
Bird Taxonomic and Functional Diversity in Three Habitats in Buenos Aires City, Argentina

Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires–IEGEBA (CONICET–UBA), Ciudad Universitaria, Pab 2, Piso 4, Buenos Aires 1426, Argentina
*
Author to whom correspondence should be addressed.
Birds 2021, 2(2), 217-229; https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020016
Submission received: 16 April 2021
/
Revised: 1 June 2021
/
Accepted: 1 June 2021
/
Published: 11 June 2021
(This article belongs to the Special Issue Feature Papers of Birds 2021)
Abstract
:Simple Summary
Urban green spaces (UGS), such as parks and wooded streets, are open areas with vegetation that provide sustainability to urban areas. The aim of this study was to analyze the variation of bird assemblages in non-wooded streets, wooded streets, and parks in Buenos Aires City, Argentina. We compared bird taxonomic and functional diversity between these habitat types. We selected five non-wooded streets, five wooded streets, and five parks. Bird surveys were performed in 100 m long and 50 m wide transects. We found that taxonomic diversity had the greatest value in the parks, followed by wooded streets, and then the non-wooded streets. Functional diversity was similar between habitats. The taxonomic and functional composition changed between habitats. The positive association between UGS and bird diversity highlights the role of UGS as biodiversity conservation sites in neotropical cities
Abstract
Urban green spaces (UGS), such as parks and wooded streets, are open areas with vegetation that provide sustainability to urban areas. However, their role in conserving bird diversity in neotropical cities has scarcely been analyzed. The aim of this study was to analyze the variation of bird assemblages in non-wooded streets, wooded streets, and parks in Buenos Aires City, Argentina. We compared the taxonomic and functional diversity between these habitat types. We selected five non-wooded streets, five wooded streets, and five parks in the city. Bird surveys were performed in 100 m long and 50 m wide transects. We found that taxonomic diversity had the greatest value in the parks, followed by wooded streets, and then the non-wooded streets. Functional diversity was similar between habitats. The taxonomic and functional composition changed between habitats. Non-wooded streets were dominated by the Rock Dove (Columba livia) and the Eared Dove (Zenaida auriculata), whereas parks had the highest abundance of the Picazuro Pigeon (Patagioenas picazuro) and the Rufous-bellied Thrush (Turdus rufiventris). Non-wooded streets were dominated by omnivorous and granivorous species, whereas parks had a higher abundance of herbivorous and frugivorous species. The positive association between UGS and bird diversity highlights the role of UGS as biodiversity conservation sites in neotropical cities.
1. Introduction
Urbanization is a process defined as the increase in human concentration, which involves the movement of people from rural to urban areas [1,2,3,4]. A report from the United Nations claimed that in 2018, 55.3% of the human population lived in urban areas, and this figure is projected to increase by 2030, while the rural population is expected to diminish [5]. This process leads to the loss of urban green areas within cities [6]. Urbanization involves intense changes in land use that persist in the long-term [6,7], which strongly and negatively impacts biodiversity [8]. Moreover, urbanization alters biodiversity composition by diminishing native species diversity and favoring widespread exotic species [7,8].
In this paper, we used birds as study taxa because they are indicators of environmental changes and they are easy to survey [9,10]. Several authors have found significant changes in bird abundance, diversity, and composition in urban areas [10,11,12,13]. Diversity has several facets like taxonomic and functional [14]. Taxonomic diversity measures species richness and relative abundance of the species within a community [15]. On the other hand, functional diversity is the variety and range of functional traits of species, which can have an effect on ecosystem functions, such as primary production, nutrient cycling, or heat transfer [16]. Several studies have shown the negative effects of urbanization on bird functional diversity [17,18,19,20], but see [21,22]. The assessment of different diversity facets allows a better approach to biodiversity status in urban ecosystems [14], and the creation of more sustainable cities [8].
Urban green spaces (UGS) are public or private open or semi-open areas with vegetation [23,24]. There are different types of UGS, such as parks and wooded streets [25,26,27]. Parks are generally composed of managed vegetation with a diversity of plants like grass, bushes, and trees [28]. They are urban biodiversity hotspots [29], probably because they have the highest green percentage cover and vegetation heterogeneity. On the other hand, vegetation quantity and heterogeneity declines when moving from wooded streets to non-wooded streets [30].
The comparison of bird assemblages between non-wooded streets, wooded streets, and urban parks has scarcely been analyzed [31]. For example, parks and wooded streets with native vegetation in Melbourne (Australia) had greater total and native bird species richness and bird abundance than wooded streets with exotic and immature vegetation [31]. Fernandez-Juricic [30] found that parks, wooded streets, and non-wooded streets in Madrid (Spain) had significant differences of bird species richness. This same authors also found that there were significant differences in bird composition between habitat types, mainly due to the different habitat requirements of different species [31]. For instance, in Argentina, some species, such as the Chalked-browed Mockingbird (Mimus saturninus), Rufous-bellied Thrush (Turdus rufiventris), and the Rufous Hornero (Furnarius rufous) need trees for nesting and short grass for foraging. On the other hand, the Green-barred Woodpecker (Colaptes melanochloros) uses trees for nesting and foraging [32]. The House Sparrow (Passer domesticus) and the Rock Dove (Columba livia) need buildings for nesting and can forage on impervious surfaces, such as asphalt streets and sidewalks [31,33,34,35]. In addition, functional diversity and functional trait composition have not been compared between parks, wooded, and non-wooded streets yet.
The analysis of bird communities in urban parks, wooded streets, and non-wooded streets is fundamental for understanding which birds occupy these habitats and, therefore, for implementing proper management actions. This study aimed to analyze bird assemblages in different habitat types in Buenos Aires City: parks, wooded streets, and non-wooded streets. We compared the taxonomic and functional diversity and taxonomic and functional composition of the three habitat types. Due to these habitats offering different amounts and varieties of resources to birds, with parks being the most suitable habitat, we expected that parks would have greater taxonomic and functional diversity than wooded or non-wooded streets. Moreover, we predicted a significant variation in species and trait composition between habitat types. Habitats with greater human disturbance, such as noise produced by car traffic, and less habitat diversity are predicted to be associated with exotic species adapted to human disturbance such as the Rock Dove and the House Sparrow. Moreover, non-wooded streets are predicted to be associated with omnivorous species that forage on the ground and have a high mass; whereas habitats with greater habitat diversity will have native birds with insectivorous diets and who forage on vegetation [32,36,37].
2. Methods
2.1. Study Area
The study was performed in Buenos Aires City (34°36′14″ S, 58°22′54″ W; 25 m. a.s.l.; almost three million inhabitants, [38]) in the metropolitan area of Buenos Aires (>14 million inhabitants, [38]). It has a temperate and seasonal climate. The mean annual temperature is 17.7 °C and the mean annual rainfall is 1214.6 mm [39]. Buenos Aires City is located in the Pampa Ondulada region, which was originally composed of a mesophytic pseudo-steppe, a wet meadow, a halophytic steppe, and a xerophytic forest [40]. Currently, the region is mainly composed of cultivated land and tree plantations.
