
Degeneration of Aortic Valves in a Bioreactor System with Pulsatile Flow

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
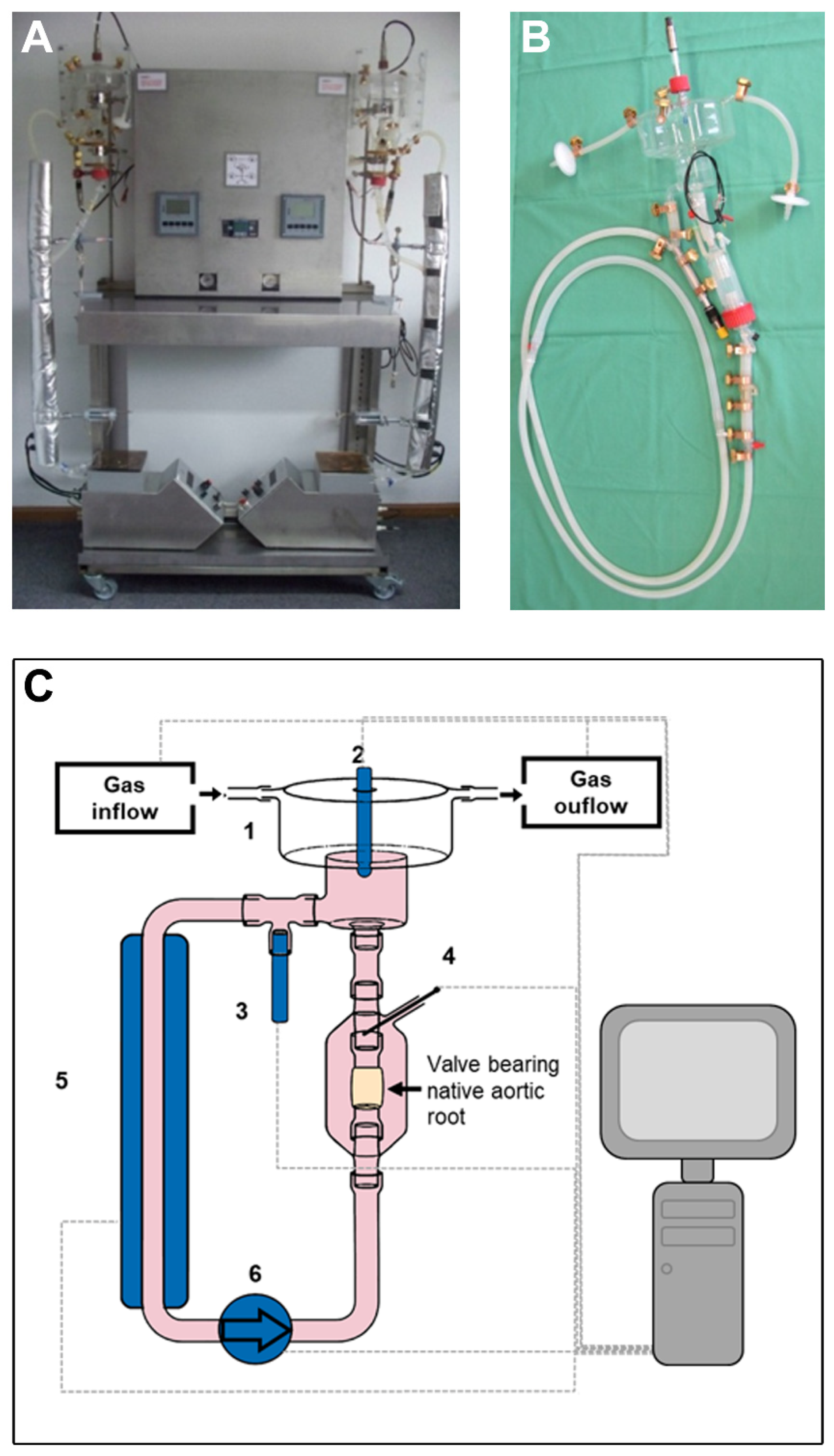
2.1. Bioreactor Design
2.2. Cultivation of Valve Bearing Native Aortic Roots
2.3. Histological Staining and Immunohistochemistry
2.4. RNA Isolation and Semi-Quantitative Real-Time PCR (qRT-PCR)
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
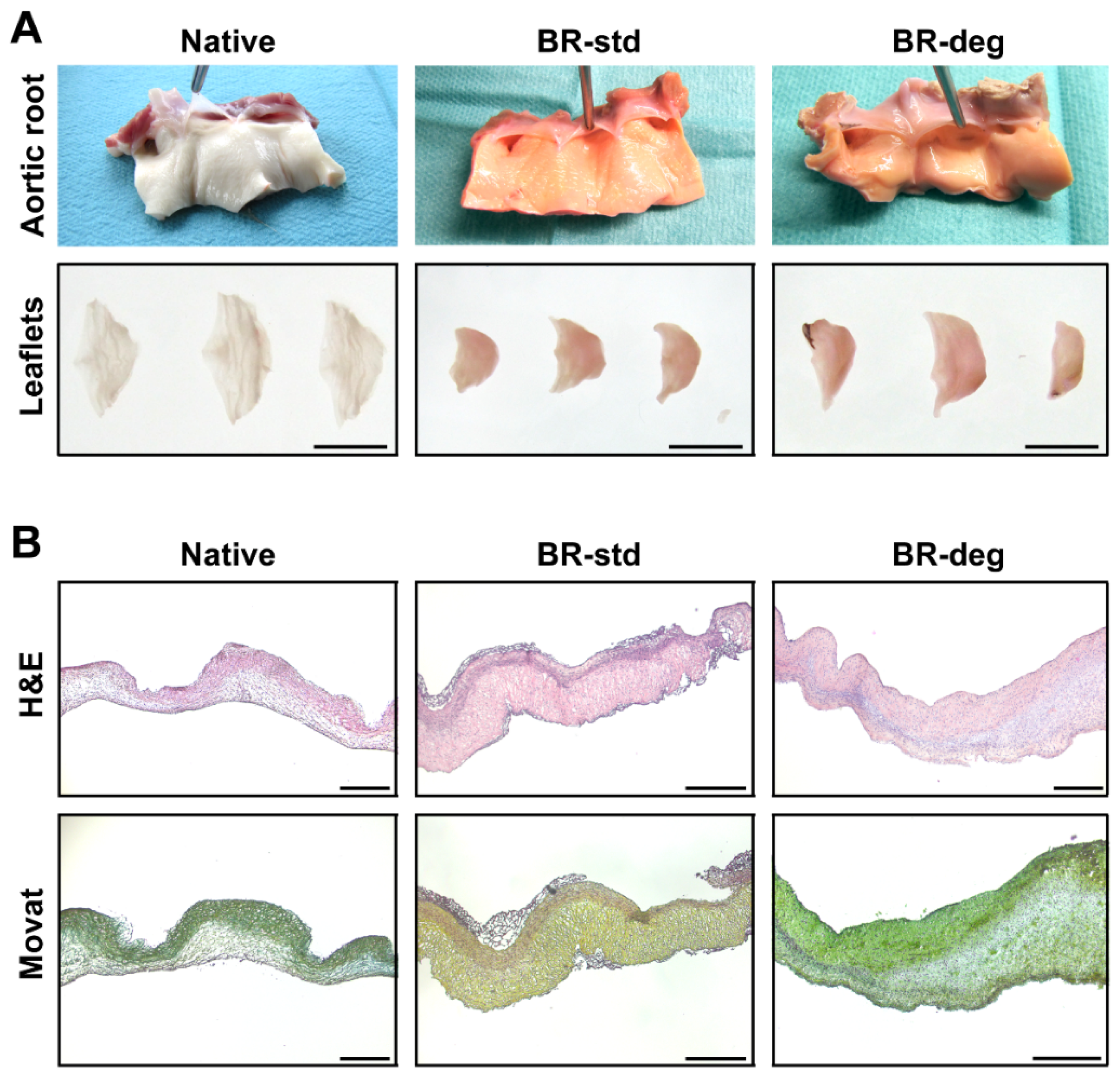
3.1. Bioreactor Function and Macroscopic Analysis
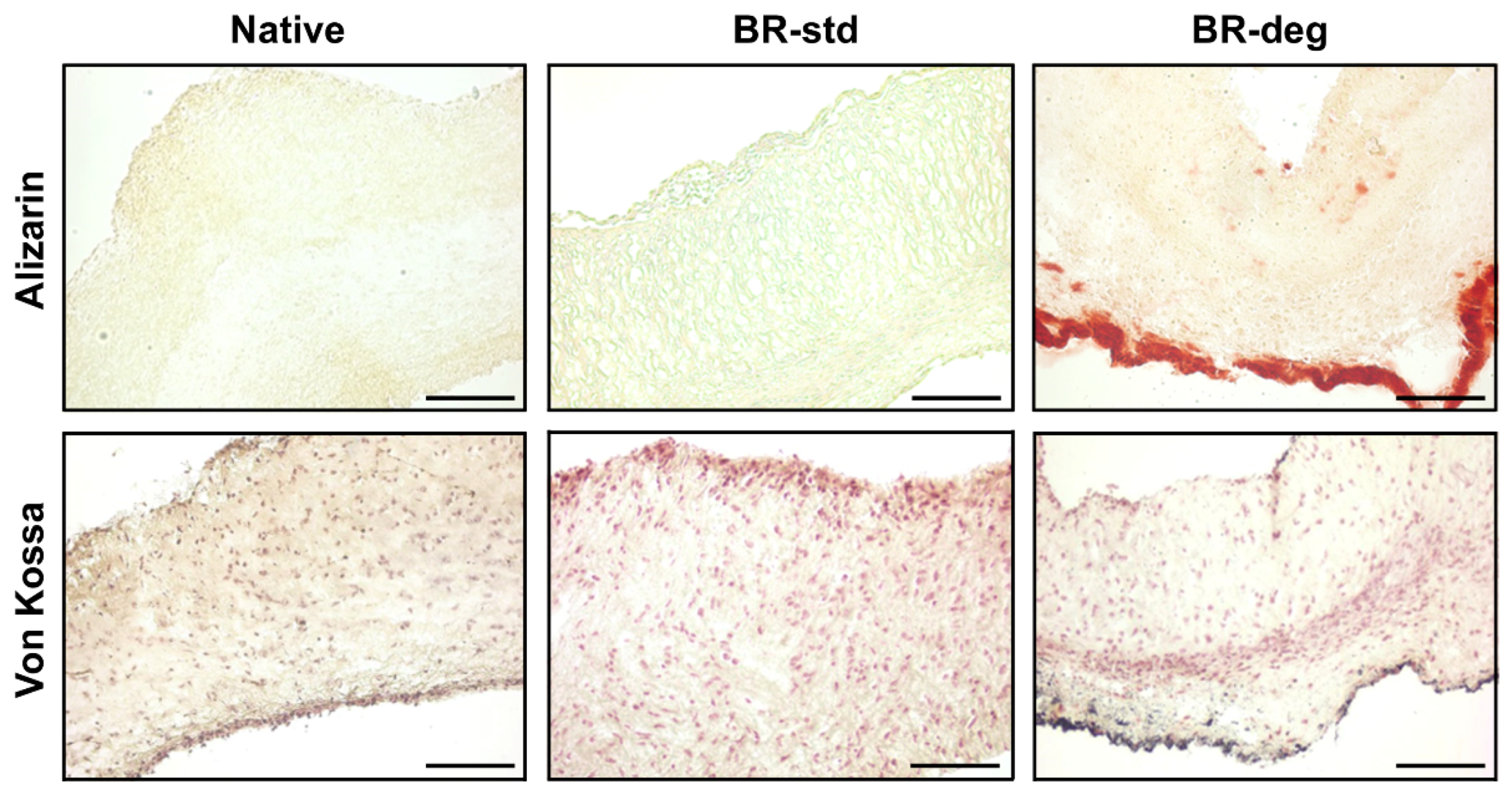
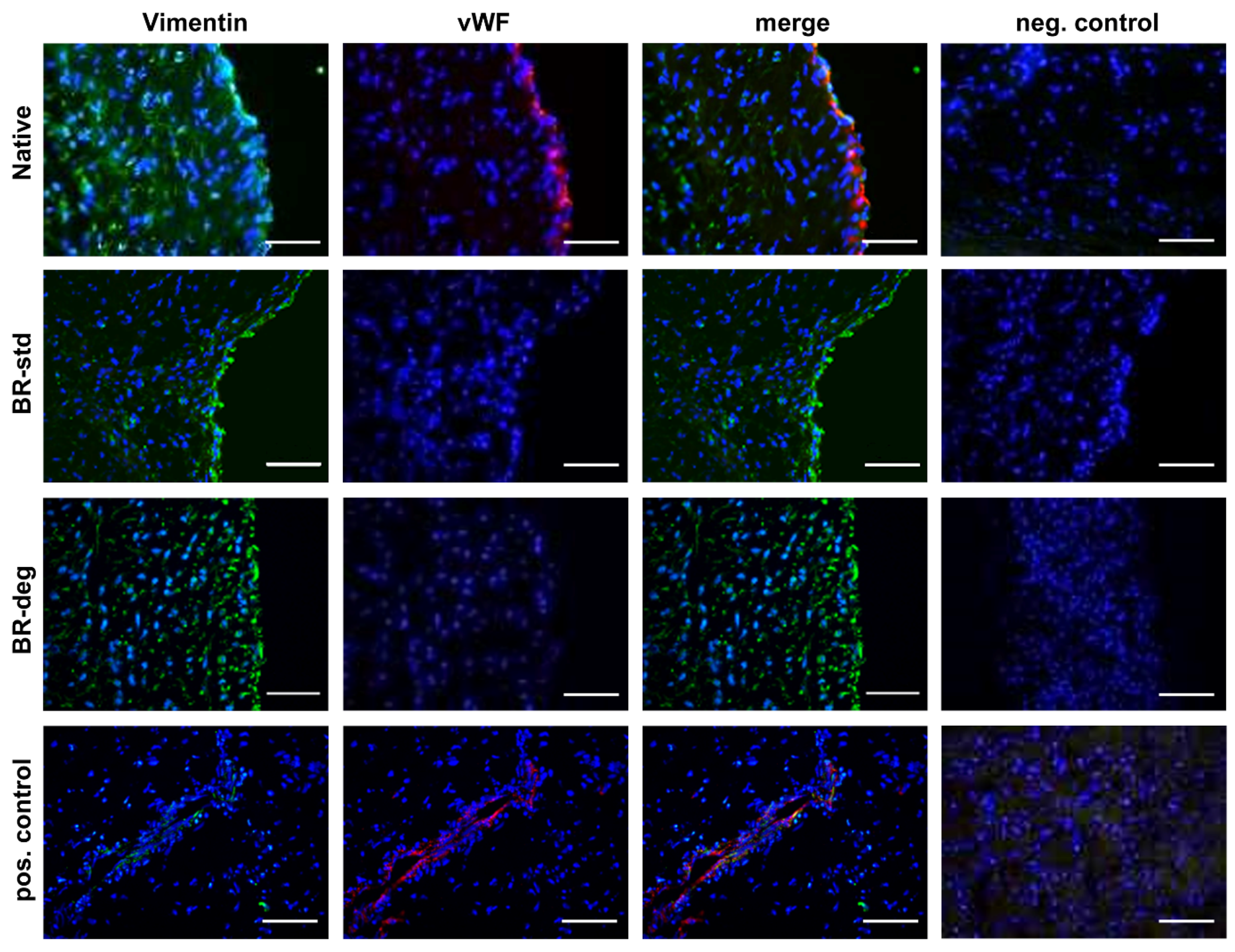
3.2. Analysis of Tissue Biomineralization and Cell Composition
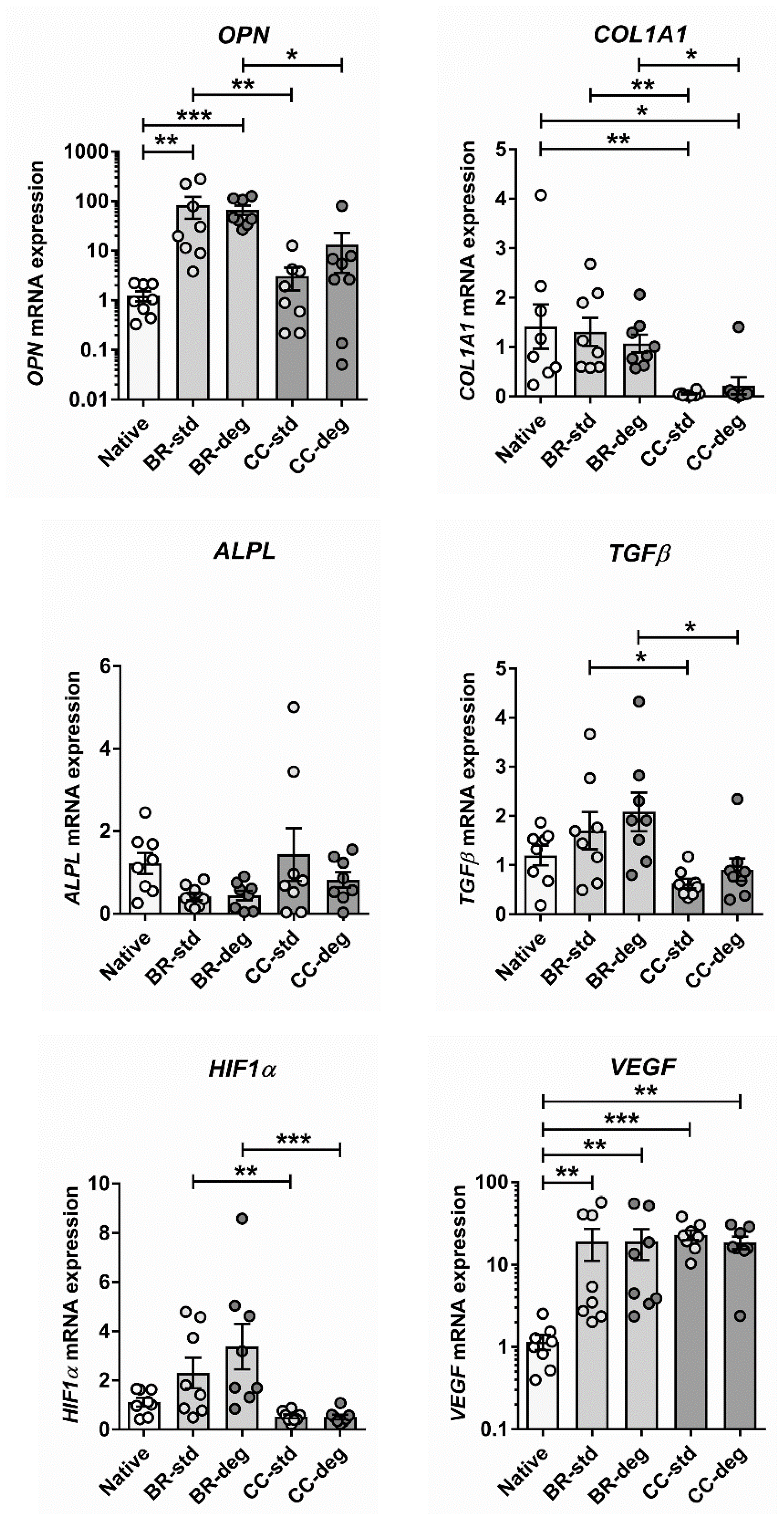
3.3. Gene Expression of Pro-Degenerative Markers
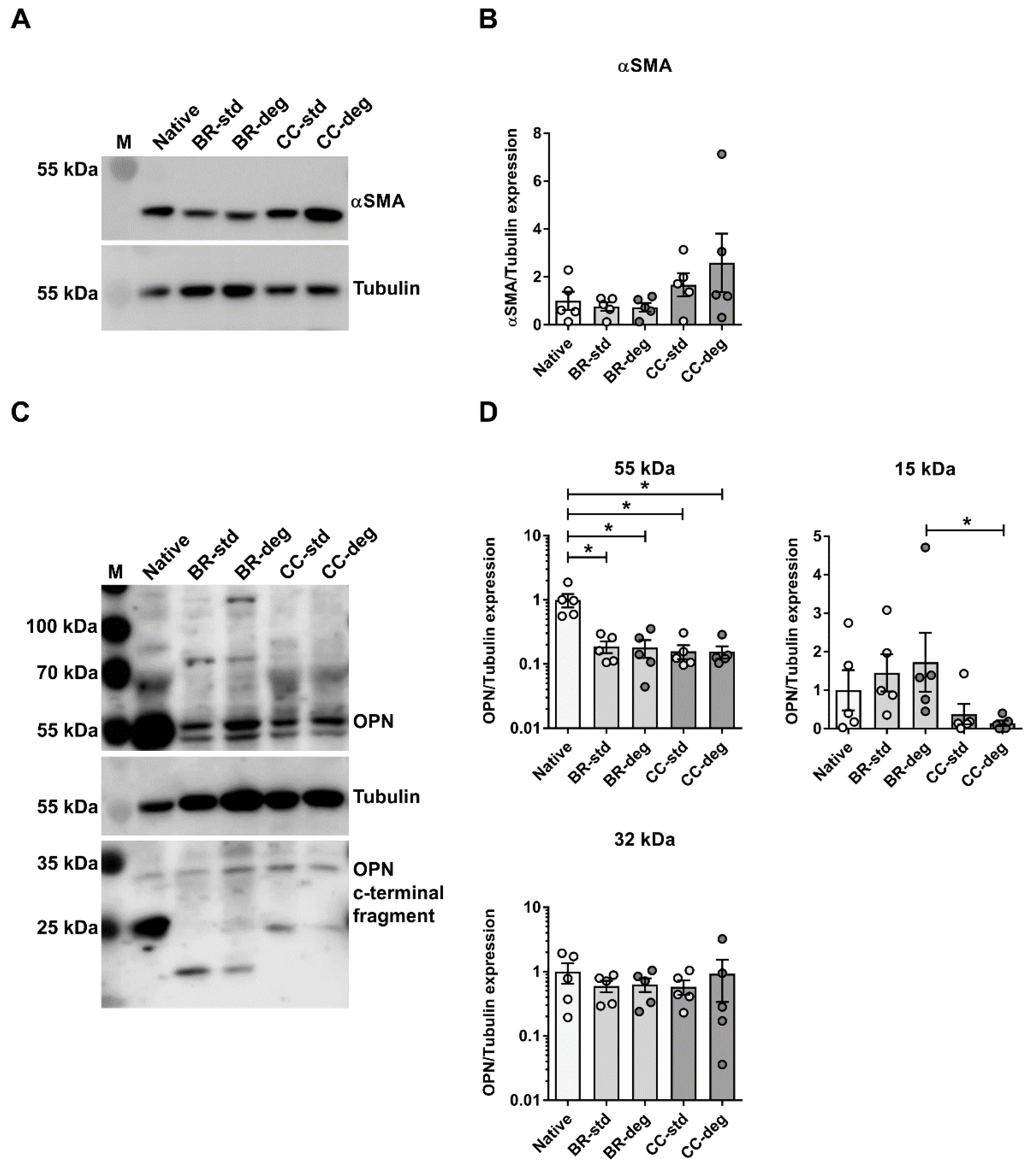
3.4. Bioreactor Mediated Changes on the Protein Level
4. Discussion
4.1. Induction of Biomineralization Due to Bioreactor Cultivation
4.2. Bioreactor Cultivation Leads to Altered Gene Expression Patterns
4.3. Altered Expression of the Osteogenic Marker OPN
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindman, B.R.; Clavel, M.-A.; Mathieu, P.; Iung, B.; Lancellotti, P.; Otto, C.M.; Pibarot, P. Calcific aortic stenosis. Nat. Rev. Dis. Primers 2016, 2, 16006. [Google Scholar] [CrossRef] [Green Version]
- Beckmann, A.; Funkat, A.-K.; Lewandowski, J.; Frie, M.; Ernst, M.; Hekmat, K.; Schiller, W.; Gummert, J.F.; Harringer, W. German Heart Surgery Report 2016: The Annual Updated Registry of the German Society for Thoracic and Cardiovascular Surgery. Thorac. Cardiovasc. Surg. 2017, 65, 505–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, R.V.; Otto, C.M. Spectrum of calcific aortic valve disease: Pathogenesis, disease progression, and treatment strategies. Circulation 2005, 111, 3316–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latif, N.; Sarathchandra, P.; Taylor, P.M.; Antoniw, J.; Yacoub, M.H. Localization and pattern of expression of extracellular matrix components in human heart valves. J. Heart Valve Dis. 2005, 14, 218–227. [Google Scholar] [PubMed]
