Prediction of Relapse in Early Stage Breast Cancer" Biomedicines 9, no. 4: 421. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040421