Hippocampal and Cerebellar Changes in Acute Restraint Stress and the Impact of Pretreatment with Ceftriaxone

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Study Design
2.2. Cognitive and Behavioral Evaluation
2.3. Biochemical Analyses
2.3.1. Quantitative Analysis of Gene Expression by Real-Time PCR
2.3.2. Complementary DNA (cDNA) Synthesis
2.3.3. Real-Time Quantitative PCR
2.3.4. Detection of GLT1 Protein by the Western Blot Technique (Using aV3 Western Workflow™ Complete System, Bio-Rad® Hercules, CA, USA):
2.4. Histopathological Examination:
2.5. Morphometric Analysis:
2.6. Transmission Electron Microscopy (TEM) Study:
2.7. Statistical Analysis:
3. Results
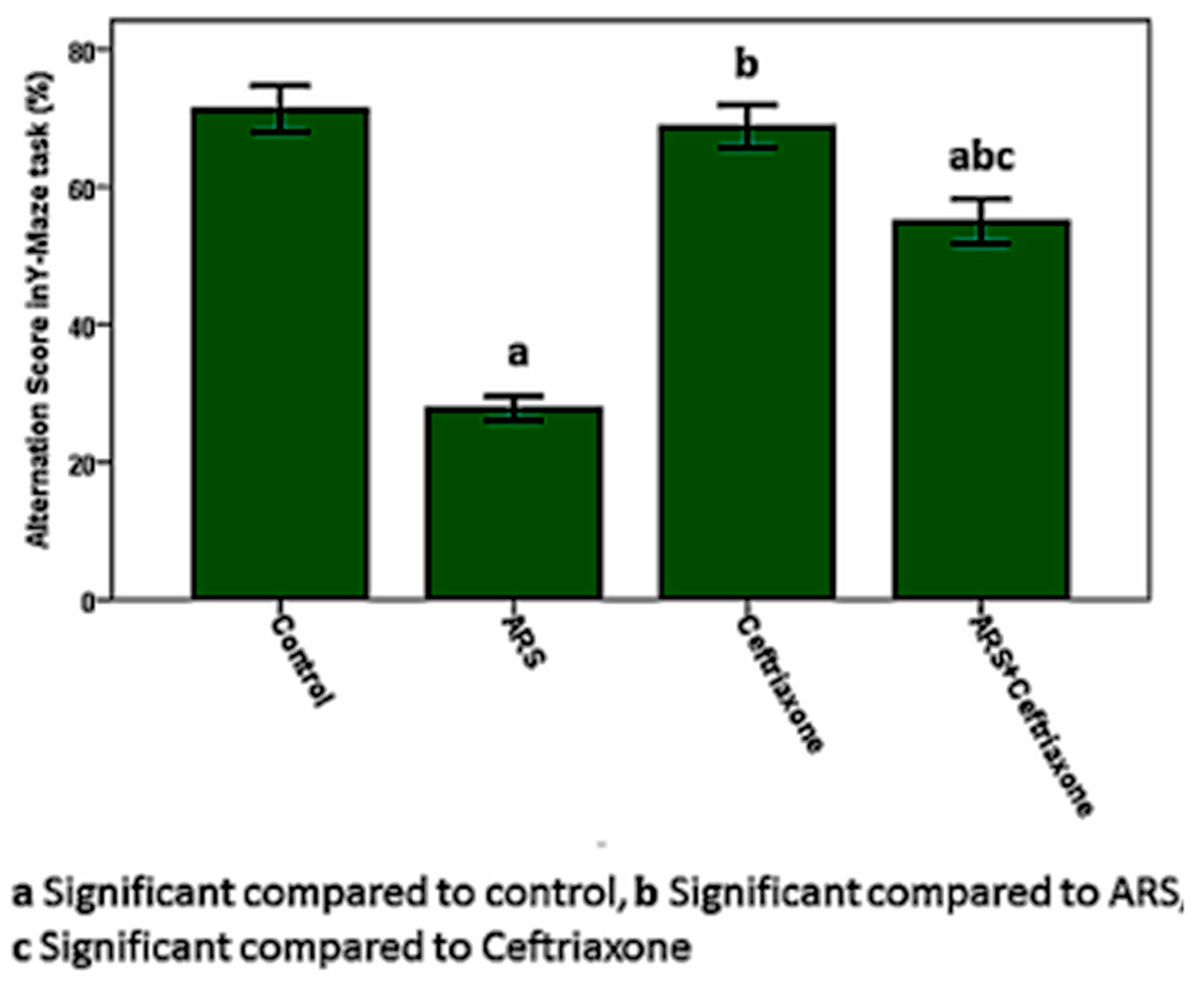
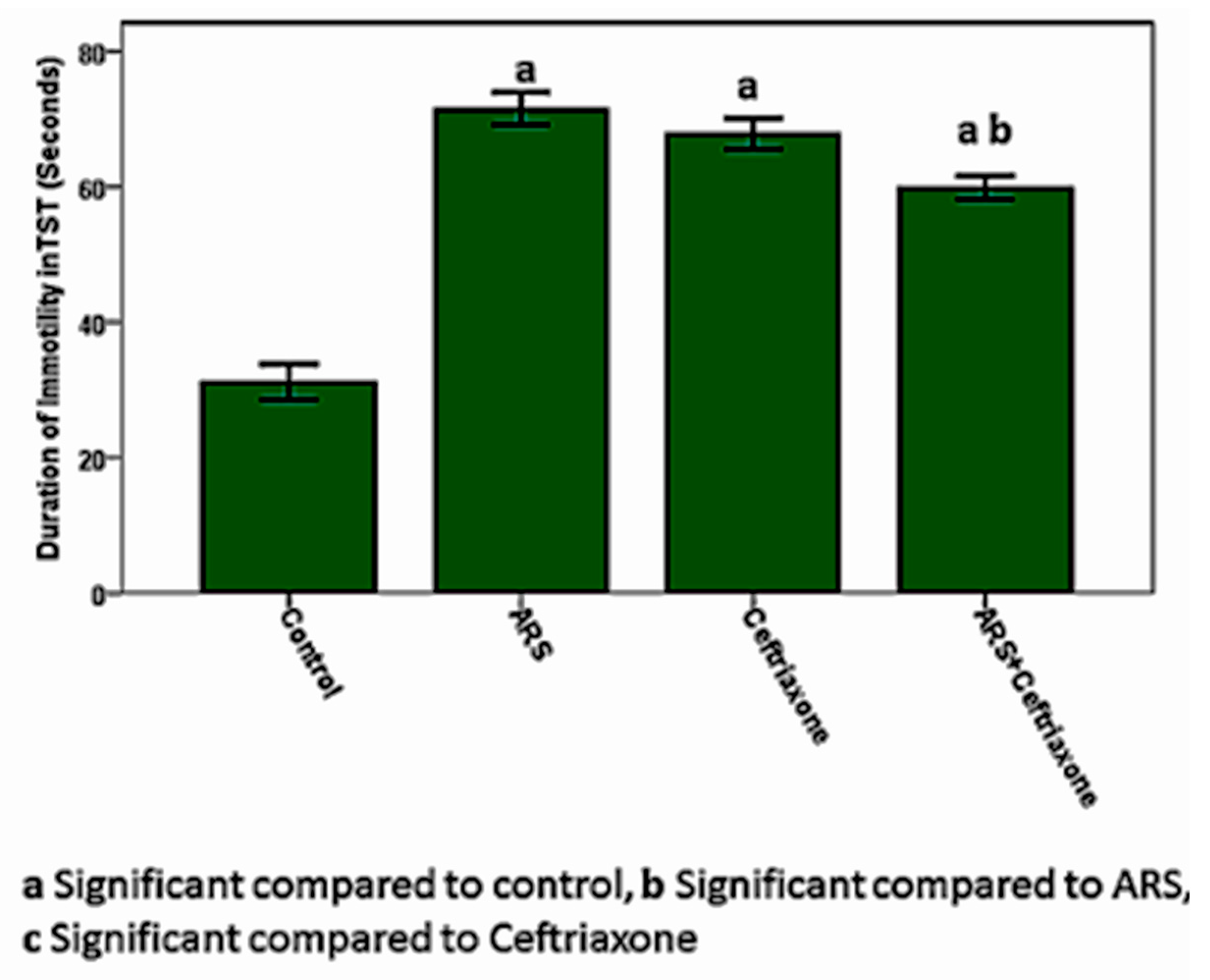
3.1. Cognitive and Behavioral Tests in the Study Groups
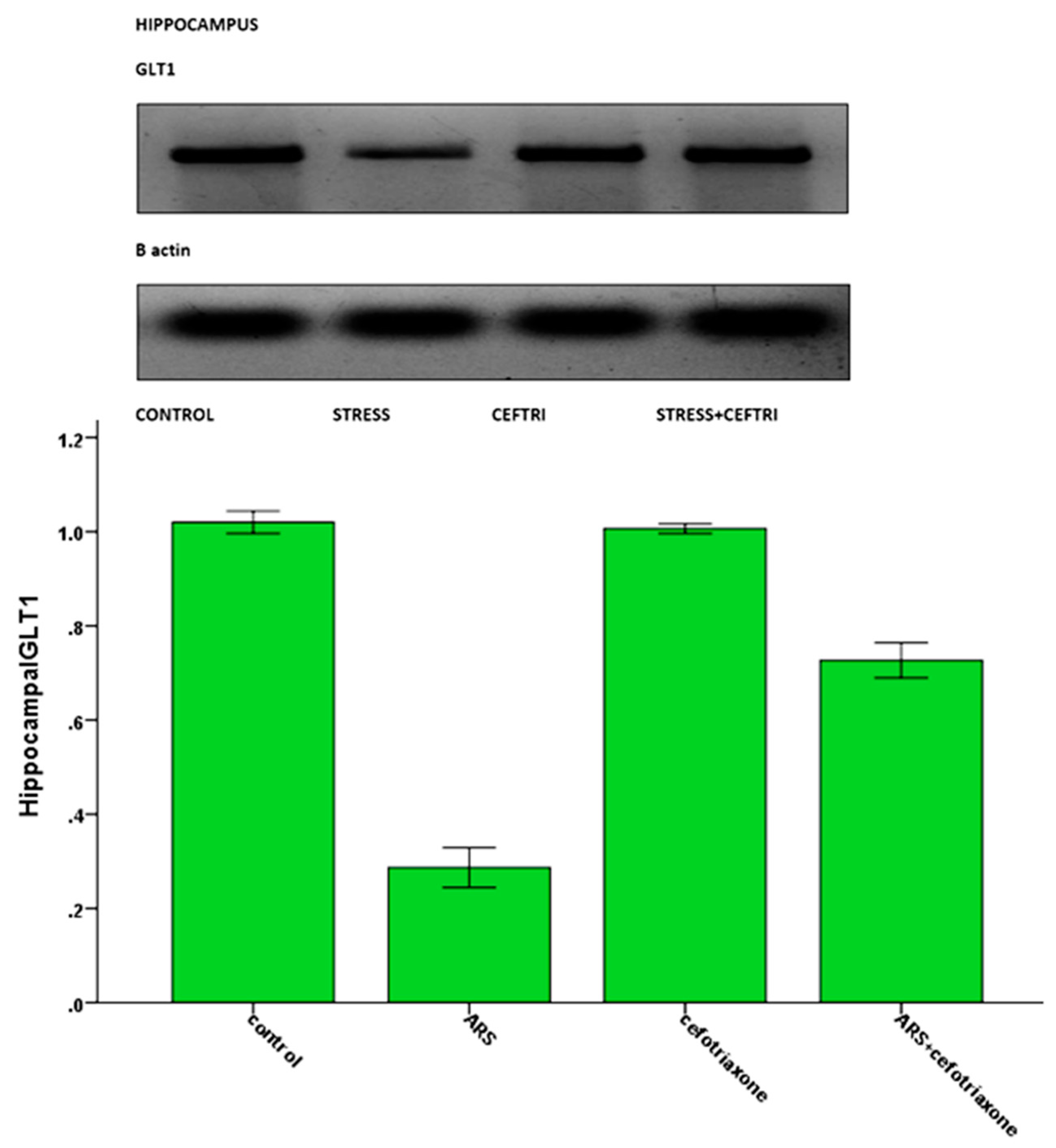
3.2. Results of Serum, Hippocampal, and Cerebellar Biochemical Analyses:
3.3. Histological and Ultrastructural Results:
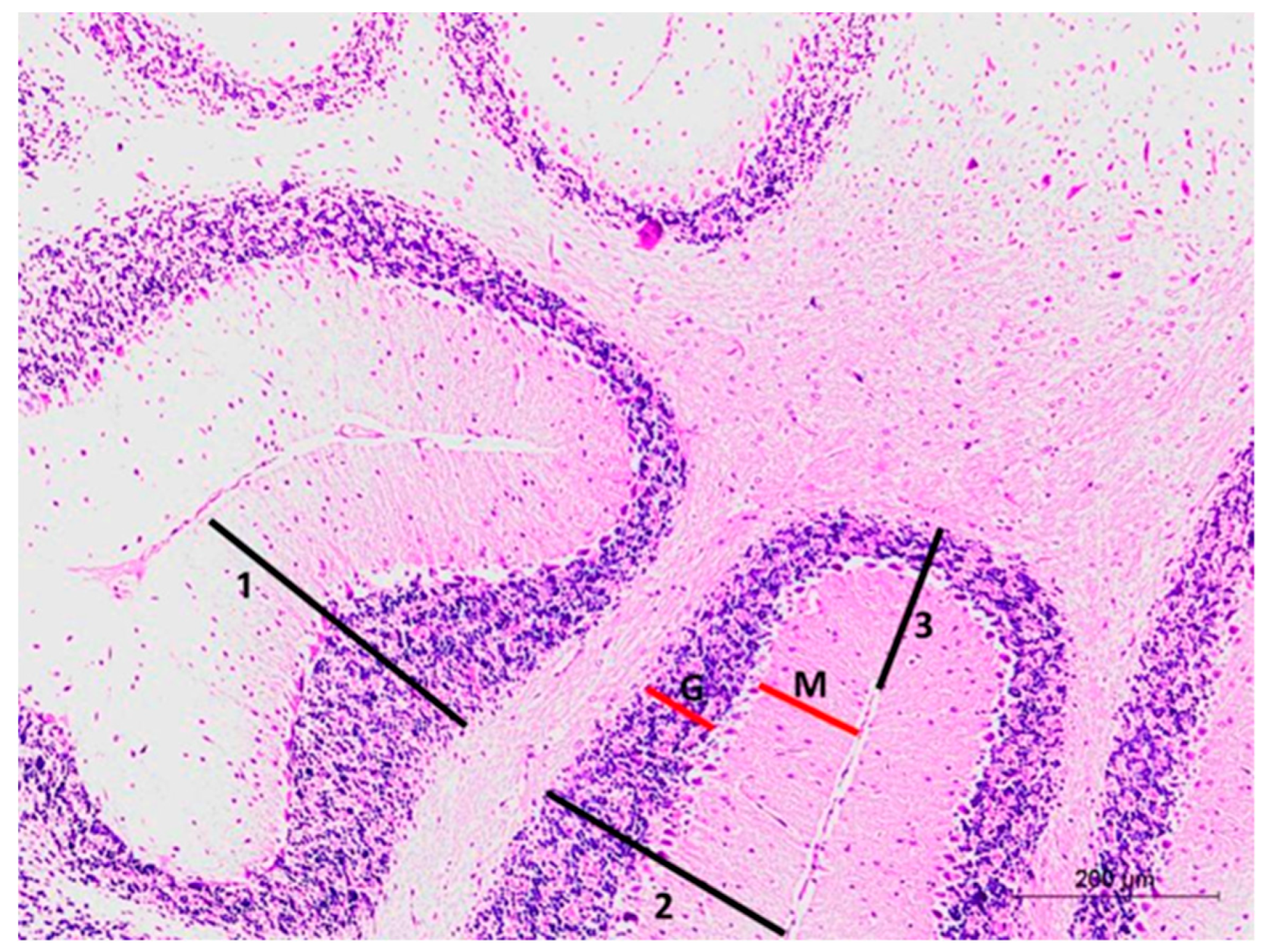
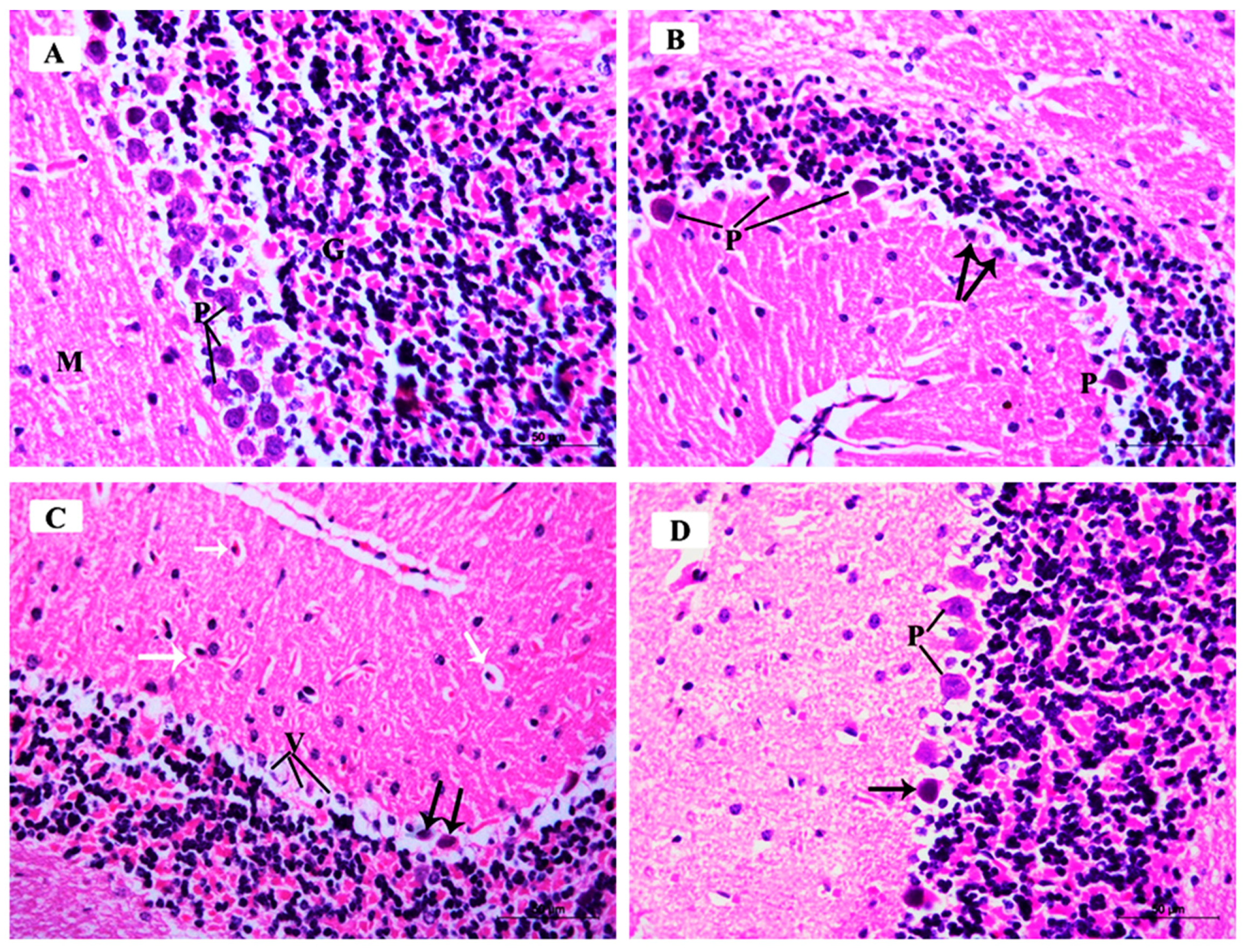
3.3.1. Histopathological Examination of the H&E-Stained Sections in the CA3 Region of the Hippocampus:
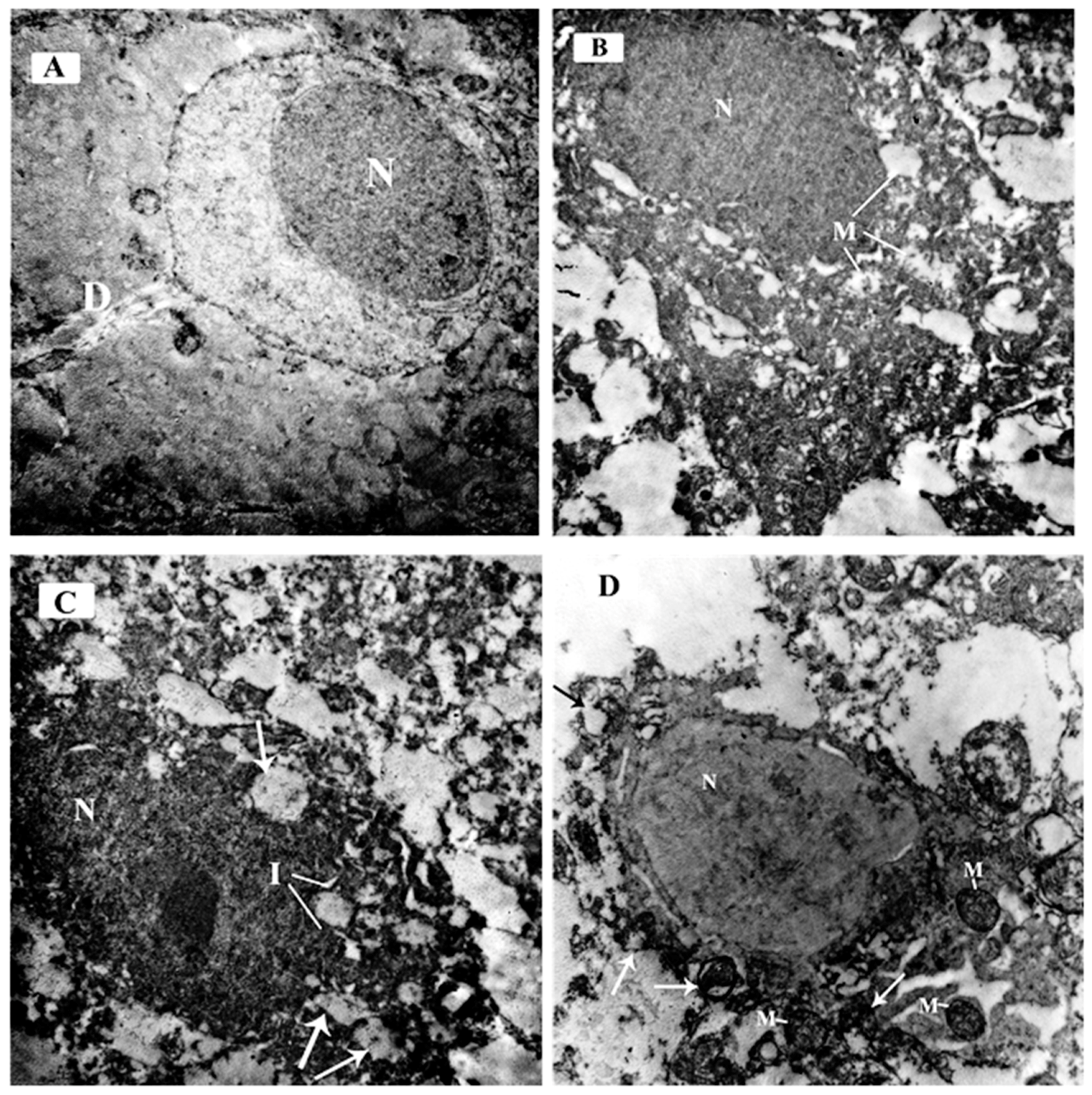
3.3.2. Ultrastructural Examination in the CA3 Region of the Hippocampus
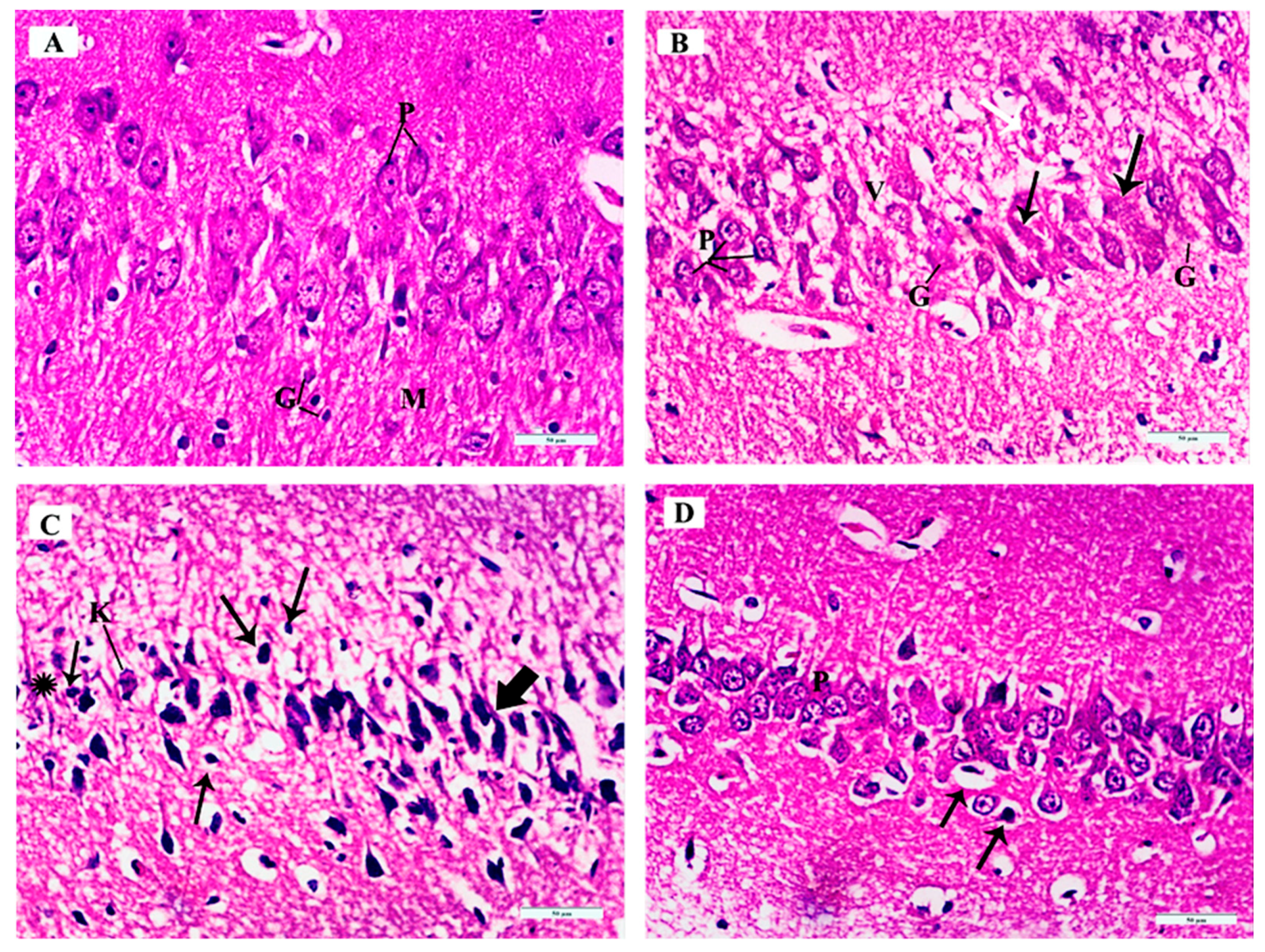
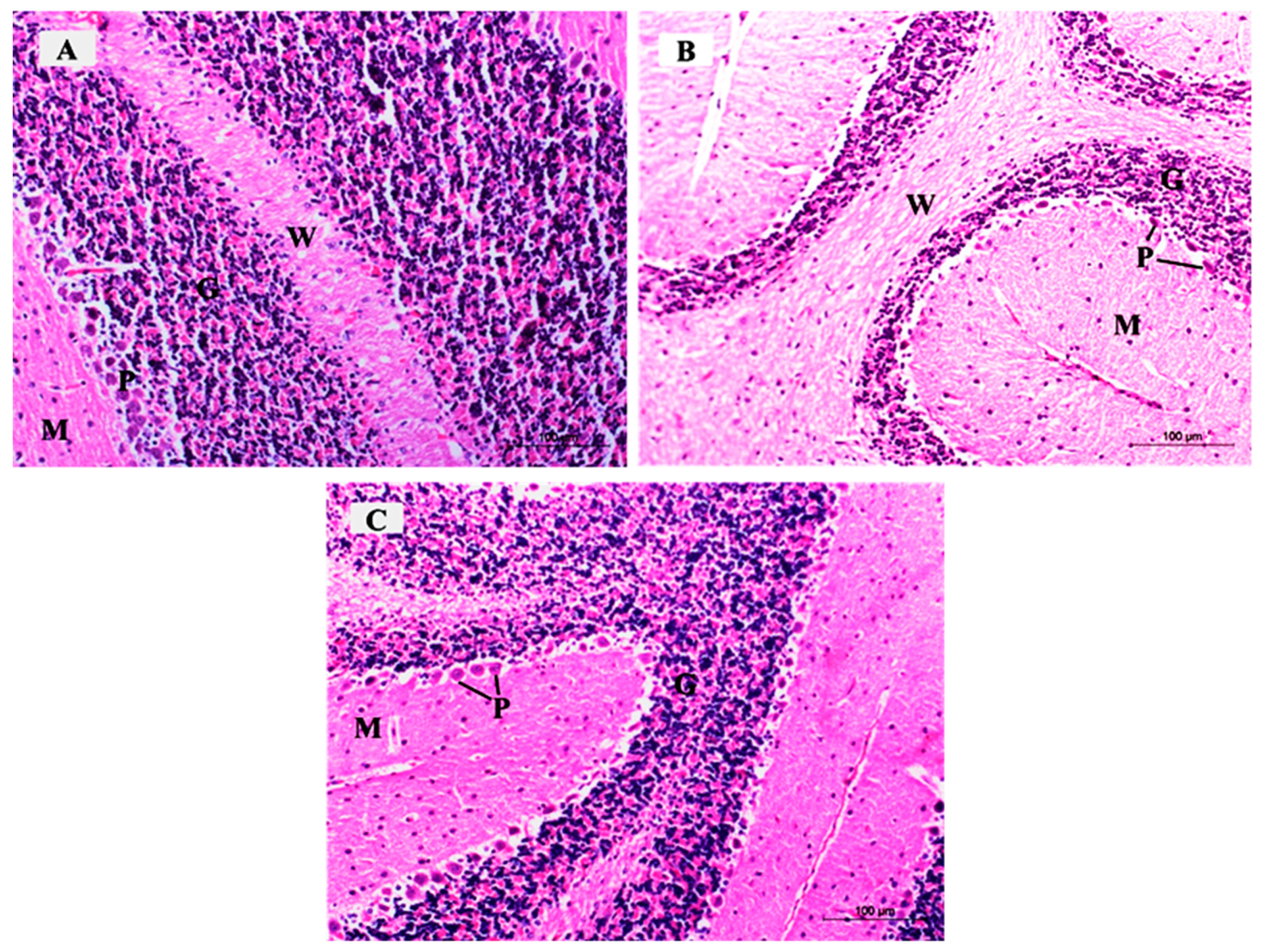
3.3.3. Histopathological Examination of H&E-Stained Sections in of the Cerebellar Cortex of Different Study Groups
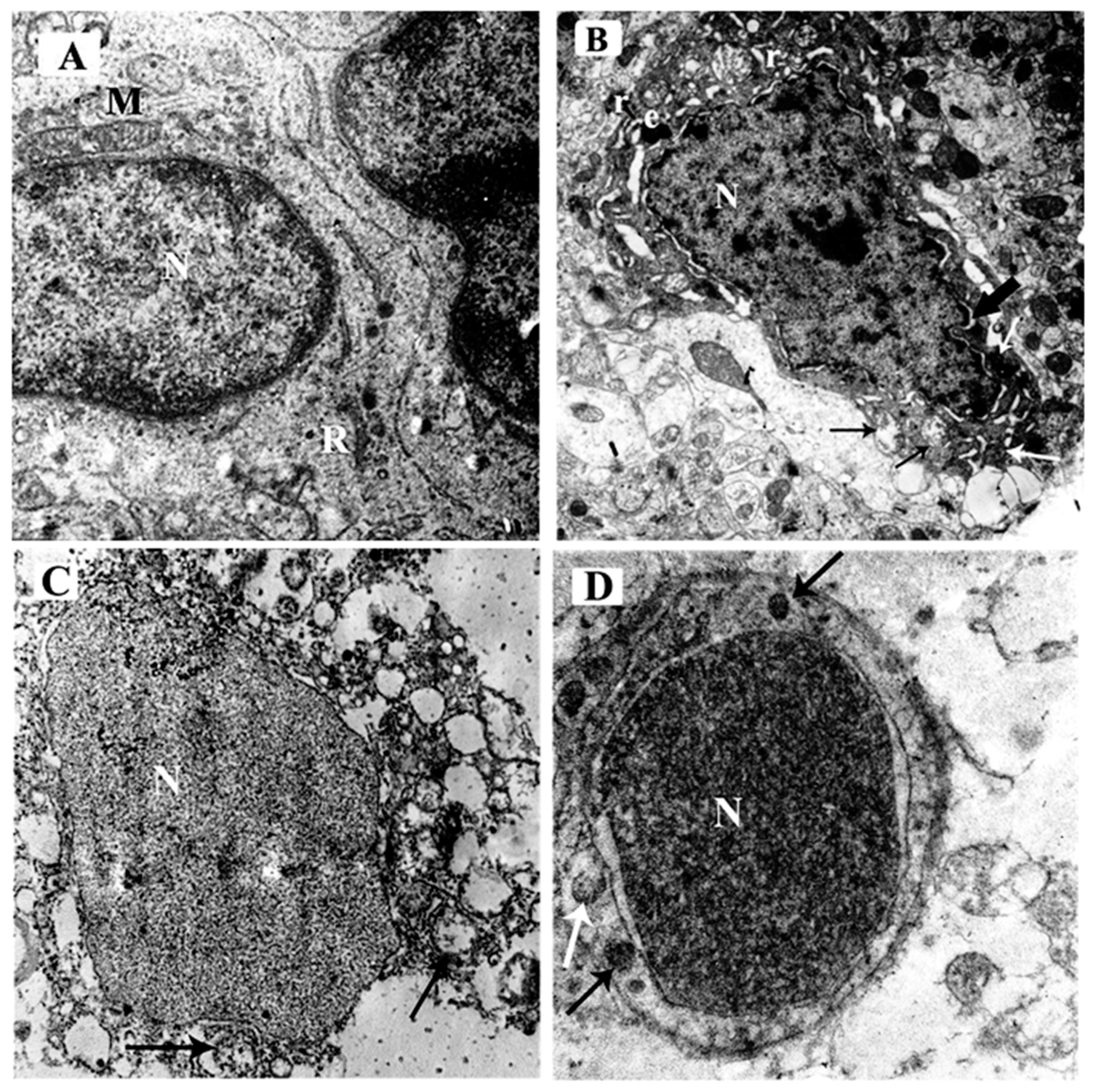
3.3.4. Ultrastructural Examination of the Sections in the Cerebellar Cortex of Different Study Groups at the Purkinje Cell Layer
3.4. Morphometric Results
3.4.1. Pyramidal Neurons Count in the CA3 Region of the Hippocampus
3.4.2. Thickness of Cerebellar Cortex Layers
3.4.3. Purkinje Cell Count
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alves, F.H.; Crestani, C.C.; Correa, F.M. The insular cortex modulates cardiovascular responses to acute restraint stress in rats. Brain Res. 2010, 1333, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, V.I. The role of the hippocampus in long-term memory: Is it memory store or comparator? J. Integr. Neurosci. 2008, 7, 117–184. [Google Scholar] [CrossRef]
- Amin, S.N.; Gamal, S.M.; Esmail, R.S.; Aziz, T.M.; Rashed, L.A. Cognitive effects of acute restraint stress in male albino rats and the impact of pretreatment with quetiapine versus ghrelin. J. Integr. Neurosci. 2014, 13, 669–692. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.N.; El-Aidi, A.A.; Ali, M.M.; Attia, Y.M.; Rashed, L.A. Modification of hippocampal markers of synaptic plasticity by memantine in animal models of acute and repeated restraint stress: Implications for memory and behavior. Neuromolecular Med. 2015, 17, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Hoche, F.; Guell, X.; Sherman, J.C.; Vangel, M.G.; Schmahmann, J.D. Cerebellar contribution to social cognition. Cerebellum 2016, 15, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Guell, X.; Gabrieli, J.D.E.; Schmahmann, J.D. Triple representation of language, working memory, social and emotion processing in the cerebellum: Convergent evidence from task and seed-based resting-state fMRI analyses in a single large cohort. Neuroimage 2018, 172, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Baldaçara, L.; Borgio, J.G.; Lacerda, A.L.; Jackowski, A.P. Cerebellum and psychiatric disorders. Rev. Bras. Psiquiatr. 2008, 30, 281–289. [Google Scholar] [CrossRef] [Green Version]
- Lipski, J.; Wan, C.K.; Bai, J.Z.; Pi, R.; Li, D.; Donnelly, D. Neuroprotective potential of ceftriaxone in in vitro models of stroke. Neuroscience 2007, 146, 617–629. [Google Scholar] [CrossRef]
- Zumkehr, J.; Rodriguez-Ortiz, C.J.; Cheng, D.; Kieu, Z.; Wai, T.; Hawkins, C.; Kilian, J.; Lim, S.L.; Medeiros, R.; Kitazawa, M. Ceftriaxone ameliorates tau pathology and cognitive decline via restoration of glial glutamate transporter in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2015, 36, 2260–2271. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.Y.; Hung, C.S.; Chang, H.M.; Liao, W.C.; Ho, S.C.; Ho, Y.J. Ceftriaxone prevents and reverses behavioral and neuronal deficits in an MPTPinduced animal model of Parkinson’s disease dementia. Neuropharmacology 2015, 91, 43–56. [Google Scholar] [CrossRef]
- Ahmad, A.; Rasheed, N.; Chand, K.; Maurya, R.; Banu, N.; Palit, G. Restraint stress-induced central monoaminergic & oxidative changes in rats & their prevention by novel Ocimum sanctum compounds. Indian J. Med. Res. 2012, 135, 548–554. [Google Scholar] [PubMed]
- Lai, P.C.; Huang, Y.T.; Wu, C.C.; Lai, C.-J.; Wang, P.J.; Chiu, T.H. Ceftriaxone attenuates hypoxic-ischemic brain injury in neonatal rats. J. Biomed. Sci. 2011, 18, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baykara, B.; Cetin, F.; Baykara, B.; Aksu, I.; Dayi, A.; Kiray, M.; Sisman, A.R.; Ozdemir, D.; Arda, M.N.; Uysal, N. Anxiety caused by traumatic brain injury correlates to decreased prefrontal cortex VEGF immunoreactivity and neuron density in immature rats. Turk. Neurosurg. 2012, 22, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Brocardo, P.S.; Budni, J.; Kaster, M.P.; Santos, A.R.S.; Rodrigues, A.L.S. Folic acid administration produces an antidepressant-like effect in mice: Evidence for the involvement of the serotonergic and noradrenergic = systems. Neuropharmacology 2008, 54, 464–473. [Google Scholar] [CrossRef]
- Arai, K.; Matsuki, N.; Ikegaya, Y.; Nishiyama, N. Deterioration of spatial learning performances in lipopolysaccharide-treated mice. Jpn. J. Pharmacol. 2001, 87, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Bancroft, J.D.; Gamble, M. (Eds.) Theory and Practice of Histological Techniques, 5th ed.; Churchill Livingstone: Edinburgh, Scotland, 2001; pp. 175–331. [Google Scholar]
- Grimaud, J.-A.; And, R. Borojevic: Intercellular formation of collagen in human liver. Cell. Mol. Biol. 1980, 26, 555–562. [Google Scholar]
