
Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia–Glioma Cross-Talk

 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
- all clinical studies reporting the role of SupTR in HGGs excluding LGG; and
- all basic research concerning microglia-glioma cross-talk in peritumoral areas of HGGs focusing on the role of microRNAs.
3. Review
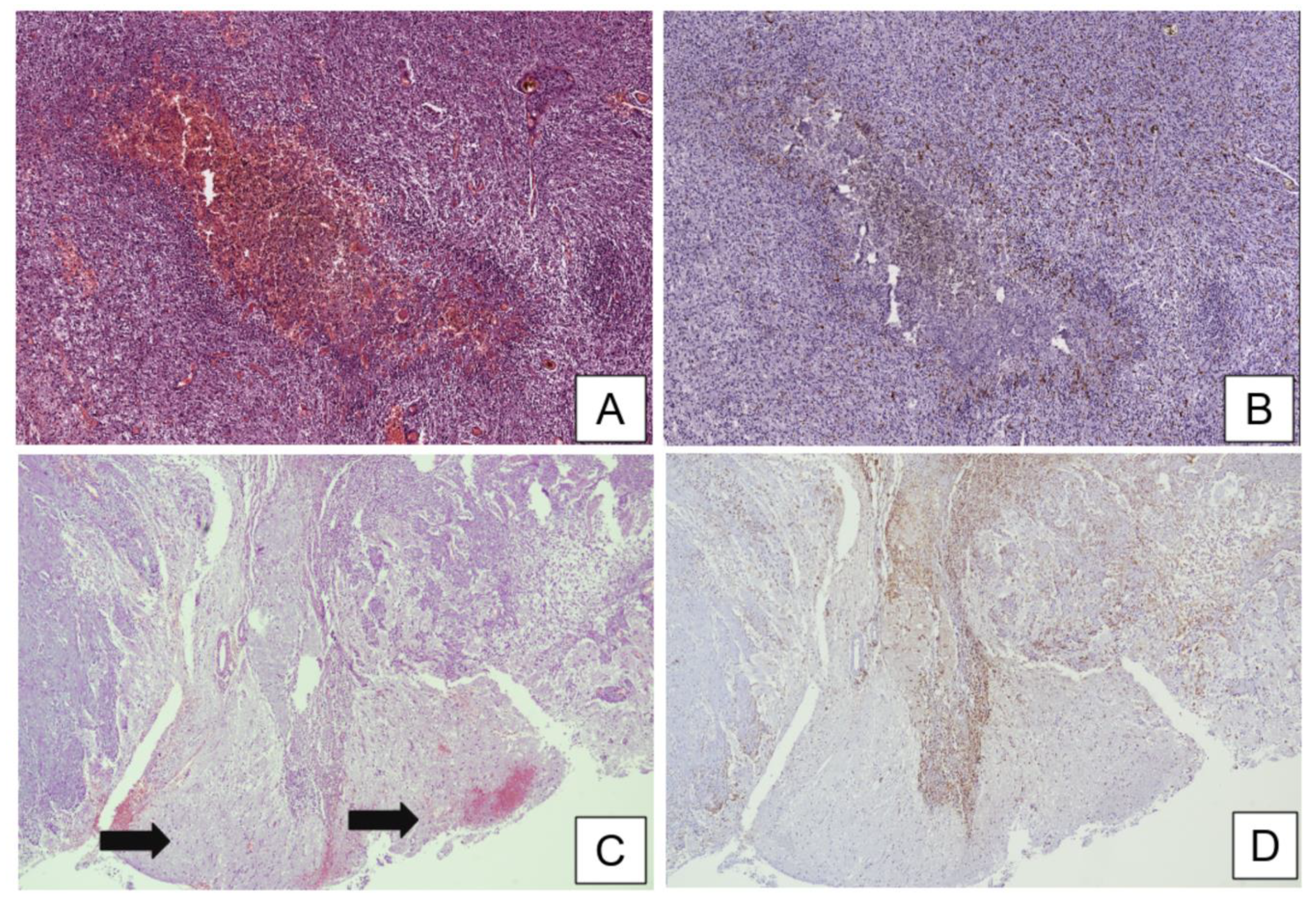
3.1. Histopathological Assessment of Microglia in the Central Nervous System (CNS) and Microglia–Glioma Cross-Talk in Peritumoral Areas
3.2. MicroRNAs (miRNAs) Are Extensively Dysregulated in GBM
3.3. miRNAs Show Specific Patterns of Expression in GBM Core and in the Peritumoral Area
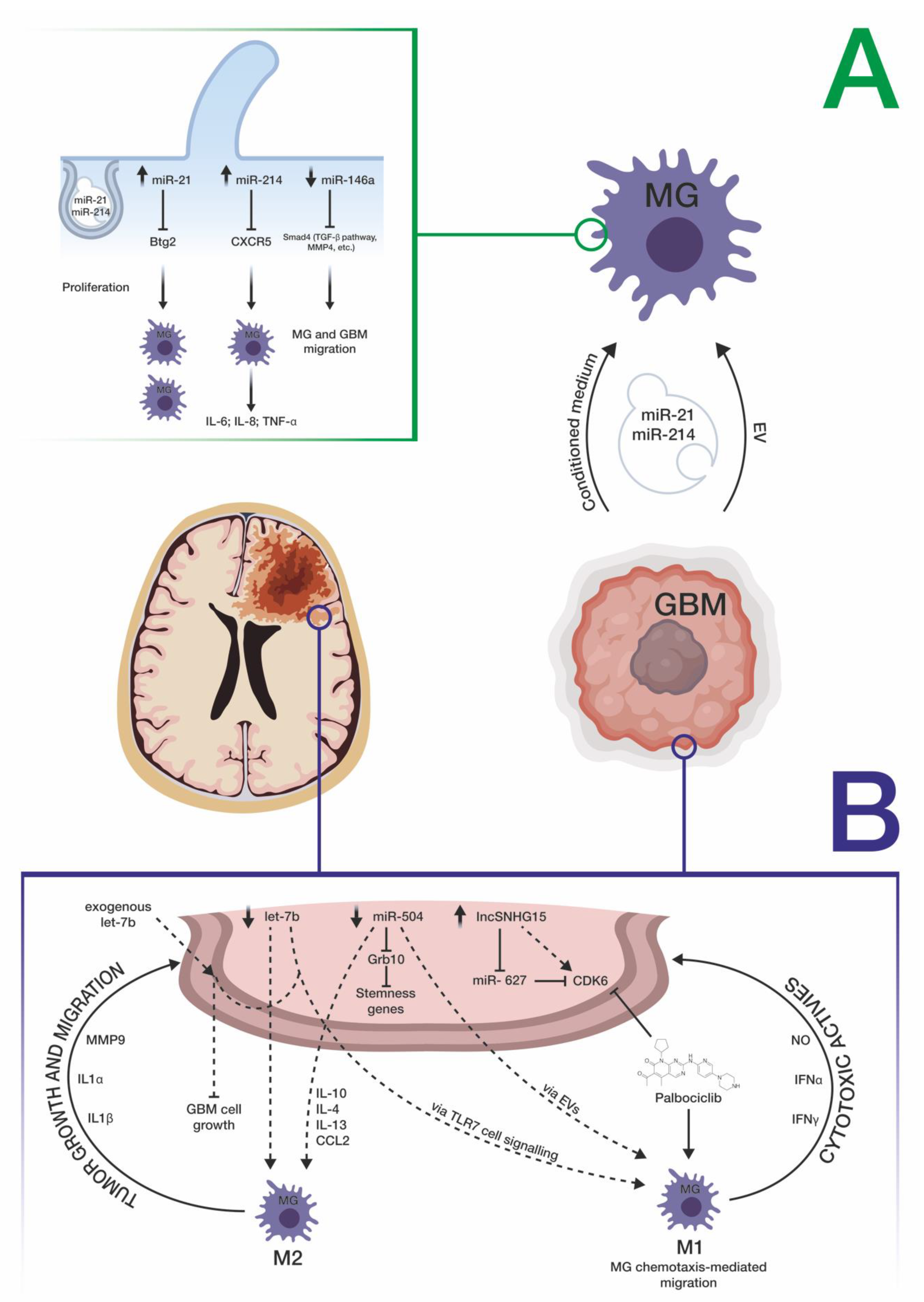
3.4. miRNAs Mediate the Cross-Talk between GBM and Microglia Cells
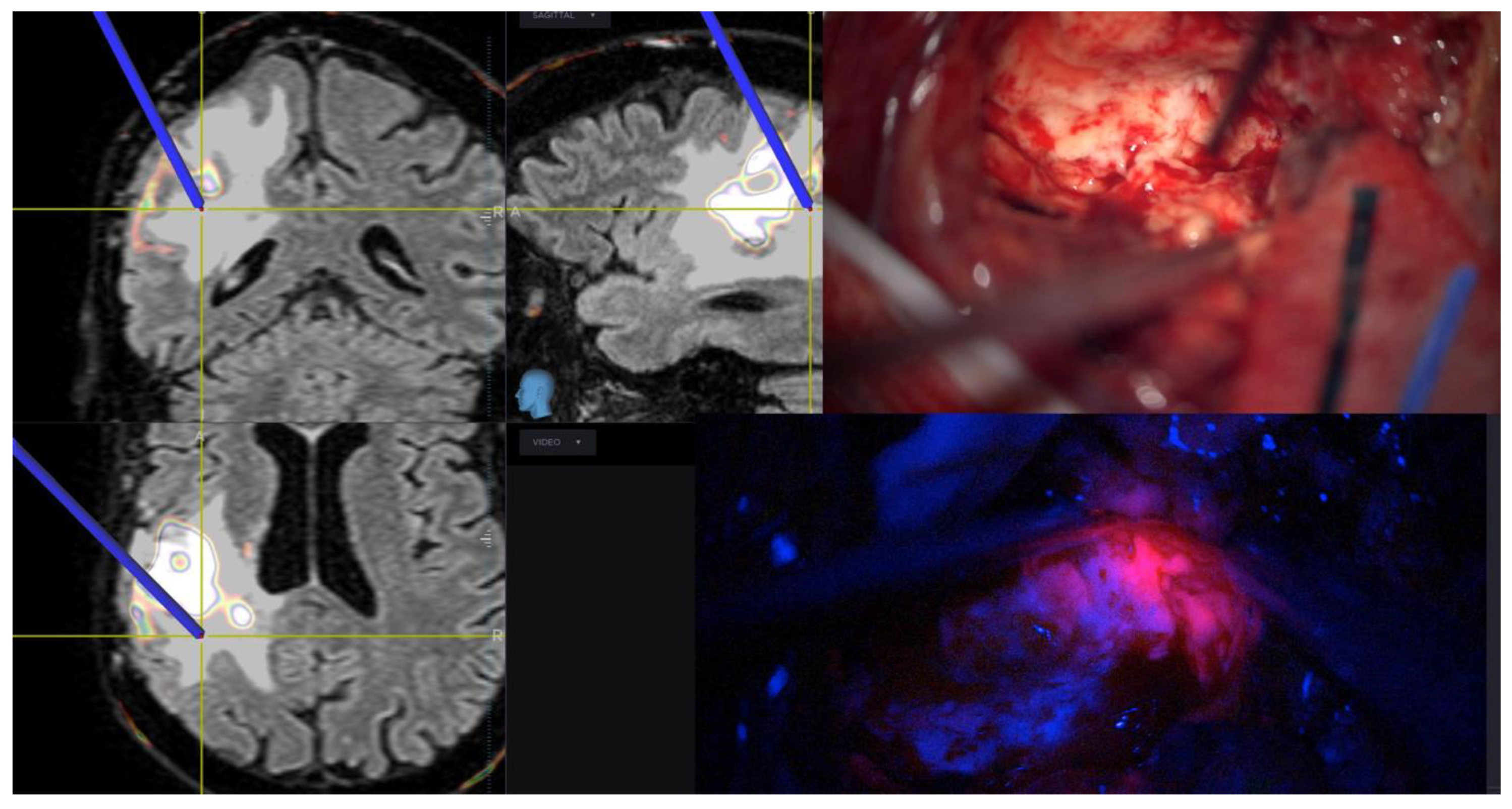
3.5. Role of Surgery in Peritumoral Infiltrated Areas
3.6. Where We Are Going
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HGGs | High-Grade Gliomas |
| SupTR | Supratotal Resection |
| EN | Enhancing Nodule |
| GBM | Glioblastoma |
| 5-ALA | 5-Aminolevulinic Acid |
| CNS | Central Nervous System |
| TILs | Tumor-Infiltrating Lymphocytes |
| GAMs | Glioma-Associated Microglia and Macrophages |
| BBB | Blood–Brain Barrier |
| H&E | Hematoxylin and Eosin |
| HC | Histochemistry |
| IHC | Immunohistochemistry |
| ECM | Extracellular Matrix |
| CCL2 | Chemokine C–C Motif Ligand 2 |
| miRNAs | MicroRNAs |
| EGFR | Epidermal Growth Factor Receptor |
| LRRFIP1 | LRR Binding FLII Interacting Protein 1 |
| DE | Differentially Expressed |
| OPC | Oligodendrocyte Precursor Cell |
| Evs | Extracellular Vesicles |
| Btg2 | BTG Anti-Proliferation Factor 2 |
| CXCR5 | C–X–C Motif Chemokine Receptor 5 |
