
Targeting of the Essential acpP, ftsZ, and rne Genes in Carbapenem-Resistant Acinetobacter baumannii by Antisense PNA Precision Antibacterials


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Antibiotic MIC
2.3. Computational Screening of Sequence Regions in the mRNA of Target Genes
2.4. BPP-PNA Synthesis
2.5. BPP-PNA MIC
2.6. Time-Kill Assay
2.7. HepG2 Cell Toxicity
2.8. Envelope Disruption
2.9. Real-Time RT-PCR
2.10. Statistics
3. Results
3.1. PNA Design for the mRNA of Target Genes
3.1.1. AcpP
3.1.2. FtsZ
3.1.3. RNase E
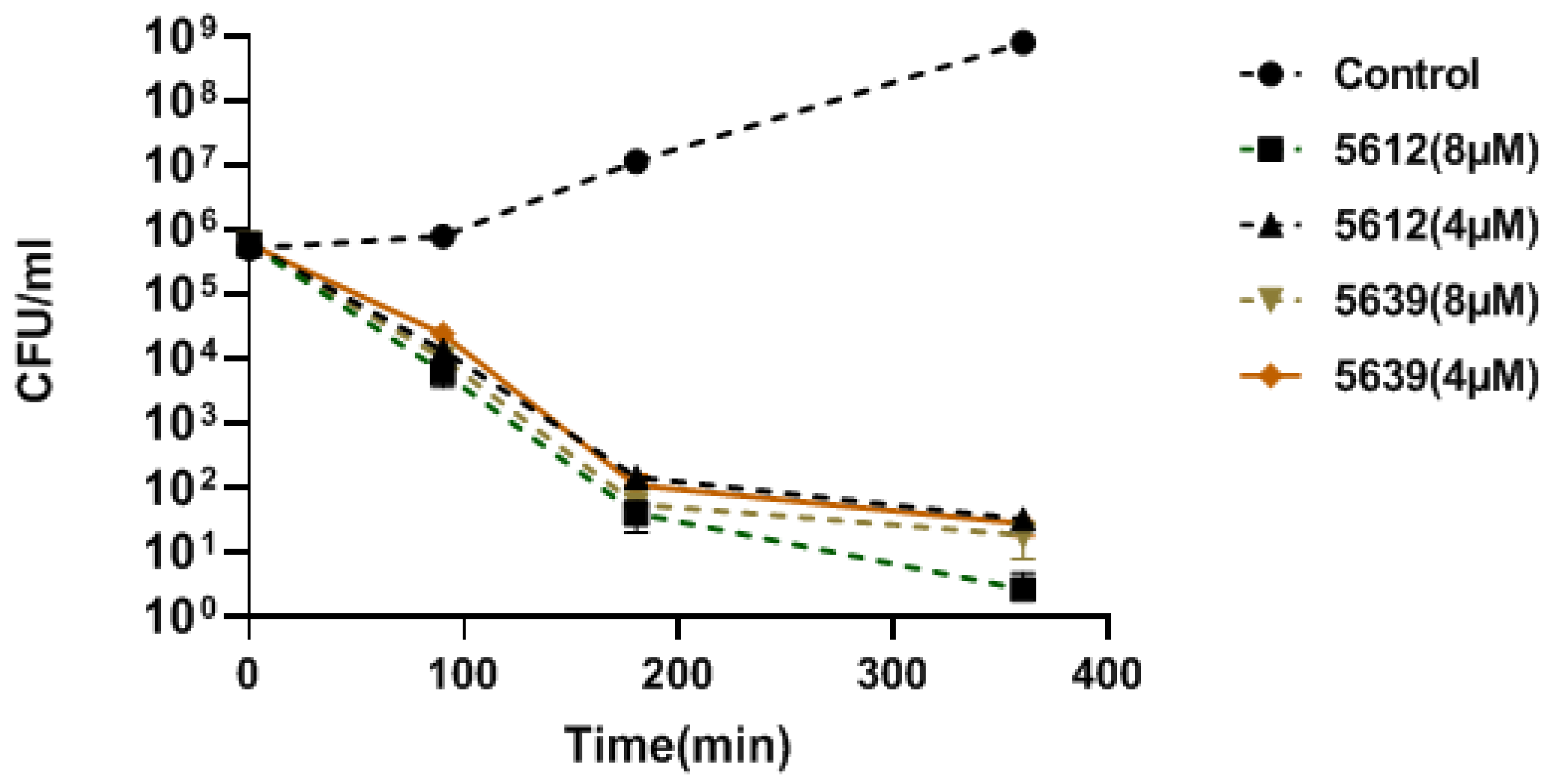
3.2. MIC and Time Kill Kinetics
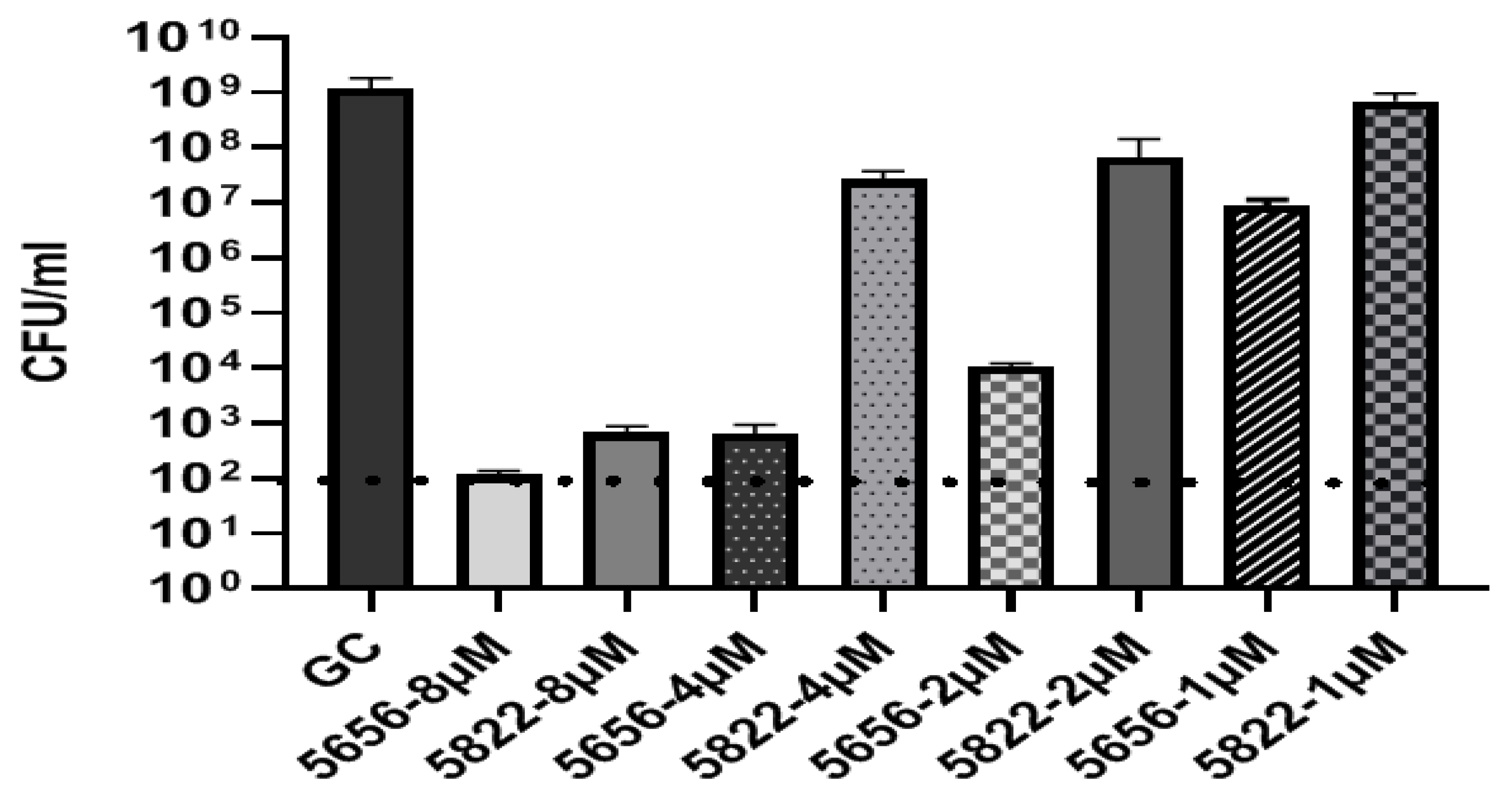
3.3. Species-Selective Activity of BPP-PNAs
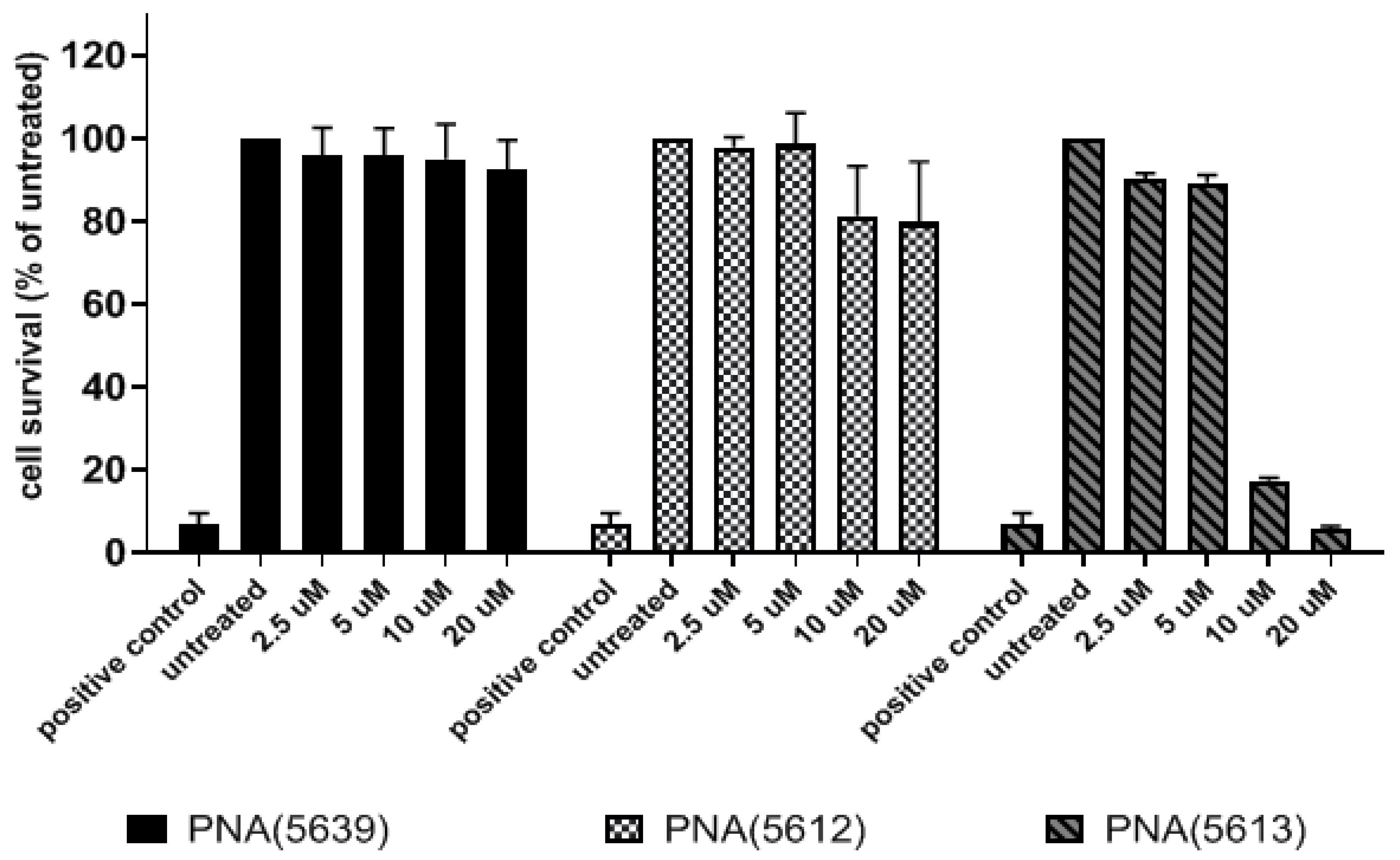
3.4. Cytotoxicity
3.5. Envelope Disruption
3.6. Effect on Target mRNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, L.-Y.; Apisarnthanarak, A.; Khan, E.; Suwantarat, N.; Ghafur, A.; Tambyah, P.A. Carbapenem-Resistant Acinetobacter baumannii and Enterobacteriaceae in South and Southeast Asia. Clin. Microbiol. Rev. 2016, 30, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and Pathophysiological Overview of Acinetobacter Infections: A Century of Challenges. Clin. Microbiol. Rev. 2016, 30, 409–447. [Google Scholar] [CrossRef] [Green Version]
- Isler, B.; Doi, Y.; Bonomo, R.A.; Paterson, D.L. New Treatment Options against Carbapenem-ResistantAcinetobacter baumanniiInfections. Antimicrob. Agents Chemother. 2018, 63. [Google Scholar] [CrossRef] [Green Version]