- Altman, G.D. Comparing Groups: Three or More Independent Groups of Observations in Practical Statistics for Medical Research; Chapman and Hall: London, UK, 2005. [Google Scholar]
- Joëls, M.; Pu, Z.; Wiegert, O.; Oitzl, M.S.; Krugers, H.J. Learning under stress: How does it work? Trends Cogn. Sci. 2006, 10, 152–158. [Google Scholar] [CrossRef]
- Tanti, A.; Rainer, Q.; Minier, F.; Surget, A.; Belzung, C. Differential environmental regulation of neurogenesis along the septo-temporal axis of the hippocampus. Neuropharmacology 2012, 63, 374–384. [Google Scholar] [CrossRef]
- LaCrosse, A.L.; O’Donovan, S.M.; Sepulveda-Orengo, M.T.; McCullumsmith, R.E.; Reissner, K.J.; Schwendt, M.; Knackstedt, L.A. Contrasting the role of xCT and GLT-1 upregulation in the ability of ceftriaxone to attenuate the cue-induced reinstatement of cocaine-seeking and normalize AMPA receptor subunit expression. J. Neurosci. 2017, 37, 3716–3717. [Google Scholar] [CrossRef] [Green Version]
- Schmahmann, J.D.; Pandya, D.N. The cerebrocerebellar system. Int. Rev. Neurobiol. 1997, 41, 31–60. [Google Scholar]
- Wei, J.; Pan, X.; Pei, Z.; Wang, W.; Qiu, W.; Shi, Z.; Xiao, G. The beta-lactam antibiotic, ceftriaxone, provides neuroprotective potential via anti-excitotoxicity and anti-inflammation response in a rat model of traumatic brain injury. J. Trauma Acute Care Surg. 2012, 73, 654–660. [Google Scholar] [CrossRef] [PubMed]
- Sousa, N. The dynamics of the stress neuromatrix. Mol. Psychiatry 2016, 21, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Ren, J.; He, N.Y.; Liu, C.; Sun, Y.H.; Jian, F.F.; Bian, L.G.; Shen, J.K.; Yan, F.H.; Pan, S.J.; et al. Volumetric magnetic resonance imaging analysis in patients with short-term remission of Cushing’s disease. Clin. Endocrinol. 2017, 87, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.M.; Young, L.J.; Plotsky, P.M.; Insel, T.R. Distribution of corticosteroid receptors in the rhesus brain: Relative absence of glucocorticoid receptors in the hippocampal formation. J. Neurosci. 2000, 20, 4657–4668. [Google Scholar] [CrossRef] [Green Version]
- Wolf, O.T.; Bauser, D.S.; Daum, I. Eyeblink conditional discrimination learning in healthy young men is impaired after stress exposure. Psychophysiology 2012, 49, 164–171. [Google Scholar] [CrossRef]
- Brown, S.P.; Safo, P.K.; Regehr, W.G. Endocannabinoids inhibit transmission at granule cell to Purkinje cell synapses by modulating three types of presynaptic calcium channels. J. Neurosci. 2004, 24, 5623–5631. [Google Scholar] [CrossRef]
- Schmahmann, J.D. Dysmetria of thought: Clinical consequences of cerebellar dysfunction on cognition and affect. Trends Cogn. Sci. 1998, 2, 362–371. [Google Scholar] [CrossRef]
- Carageorgiou, H.; Pantos, C.; Zarros, A.; Stolakis, V.; Mourouzis, I.; Cokkinos, D.; Tsakiris, S. Changes in acetylcholinesterase, Na+,K+-ATPase, and Mg2+-ATPase activities in the frontal cortex and the hippocampus of hyper- and hypothyroid adult rats. Metabolism 2007, 56, 1104–1110. [Google Scholar] [CrossRef]
