
Significance of Sex Differences in ncRNAs Expression and Function in Pregnancy and Related Complications

 , , and
, , and
Abstract
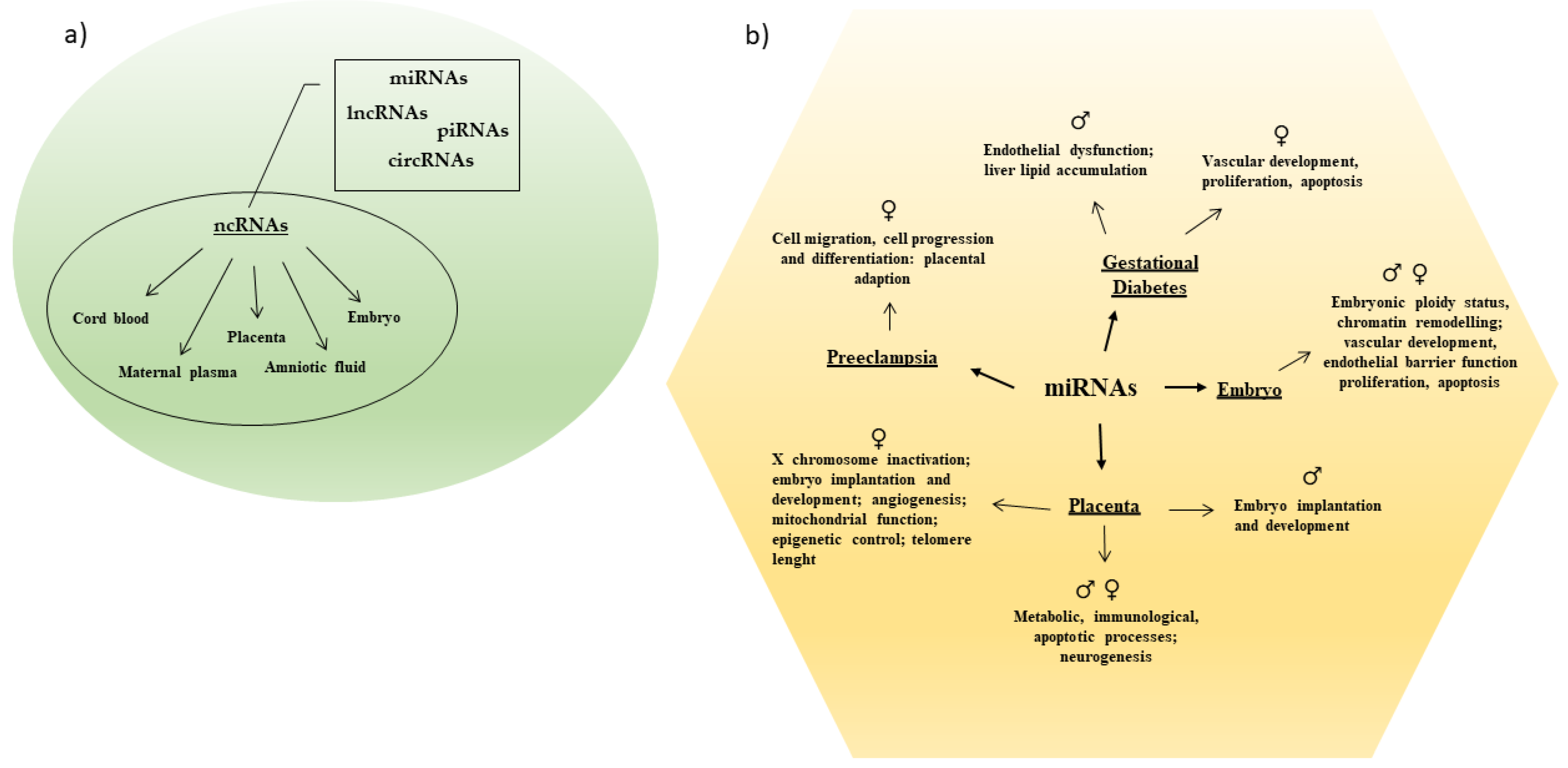
:1. Introduction
2. ncRNAs as Potential Biomarkers
3. The Importance of Sexual Dimorphism in Placental ncRNAs Expression

{kind=link}
{kind=link}
| Model | ncRNAs | Sex Regulation Female ♀ vs. Male ♂ Fetus | Targeted Genes/Processes | Study |
| Overweight and obese pregnant women | miRNAs | ♀ (↑) miR-210 | ♀ (↑) TNFα and NFκB1; mitochondrial dysfunction | [66] |
| Increasing maternal pre-pregnancy BMI | miRNAs | ♀ (↓) miR-20a, miR-34a, miR-146a, miR-210, and miR-222 | ♀♂ Cell proliferation, cell growth and invasion, inflammation, angiogenesis, and oxidative stress | [65] |
| Healthy human pregnancies | miRNAs | ♀ (↑) miR-21, miR-34a, miR-146a, miR-210, and miR-222 | ♀♂ Inflammation, oxidative stress, cell cycle, cellular senescence, and apoptosis ♀ longer placental relative telomere length | [64] |
| Human low gestational age newborns (<28 weeks gestational age) | miRNAs | ♀ (↑) 27 miRNAs, including miR-323b, miR-15a, and miR-223 ♂ (↑) 32 miRNAs, including miR-23a, miR-543, miR-495, miR-323a, miR-30a, miR-155, and miR-222 | ♀♂ Embryo implantation and development ♀ X chromosome inactivation, epigenetic control (HDAC1, HDAC8), and cell cycle progression (CDKN1B) | [69] |
| Human low gestational age newborns (<28 weeks gestational age) | miRNAs | ♀ (↑) miR-4747-5p, miR-3674, miR-4252, miR-4461, miR-1285-5p, miR-4421, miR-4728-3p, miR-6081, miR-6890-3p, iR-564, miR-1225-3p, miR-541-3,p miR-3187-5p, miR-1976, miR-548w, miR-5589-5p, miR-889-3p, miR-4734 ♂ (↑) miR-4057, (↓) miR-128 | ♀♂ Underweight status ♀ No target genes associated with underweight status ♂ regulate 43 genes related to nutrition, growth, and angiogenesis | [70] |
| Healthy human pregnancies | miRNAs | ♀ 76 (↑), and 66 (↓) miRNAs, most significant miR-361-5p | ♀♂ Metabolic, immunological, apoptotic, and neurogenesis processes underlying sex differences | [63] |
| ♀ (↑) miR-361-5p, miR-139-5p, miR-4769-3p | (↓) PCDH11X | |||
| ♀ (↑) miR-520e, miR-518d-3p, miR-331-3p, miR-423-3p, and miR-330-3p | (↓)CD99 gene | |||
| ♀ (↑) miR-4287 | (↓)DDX3Y | |||
| ♀ (↓) miR-1207-5p, miR-4530, and miR-4687-3p | (↑)KDM6A | |||
| ♀ (↓) miR-4299 | (↑)CDK16 | |||
| Healthy pregnancy from women with/without daily supplement with n-3 LCPUFA | miRNAs | ♂ (↑) basal miR-99a | ♀♂ Maternal-fetal amino acids homeostasis | [72] |
| ♀ (↑) miR-99a following supplementation | ♂ ↑ basal mTOR and SLC7A5 mRNA ♀ ↑ mTOR and SLC7A5 mRNA following supplementation | |||
| Healthy pregnancies from major ethnic groups: African Americans, European Americans, South Asians, and East Asians | miRNAs | ♂ (↑) 14 miRNAs 371a-5p,372-3p,181b-2-3p, let-7g-3p, 185-3p, 3615, and 3158-3p | ♂ glutamate receptor signaling and endocrine processes | [73] |
| ♀ (↑) 18 miRNAs | ♀ steroid hormones, estradiol, and glucocorticoid response; differentiation and metabolic processes |
4. Sex-Based ncRNAs Expression in Fetal Development
5. ncRNAs in Pregnancy Complications: Role of Fetal Sex
| Model/Tissue | ncRNAs | Sex Regulation Female♀vs. Male♂Fetus | Targeted Genes/Processes | Study |
| GDM(human feto-placental endothelial cells) | miRNAs | ♀ differently expressed: miR-195-5p, miR-145-5p, miR-494-3p, miR-656-3p, miR-590-3p, miR-299-5p, miR-342-3p, miR-134-5p, miR-574-3p, miR-let-7e-5p, miR-193a-5p, miR-409-5p, miR-24-3p, miR-17-3p, miR-376b-3p, miR-186-5p, miR-32-5p, miR-324-5p, miR-377-3p, miR-4521, miR-1307-5p, miR-335-5p | ♀ signaling cascade regulating vascular development, proliferation, and apoptosis | [91] |
| ♂ differently expressed: miR-100-5p, miR-99b-5p, miR-29b-3p, miR-136-5p | ♂ ECM receptor interaction: endothelial dysfunctions and diabetic complications | |||