- Esterly, J.S.; Griffith, M.; Qi, C.; Malczynski, M.; Postelnick, M.J.; Scheetz, M.H. Impact of Carbapenem Resistance and Receipt of Active Antimicrobial Therapy on Clinical Outcomes of Acinetobacter baumannii Bloodstream Infections. Antimicrob. Agents Chemother. 2011, 55, 4844–4849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J. An RNA biology perspective on species-specific programmable RNA antibiotics. Mol. Microbiol. 2020, 113, 550–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowska, M.; Równicki, M.; Mieczkowski, A.; Miszkiewicz, J.; Trylska, J. Antibacterial Peptide Nucleic Acids—Facts and Perspectives. Molecules 2020, 25, 559. [Google Scholar] [CrossRef] [Green Version]
- Good, L.; Awasthi, S.K.; Dryselius, R.; Larsson, O.; Nielsen, P.E. Bactericidal antisense effects of peptide–PNA conjugates. Nat. Biotechnol. 2001, 19, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Geller, B.L. Antisense antimicrobial therapeutics. Curr. Opin. Microbiol. 2016, 33, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Good, L.; Stach, J.E.M. Synthetic RNA Silencing in Bacteria? Antimicrobial Discovery and Resistance Breaking. Front. Microbiol. 2011, 2, 185. [Google Scholar] [CrossRef] [Green Version]
- Demidov, V.V.; Potaman, V.N.; Frank-Kamenetskil, M.; Egholm, M.; Buchard, O.; Sönnichsen, S.H.; Nlelsen, P.E. Stability of peptide nucleic acids in human serum and cellular extracts. Biochem. Pharmacol. 1994, 48, 1310–1313. [Google Scholar] [CrossRef]
- Dryselius, R.; Aswasti, S.K.; Rajarao, G.K.; Nielsen, P.E.; Good, L. The Translation Start Codon Region Is Sensitive to Antisense PNA Inhibition in Escherichia coli. Oligonucleotides 2003, 13, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Japoni-Nejad, A.; Farshad, S.; Van Belkum, A.; Ghaznavi-Rad, E. Novel cassette array in a class 1 integron in clinical isolates of Acinetobacter baumannii from central Iran. Int. J. Med Microbiol. 2013, 303, 645–650. [Google Scholar] [CrossRef]