- Perry, E.; Walker, M.; Grace, J.; Perry, R. Acetylcholine in mind: A neurotransmitter correlate of consciousness? Trends Neurosci. 1999, 22, 273–280. [Google Scholar] [CrossRef]
- Rinaldo, L.; Hansel, C. Muscarinic acetylcholine receptor activation blocks long-term potentiation at cerebellar parallel Fiber-Purkinje cell synapses via cannabinoid signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 11181–11186. [Google Scholar] [CrossRef] [Green Version]
- Montalban, E.; Al-Massadi, O.; Sancho-Balsells, A.; Brito, V.; de Pins, B.; Alberch, J.; Ginés, S.; Girault, J.A.; Giralt, A. Pyk2 in the amygdala modulates chronic stress sequelae via PSD-95-related micro-structural changes. Transl. Psychiatry 2019, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borisova, T.; Krisanova, N.; Himmelreich, N. Artificial gravity and functional plasticity of nerve system. L-[14C]-glutamate uptake by nerve terminals from rat cerebellum and cerebral hemispheres under hypergravity stress. J. Gravit. Physiol. 2002, 9, P25–P26. [Google Scholar] [PubMed]
- Sood, A.; Chaudhari, K.; Vaidya, V.A. Acute stress evokes sexually dimorphic, stressor-specific patterns of neural activation across multiple limbic brain regions in adult rats. Stress 2018, 21, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Li, Y.; Ma, C.; Wang, C.; Sun, Z.; Shen, Y.; Liu, L.; Li, S.; Zhang, X.; Cong, B. Restraint stress induced hyperpermeability and damage of the blood-brain barrier in the amygdala of adult rats. Front. Mol. Neurosci. 2019, 12, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, D.M.; Bennett, M.C.; Fleshner, M.; Rose, G.M. Inverted-U relationship between the level of peripheral corticosterone and the magnitude of hippocampal primed burst potentiation. Hippocampus 1992, 2, 421–430. [Google Scholar] [CrossRef]
- Alfarez, D.N.; Wiegert, O.; Joels, M.; Krugers, H.J. Corticosterone and stress reduce synaptic potentiation in mouse hippocampal slices with mild stimulation. Neuroscience 2002, 115, 1119–1126. [Google Scholar] [CrossRef]
- Krugers, H.J.; Alfarez, D.N.; Karst, H.; Parashkouhi, K.; van Gemert, N.; Joëls, M. Corticosterone shifts different forms of synaptic potentiation in opposite directions. Hippocampus 2005, 15, 697–703. [Google Scholar] [CrossRef]
- Pais-Vieira, M.; Kunicki, C.; Peres, A.; Sousa, N. Ceftriaxone modulates the acute corticosterone effects in local field potentials in the primary somatosensory cortex of anesthetized mice. Sci. Rep. 2019, 9, 20289. [Google Scholar] [CrossRef] [Green Version]
- Krzyżanowska, W.; Pomierny, B.; Bystrowska, B.; Pomierny-Chamioło, L.; Filip, M.; Budziszewska, B.; Pera, J. Ceftriaxone-and N-acetylcysteine-induced brain tolerance to ischemia: Influence on glutamate levels in focal cerebral ischemia. PLoS ONE 2017, 12, e0186243. [Google Scholar] [CrossRef] [Green Version]