- Liu, A.C.; Joag, V.R.; Gotlieb, A.I. The emerging role of valve interstitial cell phenotypes in regulating heart valve pathobiology. Am. J. Pathol. 2007, 171, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Gwanmesia, P.; Ziegler, H.; Eurich, R.; Barth, M.; Kamiya, H.; Karck, M.; Lichtenberg, A.; Akhyari, P. Opposite effects of transforming growth factor-beta1 and vascular endothelial growth factor on the degeneration of aortic valvular interstitial cell are modified by the extracellular matrix protein fibronectin: Implications for heart valve engineering. Tissue Eng. Part A 2010, 16, 3737–3746. [Google Scholar] [CrossRef]
- Hortells, L.; Sur, S.; St Hilaire, C. Cell Phenotype Transitions in Cardiovascular Calcification. Front. Cardiovasc. Med. 2018, 5, 27. [Google Scholar] [CrossRef]
- Hasan, A.; Ragaert, K.; Swieszkowski, W.; Selimović, S.; Paul, A.; Camci-Unal, G.; Mofrad, M.R.K.; Khademhosseini, A. Biomechanical properties of native and tissue engineered heart valve constructs. J. Biomech. 2014, 47, 1949–1963. [Google Scholar] [CrossRef]
- Wyss, K.; Yip, C.Y.Y.; Mirzaei, Z.; Jin, X.; Chen, J.-H.; Simmons, C.A. The elastic properties of valve interstitial cells undergoing pathological differentiation. J. Biomech. 2012, 45, 882–887. [Google Scholar] [CrossRef]
- Fisher, C.I.; Chen, J.; Merryman, W.D. Calcific nodule morphogenesis by heart valve interstitial cells is strain dependent. Biomech. Model. Mechanobiol. 2013, 12, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Farrar, E.J.; Pramil, V.; Richards, J.M.; Mosher, C.Z.; Butcher, J.T. Valve interstitial cell tensional homeostasis directs calcification and extracellular matrix remodeling processes via RhoA signaling. Biomaterials 2016, 105, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balachandran, K.; Alford, P.W.; Wylie-Sears, J.; Goss, J.A.; Grosberg, A.; Bischoff, J.; Aikawa, E.; Levine, R.A.; Parker, K.K. Cyclic strain induces dual-mode endothelial-mesenchymal transformation of the cardiac valve. Proc. Natl. Acad. Sci. USA 2011, 108, 19943–19948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, K.; Ma, X.; Chalajour, F.; Hanley, F.L.; Riemer, R.K. Critical Role of Coaptive Strain in Aortic Valve Leaflet Homeostasis: Use of a Novel Flow Culture Bioreactor to Explore Heart Valve Mechanobiology. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Rajamannan, N.M.; Sucosky, P. Defining the role of fluid shear stress in the expression of early signaling markers for calcific aortic valve disease. PLoS ONE 2013, 8, e84433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Masjedi, S.; Ferdous, Z. A study of extracellular matrix remodeling in aortic heart valves using a novel biaxial stretch bioreactor. J. Mech. Behav. Biomed. Mater. 2017, 75, 351–358. [Google Scholar] [CrossRef]