- Ellington, M.J.; Kistler, J.J.; Livermore, D.M.; Woodford, N. Multiplex PCR for rapid detection of genes encoding acquired metallo-β-lactamases. J. Antimicrob. Chemother. 2006, 59, 321–322. [Google Scholar] [CrossRef] [Green Version]
- Wayne, P.A.; CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 13th ed.; CLSI Standard M02; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Kent, W.J. BLAT---The BLAST-Like Alignment Tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goltermann, L.; Yavari, N.; Zhang, M.; Ghosal, A.; Nielsen, P.E. PNA Length Restriction of Antibacterial Activity of Peptide-PNA Conjugates in Escherichia coli Through Effects of the Inner Membrane. Front. Microbiol. 2019, 10, 1032. [Google Scholar] [CrossRef] [PubMed]
- Ghosal, A.; Nielsen, P.E. Potent Antibacterial Antisense Peptide–Peptide Nucleic Acid Conjugates Against Pseudomonas aeruginosa. Nucleic Acid Ther. 2012, 22, 323–334. [Google Scholar] [CrossRef] [Green Version]
- Roth, B.L.; Poot, M.; Yue, S.T.; Millard, P.J. Bacterial viability and antibiotic susceptibility testing with SYTOX green nucleic acid stain. Appl. Environ. Microbiol. 1997, 63, 2421–2431. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Hansen, A.M.; Bonke, G.; Larsen, C.J.; Yavari, N.; Nielsen, P.E.; Franzyk, H. Antibacterial Peptide Nucleic Acid–Antimicrobial Peptide (PNA–AMP) Conjugates: Antisense Targeting of Fatty Acid Biosynthesis. Bioconjugate Chem. 2016, 27, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Geller, B.L.; Marshall-Batty, K.; Schnell, F.J.; McKnight, M.M.; Iversen, P.L.; Greenberg, D.E. Gene-Silencing Antisense Oligomers Inhibit Acinetobacter Growth In Vitro and In Vivo. J. Infect. Dis. 2013, 208, 1553–1560. [Google Scholar] [CrossRef] [Green Version]