2.2. Study Design
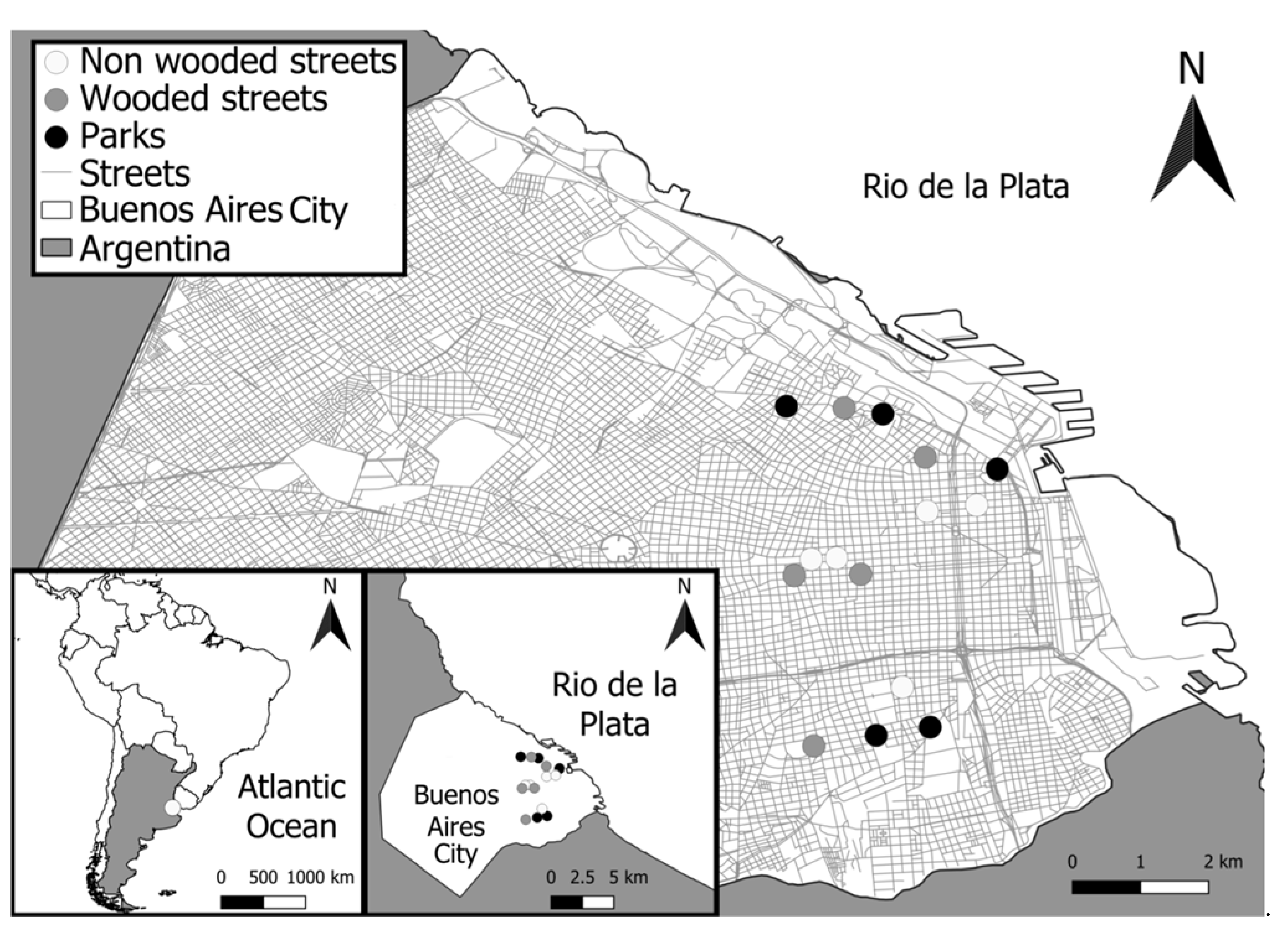
To analyze bird assemblages, we selected three types of urban habitats: five non-wooded streets (NWS); five wooded streets (WS) with trees and some bushes; and five parks composed of trees, shrubs, grass, paved pathways, and monuments that were surrounded by buildings (Figure 1). The largest park was 8.6 ha and the smallest was 1.7 ha (mean= 5.5 ha, sd= 2.5 ha). Within these three habitats, we located a 100 m long and 50 m wide transects where we performed the bird surveys (Figure 1).
2.3. Bird Surveys
Bird surveys were performed between Spring 2015 and Summer 2016 (from October to January) during three visits. These months correspond with the bird breeding period in Argentina [41]. The surveys were performed during the first four hours after dawn, on non-windy, not rainy days, either on business days or on weekends. The transects were visited randomly at different moments of the morning to avoid bias in bird detection. We registered birds that were seen or heard, and which were perching, eating, walking, or fighting with another bird on the transect area.
2.4. Response Variables and Statistical Analysis
2.4.1. Taxonomic and Functional Diversity
Taxonomic diversity was estimated by species richness and species diversity measured by the Shannon–Wiener index (H′) [42], whereas functional diversity was calculated by the functional dispersion index (FDis) [43]. Bird richness was estimated by using iNEXT [44] and COMDYN [45] software. iNEXT aims to compare diversity estimates for equally-large samples by interpolating and extrapolating total species richness [44]. Interpolation and extrapolation curves were computed for each habitat type using bird abundance data and 999 bootstraps. We used the maximum number of individuals for each species during the three visits as a measure of bird abundance. Following the recommendations of [46], curves were estimated with their respective 84% confidence limits. The confidence limits between curves were compared at a minimum sample size of individuals observed between habitats. Non-overlapping confidence areas indicated significant differences in species richness between curves (p < 0.05). On the other hand, COMDYN estimates the average richness for each transect by utilizing presence and absence data during the three visits.
The Shannon–Wiener index (H’) was estimated by using the diversity function from vegan package [47]. This index considers species richness and their relative abundance within a community [15]. Although the H’ index has been criticized [15], several authors have used it in urban bird analyses [19,35,48,49,50,51,52]. Therefore, our use of the H’ index allows comparisons with other studies.
FDis was estimated with the function dbFD from FD package [53]. Functional dispersion, or FDis index, is the distance of the traits values from each species to the centroid within a multidimensional space of all functional traits [43]. FDis can take into account species’ relative abundances and it is unaffected by species richness [43]. We used functional traits involved in obtaining resources: body size (average weight for the species (g)); clutch size (average egg number for the species); foraging sites (ground, air, foliage, or tree trunk); diet (frugivorous, granivorous, herbivorous, invertebrates, or omnivorous); and migratory status. Information about foraging site, diet, and migratory status was treated as presence/absence data. The trait values were taken from the bibliography [41,54,55,56].
Generalized Linear Models (GLMs) were performed to compare transect values of species richness, the Shannon–Wiener index, and FDis index between habitats, using the glm function from the R program. The significance of models was tested by comparing them with null models using likelihood ratio tests (LRT) with the anova function (p < 0.05). To analyze significant differences between the types of habitats, we carried out posteriori tests using the emmeans function from emmeans package [57]. For species richness, we assumed a Poisson distribution of errors and we checked for over- and sub-dispersion. For the Shannon–Wiener index and FDis, we assumed a gaussian distribution of errors, and homoscedasticity and normality were checked. Spatial auto-correlation of residuals was analyzed with the Moran index using SAM software [58], and no significant autocorrelation was found (p < 0.05).
