Effects of Juvenile or Adolescent Working Memory Experience and Inter-Alpha Inhibitor Protein Treatment after Neonatal Hypoxia-Ischemia

 , ,
, ,  and
and
Abstract
:1. Introduction
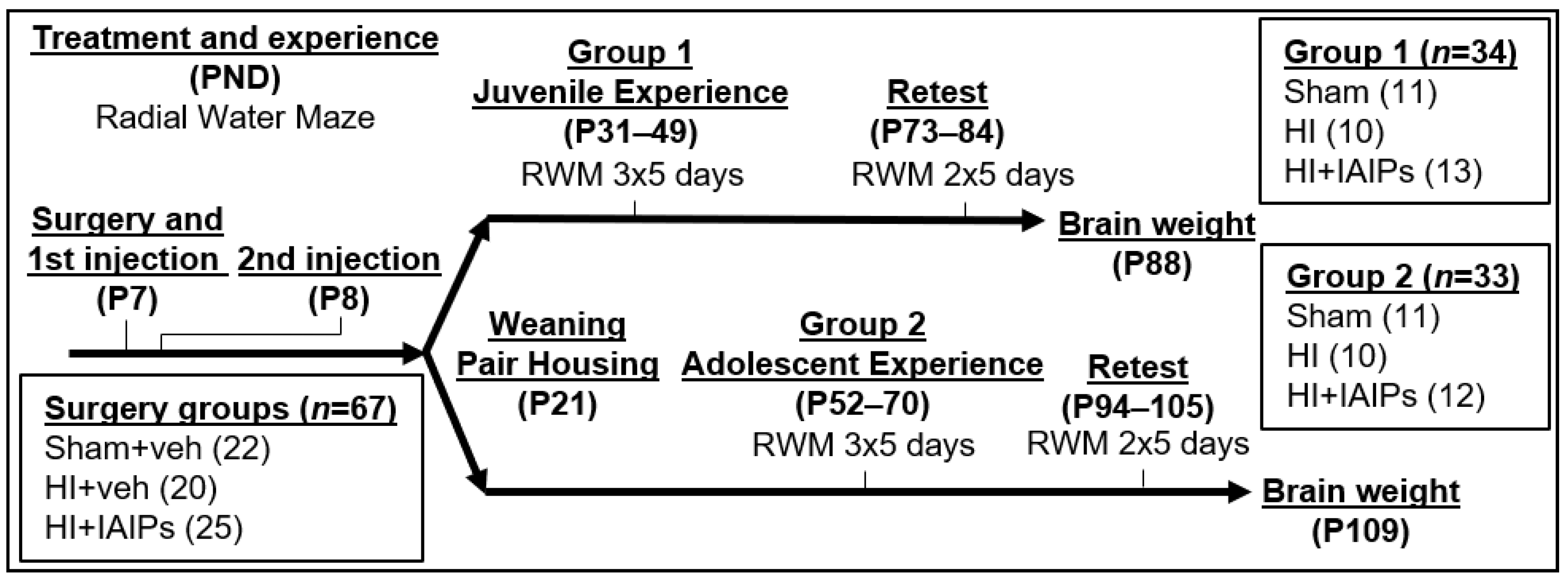
2. Materials and Methods
2.1. Animal Subjects
2.2. Surgery and Treatment
2.3. Eight Arm Radial Water Maze (RWM)
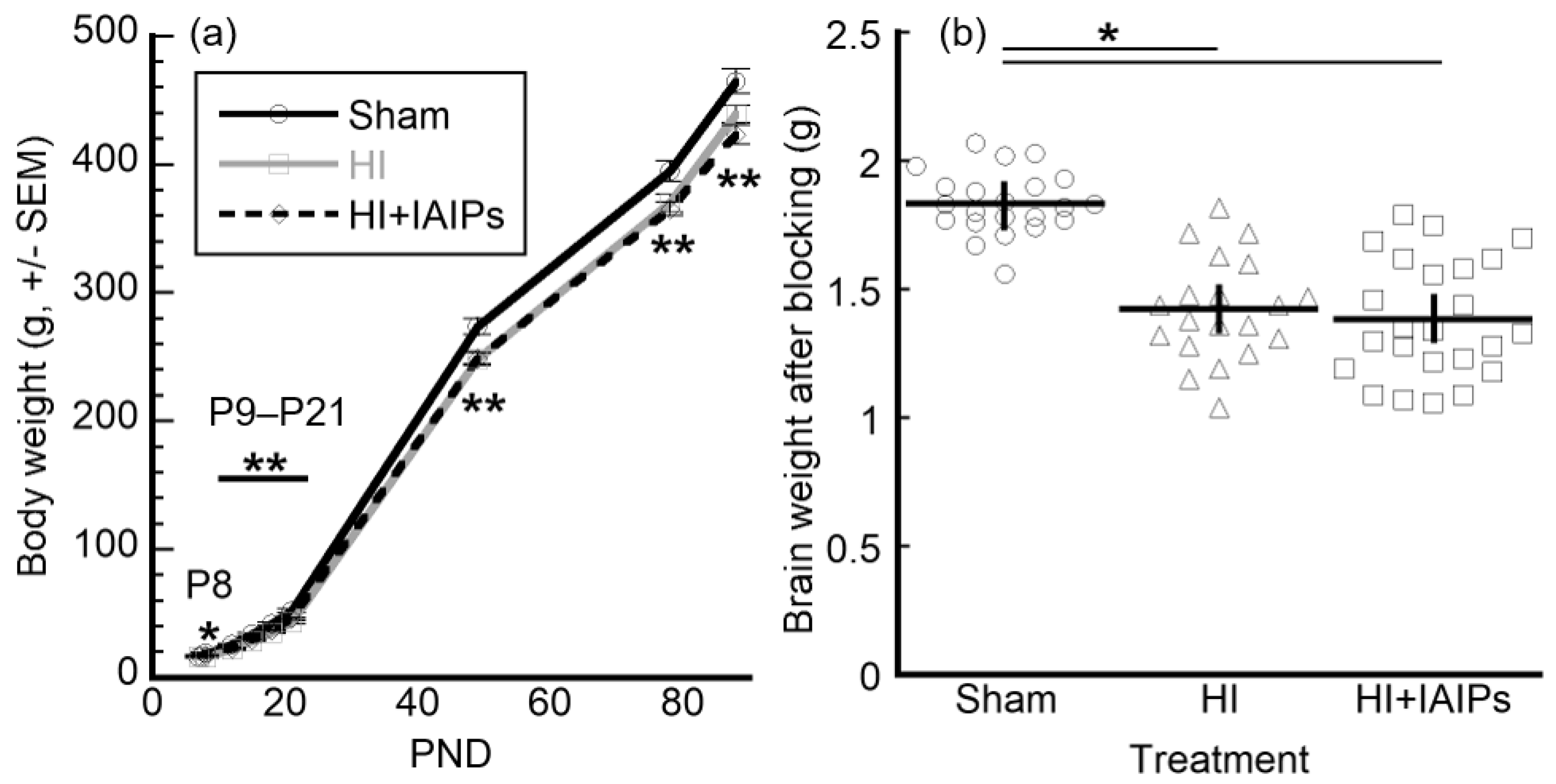
2.4. Body and Brain Weight
2.5. Statistics
3. Results
3.1. HI Exposure Resulted in Main Effects of Body and Brain Weight
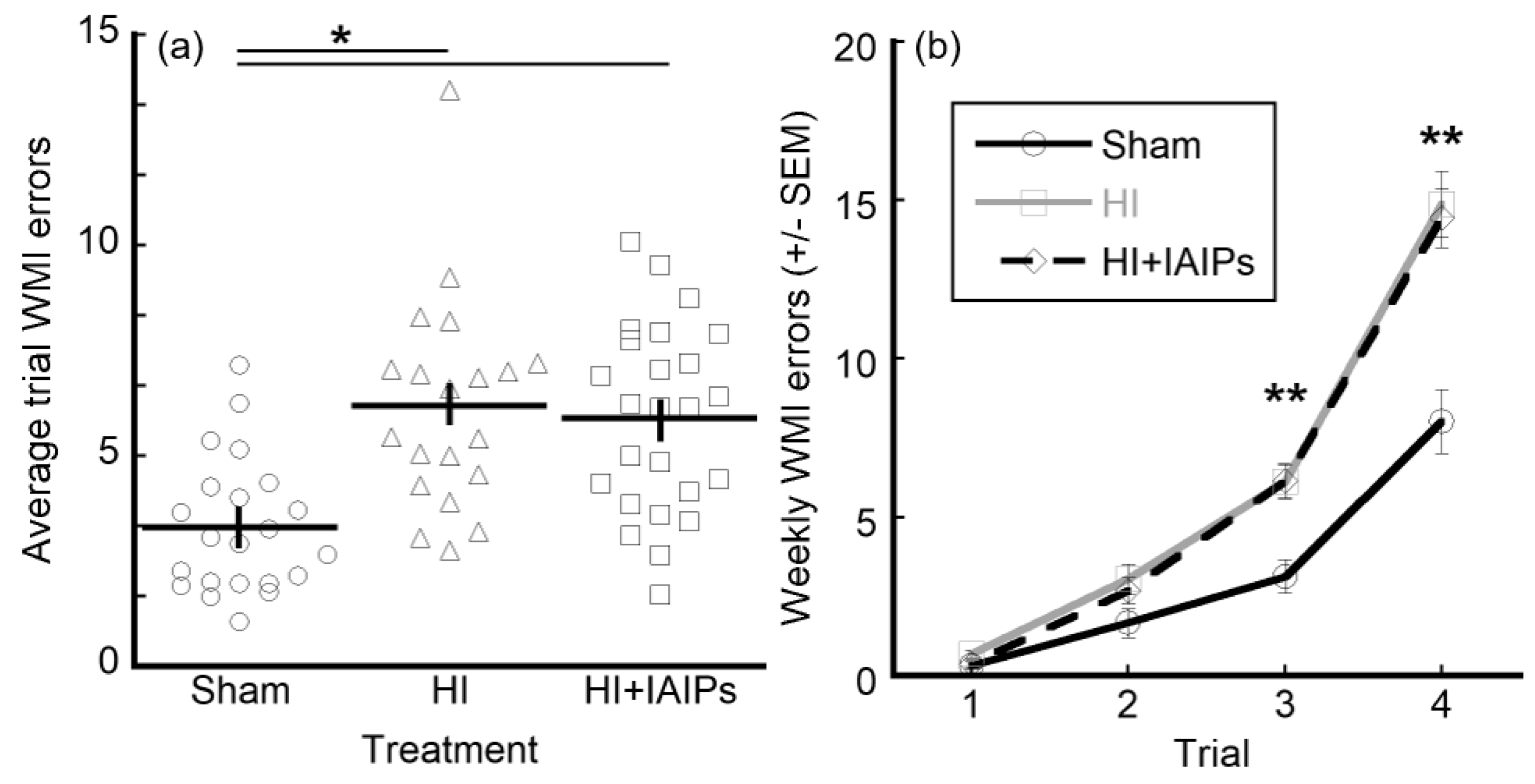
3.2. HI Exposure Increased Error Rates and Juvenile Testing Improved Performance on Retest
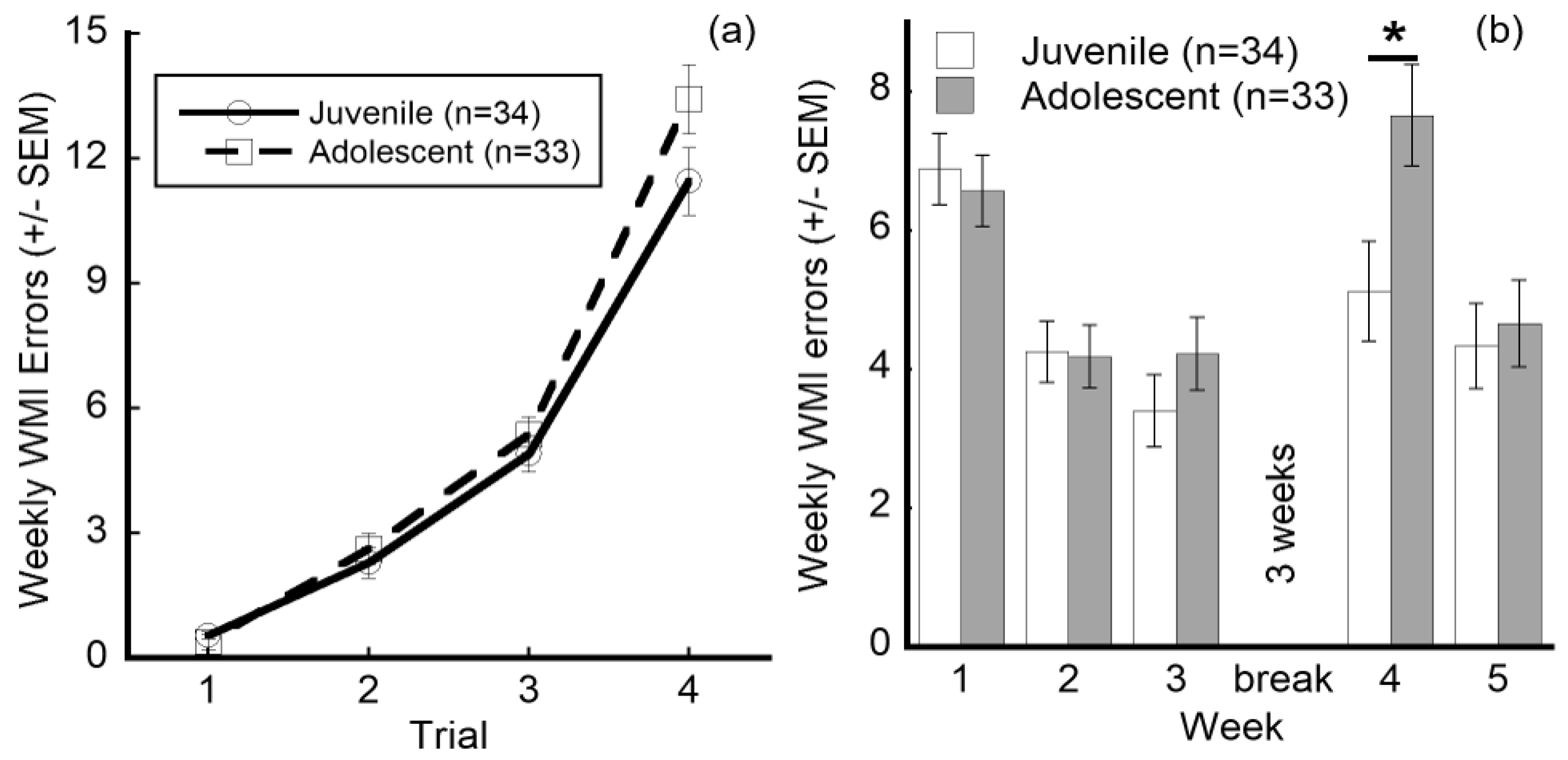
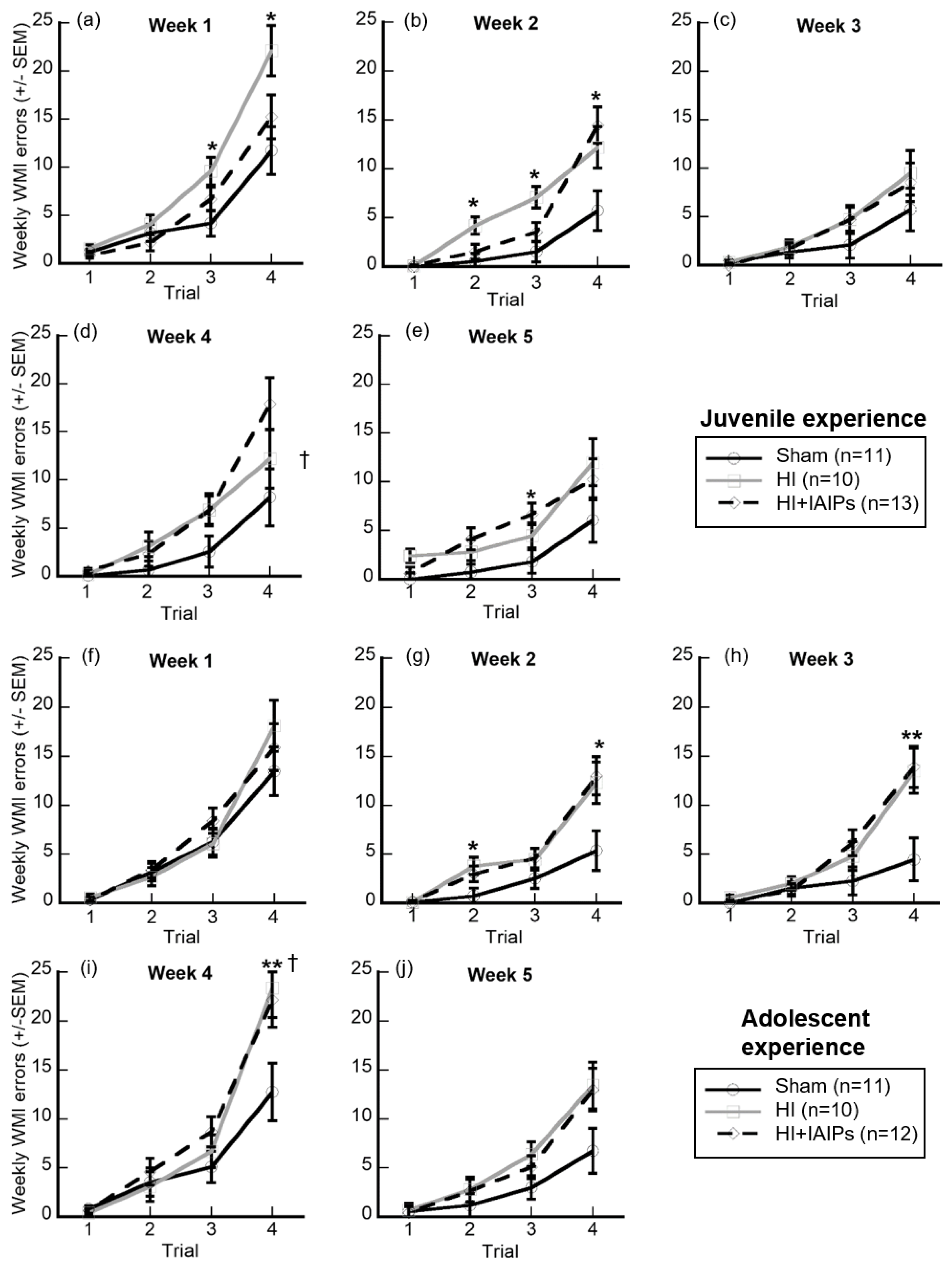
3.3. IAIP Treatment Improved Performance in Juvenile Testing. Juvenile Testing Improved Performance on Trials with High Cognitive Demand and Reduced Working Memory Errors during Retest
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, B.Y.; Castillo, M. Hypoxic-ischemic brain injury: Imaging findings from birth to adulthood. Radiographics 2008, 28, 417–439. [Google Scholar] [CrossRef] [PubMed]
- Varghese, B.; Xavier, R.; Manoj, V.C.; Priya, P.S.; Kumar, A.; Sreenivasan, V.K. Magnetic resonance imaging spectrum of perinatal hypoxic-ischemic brain injury. Ind. J. Radiol. Imaging 2016, 26, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Volpe, J.J. Brain injury in premature infants: A complex amalgam of destructive and developmental disturbances. Lancet Neurol. 2009, 8, 110–124. [Google Scholar] [CrossRef] [Green Version]
- Gopagondanahalli, K.R.; Li, J.; Fahey, M.C.; Hunt, R.W.; Jenkin, G.; Miller, S.L.; Malhotra, A. Pre-term hypoxic-ischemic encephalopathy. Front. Pediatr. 2016, 4, 114. [Google Scholar] [CrossRef] [Green Version]
- Alexander, M.; Garbus, H.; Smith, A.L.; Rosenkrantz, T.S.; Fitch, R.H. Behavioral and histological outcomes following neonatal HI injury in a preterm (P3) and term (P7) rodent model. Behav. Brain Res. 2014, 259, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Millar, L.J.; Shi, L.; Hoerder-Suabedissen, A.; Molnár, Z. Neonatal hypoxia ischaemia: Mechanisms, models, and therapeutic challenges. Front. Cell Neurosci. 2017, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Volpe, J.J. Neonatal encephalopathy: An inadequate term for hypoxic-ischemic encephalopathy. Ann. Neurol. 2012, 72, 156–166. [Google Scholar] [CrossRef]
- Shankaran, S.; Pappas, A.; McDonald, S.A.; Vohr, B.R.; Hintz, S.R.; Yolton, K.; Gustafson, K.E.; Leach, T.M.; Green, C.; Bara, R.; et al. Childhood outcomes after hypothermia for neonatal encephalopathy. NEJM 2012, 366, 2085–2092. [Google Scholar] [CrossRef]
- Pappas, A.; Shankaran, S.; McDonald, S.A.; Vohr, B.R.; Hintz, S.R.; Ehrenkranz, R.A.; Tyson, J.E.; Yolton, K.; Das, A.; Bara, R.; et al. Cognitive outcomes after neonatal encephalopathy. Pediatrics 2015, 135, e624–e634. [Google Scholar] [CrossRef] [Green Version]
- Davidson, J.O.; Wassink, G.; van den Heuij, L.G.; Bennet, L.; Gunn, A.J. Therapeutic hypothermia for neonatal hypoxic-ischemic encephalopathy—Where to from here? Front. Neurol. 2015, 6, 198. [Google Scholar] [CrossRef] [Green Version]