| GDM(fetuses rat) | miRNAs | ♂ (↓) miR-130 | ♂ (↑) PPARγ and the lipogenic genes FAS and ACC1: liver lipids accumulation | [93] |
| ♀ (↓) miR-9 and ♂(↓) miR-122 | ♀♂ (↑) PPARδ liver alterations | |||
| GDM (human amniotic fluid; 2nd trimester) | miRNAs | ♀ (↑) miR-378a-3p, miR-885-5p, miR-7-1-3p ♂ (↑) miR-199a-3p | ♀♂ (↓) ABCA1, PAK4, and INSR involved in cholesterol delivery in the liver and placenta, in hepatocyte proliferation, and fetal growth | [92] |
| PE (Plasma) | miRNAs | ♀♂ No differences in exosomal miRNAs | [100] | |
| PE (Placenta) | miRNAs | ♀ (↑) mir-138 | ♀♂ Cell migration, cell cycle progression, and cell differentiation: placental adaption | [32] |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hsu, C.N.; Tain, Y.L. The Good, the Bad, and the Ugly of Pregnancy Nutrients and Developmental Programming of Adult Disease. Nutrients 2019, 11, 894. [Google Scholar] [CrossRef] [Green Version]
- Barker, D.J. Developmental origins of adult health and disease. J. Epidemiol. Community Health 2004, 58, 114–115. [Google Scholar] [CrossRef] [Green Version]
- Clifton, V.L. Review: Sex and the human placenta: Mediating differential strategies of fetal growth and survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef]
- Vari, R.; Scazzocchio, B.; Silenzi, A.; Giovannini, C.; Masella, R. Obesity-Associated Inflammation: Does Curcumin Exert a Beneficial Role? Nutrients 2021, 13, 21. [Google Scholar] [CrossRef]
- Cowell, W.; Colicino, E.; Askowitz, T.; Nentin, F.; Wright, R.J. Fetal sex and maternal postpartum depressive symptoms: Findings from two prospective pregnancy cohorts. Biol. Sex Differ. 2021, 12, 6. [Google Scholar] [CrossRef]
- Enninga, E.A.; Nevala, W.K.; Creedon, D.J.; Markovic, S.N.; Holtan, S.G. Fetal sex-based differences in maternal hormones, angiogenic factors, and immune mediators during pregnancy and the postpartum period. Am. J. Reprod. Immunol. 2015, 73, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funaki, S.; Ogawa, K.; Ozawa, N.; Okamoto, A.; Morisaki, N.; Sago, H. Differences in pregnancy complications and outcomes by fetal gender among Japanese women: A multicenter cross-sectional study. Sci. Rep. 2020, 10, 18810. [Google Scholar] [CrossRef]
- Broere-Brown, Z.A.; Adank, M.C.; Benschop, L.; Tielemans, M.; Muka, T.; Goncalves, R.; Bramer, W.M.; Schoufour, J.D.; Voortman, T.; Steegers, E.A.P.; et al. Fetal sex and maternal pregnancy outcomes: A systematic review and meta-analysis. Biol. Sex Differ. 2020, 11, 26. [Google Scholar] [CrossRef]
- Alur, P. Sex Differences in Nutrition, Growth, and Metabolism in Preterm Infants. Front. Pediatrics 2019, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, T.; Chen, T.S.; Liang, F.W.; Kuo, P.L. Does sex matter? Association of fetal sex and parental age with pregnancy outcomes in Taiwan: A cohort study. BMC Pregnancy Childbirth 2020, 20, 348. [Google Scholar] [CrossRef] [PubMed]
- Franzago, M.; Santurbano, D.; Vitacolonna, E.; Stuppia, L. Genes and Diet in the Prevention of Chronic Diseases in Future Generations. Int. J. Mol. Sci. 2020, 21, 2633. [Google Scholar] [CrossRef]
- DiPietro, J.A.; Voegtline, K.M. The gestational foundation of sex differences in development and vulnerability. Neuroscience 2017, 342, 4–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamalidehghan, B.; Habibi, M.; Afjeh, S.S.; Shoai, M.; Alidoost, S.; Almasi Ghale, R.; Eshghifar, N.; Pouresmaeili, F. The Importance of Small Non-Coding RNAs in Human Reproduction: A Review Article. Appl. Clin. Genet. 2020, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Hadlich, F.; Abbas, M.W.; Iqbal, M.A.; Tesfaye, D.; Bouma, G.J.; Winger, Q.A.; Ponsuksili, S. MicroRNA-mRNA Networks in Pregnancy Complications: A Comprehensive Downstream Analysis of Potential Biomarkers. Int. J. Mol. Sci. 2021, 22, 2313. [Google Scholar] [CrossRef]