- Weiler, M.; Yap, C.H.; Balachandran, K.; Padala, M.; Yoganathan, A.P. Regional analysis of dynamic deformation characteristics of native aortic valve leaflets. J. Biomech. 2011, 44, 1459–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amadeo, F.; Boschetti, F.; Polvani, G.; Banfi, C.; Pesce, M.; Santoro, R. Aortic valve cell seeding into decellularized animal pericardium by perfusion-assisted bioreactor. J. Tissue Eng. Regen. Med. 2018, 12, 1481–1493. [Google Scholar] [CrossRef]
- Converse, G.L.; Buse, E.E.; Hopkins, R.A. Bioreactors and operating room centric protocols for clinical heart valve tissue engineering. Prog. Pediatric Cardiol. 2013, 35, 95–100. [Google Scholar] [CrossRef]
- Lichtenberg, A.; Tudorache, I.; Cebotari, S.; Ringes-Lichtenberg, S.; Sturz, G.; Hoeffler, K.; Hurscheler, C.; Brandes, G.; Hilfiker, A.; Haverich, A. In vitro re-endothelialization of detergent decellularized heart valves under simulated physiological dynamic conditions. Biomaterials 2006, 27, 4221–4229. [Google Scholar] [CrossRef] [Green Version]
- Lichtenberg, A.; Cebotari, S.; Tudorache, I.; Sturz, G.; Winterhalter, M.; Hilfiker, A.; Haverich, A. Flow-dependent re-endothelialization of tissue-engineered heart valves. J. Heart Valve Dis. 2006, 15, 287–293. [Google Scholar]
- Weber, A.; Barth, M.; Selig, J.I.; Raschke, S.; Dakaras, K.; Hof, A.; Hesse, J.; Schrader, J.; Lichtenberg, A.; Akhyari, P. Enzymes of the purinergic signaling system exhibit diverse effects on the degeneration of valvular interstitial cells in a 3-D microenvironment. FASEB J. 2018, 32, 4356–4369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assmann, A.; Zwirnmann, K.; Heidelberg, F.; Schiffer, F.; Horstkötter, K.; Munakata, H.; Gremse, F.; Barth, M.; Lichtenberg, A.; Akhyari, P. The degeneration of biological cardiovascular prostheses under pro-calcific metabolic conditions in a small animal model. Biomaterials 2014, 35, 7416–7428. [Google Scholar] [CrossRef]
- Balachandran, K.; Sucosky, P.; Jo, H.; Yoganathan, A.P. Elevated cyclic stretch induces aortic valve calcification in a bone morphogenic protein-dependent manner. Am. J. Pathol. 2010, 177, 49–57. [Google Scholar] [CrossRef]