- Pierrat, V.; Arnaud, C.; Resche-Rigon, M.; Bedeau-Livinec, F.; Morgan, A.S.; Marret, S. Neurodevelopmental outcome at 2 years for preterm children born at 22 to 34 weeks’ gestation in France in 2011: EPIPAGE-2 cohort study. BMJ 2017, 358, j3448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.S.; Ohlsson, A.; Perlman, M. Hypothermia to Treat Neonatal Hypoxic Ischemic Encephalopathy. Arch. Ped. Adolesc. Med. 2007, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbo, E.T.; Bartnik-Olson, B.L.; Machado, S.; Merritt, A.; Peverini, R.; Wycliffe, N.; Ashwal, S. The effect of whole-body cooling on brain metabolism following perinatal hypoxic-ischemic injury. Pediatric Res. 2012, 71, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Laptook, A.R.; McDonald, S.A.; Shankaran, S.; Stephens, B.E.; Vohr, B.R.; Guillet, R.; Higgins, R.D.; Das, A. Elevated temperature and 6- to 7-year outcome of neonatal encephalopathy. Ann. Neurol. 2013, 73, 520–528. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Trivedi, S.; Distler, A.; Liao, S.; Vesoulis, Z.; Smyser, C.; Mathur, A.M. Neurodevelopmental outcomes in neonates with mild hypoxic ischemic encephalopathy treated with therapeutic hypothermia. Am. J. Perinatol. 2019, 36, 1337–1343. [Google Scholar] [CrossRef]
- Chevin, M.; Chabrier, S.; Dinomais, M.; Bedell, B.J.; Sébire, G. Benefits of hypothermia in neonatal arterial ischemic strokes: A preclinical study. Int. J. Dev. Neurosci. 2020, 80, 257–266. [Google Scholar] [CrossRef]
- Carty, M.L.; Wixey, J.A.; Reinebrant, H.E.; Gobe, G.; Colditz, P.B.; Buller, K.M. Ibuprofin inhibits neuroinflammation and attenuates white matter damage following hypoxia-ischemia in the immature rodent brain. Brain Res. 2011, 1402, 9–19. [Google Scholar] [CrossRef]
- Wixey, J.A.; Reinebrant, H.E.; Spencer, S.J.; Buller, K.M. Efficacy of post-insult minocycline administration to alter long-term hypoxia-ischemia-induced damage to the serotinergic system in the immature rat brain. Neuroscience 2011, 182, 184–192. [Google Scholar] [CrossRef]
- Poupon-Bejuit, L.; Rocha-Ferriera, E.; Thornton, C.; Hagberg, H.; Rahim, A.A. Neuroprotective effects of diabetes drugs for the treatment of neonatal hypoxia-ischemia encephalopathy. Front. Cell Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Zhuo, L.; Kimata, K. Structure and function of inter-alpha-trypsin inhibitor heavy chains. Connect. Tissue Res. 2008, 49, 311–320. [Google Scholar] [CrossRef]
- Chaaban, H.; Shin, M.; Sirya, E.; Lim, Y.-P.; Caplan, M.; Padbury, J.F. Inter-alpha inhibitor protein level in neonates predicts necrotizing enterocolitis. J. Pediatr. 2010, 157, 757–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Rivard, L.; Naqvi, S.; Nakada, S.; Padbury, J.F.; Sanchez-Esteban, J.; Stopa, E.G.; Lim, Y.-P.; Stonestreet, B.S. Expression and localization of inter-alpha inhibitors in the rodent brain. Neuroscience 2016, 324, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spasova, M.S.; Sadowska, G.B.; Threlkeld, S.W.; Lim, Y.-P.; Stonestreet, B.S. Ontogeny of inter-alpha inhibitor proteins in ovine brain and somatic tissues. Exp. Biol. Med. (Maywood) 2014, 239, 724–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; De La Monte, S.; Hovanesian, V.; Patra, A.; Chen, X.; Chen, R.H.; Miller, M.C.; Pinar, M.H.; Lim, Y.-P.; Stopa, E.G.; et al. Ontogeny of inter-alpha inhibitor protein (IAIP) expression in human brain. J. Neurosci. Res. 2020, 98, 869–887. [Google Scholar] [CrossRef]