- Szilagyi, M.; Pos, O.; Marton, E.; Buglyo, G.; Soltesz, B.; Keseru, J.; Penyige, A.; Szemes, T.; Nagy, B. Circulating Cell-Free Nucleic Acids: Main Characteristics and Clinical Application. Int. J. Mol. Sci. 2020, 21, 6827. [Google Scholar] [CrossRef]
- Reda El Sayed, S.; Cristante, J.; Guyon, L.; Denis, J.; Chabre, O.; Cherradi, N. MicroRNA Therapeutics in Cancer: Current Advances and Challenges. Cancers 2021, 13, 2680. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jimenez, E.; Andres-Leon, E. The Implications of ncRNAs in the Development of Human Diseases. Non-Coding RNA 2021, 7, 17. [Google Scholar] [CrossRef]
- Dai, R.; Ahmed, S.A. Sexual dimorphism of miRNA expression: A new perspective in understanding the sex bias of autoimmune diseases. Ther. Clin. risk Manag. 2014, 10, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Liu, C.J.; Zhai, Z.S.; Zhang, X.; Qin, T.; Zhang, H.W. Single-Cell Non-coding RNA in Embryonic Development. Adv. Exp. Med. Biol. 2018, 1068, 19–32. [Google Scholar] [CrossRef]
- McElhinney, J.; Hasan, A.; Sajini, A.A. The epitranscriptome landscape of small noncoding RNAs in stem cells. Stem Cells 2020, 38, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Han Li, C.; Chen, Y. Small and Long Non-Coding RNAs: Novel Targets in Perspective Cancer Therapy. Curr. Genom. 2015, 16, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Kaushik, A.C.; Zhang, J. The Emerging Role of Major Regulatory RNAs in Cancer Control. Front. Oncol. 2019, 9, 920. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Imai-Sumida, M.; Tanaka, Y.; Dahiya, R. Interaction and cross-talk between non-coding RNAs. Cell. Mol. Life Sci. CMLS 2018, 75, 467–484. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Fioravanti, A.; Pirtoli, L.; Giordano, A.; Dotta, F. Crosstalk between MicroRNA and Oxidative Stress in Physiology and Pathology. Int. J. Mol. Sci. 2020, 21, 1270. [Google Scholar] [CrossRef] [Green Version]
- Reza, A.; Choi, Y.J.; Han, S.G.; Song, H.; Park, C.; Hong, K.; Kim, J.H. Roles of microRNAs in mammalian reproduction: From the commitment of germ cells to peri-implantation embryos. Biol. Rev. Camb. Philos. Soc. 2019, 94, 415–438. [Google Scholar] [CrossRef]
- Winger, E.E.; Reed, J.L.; Ji, X.; Gomez-Lopez, N.; Pacora, P.; Romero, R. MicroRNAs isolated from peripheral blood in the first trimester predict spontaneous preterm birth. PLoS ONE 2020, 15, e0236805. [Google Scholar] [CrossRef]
- Kim, S.H.; MacIntyre, D.A.; Binkhamis, R.; Cook, J.; Sykes, L.; Bennett, P.R.; Terzidou, V. Maternal plasma miRNAs as potential biomarkers for detecting risk of small-for-gestational-age births. EBioMedicine 2020, 62, 103145. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Taheri, M. The role of microRNAs in ectopic pregnancy: A concise review. Non-Coding RNA Res. 2020, 5, 67–70. [Google Scholar] [CrossRef]
- Filardi, T.; Catanzaro, G.; Mardente, S.; Zicari, A.; Santangelo, C.; Lenzi, A.; Morano, S.; Ferretti, E. Non-Coding RNA: Role in Gestational Diabetes Pathophysiology and Complications. Int. J. Mol. Sci. 2020, 21, 4020. [Google Scholar] [CrossRef]
- Leseva, M.N.; Binder, A.M.; Ponsonby, A.L.; Vuillermin, P.; Saffery, R.; Michels, K.B. Barwon Infant Study Investigator Group. Differential gene expression and limited epigenetic dysregulation at the materno-fetal interface in preeclampsia. Hum. Mol. Genet. 2020, 29, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Hirakata, S.; Siomi, M.C. piRNA biogenesis in the germline: From transcription of piRNA genomic sources to piRNA maturation. Biochim. Biophys. Acta 2016, 1859, 82–92. [Google Scholar] [CrossRef]