- Hussein, A.M.; Ghalwash, M.; Magdy, K.; Abulseoud, O.A. Beta lactams antibiotic ceftriaxone modulates seizures, oxidative stress and connexin 43 expression in hippocampus of pentylenetetrazole kindled rats. J. Epilepsy Res. 2016, 6, 8–15. [Google Scholar] [CrossRef]
- Matos-Ocasio, F.; Hernández-López, A.; Thompson, K.J. Ceftriaxone, a GLT-1 transporter activator, disrupts hippocampal learning in rats. Pharmacol. Biochem. Behav. 2014, 122, 118–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricceri, L.; Hohmann, C.; Berger-Sweeney, J. Early neonatal 192 IgG saporin induces learning impairments and disrupts cortical morphogenesis in rats. Brain Res. 2002, 954, 160–172. [Google Scholar] [CrossRef]
- Liu, A.; Niswander, L.A. Bone morphogenetic protein signalling and vertebrate nervous system development. Nat. Rev. Neurosci. 2005, 6, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Hu, Y. Bone morphogenetic protein 9 serves a protective role in response to ischemic-reperfusion in the brain by promoting ERK activation. Mol. Med. Rep. 2018, 17, 2845–2852. [Google Scholar] [CrossRef]
- Brooker, S.M.; Gobeske, K.T.; Chen, J.; Peng, C.Y.; Kessler, J.A. Hippocampal bone morphogenetic protein signaling mediates behavioral effects of antidepressant treatment. Mol. Psychiatry 2017, 22, 910–919. [Google Scholar] [CrossRef] [Green Version]
- Saftig, P.; Klumperman, J. Lysosome biogenesis and lysosomal membrane proteins: Trafficking meets function. Nat. Rev. Mol. Cell Biol. 2009, 10, 623–635. [Google Scholar] [CrossRef]
- Eskelinen, E. Roles of LAMP1 and LAMP-2 in lysosome biogenesis and autophagy. Mol. Asp. Med. 2006, 27, 495–502. [Google Scholar] [CrossRef]
- Nakashima, A.; Cheng, S.B.; Kusabiraki, T.; Motomura, K.; Aoki, A.; Ushijima, A.; Ono, Y.; Tsuda, S.; Shima, T.; Yoshino, O.; et al. Endoplasmic reticulum stress disrupts lysosomal homeostasis and induces blockade of autophagic flux in human trophoblasts. Sci. Rep. 2019, 9, 11466. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Zhou, Y.; Ma, C.; Li, S.; Cong, B. Endoplasmic reticulum stress is involved in restraint stress-induced hippocampal apoptosis and cognitive impairments in rats. Physiol. Behav. 2014, 131, 41–48. [Google Scholar] [CrossRef]
- Neckers, L.; Ivy, S.P. Heat shock protein 90. Curr. Opin. Oncol. 2003, 15, 419–424. [Google Scholar] [CrossRef]
- Grad, I.; Picard, D. The glucocorticoid responses are shaped by molecular chaperones. Mol. Cell. Endocrinol. 2007, 275, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Donato, R. S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 2001, 33, 637–668. [Google Scholar] [CrossRef]
- Mrak, R.E.; Griffinbc, W.S. The role of activated astrocytes and of the neurotrophic cytokine S100B in the pathogenesis of Alzheimer’s disease. Neurobiol. Aging 2001, 22, 915–922. [Google Scholar] [CrossRef]