- Lord, M.S.; Melrose, J.; Day, A.J.; Whitelock, J.M. The inter-α-trypsin inhibitor family: Versatile molecules in biology and pathology. J. Histochem. Cytochem. 2020, 1–21. [Google Scholar] [CrossRef]
- Opal, S.M.; Artenstein, A.W.; Cristofaro, P.A.; Jhung, J.W.; Palardy, J.E.; Parejo, N.A.; Lim, Y.P. Inter-alpha inhibitor proteins are endogenous furin inhibitors and provide protection against experimental anthrax intoxication. Infect. Immun. 2005, 73, 5101–5105. [Google Scholar] [CrossRef] [Green Version]
- Garandziotis, S.; Hollingsworth, J.W.; Ghanayem, S.; Timberlake, S.; Zhuo, L.; Kimata, K.; Schwartz, D.A. Inter-alpla-trypsin inhibitor attenuates complement activation and complement-induced lung injury. J. Immunol. 2007, 179, 4187–4192. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Zhang, L.X.; Bendelja, K.; Heath, R.; Murphy, S.; Sharma, S.; Padbury, J.F.; Lim, Y.-P. Inter-alpha inhibitor protein administration improves survival from neonatal sepsis in mice. Pediatr. Res. 2010, 68, 242–247. [Google Scholar] [CrossRef] [Green Version]
- Rice, J.E.; Vanucci, R.C. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann. Neurol. 1981, 9, 131–141. [Google Scholar] [CrossRef]
- Winerdal, M.; Winerdal, M.E.; Kinn, J.; Urmaliya, V.; Winqvist, O.; Âdén, U. Long lasting local and systemic inflammation after cerebral hypoxic ischemia in newborn mice. PLoS ONE 2012, 7, e36422. [Google Scholar] [CrossRef] [Green Version]
- Qiao, M.; Meng, S.; Scobie, K.; Foniok, T.; Tuor, U.I. Magnetic resonance imaging of differential gray versus white matter injury following a mild or moderate hypoxic-ischemic insult in neonatal rats. Neurosci. Lett. 2004, 368, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, J.R.; Zimmerman, A.J.; Pavlik, N.; Newville, J.C.; Carlin, K.; Robinson, S.; Brigman, J.L.; Northington, F.J.; Jantzie, L.L. Neonatal hypoxic-ischemic encephalopathy yields permanent deficits in learning acquisition: A preclinical touchscreen assessment. Front. Pediatr. 2020, 8, 289. [Google Scholar] [CrossRef] [PubMed]
- Disdier, C.; Zhang, J.; Fukunaga, Y.; Lim, Y.-P.; Qiu, J.; Santoso, A.; Stonestreet, B.S. Alterations in inter-alpha inhibitor protein expression after hypoxic-ischemic brain injury in neonatal rats. Int. J. Dev. Neurosci. 2018, 65, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Barrios-Anderson, A.; Chen, X.; Nakada, S.; Chen, R.; Lim, Y.-P.; Stonestreet, B.S. Inter-alpha inhibitor proteins modulate neuroinflammatory biomarkers after hypoxia-ischemia in neonatal rats. J. Neuropathol. Exp. Neurol. 2019, 78, 742–755. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Nakada, S.; Donahue, J.E.; Chen, R.H.; Tucker, R.; Qiu, J.; Lim, Y.-P.; Stopa, E.G.; Stonestreet, B.S. Neuroprotective effects of inter-alpha inhibitor proteins after hypoxic-ischemic brain injury in neonatal rats. Exp. Neurol. 2019, 317, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Threlkeld, S.W.; Gaudet, C.M.; La Rue, M.E.; Dugas, E.; Hill, C.A.; Lim, Y.