- Williams, Z.; Morozov, P.; Mihailovic, A.; Lin, C.; Puvvula, P.K.; Juranek, S.; Rosenwaks, Z.; Tuschl, T. Discovery and Characterization of piRNAs in the Human Fetal Ovary. Cell Rep. 2015, 13, 854–863. [Google Scholar] [CrossRef] [Green Version]
- Ha, H.; Song, J.; Wang, S.; Kapusta, A.; Feschotte, C.; Chen, K.C.; Xing, J. A comprehensive analysis of piRNAs from adult human testis and their relationship with genes and mobile elements. BMC Genom. 2014, 15, 545. [Google Scholar] [CrossRef] [Green Version]
- Ramat, A.; Simonelig, M. Functions of PIWI Proteins in Gene Regulation: New Arrows Added to the piRNA Quiver. Trends Genet. 2021, 37, 188–200. [Google Scholar] [CrossRef]
- Rayford, K.J.; Cooley, A.; Rumph, J.T.; Arun, A.; Rachakonda, G.; Villalta, F.; Lima, M.F.; Pratap, S.; Misra, S.; Nde, P.N. piRNAs as Modulators of Disease Pathogenesis. Int. J. Mol. Sci. 2021, 22, 2373. [Google Scholar] [CrossRef]
- Huang, X.; Wong, G. An old weapon with a new function: PIWI-interacting RNAs in neurodegenerative diseases. Transl. Neurodegener. 2021, 10, 9. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Zhang, Q.; Wang, Y.; Wang, Y.; Liu, D.; He, Y.; Wei, X.; Gu, H.; Ma, W.; Luo, W.; et al. PIWI-interacting RNA sequencing profiles in maternal plasma-derived exosomes reveal novel non-invasive prenatal biomarkers for the early diagnosis of nonsyndromic cleft lip and palate. EBioMedicine 2021, 65, 103253. [Google Scholar] [CrossRef]
- Chen, H.; Shan, G. The physiological function of long-noncoding RNAs. Non-Coding RNA Res. 2020, 5, 178–184. [Google Scholar] [CrossRef]
- Fort, V.; Khelifi, G.; Hussein, S.M.I. Long non-coding RNAs and transposable elements: A functional relationship. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118837. [Google Scholar] [CrossRef]
- Brockdorff, N. Localized accumulation of Xist RNA in X chromosome inactivation. Open Biol. 2019, 9, 190213. [Google Scholar] [CrossRef] [Green Version]
- Dey, B.K.; Mueller, A.C.; Dutta, A. Long non-coding RNAs as emerging regulators of differentiation, development, and disease. Transcription 2014, 5, e944014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, D.; Fang, C.; Deng, N.; Yao, B.; Fan, C. Long noncoding RNA expression profiling identifies MIR210HG as a novel molecule in severe preeclampsia. Life Sci. 2021, 270, 119121. [Google Scholar] [CrossRef]
- Zhang, W.; Cao, D.; Wang, Y.; Ren, W. LncRNA MEG8 is upregulated in gestational diabetes mellitus (GDM) and predicted kidney injury. J. Diabetes Its Complicat. 2021, 35, 107749. [Google Scholar] [CrossRef]
- Azari, I.; Ghafouri-Fard, S.; Omrani, M.D.; Arsang-Jang, S.; Kordi Tamandani, D.M.; Saroone Rigi, M.; Rafiee, S.; Pouresmaeili, F.; Taheri, M. Expression of Long Non-Coding RNAs in Placentas of Intrauterine Growth Restriction (IUGR) Pregnancies. Rep. Biochem. Mol. Biol. 2019, 8, 25–31. [Google Scholar]
- Luo, M.; Xiao, H.; Wang, L.; Zhao, J.; Gao, J.; Ma, W. The expression and clinical significance of three lncRNAs in patients with a missed abortion. Exp. Ther. Med. 2021, 21, 8. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Zhao, H.; Gong, L.; Xiao, X.; Zhou, Q.; Lu, H.; Cui, Y.; Xu, H.; Wu, S.; Tang, Y.; et al. Differentially expressed circular RNAs and the competing endogenous RNA network associated with preeclampsia. Placenta 2021, 103, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Di Agostino, S.; Riccioli, A.; De Cesaris, P.; Fontemaggi, G.; Blandino, G.; Filippini, A.; Fazi, F. Circular RNAs in Embryogenesis and Cell Differentiation with a Focus on Cancer Development. Front. Cell Dev. Biol. 2020, 8, 389. [Google Scholar] [CrossRef]
- Wen, G.; Zhou, T.; Gu, W. The potential of using blood circular RNA as liquid biopsy biomarker for human diseases. Protein Cell 2020. [Google Scholar] [CrossRef]