2.4.2. Taxonomic and Functional Traits Composition
Non-metric Multidimensional Scaling (NMDS) was performed to compare taxonomic compositions between habitats. NMDS is a multivariate data analysis [59], which uses dissimilarity data to ordinate sites and species in multiple dimensions. In our study we used the Bray–Curtis distance index [60], which takes into account species’ abundances, using the vegan package. To visualize composition differences between habitats, 95% confidence area ellipses were estimated in the NMDS plot using the oridiellipse function from the vegan package. Significant differences in species composition between the habitat types were tested using the adonis function from the vegan package. Differences in species abundances and their contribution to dissimilarity between habitats were analyzed with the simper (similarity percentage) function from the vegan package. We took into account those species in which the average values of contribution and standard deviation ratio were greater than one or who made a large contribution to dissimilarity and low variation between transects [61].
Fourth corner solution was used to analyze functional trait composition variation between habitats [62]. This analysis considers bird abundance and the type of habitat for each transect and functional trait values for bird species. Then, the analysis calculates interaction coefficients between functional trait values and habitat types. We used the traitglm function from the mvabund package from R program [63]. These analyses were performed in the R software [64].
3. Results
A total of 23 bird species were recorded, of which the Rock Dove and the Eared Dove were the most abundant. In parks, 20 species were observed, whereas in wooded streets 10 species were observed and in non-wooded streets six species were found.
3.1. Taxonomic and Functional Diversity
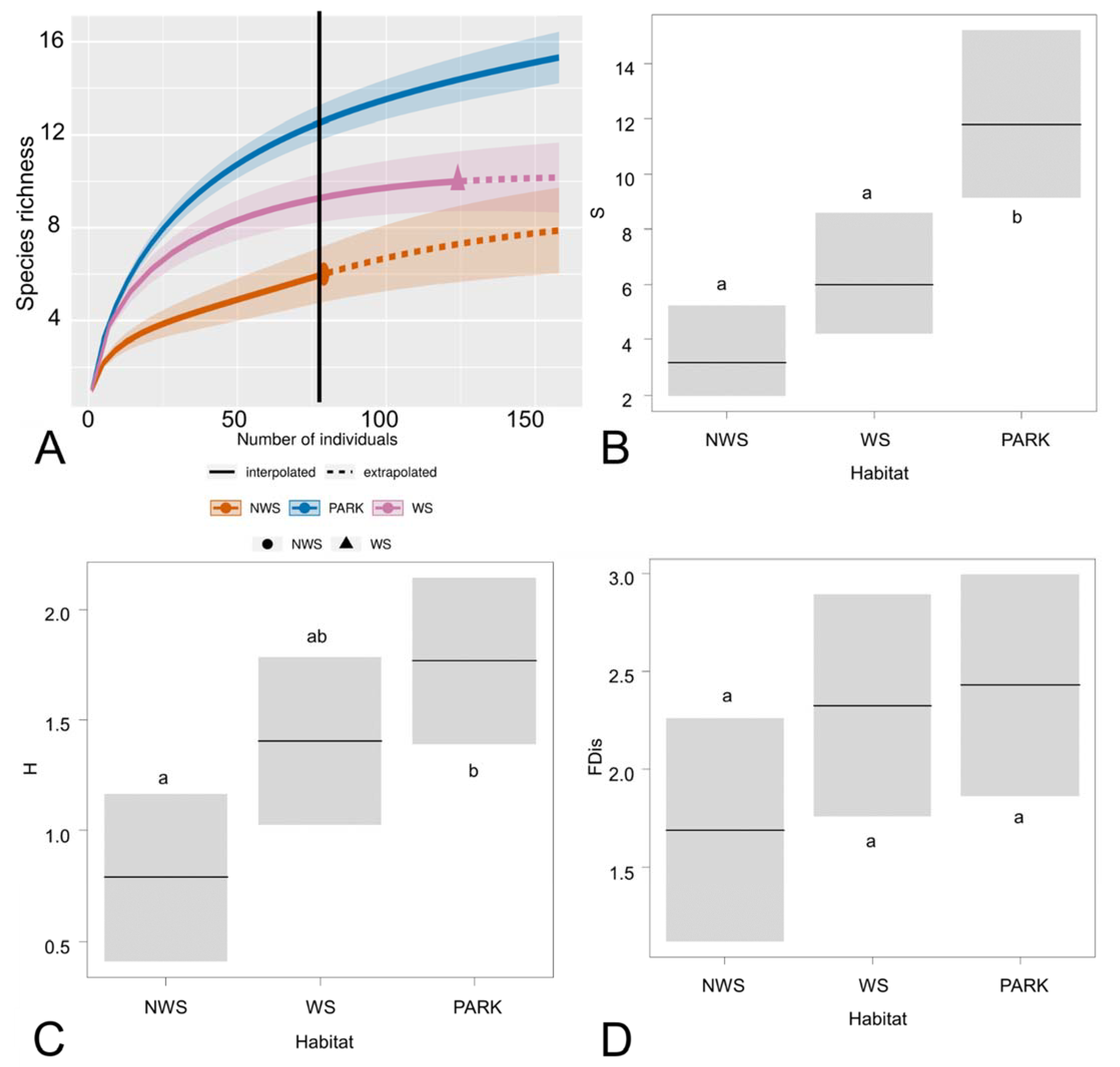
Interpolation analysis showed significant differences (p < 0.05) of species richness between habitat types (Figure 2A). Parks had the highest species richness, whereas wooded streets had an intermediate value, and the non-wooded streets had the lowest species richness (Figure 2A). Estimated species richness per transect showed that parks had more species richness than wooded and non-wooded streets (LRT = 27.32, degree of freedom (df) = 2, p = < 0.001; Table 1; Figure 2B). Parks had more species diversity (H’) than non-wooded streets (LRT = 2.44, df = 2, p = 0.001; Table 1; Figure 2C), and similar values between wooded streets and parks and between wooded and non-wooded streets. Functional dispersion index (FDis) values were similar between habitats (LRT = 1.59, df = 2, p = 0.149; Table 1; Figure 2D).