-P.; Stonestreet, B.S. Effects of inter-alpha inhibitor proteins on neonatal brain injury: Age, task and treatment dependent neurobehavioral outcomes. Exp. Neurol. 2014, 261, 424–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudet, C.M.; Lim, Y.-P.; Stonestreet, B.S.; Threlkeld, S.W. Effects of age, experience and inter-alpha inhibitor proteins on working memory and neuronal plasticity after neonatal hypoxia-ischemia. Behav. Brain Res. 2016, 302, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Threlkeld, S.W.; Lim, Y.-P.; La Rue, M.; Gaudet, C.; Stonestreet, B.S. Immuno-modulator inter-alpha inhibitor proteins ameliorate complex auditory processing deficits in rats with neonatal hypoxic-ischemic brain injury. Brain Behav. Immun. 2017, 64, 173–179. [Google Scholar] [CrossRef]
- Penley, S.C.; Gaudet, C.M.; Threlkeld, S.W. Use of an eight-arm radial water maze to assess working and reference memory following neonatal brain injury. JoVE 2013, 82, e50940. [Google Scholar] [CrossRef] [Green Version]
- Dupré, N.; Arabo, A.; Orset, C.; Maucotel, J.; Detroussel, Y.; Hauchecorne, M.; Gonzalez, B.J.; Marret, S.; Vivien, D.; Leroux, P. Neonatal cerebral hypoxia-ischemia in mice triggers age-dependent vascular effects and disabilities in adults; implication of tissue plasminogen activator (tPA). Exp. Neurol. 2020, 323, 113087. [Google Scholar] [CrossRef]
- Woodward, L.J.; Edgin, J.O.; Thompson, D.; Inder, T.E. Object working memory deficits predicted by early brain injury and development in the preterm infant. Brain 2005, 128, 2578–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luu, T.M.; Ment, L.; Allan, W.; Schneider, K.; Vohr, B.R. Executive and memory function in adolescents born very preterm. Pediatrics 2011, 127, e639–e646. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.C.C.; Kozuki, N.; Blencowe, H.; Vos, T.; Bahalim, A.; Darmstadt, G.L.; Niermeyer, S.; Ellis, M.; Robertson, N.J.; Cousens, S.; et al. Intrapartum-related neonatal encephalopathy incidence and impairment at regional and global levels for 2010 with trends from 1990. Pediatr. Res. 2013, 74, 50–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, D.M.; O’Connor, C.M.; Ryan, C.A.; Korotchikova, I.; Boylan, G.B. Early EEG grade and outcome at 6 years after mild neonatal hypoxic ischemic encephalopathy. Pediatrics 2016, 138, e20160659. [Google Scholar] [CrossRef] [Green Version]
- Schreglmann, M.; Ground, A.; Vollmer, B.; Johnson, M.J. Systematic review; long-term cognitive and behavioral outcomes of neonatal hypoxic-ischemic encephalopathy in children without cerebral palsy. Acta Pediatrica 2020, 109, 20–30. [Google Scholar] [CrossRef]
- Hintz, S.R.; Newman, J.E.; Vohr, B.R. Changing definitions of long-term follow-up: Should “long term” be even longer? Semin. Perinat. 2016, 40, 398–409. [Google Scholar] [CrossRef] [Green Version]