| TLR7 | Toll-Like Receptor 7 |
| SMAD4 | SMAD Family Member 4 |
| MMP9 | Matrix Metallopeptidase 9 |
| TMZ | Temozolomide |
| IDH-1 | Isocitrate Dehydrogenase 1 |
| RT | Radiotherapy |
| OS | Overall Survival |
| PFS | Progression Free Survival |
| TSC-mTOR | Tuberous Sclerosis Complex |
References
- Duffau, H. Long-term outcomes after supratotal resection of diffuse low-grade gliomas: A consecutive series with 11-year follow-up. Acta Neurochir. 2016, 158, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Suki, D.; Hess, K.R.; Sawaya, R. The influence of maximum safe resection of glioblastoma on survival in 1229 patients: Can we do better than gross-total resection? J. Neurosurg. 2016, 124, 977–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mampre, D.; Ehresman, J.; Pinilla-Monsalve, G.; Osorio, M.A.G.; Olivi, A.; Quinones-Hinojosa, A.; Chaichana, K.L. Extending the resection beyond the contrast-enhancement for glioblastoma: Feasibility, efficacy, and outcomes. Br. J. Neurosurg. 2018, 32, 528–535. [Google Scholar] [CrossRef]
- Pessina, F.; Navarria, P.; Cozzi, L.; Ascolese, A.M.; Simonelli, M.; Santoro, A.; Clerici, E.; Rossi, M.; Scorsetti, M.; Bello, L. Maximize surgical resection beyond contrast-enhancing boundaries in newly diagnosed glioblastoma multiforme: Is it useful and safe? A single institution retrospective experience. J. Neuro-Oncol. 2017, 135, 129–139. [Google Scholar] [CrossRef]
- Brown, J.M. Radiation damage to tumor vasculature initiates a program that promotes tumor recurrences. Int. J. Radiat. Oncol. 2020, 108, 734–744. [Google Scholar] [CrossRef]
- Dimou, J.; Beland, B.; Kelly, J. Supramaximal resection: A systematic review of its safety, efficacy and feasibility in glioblastoma. J. Clin. Neurosci. 2020, 72, 328–334. [Google Scholar] [CrossRef]