- Yang, H.; Ye, W.; Chen, R.; Zeng, F.; Long, Y.; Zhang, X.; Ma, J.; Gan, Q.; Rehemutula, R.; Zhu, C. Circulating expression of Hsa_circRNA_102893 contributes to early gestational diabetes mellitus detection. Sci. Rep. 2020, 10, 19046. [Google Scholar] [CrossRef]
- Jiang, M.; Lash, G.E.; Zhao, X.; Long, Y.; Guo, C.; Yang, H. CircRNA-0004904, CircRNA-0001855, and PAPP-A: Potential Novel Biomarkers for the Prediction of Preeclampsia. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 2576–2586. [Google Scholar] [CrossRef]
- Jia, N.; Li, J. Role of Circular RNAs in Preeclampsia. Dis. Markers 2019, 2019, 7237495. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, C.S. Sex-Specific Placental Responses in Fetal Development. Endocrinology 2015, 156, 3422–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dearden, L.; Bouret, S.G.; Ozanne, S.E. Sex and gender differences in developmental programming of metabolism. Mol. Metab. 2018, 15, 8–19. [Google Scholar] [CrossRef]
- Cowell, W.; Deyssenroth, M.; Chen, J.; Wright, R.J. Maternal stress in relation to sex-specific expression of placental genes involved in nutrient transport, oxygen tension, immune response, and the glucocorticoid barrier. Placenta 2020, 96, 19–26. [Google Scholar] [CrossRef]
- Saoi, M.; Kennedy, K.M.; Gohir, W.; Sloboda, D.M.; Britz-McKibbin, P. Placental Metabolomics for Assessment of Sex- specific Differences in Fetal Development During Normal Gestation. Sci. Rep. 2020, 10, 9399. [Google Scholar] [CrossRef] [PubMed]
- Mouillet, J.F.; Chu, T.; Sadovsky, Y. Expression patterns of placental microRNAs. Birth Defects Res. A Clin. Mol. Teratol. 2011, 91, 737–743. [Google Scholar] [CrossRef] [Green Version]
- Medina-Bastidas, D.; Guzman-Huerta, M.; Borboa-Olivares, H.; Ruiz-Cruz, C.; Parra-Hernandez, S.; Flores-Pliego, A.; Salido-Guadarrama, I.; Camargo-Marin, L.; Arambula-Meraz, E.; Estrada-Gutierrez, G. Placental Microarray Profiling Reveals Common mRNA and lncRNA Expression Patterns in Preeclampsia and Intrauterine Growth Restriction. Int. J. Mol. Sci. 2020, 21, 3597. [Google Scholar] [CrossRef]
- Ali, A.; Bouma, G.J.; Anthony, R.V.; Winger, Q.A. The Role of LIN28-let-7-ARID3B Pathway in Placental Development. Int. J. Mol. Sci. 2020, 21, 3637. [Google Scholar] [CrossRef] [PubMed]
- Majewska, M.; Lipka, A.; Paukszto, L.; Jastrzebski, J.P.; Gowkielewicz, M.; Jozwik, M.; Majewski, M.K. Preliminary RNA-Seq Analysis of Long Non-Coding RNAs Expressed in Human Term Placenta. Int. J. Mol. Sci. 2018, 19, 1894. [Google Scholar] [CrossRef] [Green Version]
- Tsamou, M.; Vrijens, K.; Wang, C.; Winckelmans, E.; Neven, K.Y.; Madhloum, N.; de Kok, T.M.; Nawrot, T.S. Genome-wide microRNA expression analysis in human placenta reveals sex-specific patterns: An ENVIRONAGE birth cohort study. Epigenetics 2021, 16, 373–388. [Google Scholar] [CrossRef]
- Tsamou, M.; Martens, D.S.; Cox, B.; Madhloum, N.; Vrijens, K.; Nawrot, T.S. Sex-specific associations between telomere length and candidate miRNA expression in placenta. J. Transl. Med. 2018, 16, 254. [Google Scholar] [CrossRef]
- Tsamou, M.; Martens, D.S.; Winckelmans, E.; Madhloum, N.; Cox, B.; Gyselaers, W.; Nawrot, T.S.; Vrijens, K. Mother’s Pre-pregnancy BMI and Placental Candidate miRNAs: Findings from the ENVIRONAGE Birth Cohort. Sci. Rep. 2017, 7, 5548. [Google Scholar] [CrossRef]
- Muralimanoharan, S.; Guo, C.; Myatt, L.; Maloyan, A. Sexual dimorphism in miR-210 expression and mitochondrial dysfunction in the placenta with maternal obesity. Int. J. Obes. 2015, 39, 1274–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anton, L.; DeVine, A.; Polyak, E.; Olarerin-George, A.; Brown, A.G.; Falk, M.J.; Elovitz, M.A. HIF-1alpha Stabilization Increases miR-210 Eliciting First Trimester Extravillous Trophoblast Mitochondrial Dysfunction. Front. Physiol. 2019, 10, 699. [Google Scholar] [CrossRef]