- Ghosal, A.; Vitali, A.; Stach, J.E.; Nielsen, P.E. Role of SbmA in the Uptake of Peptide Nucleic Acid (PNA)-Peptide Conjugates in E. coli. ACS Chem. Biol. 2012, 8, 360–367. [Google Scholar] [CrossRef]
- Mardle, C.E.; Shakespeare, T.J.; Butt, L.E.; Goddard, L.R.; Gowers, D.M.; Atkins, H.S.; Vincent, H.A.; Callaghan, A.J. A structural and biochemical comparison of Ribonuclease E homologues from pathogenic bacteria highlights species-specific properties. Sci. Rep. 2019, 9, 7952. [Google Scholar] [CrossRef] [Green Version]
- Eidem, T.M.; Roux, C.M.; Dunman, P.M. RNA decay: A novel therapeutic target in bacteria. Wiley Interdiscip. Rev. RNA 2012, 3, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Splith, K.; Neundorf, I. Antimicrobial peptides with cell-penetrating peptide properties and vice versa. Eur. Biophys. J. 2011, 40, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Guitián, M.; Vázquez-Ucha, J.C.; Álvarez-Fraga, L.; Conde-Pérez, K.; Bou, G.; Poza, M.; Beceiro, A. Antisense inhibition of lpxB gene expression in Acinetobacter baumannii by peptide–PNA conjugates and synergy with colistin. J. Antimicrob. Chemother. 2020, 75, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, Y.; Xia, Y.; Wang, L.; Liang, S. Inhibition of gene expression and growth of multidrug-resistant Acinetobacter baumannii by antisense peptide nucleic acids. Mol. Biol. Rep. 2014, 41, 7535–7541. [Google Scholar] [CrossRef]
- Rose, M.; Lapuebla, A.; Landman, D.; Quale, J. In Vitro and In Vivo Activity of a Novel Antisense Peptide Nucleic Acid Compound Against Multidrug-Resistant Acinetobacter baumannii. Microb. Drug Resist. 2019, 25, 961–965. [Google Scholar] [CrossRef]