- Altieri, R.; Melcarne, A.; Soffietti, R.; Rudá, R.; Franchino, F.; Pellerino, A.; La Rocca, G.; Sabatino, G.; Olivi, A.; Ducati, A.; et al. Supratotal resection of glioblastoma: Is less more? Surg. Technol. Int. 2019, 35, 432–440. [Google Scholar] [PubMed]
- Altieri, R.; Zenga, F.; Ducati, A.; Melcarne, A.; Cofano, F.; Mammi, M.; Di Perna, G.; Savastano, R.; Garbossa, D. Tumor location and patient age predict biological signatures of high-grade gliomas. Neurosurg. Rev. 2017, 41, 599–604. [Google Scholar] [CrossRef]
- Certo, F.; Altieri, R.; Maione, M.; Schonauer, C.; Sortino, G.; Fiumanò, G.; Tirrò, E.; Massimino, M.; Broggi, G.; Vigneri, P.; et al. FLAIRectomy in supramarginal resection of glioblastoma correlates with clinical outcome and survival analysis: A prospective, single institution, case series. Oper. Neurosurg. 2020, 20, 151–163. [Google Scholar] [CrossRef]
- Certo, F.; Stummer, W.; Farah, J.O.; Freyschlag, C.; Visocchi, M.; Morrone, A.; Altieri, R.; Toccaceli, G.; Peschillo, S.; Thomè, C.; et al. Supramarginal resection of glioblastoma: 5-ALA fluorescence, combined intraoperative strategies and correlation with survival. J. Neurosurg. Sci. 2020, 63, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, M.; Zeppa, P.; Zenga, F.; Altieri, R.; Mammi, M.; Bertero, L.; Castellano, I.; Cassoni, P.; Melcarne, A.; La Rocca, G.; et al. The post-surgical era of GBM: How molecular biology has impacted on our clinical management. A review. Clin. Neurol. Neurosurg. 