- Clark-Greuel, J.N.; Connolly, J.M.; Sorichillo, E.; Narula, N.R.; Rapoport, H.S.; Mohler, E.R., III; Gorman, J.H., III; Gorman, R.C.; Levy, R.J. Transforming growth factor-beta1 mechanisms in aortic valve calcification: Increased alkaline phosphatase and related events. Ann. Thorac. Surg. 2007, 83, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Cloyd, K.L.; El-Hamamsy, I.; Boonrungsiman, S.; Hedegaard, M.; Gentleman, E.; Sarathchandra, P.; Colazzo, F.; Gentleman, M.M.; Yacoub, M.H.; Chester, A.H.; et al. Characterization of porcine aortic valvular interstitial cell ‘calcified’ nodules. PLoS ONE 2012, 7, e48154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, G.A.; Masters, K.S.; Shah, D.N.; Anseth, K.S.; Leinwand, L.A. Valvular myofibroblast activation by transforming growth factor-beta: Implications for pathological extracellular matrix remodeling in heart valve disease. Circ. Res. 2004, 95, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Benton, J.A.; Kern, H.B.; Leinwand, L.A.; Mariner, P.D.; Anseth, K.S. Statins block calcific nodule formation of valvular interstitial cells by inhibiting alpha-smooth muscle actin expression. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1950–1957. [Google Scholar] [CrossRef] [Green Version]
- Jian, B.; Narula, N.; Li, Q.; Mohler, E.R.; Levy, R.J. Progression of aortic valve stenosis: TGF-β1 is present in calcified aortic valve cusps and promotes aortic valve interstitial cell calcification via apoptosis. Ann. Thorac. Surg. 2003, 75, 457–465. [Google Scholar] [CrossRef]
- Kennedy, J.A.; Hua, X.; Mishra, K.; Murphy, G.A.; Rosenkranz, A.C.; Horowitz, J.D. Inhibition of calcifying nodule formation in cultured porcine aortic valve cells by nitric oxide donors. Eur. J. Pharmacol. 2009, 602, 28–35. [Google Scholar] [CrossRef]
- Sucosky, P.; Balachandran, K.; Elhammali, A.; Jo, H.; Yoganathan, A.P. Altered shear stress stimulates upregulation of endothelial VCAM-1 and ICAM-1 in a BMP-4- and TGF-beta1-dependent pathway. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Sewell-Loftin, M.-K.; Brown, C.B.; Baldwin, H.S.; Merryman, W.D. A novel technique for quantifying mouse heart valve leaflet stiffness with atomic force microscopy. J. Heart Valve Dis. 2012, 21, 513–520. [Google Scholar] [PubMed]