- Carpousis, A.J. The RNA Degradosome of Escherichia coli: An mRNA-Degrading Machine Assembled on RNase E. Annu. Rev. Microbiol. 2007, 61, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, D.E.; Marshall-Batty, K.R.; Brinster, L.R.; Zarember, K.A.; Shaw, P.A.; Mellbye, B.L.; Iversen, P.L.; Holland, S.M.; Geller, B.L. Antisense Phosphorodiamidate Morpholino Oligomers Targeted to an Essential Gene Inhibit Burkholderia cepacia Complex. J. Infect. Dis. 2010, 201, 1822–1830. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.J.; Sturge, C.R.; Moustafa, D.A.; Daly, S.M.; Marshall-Batty, K.R.; Felder, C.F.; Zamora, D.; Yabe-Gill, M.; Labandeira-Rey, M.; Bailey, S.M.; et al. Inhibition of Pseudomonas aeruginosa by Peptide-Conjugated Phosphorodiamidate Morpholino Oligomers. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkowsky, G.; Lemster, A.-L.; Pappesch, R.; Jacob, A.; Krüger, S.; Schröder, A.; Kreikemeyer, B.; Patenge, N. Influence of Different Cell-Penetrating Peptides on the Antimicrobial Efficiency of PNAs in Streptococcus pyogenes. Mol. Ther. Nucleic Acids 2019, 18, 444–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yavari, N.; Goltermann, L.; Nielsen, P.E. Uptake, Stability, and Activity of Antisense Anti-acpP PNA-Peptide Conjugates in Escherichia coli and the Role of SbmA. ACS Chem. Biol. 2021, 16, 471–479. [Google Scholar] [CrossRef] [PubMed]









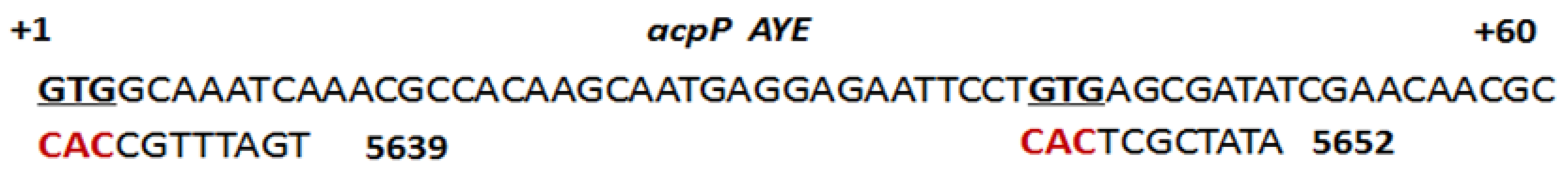{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PNA No. | Sequence N→C | Target Position | Peptide | MIC (µM) | ||||
|---|---|---|---|---|---|---|---|---|
| ATCC 19606 | AC44 | AC46 | SSI 1104 | AC2 | ||||
| 5652 (m) * | ATATCGCTCAC | +40 to +50 b | H-(KFF)3K-eg1 | 4 | 4 | 4 | 4 | 4 |
| 5653 (mm) * | ATACCGCTTAC | H-(KFF)3K-eg1 | 4 | 4 | 4 | 4 | 4 | |