2018, 170, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Ross, J.L.; Cooper, L.A.; Kong, J.; Gutman, D.; Williams, M.; Tucker-Burden, C.; McCrary, M.R.; Bouras, A.; Kaluzova, M.; Dunn, W.D.; et al. 5-Aminolevulinic acid guided sampling of glioblastoma microenvironments identifies pro-survival signaling at infiltrative margins. Sci. Rep. 2017, 7, 15593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müther, M.; Koch, R.; Weckesser, M.; Sporns, P.; Schwindt, W.; Stummer, W. 5-aminolevulinic acid fluorescence-guided resection of 18F-FET-PET positive tumor beyond gadolinium enhancing tumor improves survival in glioblastoma. Neurosurgery 2019, 85, E1020–E1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaşargil, M.G.; Kadri, P.A.; Yaşargil, D.C. Microsurgery for malignant gliomas. J. Neuro-Oncol. 2004, 69, 67–81. [Google Scholar] [CrossRef]
- Juliano, J.; Gil, O.; Hawkins-Daarud, A.; Noticewala, S.; Rockne, R.C.; Gallaher, J.; Massey, S.C.; Sims, P.A.; Anderson, A.R.A.; Swanson, K.R.; et al. Comparative dynamics of microglial and glioma cell motility at the infiltrative margin of brain tumours. J. R. Soc. Interface 2018, 15, 20170582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Fridman, W.H.; Pagès, F.; Sautès-Fridman, C.; Galon, J. The immune contexture in human tumours: Impact on clinical outcome. Nat. Rev. Cancer 2012, 12, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Berghoff, A.S.; Kiesel, B.; Widhalm, G.; Rajky, O.; Ricken, G.; Wöhrer, A.; Dieckmann, K.; Filipits, M.; Brandstetter, A.; Weller, M.; et al. Programmed death ligand 1 expression and tumor-infiltrating lymphocytes in glioblastoma. Neuro-Oncology 2015, 17, 1064–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorsson, V.; Gibbs, D.L.; Brown, S.D.; Wolf, D.; Bortone, D.S.; Ou Yang, T.-H.; Porta-Pardo, E.; Gao, G.F.; Plaisier, C.L.; Eddy, J.A.; et al. The immune landscape of cancer. Immunity 2019, 51, 411–412. [Google Scholar] [CrossRef] [Green Version]