- Frazier, S.; McBride, M.W.; Mulvana, H.; Graham, D. From animal models to patients: The role of placental microRNAs, miR-210, miR-126, and miR-148a/152 in preeclampsia. Clin. Sci. 2020, 134, 1001–1025. [Google Scholar] [CrossRef]
- Eaves, L.A.; Phookphan, P.; Rager, J.E.; Bangma, J.; Santos, H.P., Jr.; Smeester, L.; O’Shea, T.M.; Fry, R.C. A role for microRNAs in the epigenetic control of sexually dimorphic gene expression in the human placenta. Epigenomics 2020, 12, 1543–1558. [Google Scholar] [CrossRef]
- Clark, J.; Eaves, L.A.; Gaona, A.R.; Santos, H.P.; Smeester, L.; Bangma, J.T.; Rager, J.E.; O’Shea, T.M.; Fry, R.C. Pre-pregnancy BMI-associated miRNA and mRNA expression signatures in the placenta highlight a sexually-dimorphic response to maternal underweight status. Sci. Rep. 2021, 11, 15743. [Google Scholar] [CrossRef] [PubMed]
- Sedlmeier, E.M.; Brunner, S.; Much, D.; Pagel, P.; Ulbrich, S.E.; Meyer, H.H.; Amann-Gassner, U.; Hauner, H.; Bader, B.L. Human placental transcriptome shows sexually dimorphic gene expression and responsiveness to maternal dietary n-3 long-chain polyunsaturated fatty acid intervention during pregnancy. BMC Genom. 2014, 15, 941. [Google Scholar] [CrossRef] [Green Version]
- Sedlmeier, E.M.; Meyer, D.M.; Stecher, L.; Sailer, M.; Daniel, H.; Hauner, H.; Bader, B.L. Fetal sex modulates placental microRNA expression, potential microRNA-mRNA interactions, and levels of amino acid transporter expression and substrates: INFAT study subpopulation analysis of n-3 LCPUFA intervention during pregnancy and associations with offspring body composition. BMC Mol. Cell Biol. 2021, 22, 15. [Google Scholar] [CrossRef]
- Guo, S.; Huang, S.; Jiang, X.; Hu, H.; Han, D.; Moreno, C.S.; Fairbrother, G.L.; Hughes, D.A.; Stoneking, M.; Khaitovich, P. Variation of microRNA expression in the human placenta driven by population identity and sex of the newborn. BMC Genom. 2021, 22, 286. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, A.; Drapkina, Y.; Fedorov, I.; Chagovets, V.; Makarova, N.; Shamina, M.; Kalinina, E.; Sukhikh, G. Small Noncoding RNA Signatures for Determining the Developmental Potential of an Embryo at the Morula Stage. Int. J. Mol. Sci. 2020, 21, 9399. [Google Scholar] [CrossRef]
- Paul, A.B.M.; Sadek, S.T.; Mahesan, A.M. The role of microRNAs in human embryo implantation: A review. J. Assist. Reprod. Genet. 2019, 36, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Salas-Huetos, A.; James, E.R.; Aston, K.I.; Jenkins, T.G.; Carrell, D.T.; Yeste, M. The Expression of miRNAs in Human Ovaries, Oocytes, Extracellular Vesicles, and Early Embryos: A Systematic Review. Cells 2019, 8, 1564. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhn, S.; Ali, A.; Hossain, M.; Hoelker, M.; Salilew-Wondim, D.; Anthony, R.V.; Tesfaye, D. MicroRNA-Mediated Gene Regulatory Mechanisms in Mammalian Female Reproductive Health. Int. J. Mol. Sci. 2021, 22, 938. [Google Scholar] [CrossRef] [PubMed]
- Rosenbluth, E.M.; Shelton, D.N.; Sparks, A.E.; Devor, E.; Christenson, L.; Van Voorhis, B.J. MicroRNA expression in the human blastocyst. Fertil. Steril. 2013, 99, 855–861. [Google Scholar] [CrossRef]
- Ciaudo, C.; Jay, F.; Okamoto, I.; Chen, C.J.; Sarazin, A.; Servant, N.; Barillot, E.; Heard, E.; Voinnet, O. Correction: RNAi-Dependent and Independent Control of LINE1 Accumulation and Mobility in Mouse Embryonic Stem Cells. PLoS Genet. 2015, 11, e1005247. [Google Scholar] [CrossRef] [PubMed]