- DeMaster, D.; Bick, J.; Johnson, U.; Montroy, J.J.; Landry, S.; Duncan, A.F. Nurturing the preterm infant brain: Leveraging neuroplasticity to improve neurobehavioral outcomes. Pediatr. Res. 2019, 85, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Als, H.; Duffy, F.; McAnulty, G.; Rivkin, M.; Sridhar, V.; Mulkern, R.; Warfield, S.; Huppi, P.; Butler, S.; Conneman, N.; et al. Early Experience Alters Brain Function and Structure. Pediatrics 2004, 133, 846. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.O.; Strapasson, A.C.P.; Nabinger, P.M.; Achaval, M.; Netto, C.A. Early enriched housing results in partial recovery of memory deficits in female, but not in male, rats after neonatal hypoxia-ischemia. Brain Res. 2008, 1218, 257–268. [Google Scholar] [CrossRef]
- Rojas, J.J.; Deniz, B.F.; Miguel, P.M.; Diaz, R.; Hermel, E.E.S.; Achaval, M.; Netto, C.A.; Pereira, L.O. Effects of daily environmental enrichment on behavior and dendritic spine density in hippocampus following neonatal hypoxia-ischemia in the rat. Exp. Neurol. 2013, 241, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Forbes, T.A.; Goldstein, E.Z.; Dupree, J.L.; Jablonska, B.; Scafidi, J.; Adams, K.L.; Imamura, Y.; Hashimoto-Torii, K.; Gallo, V. Environmental enrichment ameliorates perinatal brain injury and promotes functional white matter recovery. Nat. Commun. 2020, 11, 964. [Google Scholar] [CrossRef] [PubMed]
- van Wassenaer-Leemhuis, A.G.; Jeukens-Visser, M.; van Hus, J.W.P.; Meijssen, D.; Wolf, M.-J.; Kok, J.H.; Nollet, F.; Koldewijn, K. Rethinking preventative post-discharge intervention programmes for very preterm infants and their parents. Dev. Med. Child Neurol. 2016, 58, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Threlkeld, S.W.; Hill, C.A.; Rosen, G.D.; Fitch, R.H. Early acoustic discrimination experience ameliorates auditory processing deficits in male rats with cortical developmental disruption. Int. J. Dev. Neurosci. 2009, 27, 321–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, A.; Lebessi, A.; Sowinski, P.; Hodges, H. Comparison of effects of global cerebral ischaemia on spatial learning in the standard and radial water maze: Relationship of hippocampal damage to performance. Behav. Brain Res. 1997, 85, 93–115. [Google Scholar] [CrossRef]
- Shukitt-Hale, B.; McEwen, J.J.; Szprengiel, A.; Joseph, J.A. Effect of age on radial arm water maze-a test of spatial learning and memory. Neurobiol. Aging 2004, 25, 223–229. [Google Scholar] [CrossRef]
- Astur, R.S.; Tropp, J.; Sava, S.; Constable, R.T.; Markus, E.J. Sex differences and correlations in a virtual Morris water task, a virtual radial arm maze, and mental rotation. Behav. Brain Res. 2004, 151, 103–115. [Google Scholar] [CrossRef]
- Mennenga, S.E.; Baxter, L.C.; Grunfield, I.S.; Brewer, G.A.; Aiken, L.S.; Engler-Chiurazzi, E.B.; Camp, B.W.; Acosta, J.I.; Braden, B.B.; Schaefer, K.R.; et al. Navigating to new frontiers in behavioral neuroscience: Traditional neuropsychological tests predict human performance on a rodent-inspired radial-arm maze. Front. Behav. Neurosci. 2014, 8, 294. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.A.; Threlkeld, S.W.; Fitch, R.H. Early testosterone modulated sex differences in behavioral outcome following neonatal hypoxia ischemia in rats. Int. J. Dev. Neurosci. 2011, 29, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.L.; Alexander, M.; Rosenkrantz, T.S.; Sadek, M.I.; Fitch, R.H. Sex differences in behavioral outcome following neonatal hypoxia ischemia: Insights from a clinical meta-analysis and a rodent model of induced hypoxic ischemic brain injury. Exp. Neurol. 2014, 254, 54–67. [Google Scholar] [CrossRef]