- Schiffer, D.; Annovazzi, L.; Casalone, C.; Corona, C.; Mellai, M. Glioblastoma: Microenvironment and niche concept. Cancers 2018, 11, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiavari, M.; Ciotti, G.M.P.; Navarra, P.; Lisi, L. Pro-inflammatory activation of a new immortalized human microglia cell line. Brain Sci. 2019, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Arcuri, C. Microglia-glioma cross-talk a two way approach to new strategies against glioma. Front. Biosci. 2017, 22, 268–309. [Google Scholar] [CrossRef] [Green Version]
- Wolf, S.A.; Boddeke, H.W.G.M.; Kettenmann, H. Microglia in physiology and disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef] [PubMed]
- Zeiner, P.S.; Preusse, C.; Golebiewska, A.; Zinke, J.; Iriondo, A.; Muller, A.; Kaoma, T.; Filipski, K.; Müller-Eschner, M.; Bernatz, S.; et al. Distribution and prognostic impact of microglia/macrophage subpopulations in gliomas. Brain Pathol. 2018, 29, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Carola, V.; Viscomi, M.T. Microglia. Semin. Cell Dev. Biol. 2019, 94, 94–95. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, D.; Li, G. The role of microglia in viral encephalitis: A review. J. Neuroinflammation 2019, 16, 76. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhang, S. Microgliosis in the injured brain. Neuroscientist 2016, 22, 165–170. [Google Scholar] [CrossRef]
- Glioma-Derived IL-33 Orchestrates an Inflammatory Brain Tumor Microenvironment that Accelerates Glioma Progression. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/s41467-020-18569-4 (accessed on 30 November 2020).