3.2. Taxonomic and Functional Traits Composition
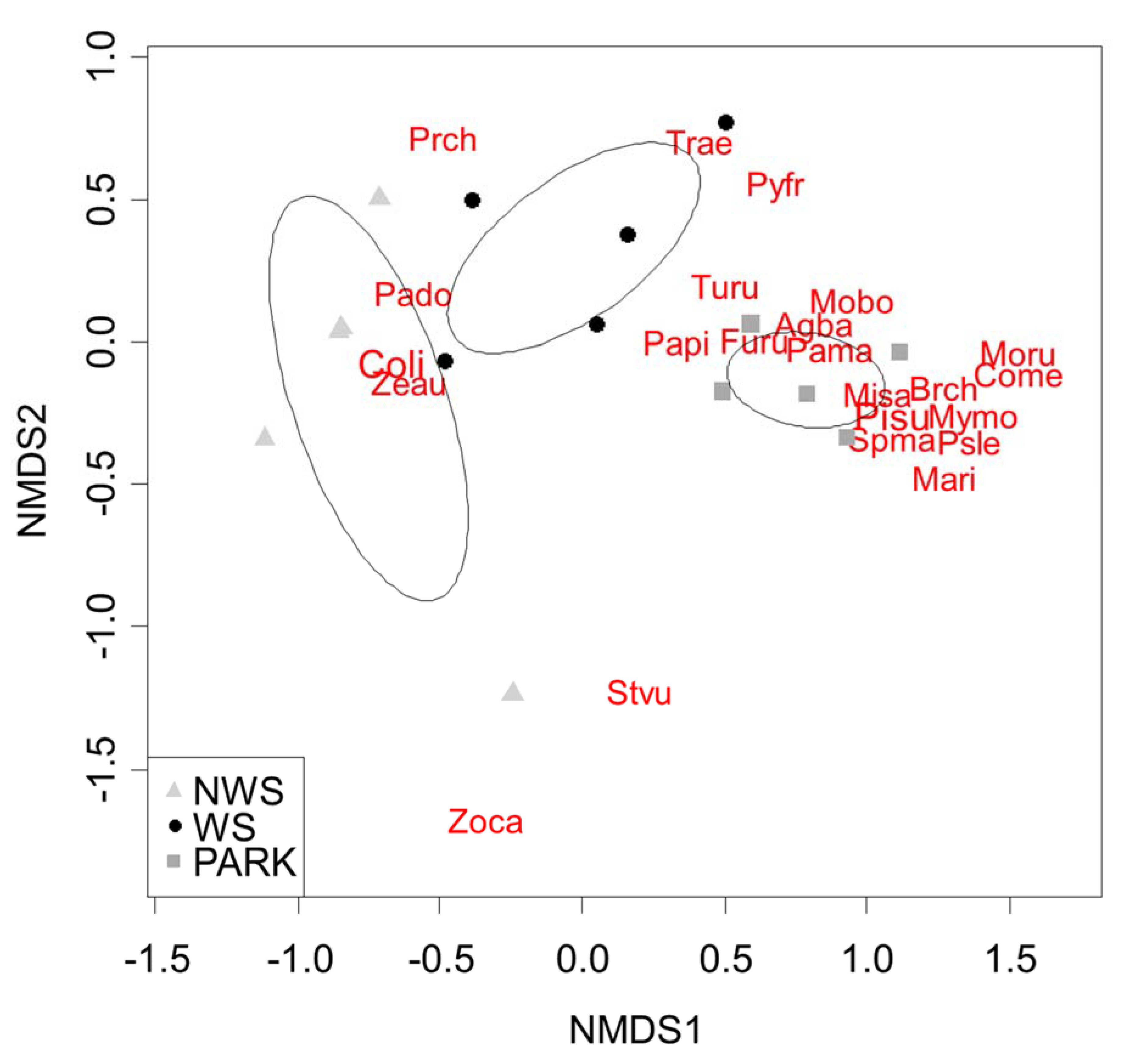
The adonis analysis showed significant differences in species composition between habitats (F model = 4.686, p = 0.001, r2 = 0.44). The ordiellipses confirmed that the three habitat types differed in terms of species composition (Figure 3). Parks had more abundance of the Grayish Baywing (Agelaioides badius), the Yellow-chevroned Parakeet (Brotogeris chiriri), the Green-barred Woodpecker, the Rufous Hornero, the Cattle Tyrant (Machetornis rixosa), the Chalk-browed Mockingbird, the Shiny Cowbird (Molothrus bonariensis), the Screaming Cowbird (Molothrus rufoaxillaris), the Monk Parakeet (Myiopsitta monachus), the Great Kiskadee (Pitangus sulphuratus), and the White-eyed Parakeet (Psittacara leucophthalma). The species associated with wooded streets were the Picazuro Pigeon, the Rufous-bellied Thrush, and the House Wren (Troglodytes aedon). The Rock Dove, the Eared Dove, the House Sparrow, and the Grey-breasted Martin (Progne chalybea), meanwhile, were related to non-wooded and wooded streets (Figure 3).
The similarity percentage analysis (simper) showed that the Rock Dove contributed the most to the dissimilarity of species among habitat types (Table 2). The Rock Dove had a greater average abundance in parks, followed by non-wooded streets, and then wooded streets. The House Sparrow had more abundance in wooded streets than in parks or non-wooded streets (Table 2). Native species like the Rufous Hornero, the Great Kiskadee, the Shiny Cowbird, and the Chalk-browed Mockingbird were found in parks and were absent in non-wooded streets.
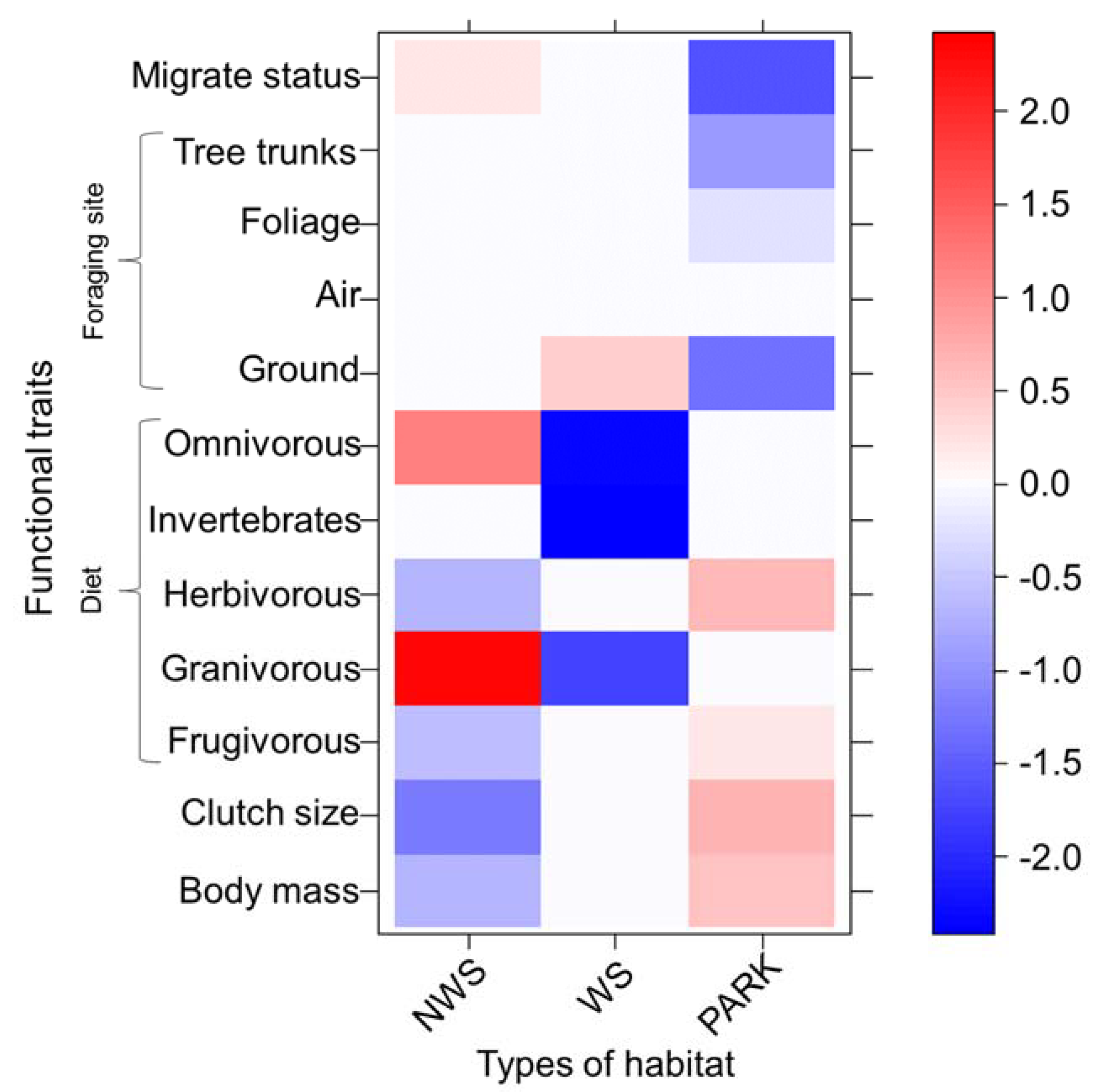
Migratory status was negatively related with parks (Figure 4). Tree trunk and ground foragers were negatively related with parks, whereas air foragers had no relation with any habitat type (Figure 4). Wooded streets were positively related with ground foragers. Omnivorous and granivorous birds were positively related with non-wooded streets, and they were negatively related with wooded streets (Figure 4). Frugivorous and herbivorous birds were positively related with parks and they were negatively related with non-wooded streets. Invertebrate feeders were strongly and negatively related with wooded streets. Bird species with greater clutch size and body mass were positively related with parks and the opposite pattern was found in non-wooded streets (Figure 4).