- Waddell, J.; Hanscom, M.; Edwards, N.S.; McKenna, M.C.; McCarthy, M.M. Sex differences in cell genesis, hippocampal volume and behavioral outcomes in a rat model of neonatal HI. Exp. Neurol. 2016, 275, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Bimonte, H.; Hyde, L.; Hoplight, B.; Denenberg, V. In two species, females exhibit superior working memory and inferior reference memory on the water radial arm maze. Physiol. Behav. 2000, 70, 311–317. [Google Scholar] [CrossRef]
- Bimonte-Nelson, H.A.; Singleton, R.S.; Nelson, M.A.; Eckman, C.B.; Barber, J.; Scott, T.Y.; Granholm, A.-C.E. Testosterone, but not nonaromatizable dihydrotestosterone, improves working memory and alters nerve growth factor levels in aged male rats. Exp. Neurol. 2003, 181, 301–312. [Google Scholar] [CrossRef]
- Hoh, T.E.; Kolb, B.; Eppel, A.; Vanderwolf, C.H.; Cain, D.P. Role of the neocortex in the water maze task in the rat: A detailed behavioral and Golgi-Cox analysis. Brain Behav. Res. 2003, 138, 81–94. [Google Scholar] [CrossRef]
- Frick, K.M.; Baxter, M.G.; Markowska, A.L.; Olton, D.S.; Price, D.L. Age-related spatial reference and working memory deficites assessed in the water maze. Neurobiol. Aging 1995, 16, 149–160. [Google Scholar] [CrossRef]
- Berchtold, N.C.; Castello, N.; Cotman, C.W. Exercise and time-dependent benefits of learning and memory. Neuroscience 2010, 167, 588–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.-W.; Lin, Y.-C.; Wang, S.-T.; Yeh, T.-F.; Huang, C.-C. Hypoxic/ischemic and infectious events have cumulative effects on the risk of cerebral palsy in very-low-birth-weight preterm infants. Neonatology 2014, 106, 209–215. [Google Scholar] [CrossRef]
- Stoll, B.J.; Hansen, N.J.; Adams-Chapman, I.; Fanaroff, A.A.; Hintz, S.R.; Vohr, B.; Higgins, R.D. Neurodevelopmental and growth impairment among extremely low-birth-weight infants with neonatal infection. JAMA 2004, 292, 2357–2365. [Google Scholar] [CrossRef] [Green Version]
- Aanes, S.; Bjuland, K.J.; Winsnes Lærum, A.M.; Wieder, S.; Evensen, K.A.I.; Indredavik, M.; Brubakk, A.M.; Håberg, A.; Lohaugen, G.C.; Skranes, J. Relationship between hippocampal subfield volumes and memory function in adults born preterm with very low birth weight (VLBW). Clin. Obstet. Gynecol. Reprod. Med. 2020, 6, 1–11. [Google Scholar] [CrossRef]
- Ehrenkranz, R.A.; Dusick, A.M.; Vohr, B.R.; Wright, L.L.; Wrage, L.A.; Poole, W.K. Growth in the neonatal intensive care unit influences neurodevelopmental and growth outcomes of extremely low birth weight infants. Pediatrics 2006, 117, 1253–1261. [Google Scholar] [CrossRef] [Green Version]