- Cvitic, S.; Strutz, J.; Appel, H.M.; Weiss, E.; Brandl, W.T.; Thuringer, A.; Bernhart, E.M.; Lassance, L.; Wadsack, C.; Schliefsteiner, C.; et al. Sexual dimorphism of miRNA signatures in feto-placental endothelial cells is associated with altered barrier function and actin organization. Clin. Sci. 2020, 134, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.K.; Gunel, T.; Gumusoglu, E.; Benian, A.; Aydinli, K. MicroRNA expression profiling in placenta and maternal plasma in early pregnancy loss. Mol. Med. Rep. 2018, 17, 4941–4952. [Google Scholar] [CrossRef] [PubMed]
- Joglekar, M.V.; Wong, W.K.M.; Ema, F.K.; Georgiou, H.M.; Shub, A.; Hardikar, A.A.; Lappas, M. Postpartum circulating microRNA enhances prediction of future type 2 diabetes in women with previous gestational diabetes. Diabetologia 2021, 64, 1516–1526. [Google Scholar] [CrossRef]
- Scalavino, V.; Liso, M.; Cavalcanti, E.; Gigante, I.; Lippolis, A.; Mastronardi, M.; Chieppa, M.; Serino, G. miR-369-3p modulates inducible nitric oxide synthase and is involved in regulation of chronic inflammatory response. Sci. Rep. 2020, 10, 15942. [Google Scholar] [CrossRef]
- Hromadnikova, I.; Kotlabova, K.; Dvorakova, L.; Krofta, L. Postpartum profiling of microRNAs involved in pathogenesis of cardiovascular/cerebrovascular diseases in women exposed to pregnancy-related complications. Int. J. Cardiol. 2019, 291, 158–167. [Google Scholar] [CrossRef]
- Sorensen, A.E.; van Poppel, M.N.M.; Desoye, G.; Damm, P.; Simmons, D.; Jensen, D.M.; Dalgaard, L.T.; The, D. ALI Core Investigator Group. The Predictive Value of miR-16, -29a and -134 for Early Identification of Gestational Diabetes: A Nested Analysis of the DALI Cohort. Cells 2021, 10, 170. [Google Scholar] [CrossRef]
- Sun, D.G.; Tian, S.; Zhang, L.; Hu, Y.; Guan, C.Y.; Ma, X.; Xia, H.F. The miRNA-29b Is Downregulated in Placenta During Gestational Diabetes Mellitus and May Alter Placenta Development by Regulating Trophoblast Migration and Invasion Through a HIF3A-Dependent Mechanism. Front. Endocrinol. 2020, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.B.; Chernausek, S.D.; Teague, A.M.; Bard, D.E.; Tryggestad, J.B. Maternal diabetes alters microRNA expression in fetal exosomes, human umbilical vein endothelial cells and placenta. Pediatric Res. 2021, 89, 1157–1163. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Zhang, L.; Lin, Y.; Li, Z.; Xu, J.; Shi, Z.; Chen, Z.; Ma, J.; Wen, J. Circular RNA expression profiles in umbilical cord blood exosomes from normal and gestational diabetes mellitus patients. Biosci. Rep. 2020, 40, BSR20201946. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Wu, Y.; Li, Z.; Wang, D.; Cai, S.; Wang, Z. The role of circular RNA circ_0008285 in gestational diabetes mellitus by regulating the biological functions of trophoblasts. Biol. Res. 2021, 54, 14. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, Y.; Xu, L.; Long, W.; Shan, C.; Ding, H.; You, L.; Zhao, C.; Shi, Z. High expression of an unknown long noncoding RNA RP11-290L1.3 from GDM macrosomia and its effect on preadipocyte differentiation. Endocr. Connect. 2021, 10, 191–204. [Google Scholar] [CrossRef] [PubMed]
- Strutz, J.; Cvitic, S.; Hackl, H.; Kashofer, K.; Appel, H.M.; Thuringer, A.; Desoye, G.; Koolwijk, P.; Hiden, U. Gestational diabetes alters microRNA signatures in human feto-placental endothelial cells depending on fetal sex. Clin. Sci. 2018, 132, 2437–2449. [Google Scholar] [CrossRef]
- Joshi, A.; Azuma, R.; Akumuo, R.; Goetzl, L.; Pinney, S.E. Gestational diabetes and maternal obesity are associated with sex-specific changes in miRNA and target gene expression in the fetus. Int. J. Obes. 2020, 44, 1497–1507. [Google Scholar] [CrossRef]
- Fornes, D.; White, V.; Higa, R.; Heinecke, F.; Capobianco, E.; Jawerbaum, A. Sex-dependent changes in lipid metabolism, PPAR pathways and microRNAs that target PPARs in the fetal liver of rats with gestational diabetes. Mol. Cell. Endocrinol. 2018, 461, 12–21. [Google Scholar] [CrossRef]