4. Discussion
Our results showed changes in taxonomic diversity and species and functional trait composition between habitat types. Taxonomic diversity had the greatest value in the parks, the wooded streets had an intermediate value, and the non-wooded streets had the lowest value. However, the sample size of the three habitats surveyed was small. Therefore, our results should be taken with caution.
The pattern found for taxonomic diversity agrees with [30], which obtained a similar pattern for species richness in Madrid, Spain. In our study the parks were composed of different vegetation strata (grass, shrubs, and trees), whereas the wooded streets were dominated by the arboreal stratum, and mainly composed of Ash (Fraxinus pennsylvanica) and Ficus (Ficus benjamina), an exotic tree species in Argentina. On the other hand, non-wooded streets have poor vegetation structures such as balcony pots or in some cases no vegetation at all. Several studies have found a positive relation between vegetation cover and bird species richness [65,66].
Rarefaction curves showed significant differences in species richness between habitat types. However, if we consider the mean estimated species richness of sampling units, wooded and non-wooded streets did not have significant differences between them. These patterns could be because the rarefaction curves preserve the heterogeneity of species composition between the transects. On the other hand, species diversity was similar between parks and wooded streets. This result could be related to the high abundance of the Rock Dove and Eared Dove in urban parks, which resulted in less even communities. Although the wooded streets had fewer species than urban parks, the former had more equitable abundances between species.
Contrary to expectations, the functional diversity did not differ significantly between habitat types. The index used, FDis, considers both the variety of traits and the abundance of the species. Therefore, the dominance of the Rock Dove in the parks could have influenced their functional diversity values. On the other hand, although in the non-wooded streets only six species were observed, the presence of the Gray-breasted Martin, the common starling (Sturnus vulgaris), and the Rufous-collared Sparrow (Zonotrichia capensis) could include novel functions, such as migration, air and foliage foraging sites, and insectivory and herbivory [62], thus increasing the FDis values.
The ordination analysis demonstrated that habitats had different species composition. The species that most contributed to the dissimilarity between habitats were the Rock Dove, the Eared Dove, the House Sparrow, the Picazuro Pigeon, and the Rufous-bellied Thrush. The Rock Dove, the Eared Dove, and the House Sparrow predominated in all habitats and a similar pattern was found by [67]. The Picazuro Pigeon and the Rufous-bellied Thrush were found in parks and wooded streets. These two species need trees to nest [41], and in the case of the Rufous-bellied Thrush, it can find insects in the trees. In this sense, it has been shown that trees are positively related with the number of arthropods [36]. The large number of granivores, such as the Rock Dove, the Eared Dove and the Picazuro Pigeon in parks and wooded streets, already recorded by [68] in squares, may be because the large portions of lawn or bare ground under leafy trees increase their foraging success [68]. On the other hand, in parks there was a greater abundance of species that nest in trees and feed on lawns, such as the Rufous Hornero, the Shiny Cowbird and the Chalk-browed Mockingbird.
The fourth corner analysis showed that trait composition varied between habitats. Migratory birds were negatively related to parks. The only migratory bird observed was the Gray-breasted Martin, which is related with high buildings in the urban center and usually nests in holes in buildings [35]. In this study, the Gray-breasted Martin occupied wooded and non-wooded streets, but other authors have found it in parks mostly surrounded by dense urbanization [69]. The positive association between the Grey-breasted Martin and streets may be due to the availability of nesting places. Most of the birds sampled were ground feeders. Ground feeders have been associated with urban environments [32] and the visibility provided by the asphalt could help these birds to find their food [35]. Omnivorous and granivorous birds were positively related with non-wooded streets and negatively related with wooded streets, traits related to the Rock Dove and the Eared Dove, respectively, the most abundant birds sampled and related to non-wooded streets in this study. Omnivorous and granivorous birds are the most common in urban areas [70]. Frugivorous, herbivorous, and invertebrate feeders were negatively related to non-wooded streets and positively related to parks, suggesting the importance of vegetated habitats for birds with these traits.
5. Conclusions
The results obtained show the importance of parks and wooded streets as they conserve several facets of bird diversity in a big neotropical city. Therefore, to improve the presence of birds in urban areas, the conservation of green areas such as parks and wooded streets and the afforestation of non-wooded streets are essential. This study should be a starting point for analyzing other aspects of the relationship between bird communities and urban habitats. For example, future studies should analyze the role of density-dependence in parks on wooded street use by birds [71], or the relation between urban park features and bird communities.