- Bona, E.; Johansson, B.B.; Hagberg, H. Sensorimotor function and neuropathology five to six weeks after hypoxia-ischemia in seven-day-old rats. Pediatr. Res. 1997, 42, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Lubics, A.; Reglodi, D.; Tamás, A.; Kiss, P.; Szalai, M.; Szalontay, L.; Lengvári, I. Neurological reflexes and early motor behavior in rats subjected to neonatal hypoxic-ischemic injury. Behav. Brain Res. 2005, 157, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Farias Sanches, E.; Sarmento Arteni, N.; Spindler, C.; Moysés, F.; Rodrigues Siqueira, I.; Luis Perry, M.; Alexandre Netto, C. Effects of pre- and postnatal protein malnutrition in hypoxic-ischemic rats. Brain Res. 2012, 1438, 85–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, B.P.; Nedelcu, J.; Martin, E. Delayed postischemic hypothermia improves long-term behavioral outcome after cerebral hypoxia-ischemia in neonatal rats. Pediatr. Res. 2002, 51, 354–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borjini, N.; Sivilia, S.; Guiliani, A.; Fernandez, M.; Giardino, L.; Facchinetti, F.; Calzá, L. Potential biomarkers for neuroinflammation and neurodegeneration at short and long term after neonatal hypoxic-ischemic insult in rat. J. Neuroinflamm. 2019, 9, 194. [Google Scholar] [CrossRef]
- Patki, G.; Li, L.; Allam, F.; Solanki, N.; Dao, A.T.; Alkadhi, K.; Salim, S. Moderate treadmill exercise rescues anxiety and depression-like behavior as well as memory impairment in a rat model of posttraumatic stress disorder. Physiol. Behav. 2014, 130, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Sparling, J.E.; Baker, S.L.; Bielajew, C. Effects of combined pre- and post-natal enrichment on anxiety-like, social, and cognitive behaviors in juvenile and adult rat offspring. Behav. Brain Res. 2018, 353, 40–50. [Google Scholar] [CrossRef]
- Figuierêdo do Bonfim, T.H.; Leite Tavares, R.; Araújo de Vasconcelos, M.H.; Gouveia, M.; Campos Nunes, P.; Soares, N.L.; Coutinho Alves, R.; Pinto de Carvalho, J.L.; Alves, A.F.; de Alencar Pereira, R.; et al. Potentially obesogenic diets alter metabolic and neurobehavioural parameters in Wistar rats: A comparison between two dietary models. J. Affect Disord. 2020, 279, 451–461. [Google Scholar] [CrossRef]
- Hagberg, H.; Mallard, C.; Ferriero, D.M.; Vannucci, S.J.; Levison, S.W.; Vexler, Z.S.; Gressens, P. The role of inflammation in perinatal brain injury. Nat. Rev. Neurol. 2015, 11, 192–208. [Google Scholar] [CrossRef]
- Mirza, M.A.; Ritzel, R.; Xu, Y.; McCullough, L.D.; Liu, F. Sexually dimorphic outcomes and inflammatory responses in hypoxic-ischemic encephalopathy. J. Neuroinflamm. 2015, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Villapol, S.; Faivre, V.; Joshi, P.; Moretti, R.; Besson, V.C.; Charriaut-Marlangue, C. Early sex differences in the immune-inflammatory responses to neonatal ischemic stroke. Int. J. Mol. Sci. 2019, 20, 3809. [Google Scholar] [CrossRef] [Green Version]
- Logsdon, A.F.; Erickson, M.A.; Chen, X.; Qiu, J.; Lim, Y.-P.; Stonestreet, B.S.; Banks, W.A. Inter-alpha inhibitor proteins attenuate lipopolysaccharide-induced blood-brain barrier disruption and downregulate circulating interleukin 6 in mice. J. Cereb. Blood Flow Metab. 2020, 40, 1090–1102. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor (n) | Average WMI Errors | Significance [Test, p] | |
|---|---|---|---|
| Exposure/Treatment | * Main effect [RMANOVA, p < 0.001] | ||
| Sham + Vehicle (n = 22) | 3.289 | ||
| HI + Vehicle (n = 20) | 6.188 * | * vs. Sham [t-test, p < 0.001] | |
| HI + IAIPs (n = 25) | 5.909 * | * vs. Sham [t-test, p < 0.001] | |