- Merryman, W.D.; Huang, H.-Y.S.; Schoen, F.J.; Sacks, M.S. The effects of cellular contraction on aortic valve leaflet flexural stiffness. J. Biomech. 2006, 39, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Haskett, D.; Johnson, G.; Zhou, A.; Utzinger, U.; Vande Geest, J. Microstructural and biomechanical alterations of the human aorta as a function of age and location. Biomech. Model. Mechanobiol. 2010, 9, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Sucosky, P. Computational comparison of regional stress and deformation characteristics in tricuspid and bicuspid aortic valve leaflets. Int. J. Numer. Method Biomed. Eng. 2017, 33. [Google Scholar] [CrossRef]
- Otto, C.M.; Kuusisto, J.; Reichenbach, D.D.; Gown, A.M.; O’Brien, K.D. Characterization of the early lesion of ‘degenerative’ valvular aortic stenosis. Histological and immunohistochemical studies. Circulation 1994, 90, 844–853. [Google Scholar] [CrossRef] [Green Version]
- Sapp, M.C.; Krishnamurthy, V.K.; Puperi, D.S.; Bhatnagar, S.; Fatora, G.; Mutyala, N.; Grande-Allen, K.J. Differential cell-matrix responses in hypoxia-stimulated aortic versus mitral valves. J. R. Soc. Interface 2016, 13. [Google Scholar] [CrossRef] [Green Version]
- Perrotta, I.; Moraca, F.M.; Sciangula, A.; Aquila, S.; Mazzulla, S. HIF-1alpha and VEGF: Immunohistochemical Profile and Possible Function in Human Aortic Valve Stenosis. Ultrastruct. Pathol. 2015, 39, 198–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.S.; Kiriakidis, S.; Sandison, A.; Paleolog, E.M.; Davies, A.H. Hypoxia-inducible factor pathway and diseases of the vascular wall. J. Vasc. Surg. 2013, 58, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Masson, N.; Willam, C.; Maxwell, P.H.; Pugh, C.W.; Ratcliffe, P.J. Independent function of two destruction domains in hypoxia-inducible factor-alpha chains activated by prolyl hydroxylation. EMBO J. 2001, 20, 5197–5206. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.C.; Gotlieb, A.I. Transforming growth factor-beta regulates in vitro heart valve repair by activated valve interstitial cells. Am. J. Pathol. 2008, 173, 1275–1285. [Google Scholar] [CrossRef] [Green Version]
- Rajamannan, N.M.; Subramaniam, M.; Rickard, D.; Stock, S.R.; Donovan, J.; Springett, M.; Orszulak, T.; Fullerton, D.A.; Tajik, A.J.; Bonow, R.O.; et al. Human aortic valve calcification is associated with an osteoblast phenotype. Circulation 2003, 107, 2181–2184. [Google Scholar] [CrossRef] [PubMed]