Author Contributions
L.M.L.: Conceptualization. F.E.C., L.M.L.: Methodology. F.E.C.: Software, data curation, formal analysis, writing—original draft preparation, visualization. F.E.C., L.M.L.: Investigation. L.M.L.: Supervision. F.E.C., L.M.L.: Writing—reviewing and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This study received no external founding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We thank the editor and two anonymous reviewers for the constructive comments that greatly improved the first draft of the manuscript. Paloma Garcia-Orza for improving the writing of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tisdale, H. The process of urbanization. Soc. Forces 1942, 20, 311–316. [Google Scholar] [CrossRef]
- Shen, L.; Peng, Y.; Zhang, X.; Wu, Y. An alternative model for evaluating sustainable urbanization. Cities 2012, 29, 32–39. [Google Scholar] [CrossRef]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef] [Green Version]
- McDonnell, M.J.; MacGregor-Fors, I. The ecological future of cities. Science 2016, 352, 936–938. [Google Scholar] [CrossRef] [PubMed]
- United Nations; Department of Economic and Social Affairs; Social Affairs, P.D. The World’s Cities in 2018; United Nation: New York, NY, USA, 2018. [Google Scholar]
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Clilliers, S.; Clarkson, B.D.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B Boil. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Becker, P.H. Biomonitoring with birds. In Trace Metals and Other Contaminants in the Environment; Elsevier: Amsterdam, The Netherlands, 2003; Volume 6, pp. 677–736. [Google Scholar]
- Faeth, S.H.; Bang, C.; Saari, S. Urban biodiversity: Patterns and mechanisms. Ann. N. Y. Acad. Sci. 2011, 1223, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, B.J.; FitzGibbon, S.I.; Wilson, R.S. New urban developments that retain more remnant trees have greater bird diversity. Landsc. Urban Plan. 2015, 136, 122–129. [Google Scholar] [CrossRef]
- Pena, J.C.D.C.; Martello, F.; Ribeiro, M.C.; Armitage, R.; Young, R.; Rodrigues, M. Street trees reduce the negative effects of urbanization on birds. PLoS ONE 2017, 12, e0174484. [Google Scholar] [CrossRef] [Green Version]
- Carbó-Ramírez, P.; Zuria, I. The value of small urban greenspaces for birds in a Mexican city. Landsc. Urban Plan. 2011, 100, 213–222. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Tucker, C.M. Difficult decisions: Strategies for conservation prioritization when taxonomic, phylogenetic and functional diversity are not spatially congruent. Biol. Conserv. 2018, 225, 128–133. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Wiley: Hoboken, NJ, USA, 2004; ISBN 0632056339. [Google Scholar]
- Díaz, S.; Cabido, M. Vive la différence: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Schütz, C.; Schulze, C.H. Functional diversity of urban bird communities: Effects of landscape composition, green space area and vegetation cover. Ecol. Evol. 2015, 5, 5230–5239. [Google Scholar] [CrossRef] [PubMed]
- La Sorte, F.A.; Lepczyk, C.A.; Aronson, M.F.J.; Goddard, M.A.; Hedblom, M.; Katti, M.; MacGregor-Fors, I.; Mörtberg, U.; Nilon, C.H.; Warren, P.S.; et al. The phylogenetic and functional diversity of regional breeding bird assemblages is reduced and constricted through urbanization. Divers. Distrib. 2018, 24, 928–938. [Google Scholar] [CrossRef] [Green Version]
- Leveau, L.M.; Isla, F.I.; Bellocq, M.I. From town to town: Predicting the taxonomic, functional and phylogenetic diversity of birds using NDVI. Ecol. Indic. 2020, 119, 106703. [Google Scholar] [CrossRef]
- Matuoka, M.A.; Benchimol, M.; de Almeida-Rocha, J.M.; Morante-Filho, J.C. Effects of anthropogenic disturbances on bird functional diversity: A global meta-analysis. Ecol. Indic. 2020, 116, 106471. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Su, T.; Zhou, B.; Moravec, D.; Šímová, P.; Liang, W. Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing’s urban parks: Effects of land use and vegetation structure. Urban For. Urban Green. 2017, 23, 84–92. [Google Scholar] [CrossRef]
- Hagen, E.O.; Hagen, O.; Ibáñez-Álamo, J.D.; Petchey, O.L.; Evans, K.L. Impacts of urban areas and their characteristics on avian functional diversity. Front. Ecol. Evol. 2017, 5, 84. [Google Scholar] [CrossRef] [Green Version]
- Taylor, L.; Hochuli, D.F. Defining greenspace: Multiple uses across multiple disciplines. Landsc. Urban Plan. 2017, 158, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.M.A. Urban green spaces and an integrative approach to sustainable environment. J. Environ. Prot. 2011, 2, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Karade, R.; Kuchi, V.; Kabir, J. The role of green space for sustainable landscape development in urban areas. Acta Hortic. 2017, 1181, 73–76. [Google Scholar] [CrossRef]
- Wolch, J.R.; Byrne, J.; Newell, J.P. Urban green space, public health, and environmental justice: The challenge of making cities ‘just green enough’. Landsc. Urban Plan. 2014, 125, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Byrne, J.; Pickering, C. A systematic quantitative review of urban tree benefits, costs, and assessment methods across cities in different climatic zones. Urban For. Urban Green. 2012, 11, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Bolund, P.; Hunhammar, S. Ecosystem services in urban areas. Ecol. Econ. 1999, 29, 293–301. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Jokimäki, J. A habitat island approach to conserving birds in urban landscapes: Case studies from southern and northern Europe. Biodivers. Conserv. 2001, 10, 2023–2043. [Google Scholar] [CrossRef]
- Fernández-Juricic, E. Avifaunal use of wooded streets in an urban landscape. Conserv. Biol. 2000, 14, 513–521. [Google Scholar] [CrossRef]
- White, J.G.; Antos, M.J.; Fitzsimons, J.; Palmer, G. Non-uniform bird assemblages in urban environments: The influence of streetscape vegetation. Landsc. Urban Plan. 2005, 71, 123–135. [Google Scholar] [CrossRef]
- Leveau, L.M. Bird traits in urban–rural gradients: How many functional groups are there? J. Ornithol. 2013, 154, 655–662. [Google Scholar] [CrossRef]
- Blair, R.B. Land use and avian species diversity along an urban gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar] [CrossRef]
- Rolando, A.; Maffei, G.; Pulcher, C.; Giuso, A. Avian community structure along an urbanization gradient. Ital. J. Zool. 1997, 64, 341–349. [Google Scholar] [CrossRef]
- Leveau, L.M.; Leveau, C.M. Comunidades de aves en un gradiente urbano de la ciudad de Mar del Plata, Argentina. Hornero 2004, 19, 13–21. [Google Scholar]
- Recher, H.F.; Majer, J.; Ganesh, S. Eucalypts, insects and birds: On the relationship between foliar nutrients and species richness. For. Ecol. Manag. 1996, 85, 177–195. [Google Scholar] [CrossRef]
- Leveau, L.M. Relaciones aves-habitat en el sector suburbano de Mar del Plata, Argentina. Ornitol. Neotrop. 2013, 24, 201–212. [Google Scholar]
- Instituto Nacional de Estadística y Censos (INDEC). Estadísticas Básicas. Total de Personas en el Hogar. Available online: https://redatam.indec.gob.ar/argbin/RpWebEngine.exe/PortalAction?&MODE=MAIN&BASE=CPV2010B&MAIN=WebServerMain.inl&_ga=2.160198014.2143654589.1623172429-797322923.1623172429 (accessed on 8 June 2021).
- Servicio Meteorologico Nacional. Valores Medios de Temperatura y Precipitación-Capital Federal. Available online: http://www3.smn.gov.ar/serviciosclimaticos/?mod=elclima&id=5&var=capitalfederal (accessed on 11 July 2019).
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.M.; Barberis, I.; Maturo, H.M.; Aragón, R.; Campanello, P.I.; Prado, D.; et al. Unidades de vegetación de la Argentina. Ecol. Austral. 2018, 28, 040–063. [Google Scholar] [CrossRef] [Green Version]
- de la Peña, M.R. Nidos y Reproducción de las Aves Argentinas; Ediciones Biológica: Santa Fe, Argentina, 2013. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Ma, K.H.; Hsieh, T.C. iNEXT (iNterpolation and EXTrapolation). Available online: https://chao.shinyapps.io/iNEXTOnline/ (accessed on 7 February 2016).
- Hines, J.E.; Boulinier, T.; Nichols, J.D.; Sauer, J.R.; Pollock, K.H. COMDYN: Software to study the dynamics of animal communities using a capture—Recapture approach. Bird Study 1999, 46, S209–S217. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Payton, M.E. Contrasting diversity values: Statistical inferences based on overlapping confidence intervals. PLoS ONE 2013, 8, e56794. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package “Vegan”. Community Ecology Package. Available online: https://github.com/vegandevs/vegan (accessed on 8 June 2021).