| Experience Group | NS Main effect [RMANOVA, p > 0.05] | ||
| Juvenile (n = 34) | 4.798 | ||
| Adolescent (n = 33) | 5.458 | ||
| Exposure × Treatment Interaction | Juvenile | Adolescent | NS Interaction effect [RMANOVA, p > 0.05] |
| Sham + Vehicle | 2.882 (n = 11) | 3.695 (n = 11) | |
| HI + Vehicle | 6.075 * (n = 10) | 6.300 * (n = 10) | * vs. Sham [t-test, p < 0.001] |
| HI + IAIPs | 5.438 * (n = 13) | 6.379 * (n = 12) | * vs. Sham [t-test, p < 0.001] |
| Factor 1 (n) | Factor 2 Average WMI Errors | Significance [Test, p] | ||||
|---|---|---|---|---|---|---|
| Exposure/Treatment | Trial | * Interaction effect [RMANOVA, p < 0.001] | ||||
| 1 | 2 | 3 | 4 | |||
| Sham + Vehicle (n = 22) | 0.336 | 1.664 | 3.136 | 8.018 | ||
| HI + Vehicle (n = 20) | 0.700 | 3.050 | 6.130 * | 14.870 * | * vs. Sham in trials 3 and 4 [t-tests, p < 0.001] | |
| HI + IAIPs (n = 25) | 0.374 | 2.685 | 6.138 * | 14.438 * | * vs. Sham in trials 3 and 4 [t-tests, p < 0.001] | |
| Experience Group | Trial | * Interaction effect [RMANOVA, p < 0.05] | ||||
| 1 | 2 | 3 | 4 | |||
| Juvenile (n = 34) | 0.548 | 2.293 | 4.896 | 11.456 | ||
| Adolescent (n = 33) | 0.392 | 2.639 | 5.373 | 13.428 | NS [t-tests, p > 0.05] | |
| Experience Group (total) | Week | * Interaction effect [RMANOVA, p < 0.001] | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| Juvenile (n = 34) | 6.883 | 4.253 | 3.398 | 5.122 | 4.336 | |
| Adolescent (n = 33) | 6.571 | 4.181 | 4.223 | 7.655 * | 4.660 | * vs. juvenile in wk4 [t-tests, p < 0.05] |
| Sham + Vehicle | ||||||
| Juvenile (n = 11) | 5.068 | 1.955 | 2.364 | 2.863 | 2.159 | |
| Adolescent (n = 11) | 5.818 | 2.182 | 2.068 | 5.545 * | 2.864 | * vs. juvenile in wk4 [t-tests, p < 0.05] |
| HI + Vehicle | ||||||
| Juvenile (n = 10) | 9.350 | 5.900 | 4.100 | 5.600 | 5.425 | |
| Adolescent (n = 10) | 6.875 | 5.175 | 5.225 | 8.400 * | 5.825 | * vs. juvenile in wk4 [t-tests, p < 0.05] |
| HI + IAIPs | ||||||
| Juvenile (n = 13) | 6.231 | 4.903 | 3.731 | 6.903 | 5.423 | |
| Adolescent (n = 12) | 7.021 | 5.188 | 5.375 | 9.021 * | 5.291 | * vs. juvenile in wk4 [t-tests, p < 0.05] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bradford, A.; Hernandez, M.; Kearney, E.; Theriault, L.; Lim, Y.-P.; Stonestreet, B.S.; Threlkeld, S.W. Effects of Juvenile or Adolescent Working Memory Experience and Inter-Alpha Inhibitor Protein Treatment after Neonatal Hypoxia-Ischemia. Brain Sci. 2020, 10, 999. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10120999
Bradford A, Hernandez M, Kearney E, Theriault L, Lim Y-P, Stonestreet BS, Threlkeld SW. Effects of Juvenile or Adolescent Working Memory Experience and Inter-Alpha Inhibitor Protein Treatment after Neonatal Hypoxia-Ischemia. Brain Sciences. 2020; 10(12):999. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10120999
Chicago/Turabian StyleBradford, Aaron, Miranda Hernandez, Elaine Kearney, Luke Theriault, Yow-Pin Lim, Barbara S. Stonestreet, and Steven W. Threlkeld. 2020. "Effects of Juvenile or Adolescent Working Memory Experience and Inter-Alpha Inhibitor Protein Treatment after Neonatal Hypoxia-Ischemia" Brain Sciences 10, no. 12: 999. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci10120999