- Bettinger, I.; Thanos, S.; Paulus, W. Microglia promote glioma migration. Acta Neuropathol. 2002, 103, 351–355. [Google Scholar] [CrossRef]
- Coniglio, S.J.; Segall, J.E. Review: Molecular mechanism of microglia stimulated glioblastoma invasion. Matrix Biol. 2013, 32, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, A.C.C.; Badie, B. Microglia and macrophages in malignant gliomas: Recent discoveries and implications for promising therapies. Clin. Dev. Immunol. 2013, 2013, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Penfield, W. Microglia and the process of phagocytosis in gliomas. Am. J. Pathol. 1925, 1, 77. [Google Scholar]
- Zhang, J.; Sarkar, S.; Cua, R.; Zhou, Y.; Hader, W.; Yong, V.W. A dialog between glioma and microglia that promotes tumor invasiveness through the CCL2/CCR2/interleukin-6 axis. Carcinogenesis 2012, 33, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Lisi, L.; Stigliano, E.; Lauriola, L.; Navarra, P.; Russo, C.D. Proinflammatory-activated glioma cells induce a switch in microglial polarization and activation status, from a predominant M2b phenotype to a mixture of M1 and M2a/B polarized cells. ASN Neuro 2014, 6, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Barbagallo, D.; Vittone, G.; Romani, M.; Purrello, M. Noncoding RNAs in health and disease. Int. J. Genom. 2018, 2018, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parodi, F.; Carosio, R.; Ragusa, M.; Di Pietro, C.; Maugeri, M.; Barbagallo, D.; Sallustio, F.; Allemanni, G.; Pistillo, M.P.; Casciano, I.; et al. Epigenetic dysregulation in neuroblastoma: A tale of miRNAs and DNA methylation. Biochim. Biophys. Acta 2016, 1859, 1502–1514. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, M.; Barbagallo, C.; Brex, D.; Caponnetto, A.; Cirnigliaro, M.; Battaglia, R.; Barbagallo, D.; Di Pietro, C.; Purrello, M. Molecular crosstalking among noncoding RNAs: A new network layer of genome regulation in cancer. Int. J. Genom. 2017, 2017, 4723193. [Google Scholar] [CrossRef]
- Ragusa, M.; Bosco, P.; Tamburello, L.; Barbagallo, C.; Condorelli, A.G.; Tornitore, M.; Spada, R.S.; Barbagallo, D.; Scalia, M.; Elia, M.; et al. miRNAs plasma profiles in vascular dementia: Biomolecular data and biomedical implications. Front. Cell. Neurosci. 2016, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Ciafrè, S.; Galardi, S.; Mangiola, A.; Ferracin, M.; Liu, C.-G.; Sabatino, G.; Negrini, M.; Maira, G.; Croce, C.; Farace, M. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem. Biophys. Res. Commun. 2005, 334, 1351–1358. [Google Scholar] [CrossRef]
- Chan, J.A.; Krichevsky, A.M.; Kosik, K.S. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res. 2005, 65, 6029–6033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillies, J.K.; Lorimer, I.A.J. Regulation of p27Kip1by miRNA 221/222 in glioblastoma. Cell Cycle 2007, 6, 2005–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukiw, W.J.; Cui, J.G.; Li, Y.Y.; Culicchia, F. Up-regulation of micro-RNA-221 (miRNA-221; chr Xp11.3) and caspase-3 accompanies down-regulation of the survivin-1 homolog BIRC1 (NAIP) in glioblastoma multiforme (GBM). J. Neuro-Oncol. 