| 5639 (m) * | TGATTTGCCAC | +1 to +11 a | H-(KFF)3K-eg1 | 2 | 1 | 1 | 2 | 1–2 |
| 5640 (mm) * | TGACTTGTCAC | H-(KFF)3K-eg1 | 4 | 4 | 4 | 4 | 4 | |
| 5639 (m) ** | TGATTTGCCAC | +1 to +11 a | H-(KFF)3K-eg1 | 4 | 4 | 4 | 4 | 4 |
| 5640 (mm) ** | TGACTTGTCAC | H-(KFF)3K-eg1 | 4 | 4 | 4 | 4 | 4 | |
| 5823 (m) * | TGATTTGCCAC | +1 to +11 a | H-(kff)3k-eg1 c | 4 | 4 | 4 | 4 | 4 |
| 5824 (mm) * | TGACTTGTCAC | H-(kff)3k-eg1 c | 8 | 4 | 4 | 8 | 8 |
| PNA No | Sequence N→C | Target Position | Peptide | MIC (µM) | ||
|---|---|---|---|---|---|---|
| ATCC 19606 | AC44 | SSI1104 | ||||
| 5612 (m) | GAGGCCATGAC | −3 to +8 | H-(KFF)3K-eg1 | 4 | 8 | 4 |
| 5615 (mm) | GAGTCCAGGAC | H-(KFF)3K-eg1 | 8 | 16 | 16 | |
| 5613 (m) | GAGGCCATGAC | −3 to +8 | H-(R-Ahx-R)4-Ahx | >16 | >16 | nd |
| 5616 (mm) | GAGTCCAGGAC | H-(R-Ahx-R)4-Ahx | >16 | >16 | nd | |
| 5657 (m) | ATGAGGCCATG | −1 to +10 | H-(KFF)3K-eg1 | >16 | 16 | nd |
| 5658 (m) | GGCCATGACCT | −5 to +6 | H-(KFF)3K-eg1 | 4 | 8 | 8 |
| Strains | H-(KFF)3K-eg1-ftsZ PNA(5612) | H-(KFF)3K-eg1-ftsZ PNA(5615) |
|---|---|---|
| ATCC19606 | 4 µM | 8 µM |
| AC1 | 4 µM | 16 µM |
| AC2 | 2 µM | 16 µM |
| AC3 | 4 µM | 8 µM |
| AC4 | 8 µM | 16 µM |
| AC5 | 4 µM | 16 µM |
| AC6 | 4 µM | 8 µM |
| AC7 | 4 µM | 16 µM |
| AC8 | 8 µM | 16 µM |
| AC9 | 2 µM | 16 µM |
| AC44 | 8 µM | 16 µM |
| AC46 | 8 µM | 16 µM |
| SSI1104 | 4 µM | 16 µM |
| MIC50 | 4 µM | 16 µM |
| PNA No | Sequence N→C | Target Position | Peptide | MIC (µM) | ||||
|---|---|---|---|---|---|---|---|---|
| ATCC 19606 | AC44 | AC46 | SSI 1104 | AC2 | ||||
| 5637 (m) | ACGTTTCATGG | −2 to +9 | H-(KFF)3K-eg1 | 1 | 1 | 1 | nd | nd |
| 5638 (mm) | ACGATTCTTGG | H-(KFF)3K-eg1 | 2 | 2 | 1 | nd | nd | |