- Rutkovskiy, A.; Malashicheva, A.; Sullivan, G.; Bogdanova, M.; Kostareva, A.; Stenslokken, K.-O.; Fiane, A.; Vaage, J. Valve Interstitial Cells: The Key to Understanding the Pathophysiology of Heart Valve Calcification. J. Am. Heart Assoc. 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Rattazzi, M.; Iop, L.; Faggin, E.; Bertacco, E.; Zoppellaro, G.; Baesso, I.; Puato, M.; Torregrossa, G.; Fadini, G.P.; Agostini, C.; et al. Clones of interstitial cells from bovine aortic valve exhibit different calcifying potential when exposed to endotoxin and phosphate. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 2165–2172. [Google Scholar] [CrossRef]
- Balachandran, K.; Konduri, S.; Sucosky, P.; Jo, H.; Yoganathan, A.P. An ex vivo study of the biological properties of porcine aortic valves in response to circumferential cyclic stretch. Ann. Biomed. Eng. 2006, 34, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Warnock, J.N.; He, Z.; Hilbert, S.L.; Yoganathan, A.P. Cyclic pressure affects the biological properties of porcine aortic valve leaflets in a magnitude and frequency dependent manner. Ann. Biomed. Eng. 2004, 32, 1461–1470. [Google Scholar] [CrossRef]
- Thayer, P.; Balachandran, K.; Rathan, S.; Yap, C.H.; Arjunon, S.; Jo, H.; Yoganathan, A.P. The Effects of Combined Cyclic Stretch and Pressure on the Aortic Valve Interstitial Cell Phenotype. Ann. Biomed. Eng. 2011, 39, 1654–1667. [Google Scholar] [CrossRef] [Green Version]
- Orimo, H. The mechanism of mineralization and the role of alkaline phosphatase in health and disease. J. Nippon Med. Sch. 2010, 77, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Gould, R.A.; Chin, K.; Santisakultarm, T.P.; Dropkin, A.; Richards, J.M.; Schaffer, C.B.; Butcher, J.T. Cyclic strain anisotropy regulates valvular interstitial cell phenotype and tissue remodeling in three-dimensional culture. Acta Biomater. 2012, 8, 1710–1719. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Nielsen, M.S.; Haselmann, K.F.; Petersen, T.E.; Sørensen, E.S. Post-translationally modified residues of native human osteopontin are located in clusters: Identification of 36 phosphorylation and five O-glycosylation sites and their biological implications. Biochem. J. 2005, 390, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainger, R.; Grau, J.B.; Poggio, P.; Branchetti, E.; Bavaria, J.E.; Gorman, J.H., 