- Huhtalo, H.; Järvinen, O. Quantitative composition of the urban bird community in Tornio, Northern Finland. Bird Study 1977, 24, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Clergeau, P.; Savard, J.-P.L.; Mennechez, G.; Falardeau, G. Bird abundance and diversity along an urban-rural gradient: A comparative study between two cities on different continents. Condor 1998, 100, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, R.; Marzluff, J.M. Importance of reserve size and landscape context to urban bird conservation. Conserv. Biol. 2004, 18, 733–745. [Google Scholar] [CrossRef]
- Sengupta, S.; Mondal, M.; Basu, P. Bird species assemblages across a rural urban gradient around Kolkata, India. Urban Ecosyst. 2014, 17, 585–596. [Google Scholar] [CrossRef]
- Panda, B.P.; Prusty, B.A.K.; Panda, B.; Pradhan, A.; Parida, S.P. Habitat heterogeneity influences avian feeding guild composi-tion in urban landscapes: Evidence from Bhubaneswar, India. Ecol. Process. 2021, 10, 31. [Google Scholar] [CrossRef]
- Laliberté, E.; Legendre, P.; Shipley, B. Package “FD”. Measuring Functional Diversity (FD) from Multiple Traits, and Other Tools for Functional Ecology. Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 8 June 2021).
- de la Peña, M.R. Aves Argentinas: Descripción, comportamiento, reproducción y distribución: Trogonidae a Furnariidae. In Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie); El Museo: Santa Fe, Argentina, 2016; Volume 20, pp. 1–620. [Google Scholar]
- de la Peña, M.R. Aves Argentinas: Descripción, comportamiento, reproducción y distribución: Tyrannidae a Turdidae. In Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie); El Museo: Santa Fe, Argentina, 2016; Volume 21, pp. 1–633. [Google Scholar]
- de la Peña, M.R. Aves Argentinas: Descripción, comportamiento, reproducción y distribución: Mimidae a Passeridae. In Comunicaciones del Museo Provincial de Ciencias Naturales “Florentino Ameghino” (Nueva Serie); El Museo: Santa Fe, Argentina, 2016; Volume 21, pp. 1–564. [Google Scholar]
- Lenth, R.; Buerkner, P.; Herve, M.; Love, J.; Riebl, H.; Singmann, H. Package “Emmeans”: Estimated Marginal Means, aka Least-Squares Means. Available online: https://github.com/rvlenth/emmeans (accessed on 8 June 2021).
- Rangel, T.F.; Diniz-Filho, J.A.F.; Bini, L.M. SAM: A comprehensive application for spatial analysis in macroecology. Ecography 2010, 33, 46–50. [Google Scholar] [CrossRef]
- Hair, J.F.; Anderson, R.E.; Tatham, R.L.; Black, W.C. Análisis Multivariante; Prentice Hall: Madrid, Spain, 1999; ISBN 9788578110796. [Google Scholar]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Zhou, D.; Kong, H.; Chu, L.M. Do avian communities vary with season in highly urbanized. Wilson J. Ornithol. 2014, 126, 69–80. [Google Scholar] [CrossRef]
- Brown, A.M.; Warton, D.I.; Andrew, N.; Binns, M.; Cassis, G.; Gibb, H. The fourth-corner solution—Using predictive models to understand how species traits interact with the environment. Methods Ecol. Evol. 2014, 5, 344–352. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Eddelbuettel, D.; Warton, D.; Byrnes, J.; Silva, S.; Niku, J.; Renner, I.; Wright, S.; Package “Mvabund”. Statistical Methods for Analysing Multivariate Abundance Data. 2020. Available online: https://rdrr.io/cran/mvabund/man/mvabund-package.html#heading-2 (accessed on 8 June 2021).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Development Core Team: Viena, Austria, 2016. [Google Scholar]
- Faggi, A.; Caula, S. “Green” or “Gray”? Infrastructure and bird ecology in urban Latin America. In Avian Ecology in Latin American Cityscapes; MacGregor-Fors, I., Escobar-Ibáñez, J.F., Eds.; Springer International Publishing: New York, NY, USA, 2017; pp. 79–96. [Google Scholar]
- MacGregor-Fors, I.; Schondube, J.E. Gray vs. green urbanization: Relative importance of urban features for urban bird communities. Basic Appl. Ecol. 2011, 12, 372–381. [Google Scholar] [CrossRef]
- Leveau, C.M.; Leveau, L.M. Ensambles de aves en calles arboladas de tres ciudades del sudeste de la provincia de Buenos Aires, Argentina. Hornero 2006, 21, 25–30. [Google Scholar]
- Perepelizin, P.V.; Faggi, A.M. Diversidad de aves en tres barrios de la ciudad de Buenos Aires, Argentina. Multequina 2009, 18, 71–85. [Google Scholar]
- Faggi, A.; Perepelizin, P.V. Riqueza de aves a lo largo de un gradiente de urbanización en la ciudad de Buenos Aires. Rev. Mus. Argent. Cienc. Nat. Nueva Ser. 2006, 8, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E. Density-dependent habitat selection of corridors in a fragmented landscape. IBIS 2001, 143, 278–287. [Google Scholar] [CrossRef]
Figure 1.
Location of the transects within Buenos Aires city, Argentina.

Figure 2.
Bird taxonomic and functional diversity in different urban habitats. (A) Rarefaction curves estimated by the iNEXT software. The vertical black line in the rarefaction curve indicates the minimum number of individuals that we used to compare species richness between habitat types, and it corresponds to the greatest interpolated species richness of NWS. (B) GLM plot of species richness (S) estimated by COMDYN. (C) GLM plot of species diversity (H′). (D) GLM plot of Functional dispersion index (FDis). NWS = Non-Wooded Streets, WS = Wooded Streets. In (B–D), the horizontal black line represents the average value of the response variable for each habitat type and the grey bands are 95% confidence intervals. The different letters (a,b) indicate significant differences between the habitat types (p < 0.05).
Figure 2.
Bird taxonomic and functional diversity in different urban habitats. (A) Rarefaction curves estimated by the iNEXT software. The vertical black line in the rarefaction curve indicates the minimum number of individuals that we used to compare species richness between habitat types, and it corresponds to the greatest interpolated species richness of NWS. (B) GLM plot of species richness (S) estimated by COMDYN. (C) GLM plot of species diversity (H′). (D) GLM plot of Functional dispersion index (FDis). NWS = Non-Wooded Streets, WS = Wooded Streets. In (B–D), the horizontal black line represents the average value of the response variable for each habitat type and the grey bands are 95% confidence intervals. The different letters (a,b) indicate significant differences between the habitat types (p < 0.05).

Figure 3.
Non-metric multidimensional scaling (NMDS) ordination based on species abundances within different habitat types (NWS = non-wooded streets; WS = wooded streets; and parks) in Buenos Aires city (stress = 0.095). Agba = Agelaioides badius; Brch = Brotogeris chiriri Coli = Columba livia; Come = Colaptes melanochloros; Furu = Furnarius rufus; Mari = Machitornis rixosa; Misa = Mimus saturninus; Mobo = Molothrus bonaeriensis; Moru = Molothrus rufoaxilaris; Mymo = Myiopsitta monachus; Pado = Passer domesticus, Pama = Patagioenas maculosa; Papi = Patagioenas picazuro; Pisu = Pitangus sulphuratus; Prch = Progne chalybea; Psle = Psittacara leucophthalma; Pyfr = Pyrrhura frontalis, Spma = Spinus magellanica; Stvu = Sturnus vulgaris; Trae = Troglodytes aedon; Turu = Turdus rufiventris; Zeau = Zenaida auriculata; Zoca = Zonotrichia capensis.