| 5654 (m) | ATACGTTTCAT | +1 to +11 | H-(KFF)3K-eg1 | 2 | 1 | 2 | 2 | 2 |
| 5821 (mm) | ATCCGTTTAAT | H-(KFF)3K-eg1 | 4 | 1 | 2 | 2 | 2 | |
| 5656 (m) | TTTCATGGGTG | −5 to +6 | H-(KFF)3K-eg1 | 2 | 2 | 2 | 2 | 2 |
| 5822 (mm) | TTTGATGTGTG | H-(KFF)3K-eg1 | 8 | 8 | 8 | 8 | 8 | |
| 5655 (m) | GTTTCATGGGT | −4 to +7 | H-(KFF)3K-eg1 | 4 | 8 | nd | nd | 8 |
| Target | PNA | Sequence * | MIC (µM) | MIC (µM) | MIC (µM) | MIC (µM) |
|---|---|---|---|---|---|---|
| P. aeruginosa (PAO1) | E. coli K-12 MG1655 | A. baumannii ATCC 19606 | A. baumannii SSI1104 | |||
| ftsZ (m) | 5612 | GAGGCCATGAC | >16 µM | >16 µM | 4 µM | 4 µM |
| acpP (m) | 5639 | TGATTTGCCAC | >16 µM | >16 µM | 2 µM | 2 µM |
| acpP (m) | 5652 | ATATCGCTCAC | 16 µM | 16 µM | 4 µM | 4 µM |
| ftsZ (mm) # | 3965 | TTCTAACAAAGT | nd | 16 µM | 16 µM | ≥16 µM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nejad, A.J.; Shahrokhi, N.; Nielsen, P.E. Targeting of the Essential acpP, ftsZ, and rne Genes in Carbapenem-Resistant Acinetobacter baumannii by Antisense PNA Precision Antibacterials. Biomedicines 2021, 9, 429. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040429
Nejad AJ, Shahrokhi N, Nielsen PE. Targeting of the Essential acpP, ftsZ, and rne Genes in Carbapenem-Resistant Acinetobacter baumannii by Antisense PNA Precision Antibacterials. Biomedicines. 2021; 9(4):429. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040429
Chicago/Turabian StyleNejad, Alireza Japoni, Nader Shahrokhi, and Peter E. Nielsen. 2021. "Targeting of the Essential acpP, ftsZ, and rne Genes in Carbapenem-Resistant Acinetobacter baumannii by Antisense PNA Precision Antibacterials" Biomedicines 9, no. 4: 429. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9040429