- Hagmeyer, S.; Cristóvão, J.S.; Mulvihill, J.J.; Boeckers, T.M.; Gomes, C.M.; Grabrucker, A.M. Zinc binding to S100B affords regulation of trace metal homeostasis and excitotoxicity in the brain. Front. Mol. Neurosci. 2017, 10, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghandour, M.S.; Langley, O.K.; Vincendon, G.; Gombos, G.; Filippi, D.; Limozin, N.; Dalmasso, D.; Laurent, G. Immunochemical and immunohistochemical study of carbonic anhydrase II in adult rat cerebellum: A marker for oligodendrocytes. Neuroscience 1980, 5, 559–571. [Google Scholar] [CrossRef]
- Shi, C.; Uda, Y.; Dedic, C.; Azab, E.; Sun, N.; Hussein, A.I.; Petty, C.A.; Fulzele, K.; Mitterberger-Vogt, M.C.; Zwerschke, W.; et al. Carbonic anhydrase III protects osteocytes from oxidative stress. FASEB 2018, 32, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Mitoma, H.; Manto, M.; Hampe, C.S. Pathogenic roles of glutamic acid decarboxylase 65 autoantibodies in cerebellar ataxias. J. Immunol. Res. 2017, 2017, 2913297. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence | |
|---|---|
| BMP-9 | Forward: 5’-- TTCAGGATGAGGGCTGGGAG -3’ Reverse: 5’- GGATGTCTTCACAAGCACGGTC 3’ |
| Lamp1 | Forward: 5’- GTCCTCATCGTCCTCATTGC -3 Reverse: 5’- CTGATAGCCGGCGTGACT-3 |
| S100 | Forward: 5’ TGCTGTGGTTGGCATTTT TC 3’ Reverse: 5’ AGGCTGCGCAGCTTGGCCAT 3’ |
| CAII | Forward: 5’ TGCCCTCAGTTTGTGCAGAATA 3’ Reverse: 5’ CCAACGCAAGGAACTCTTCGA3’ |
| HSP90 | Forward:5’--GGTCATCTTGCTGTACGAAA -3 Reverse: 5′- GGTGGCATTTCTTCAGTTAC-3 |
| GLT- 1 | Forward: 5-GAAAAAACCCATTCTCCTTTTT-3 Reverse:5- CCGACTGGGAGGACGAATC-3 |
| GAD | Forward: 5-GAAAAAACCCATTCTCCTTTTT-3 Reverse: 5- CCGACTGGGAGGACGAATC-3 |
| Beta actin | Forward: 5’--GGTCGGTGTGAACGGATTTGG -3 Reverse: 5′- ATGTAGGCCATGAGGTCCACC-3 |
| Test | Control Group | ARS Group | Ceftriaxone Group | ARS + Ceftriaxone Group | |
|---|---|---|---|---|---|
| Open field | Number of crossed lines | 81.17 ± 2.483 | 19.17 ± 2.137 a | 70.50 ± 5.612 a,b | 82.83 ± 3.971 b,c |
| Central Square entry Frequency | 8.50 ± 1.049 | 1.33 ± 0.816 a | 7.83 ± 1.169 b | 7.17 ± 1.169 b | |
| Central Square Duration (Seconds) | 20.17 ± 1.941 | 3.33 ± 1.211 a | 16.83 ± 1.472 a,b | 12.67 ± 1.366 a,b,c | |
| Rearing Frequency | 5.17 ± 0.983 | 14.17 ± 1.722 a | 4.33 ± 1.751 b | 10.83 ± 1.472 a,b | |
| Grooming Frequency | 2.33 ± 0.816 | 9.00 ± 1.095 a | 2.00 ± 1.095 b | 4.33 ± 0.816 a,b,c | |
| Freezing Frequency | 2.00 ± 0.632 | 8.83 ± 1.169 a | 2.50 ± 0.548 b | 5.00 ± 1.095 a,b,c | |
| Y-Maze | total number of arm entries | 26 ± 2.832 | 14 ± 2.22 a | 24 ± 1.89 b | 19 ± 2.53 a,b,c |
| Alternation score (%) | 75 ± 3.410 | 28 ± 1.260 a | 73 ± 3.002 b | 56 ± 3.1 a,b,c |
| Control Group | ARS Group | Ceftriaxone Group | ARS + Ceftriaxone Group | |
|---|---|---|---|---|
| Serum TNF (pg/mL) | 14.433 ± 0.4131 | 112.367 ± 0.16.0814 a | 16.367 ± 0.2.9588 b | 52.700 ± 0.22.0782 a,b,c |
| Serum IL6 (pg/mL) | 21.400 ± 0.2280 | 95.267 ± 20.5213 a | 28.133 ± 2.0675 b | 60.70 ± 4.3964 a,b,c |