- Chen, D.; He, B.; Zheng, P.; Wang, S.; Zhao, X.; Liu, J.; Yang, X.; Cheng, W. Identification of mRNA-, circRNA- and lncRNA-Associated ceRNA Networks and Potential Biomarkers for Preeclampsia from Umbilical Vein Endothelial Cells. Front. Mol. Biosci. 2021, 8, 652250. [Google Scholar] [CrossRef]
- Ping, Z.; Ai, L.; Shen, H.; Zhang, X.; Jiang, H.; Song, Y. Identification and comparison of circular RNAs in preeclampsia. PeerJ 2021, 9, e11299. [Google Scholar] [CrossRef]
- Gai, S.; Sun, L.; Wang, H.; Yang, P. Circular RNA hsa_circ_0007121 regulates proliferation, migration, invasion, and epithelial-mesenchymal transition of trophoblast cells by miR-182-5p/PGF axis in preeclampsia. Open Med. 2020, 15, 1061–1071. [Google Scholar] [CrossRef]
- Pankiewicz, K.; Fijalkowska, A.; Issat, T.; Maciejewski, T.M. Insight into the Key Points of Preeclampsia Pathophysiology: Uterine Artery Remodeling and the Role of MicroRNAs. Int. J. Mol. Sci. 2021, 22, 3132. [Google Scholar] [CrossRef]
- Kolkova, Z.; Holubekova, V.; Grendar, M.; Nachajova, M.; Zubor, P.; Pribulova, T.; Loderer, D.; Zigo, I.; Biringer, K.; Hornakova, A. Association of Circulating miRNA Expression with Preeclampsia, Its Onset, and Severity. Diagnostics 2021, 11, 476. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Zhu, F.; Ding, Y. Differential microRNA expression profile in the plasma of preeclampsia and normal pregnancies. Exp. Ther. Med. 2019, 18, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ouyang, Y.; Sadovsky, E.; Parks, W.T.; Chu, T.; Sadovsky, Y. Unique microRNA Signals in Plasma Exosomes from Pregnancies Complicated by Preeclampsia. Hypertension 2020, 75, 762–771. [Google Scholar] [CrossRef]


| Model/Tissue | ncRNAs | Sex Regulation Female ♀ vs. Male ♂ Fetus | Targeted Genes/Processes | Study |
| Cryopreserved human embryos (day 5 of development) | miRNAs | ♀ (↑) miR-182, miR-206, miR-500, miR-601, miR-604, miR-875-5p ♂ (↑) miR-140-5p, miR-149, miR-151-5p, miR-26b, miR-31, miR-362-3p, miR-512-3p, miR-512-5p, miR-518d-5p, miR-518e, miR-525-3p, miR-886-3p, miR-886-5p, miR-92a | ♀♂ Embryo development, cell cycle, and apoptosis | [78] |
| Mouse embryonic stem cells | miRNAs | ♂ (↑) miR-302a, miR-302b, miR-302c, miR-302d, at differentiation day 5 ♂ (↓) miR-302, at differentiation day 10 | ♀♂ Chromatin remodeling; proliferation, differentiation, and cell fate determination | [79] |
| Human feto-placental endothelial cells from healthy pregnancies | miRNAs | ♀ (↑) miR-23a-3p, miR-222-5p, miR-181a-3p, miR-151a-3p, miR-4286 | ♀♂ Endothelial barrier function: adherent junction, ECM receptor interaction, and focal adhesion; | [80] |
| ♂ (↑) miR-29b-3p, miR-15b-5p, miR-431-5p, miR345-5p | ♂ cells have increased barrier resistance compared with female cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varì, R.; Scazzocchio, B.; Filardi, T.; Citarella, A.; Bellenghi, M.; Masella, R.; Santangelo, C. Significance of Sex Differences in ncRNAs Expression and Function in Pregnancy and Related Complications. Biomedicines 2021, 9, 1509. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111509
Varì R, Scazzocchio B, Filardi T, Citarella A, Bellenghi M, Masella R, Santangelo C. Significance of Sex Differences in ncRNAs Expression and Function in Pregnancy and Related Complications. Biomedicines. 2021; 9(11):1509. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111509
Chicago/Turabian StyleVarì, Rosaria, Beatrice Scazzocchio, Tiziana Filardi, Anna Citarella, Maria Bellenghi, Roberta Masella, and Carmela Santangelo. 2021. "Significance of Sex Differences in ncRNAs Expression and Function in Pregnancy and Related Complications" Biomedicines 9, no. 11: 1509. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9111509