3rd; Gorman, R.C.; Ferrari, G. Dephosphorylation of circulating human osteopontin correlates with severe valvular calcification in patients with calcific aortic valve disease. Biomarkers 2012, 17, 111–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jono, S.; Peinado, C.; Giachelli, C.M. Phosphorylation of osteopontin is required for inhibition of vascular smooth muscle cell calcification. J. Biol. Chem. 2000, 275, 20197–20203. [Google Scholar] [CrossRef] [Green Version]
- Higashikawa, F.; Eboshida, A.; Yokosaki, Y. Enhanced biological activity of polymeric osteopontin. FEBS Lett. 2007, 581, 2697–2701. [Google Scholar] [CrossRef] [Green Version]
- Nishimichi, N.; Hayashita-Kinoh, H.; Chen, C.; Matsuda, H.; Sheppard, D.; Yokosaki, Y. Osteopontin undergoes polymerization in vivo and gains chemotactic activity for neutrophils mediated by integrin alpha9beta1. J. Biol. Chem. 2011, 286, 11170–11178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnihotri, R.; Crawford, H.C.; Haro, H.; Matrisian, L.M.; Havrda, M.C.; Liaw, L. Osteopontin, a novel substrate for matrix metalloproteinase-3 (stromelysin-1) and matrix metalloproteinase-7 (matrilysin). J. Biol. Chem. 2001, 276, 28261–28267. [Google Scholar] [CrossRef] [Green Version]
- Kon, S.; Nakayama, Y.; Matsumoto, N.; Ito, K.; Kanayama, M.; Kimura, C.; Kouro, H.; Ashitomi, D.; Matsuda, T.; Uede, T. A novel cryptic binding motif, LRSKSRSFQVSDEQY, in the C-terminal fragment of MMP-3/7-cleaved osteopontin as a novel ligand for α9β1 integrin is involved in the anti-type II collagen antibody-induced arthritis. PLoS ONE 2014, 9, e116210. [Google Scholar] [CrossRef]
- Passmore, M.; Nataatmadja, M.; Fung, Y.L.; Pearse, B.; Gabriel, S.; Tesar, P.; Fraser, J.F. Osteopontin alters endothelial and valvular interstitial cell behaviour in calcific aortic valve stenosis through HMGB1 regulation. Eur. J. Cardiothorac. Surg. 2015, 48, e20–e29. [Google Scholar] [CrossRef] [Green Version]
- Zhiduleva, E.V.; Irtyuga, O.B.; Shishkova, A.A.; Ignat’eva, E.V.; Kostina, A.S.; Levchuk, K.A.; Golovkin, A.S.; Rylov, A.Y.; Kostareva, A.A.; Moiseeva, O.M.; et al. Cellular Mechanisms of Aortic Valve Calcification. Bull. Exp. Biol. Med. 2018, 164, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Grau, J.B.; Poggio, P.; Sainger, R.; Vernick, W.J.; Seefried, W.F.; Branchetti, E.; Field, B.C.; Bavaria, J.E.; Acker, M.A.; Ferrari, G. Analysis