| Serum cortisol (ug/mL) | 4.1850 ± 0.22941 | 9.810 ± 0.80811 a | 3.6867 ± 0.39450 | 5.470 ± 0.43626 b,c |
| Hippocampal BMP9 (pg/mg protein) | 0.9833 ± 0.07528 | 0.483 ± 0.26838 a | 1.016 ± 0.01862 b | 0.8733 ± 0.09812 b,c |
| Hippocampal HSP90 (pg/mg protein) | 1.100 ± 0.12649 | 5.0067 ± 0.98392 a | 1.010 ± 0.00894 b | 1.800 ± 0.26833 b |
| Hippocampal LAMP1 (ng/mg protein) | 1.050 ± 0.19748 | 0.460 ± 0.26668 a | 1.0167 ± 0.02582 b | 0.880 ± 0.16757 b |
| Hippocampal GLT1(PCR) | 1.0067 ± 0.01 | 0.1867 ± 0.02 a | 1.0367 ± 0.03 b | 0.6233 ± 0.03 a,b,c |
| Hippocampal GLT1(WB) | 1.02 ± 0.023 | 0.2867 ± 0.042 a | 1.0067 ± 0.0103 b | 0.7267 ± 0.037 a,b,c |
| Cerebellar S100 B (pg/mg protein) | 6.133 ± 0.2251 | 16.133 ± 5.6811 a | 6.133 ± 0.7711 b | 7.633 ± 1.1911 b |
| Cerebellar Carbonic Anhydrase (ng/mg protein) | 2.3167 ± 0.37639 | 8.700 ± 1.35351 a | 2.6033 ± 0.32745 b | 5.300 ± 0.77974 a,b,c |
| Cerebellar GLT1 (PCR) | 1.0033 ± 0.005 | 0.3267 ± 0.0981 a | 1.0067 ± 0.0103 b | 0.7333 ± 0.0403 a,b,c |
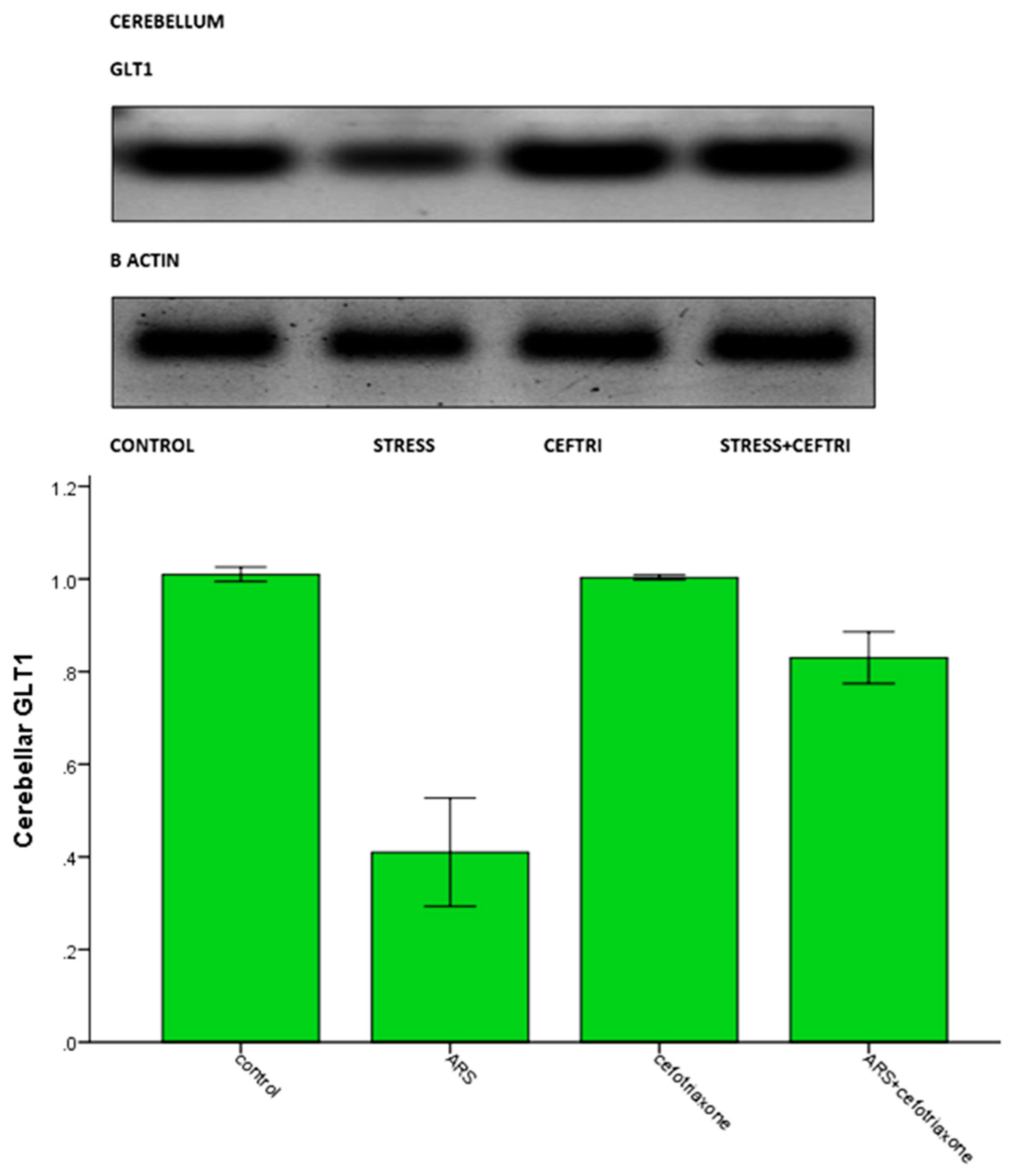
| Cerebellar GLT1 (WB) | 1.010 ± 0.0154 | 0.4100 ± 0.1169 a | 1.0033± 0.0051 b | 0.830 ± 0.0558 a,b,c |
| Cerebellar GAD (PCR) | 1.013 ± 0.0206 | 4.6000 ± 0.3224 a | 1.0133 ± 0.0206 b | 2.6367 ± 0.31879 a,b,c |
| Groups | Mean ± SD | Pairwise Comparisons with Other Groups | |
|---|---|---|---|
| Control | 41.83 ± 6.21 | ARS | p < 0.0015 |
| Cef. | p < 0.7194 | ||
| ARS+Cef | p < 0.1359 | ||
| ARS | 26.00 ± 6.42 | Control | p < 0.0015 |
| Cef. | p < 0.0027 | ||
| ARS+Cef | p < 0.0107 | ||
| Ceftriaxone | 40.50 ± 6.28 | Control | p < 0.7194 |
| ARS | p < 0.0027 | ||
| ARS+Cef | p < 0.2549 | ||
| ARS + Ceftriaxone | 36.50 ± 5.13 | Control | p < 0.1359 |
| ARS | p < 0.0107 | ||
| Cef. | p < 0.2549 | ||
| Groups | Mean ± SD | Pairwise Comparisons with Other Groups | Mean ± SD | Pairwise Comparisons with Other Groups | Mean ± SD | Pairwise Comparisons with Other Groups | |||
|---|---|---|---|---|---|---|---|---|---|
| At the Folium Surface | Facing the Fissure | At the Fissure Base | |||||||
| Control | 83.15± 5.77 | ARS | p < 0.0020 | 125.0 ± 5.49 | ARS | p < 0.0043 | 131.18 ± 5.99 | ARS | p < 0.0005 |
| Cef. | p < 0.9679 | Cef. | p < 0.6151 | Cef. | p < 0.8693 | ||||
| ARS+Cef | p < 0.4147 | ARS+Cef | p < 0.2207 | ARS+Cef | p < 0.1152 | ||||
| ARS | 98.74± 7.17 | Control | p < 0.0020 | 138.28 ± 6.90 | Control | p < 0.0043 | 147.88 ± 5.55 | Control | p < 0.0005 |
| Cef. | p < 0.0019 | Cef. | p < 0.0110 | Cef. | p < 0.0003 | ||||
| ARS+Cef | p < 0.0116 | ARS+Cef | p < 0.0298 | ARS+Cef | p < 0.0060 | ||||
| Cef. | 83.28± 5.57 | Control | p < 0.9679 | 126.76 ± 5.87 | Control | p < 0.6151 | 130.62 ± 5.42 | Control | p < 0.8693 |
| ARS | p < 0.0019 | ARS | p < 0.0110 | ARS | p < 0.0003 | ||||
| ARS+Cef | p < 0.4280 | ARS+Cef | p < 0.4755 | ARS+Cef | p < 0.1160 | ||||
| ARS + Ceftriaxone | 86.26± 6.85 | Control | p < 0.4147 | 129.18 ± 5.46 | Control | p < 0.2207 | 136.88 ± 5.44 | Control | p < 0.1152 |