Figure 3.
Non-metric multidimensional scaling (NMDS) ordination based on species abundances within different habitat types (NWS = non-wooded streets; WS = wooded streets; and parks) in Buenos Aires city (stress = 0.095). Agba = Agelaioides badius; Brch = Brotogeris chiriri Coli = Columba livia; Come = Colaptes melanochloros; Furu = Furnarius rufus; Mari = Machitornis rixosa; Misa = Mimus saturninus; Mobo = Molothrus bonaeriensis; Moru = Molothrus rufoaxilaris; Mymo = Myiopsitta monachus; Pado = Passer domesticus, Pama = Patagioenas maculosa; Papi = Patagioenas picazuro; Pisu = Pitangus sulphuratus; Prch = Progne chalybea; Psle = Psittacara leucophthalma; Pyfr = Pyrrhura frontalis, Spma = Spinus magellanica; Stvu = Sturnus vulgaris; Trae = Troglodytes aedon; Turu = Turdus rufiventris; Zeau = Zenaida auriculata; Zoca = Zonotrichia capensis.

Figure 4.
Fourth corner solution results. Functional traits were: body mass; clutch size; diet (frugivorous (fruit feeders), granivorous (seed feeders), herbivorous (plant material feeders), invertebrates (invertebrate feeders), and omnivorous (those that feed of more than three diet items)); foraging site (tree trunks, foliage, air, and ground). The scale bar indicates the interaction coefficients and the strength of the association between traits and habitats. Red = positive relation. Blue = negative relation. NWS = non-wooded streets, WS = wooded streets.
Figure 4.
Fourth corner solution results. Functional traits were: body mass; clutch size; diet (frugivorous (fruit feeders), granivorous (seed feeders), herbivorous (plant material feeders), invertebrates (invertebrate feeders), and omnivorous (those that feed of more than three diet items)); foraging site (tree trunks, foliage, air, and ground). The scale bar indicates the interaction coefficients and the strength of the association between traits and habitats. Red = positive relation. Blue = negative relation. NWS = non-wooded streets, WS = wooded streets.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Generalized linear model results showing differences in species richness, Shannon–Wiener index (H’), and functional dispersion (FDis) between habitat types. Habitat types: non-wooded streets (Intercept), wooded streets (WS), and parks; z/t indicates z or t tests.
Table 1.
Generalized linear model results showing differences in species richness, Shannon–Wiener index (H’), and functional dispersion (FDis) between habitat types. Habitat types: non-wooded streets (Intercept), wooded streets (WS), and parks; z/t indicates z or t tests.
| Variable | Estimate | Std. Error | z/t | p Value |
|---|---|---|---|---|
| Species richness | ||||
| Intercept | 1.163 | 0.250 | 4.653 | <0.001 |
| WS | 0.629 | 0.310 | 2.031 | 0.042 |
| Parks | 1.305 | 0.282 | 4.630 | <0.001 |
| Species diversity | ||||
| Intercept | 0.788 | 0.193 | 4.082 | 0.002 |
| WS | 0.616 | 0.273 | 2.258 | 0.043 |
| Parks | 0.978 | 0.273 | 3.583 | 0.004 |
| Fdis | ||||
| Intercept | 1.692 | 0.289 | 5.855 | <0.001 |
| WS | 0.632 | 0.409 | 1.548 | 0.148 |
| Parks | 0.738 | 0.409 | 1.805 | 0.096 |
Table 2.
Similarity percentage analysis (simper) between habitat types, showing the species percentage contribution to the dissimilarity of species composition (% contribution), and the average abundance of those species with a large contribution to dissimilarity (see Methods).
Table 2.
Similarity percentage analysis (simper) between habitat types, showing the species percentage contribution to the dissimilarity of species composition (% contribution), and the average abundance of those species with a large contribution to dissimilarity (see Methods).
| Wooded Streets–Non-Wooded Streets | |||
| Species | % Contribution | Wooded Streets | Non-Wooded Streets |
| Rock Dove (Columba livia) | 28.13 | 7.8 | 11.0 |
| House Sparrow (Passer domesticus) | 12.11 | 2.6 | 1.2 |
| Picazuro Pigeon (Patagioenas picazuro) | 8.67 | 1.6 | 0.0 |
| Rufous-bellied Thrush (Turdus rufiventris) | 7.45 | 1.4 | 0.0 |
| Wooded streets–Parks | |||
| Species | % Contribution | Wooded streets | Parks |
| Rock Dove (Columba livia) | 36.41 | 7.8 | 42.0 |
| Eared Dove (Zenaida auriculata) | 18.32 | 9.0 | 16.0 |
| Picazuro Pigeon (Patagioneas picazuro) | 7.35 | 1.6 | 7.4 |
| House Sparrow (Passer domesticus) | 3.65 | 2.6 | 1.8 |
| Great Kiskadee (Pitangus sulphuratus) | 2.9 | 0.0 | 2.0 |
| Rufous-bellied Thrush (Turdus rufiventris) | 2.69 | 1.4 | 3.2 |
| Rufous Hornero (Furnarius rufus) | 2.61 | 0.8 | 2.8 |
| Shiny Cowbird (Molothrus bonariensis) | 2.48 | 0.2 | 2.0 |
| Chalk-browed Mockingbird (Mimus saturninus) | 2.42 | 0.0 | 1.8 |
| Non-wooded streets–Parks | |||
| Species | % Contribution | Non-wooded streets | Parks |
| Rock Dove (Columba livia) | 34.43 | 11.0 | 42.0 |
| Eared Dove (Zenaida auriculata) | 17.86 | 3.0 | 16.0 |
| Picazuro Pigeon (Patagioneas picazuro) | 9.44 | 0.0 | 7.4 |
| Rufous-bellied Thrush (Turdus rufiventris) | 4.65 | 0.0 | 3.2 |
| Rufous Hornero (Furnarius rufus) | 3.7 | 0.0 | 2.8 |
| Great Kiskadee (Pitangus sulphuratus) | 2.85 | 0.0 | 2.0 |
| House Sparrow (Passer domesticus) | 2.74 | 1.2 | 1.8 |
| Shiny Cowbird (Molothrus bonariensis) | 2.5 | 0.0 | 2.0 |
| Chalk-browed Mockingbird (Mimus saturninus) | 2.35 | 0.0 | 1.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Curzel, F.E.; Leveau, L.M. Bird Taxonomic and Functional Diversity in Three Habitats in Buenos Aires City, Argentina. Birds 2021, 2, 217-229. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020016
AMA Style
Curzel FE, Leveau LM. Bird Taxonomic and Functional Diversity in Three Habitats in Buenos Aires City, Argentina. Birds. 2021; 2(2):217-229. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020016
Chicago/Turabian StyleCurzel, Florencia E., and Lucas M. Leveau. 2021. "Bird Taxonomic and Functional Diversity in Three Habitats in Buenos Aires City, Argentina" Birds 2, no. 2: 217-229. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2020016