of osteopontin levels for the identification of asymptomatic patients with calcific aortic valve disease. Ann. Thorac. Surg. 2012, 93, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahles, F.; Findeisen, H.M.; Bruemmer, D. Osteopontin: A novel regulator at the cross roads of inflammation, obesity and diabetes. Mol. Metab. 2014, 3, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Pohjolainen, V.; Taskinen, P.; Soini, Y.; Rysa, J.; Ilves, M.; Juvonen, T.; Ruskoaho, H.; Leskinen, H.; Satta, J. Noncollagenous bone matrix proteins as a part of calcific aortic valve disease regulation. Hum. Pathol. 2008, 39, 1695–1701. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene of Interest | Forward Primer | Reverse Primer |
|---|---|---|
| Ribosomal protein L29 (RPL29) | CCAAGTCCAAGAACCACACC | TATCGTTGTGATCGGGGTTT |
| Osteopontin (OPN) | GATGGCCGAGGTGATAGTGT | TCGTCTTCTTAGGTGCGTCA |
| Collagen type 1 alpha 1 (COL1A1) | AAGACATCCCACCAGTCACC | TAAGTTCGTCGCAGATCACG |
| Alkaline phosphatase (ALPL) | GACATCGCCTACCAGCTCAT | CACATCGGTTCTGTTCTTGG |
| Hypoxia-inducible factor 1 alpha (HIF1α) | GAAGCAAAGAATCCATTTTCCA | TGGTGACAACTGATCGAAGG |
| Transforming growth factor beta (TGFβ) | GAGCCAGAGGCGGACTACTA | TCGGACGTGTTGAAGAACAT |
| Vascular endothelial growth factor (VEGF) | CGGATCAAACCTCACCAAAG | AAATGCTTTCTCCGCTCTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niazy, N.; Barth, M.; Selig, J.I.; Feichtner, S.; Shakiba, B.; Candan, A.; Albert, A.; Preuß, K.; Lichtenberg, A.; Akhyari, P. Degeneration of Aortic Valves in a Bioreactor System with Pulsatile Flow. Biomedicines 2021, 9, 462. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9050462
Niazy N, Barth M, Selig JI, Feichtner S, Shakiba B, Candan A, Albert A, Preuß K, Lichtenberg A, Akhyari P. Degeneration of Aortic Valves in a Bioreactor System with Pulsatile Flow. Biomedicines. 2021; 9(5):462. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9050462
Chicago/Turabian StyleNiazy, Naima, Mareike Barth, Jessica I. Selig, Sabine Feichtner, Babak Shakiba, Asya Candan, Alexander Albert, Karlheinz Preuß, Artur Lichtenberg, and Payam Akhyari. 2021. "Degeneration of Aortic Valves in a Bioreactor System with Pulsatile Flow" Biomedicines 9, no. 5: 462. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9050462