| ARS | p < 0.0116 | ARS | p < 0.0298 | ARS | p < 0.0060 | ||||
| Ceftriaxone | p < 0.4280 | Ceftriaxone | p < 0.4755 | Ceftriaxone | p < 0.1160 | ||||
| Groups | Mean ± SD | Pairwise Comparisons with Other Groups | Mean ± SD | Pairwise Comparisons with Other Groups | Mean ± SD | Pairwise Comparisons with Other Groups | |||
|---|---|---|---|---|---|---|---|---|---|
| At the Folium Surface | Facing the Fissure | At the Fissure Base | |||||||
| Control | 122.67± 4.73 | ARS | p < 0.0002 | 99.20± 5.87 | ARS | p < 0.0004 | 73.20± 5.93 | ARS | p < 0.0005 |
| Cef. | p < 0.8941 | Cef. | p < 0.7739 | Cef. | p < 0.9376 | ||||
| ARS+Cef | p < 0.3302 | ARS+Cef | p < 0.2117 | ARS+Cef | p < 0.2181 | ||||
| ARS | 104.21± 6.28 | Control | p < 0.0002 | 82.20± 5.37 | Control | p < 0.0004 | 54.18± 7.14 | Control | p < 0.0005 |
| Cef. | p < 0.0001 | Cef. | p < 0.0001 | Cef. | p < 0.0002 | ||||
| ARS+Cef | p < 0.0009 | ARS+Cef | p < 0.0044 | ARS+Cef | p < 0.0046 | ||||
| Cef. | 123.01± 3.85 | Control | p < 0.8941 | 98.36± 3.68 | Control | p < 0.7739 | 73.44± 4.29 | Control | p < 0.9376 |
| ARS | p < 0.0001 | ARS | p < 0.0001 | ARS | p < 0.0002 | ||||
| ARS+Cef | p < 0.2434 | ARS+Cef | p < 0.2242 | ARS+Cef | p < 0.1499 | ||||
| ARS+ Cef | 119.72± 5.25 | Control | p < 0.3302 | 94.52± 6.27 | Control | p < 0.2117 | 68.48± 6.50 | Control | p < 0.2181 |
| ARS | p < 0.0009 | ARS | p < 0.0044 | ARS | p < 0.0046 | ||||
| Cef. | p < 0.2434 | Cef. | p < 0.2242 | Cef. | p < 0.1499 | ||||
| Groups | Mean ± SD | Pairwise Comparisons with Other Groups | |
|---|---|---|---|
| Control | 21.33 ± 2.80 | ARS | p < 0.0001 |
| Cef. | p < 0.4969 | ||
| ARS+Cef | p < 0.0658 | ||
| ARS | 10.83 ± 3.19 | Control | p < 0.0001 |
| Cef. | p < 0.0004 | ||
| ARS+Cef | p < 0.0031 | ||
| Ceftriaxone | 20.17 ± 2.93 | Control | p < 0.4969 |
| ARS | p < 0.0004 | ||
| ARS+Cef | p < 0.2069 | ||
| ARS + Ceftriaxone | 17.83 ± 3.06 | Control | p < 0.0658 |
| ARS | p < 0.0031 | ||
| Cef. | p < 0.2069 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amin, S.N.; Hassan, S.S.; Khashaba, A.S.; Youakim, M.F.; Latif, N.S.A.; Rashed, L.A.; Yassa, H.D. Hippocampal and Cerebellar Changes in Acute Restraint Stress and the Impact of Pretreatment with Ceftriaxone. Brain Sci. 2020, 10, 193. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10040193
Amin SN, Hassan SS, Khashaba AS, Youakim MF, Latif NSA, Rashed LA, Yassa HD. Hippocampal and Cerebellar Changes in Acute Restraint Stress and the Impact of Pretreatment with Ceftriaxone. Brain Sciences. 2020; 10(4):193. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10040193
Chicago/Turabian StyleAmin, Shaimaa N., Sherif S. Hassan, Ahmed S. Khashaba, Magdy F. Youakim, Noha S. Abdel Latif, Laila A. Rashed, and Hanan D. Yassa. 2020. "Hippocampal and Cerebellar Changes in Acute Restraint Stress and the Impact of Pretreatment with Ceftriaxone" Brain Sciences 10, no. 4: 193. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10040193