2008, 91, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Silber, J.; Lim, D.A.; Petritsch, C.K.; Persson, A.I.; Maunakea, A.K.; Yu, M.; Vandenberg, S.R.; Ginzinger, D.G.; James, C.D.; Costello, J.F.; et al. miR-124 and miR-137 inhibit proliferation of glioblastoma multiforme cells and induce differentiation of brain tumor stem cells. BMC Med. 2008, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Kefas, B.; Godlewski, J.; Comeau, L.; Li, Y.; Abounader, R.; Hawkinson, M.; Lee, J.; Fine, H.; Chiocca, E.A.; Lawler, S.; et al. microRNA-7 inhibits the epidermal growth factor receptor and the akt pathway and is down-regulated in glioblastoma. Cancer Res. 2008, 68, 3566–3572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skog, J.; Würdinger, T.; Van Rijn, S.; Meijer, D.H.; Gainche, L.; Curry, W.T., Jr.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Roth, P.; Wischhusen, J.; Happold, C.; Chandran, P.A.; Hofer, S.; Eisele, G.; Weller, M.; Keller, A. A specific miRNA signature in the peripheral blood of glioblastoma patients. J. Neurochem. 2011, 118, 449–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teplyuk, N.M.; Mollenhauer, B.; Gabriely, G.; Giese, A.; Kim, E.; Smolsky, M.; Kim, R.Y.; Saria, M.G.; Pastorino, S.; Kesari, S.; et al. MicroRNAs in cerebrospinal fluid identify glioblastoma and metastatic brain cancers and reflect disease activity. Neuro-Oncol. 2012, 14, 689–700. [Google Scholar] [CrossRef] [Green Version]
- Regazzo, G.; Terrenato, I.; Spagnuolo, M.; Carosi, M.; Cognetti, G.; Cicchillitti, L.; Sperati, F.; Villani, V.; Carapella, C.; Piaggio, G.; et al. A restricted signature of serum miRNAs distinguishes glioblastoma from lower grade gliomas. J. Exp. Clin. Cancer Res. 2016, 35, 124. [Google Scholar] [CrossRef] [Green Version]
- D’Urso, P.I.; D’Urso, O.F.; Gianfreda, C.D.; Mezzolla, V.; Storelli, C.; Marsigliante, S. miR-15b and miR-21 as circulating biomarkers for diagnosis of glioma. Curr. Genom. 2015, 16, 304–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parvizhamidi, M.; Haddad, G.; Ostadrahimi, S.; Ostadrahimi, N.; Sadeghi, S.; Fayaz, S.; Fard-Esfahani, P. Circulating miR-26a and miR-21 as biomarkers for glioblastoma multiform. Biotechnol. Appl. Biochem. 2019, 66, 261–265. [Google Scholar] [CrossRef]
- Ebrahimkhani, S.; Vafaee, F.; Hallal, S.; Wei, H.; Lee, M.Y.T.; Young, P.E.; Satgunaseelan, L.; Beadnall, H.; Barnett, M.H.; Shivalingam, B.; et al. Deep sequencing of circulating exosomal microRNA allows non-invasive glioblastoma diagnosis. NPJ Precis. Oncol. 2018, 2, 28. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, W.; Yang, Y.; Lu, Y.; He, C.; Hu, G.; Liu, H.; Chen, J.; He, J.; Yu, H. MicroRNA-21 targets LRRFIP1 and contributes to VM-26 resistance in glioblastoma multiforme. Brain Res. 2009, 1286, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Ujifuku, K.; Mitsutake, N.; Takakura, S.; Matsuse, M.; Saenko, V.; Suzuki, K.; Hayashi, K.; Matsuo, T.; Kamada, K.; Nagata, I.; et al. miR-195, miR-455-3p and miR-10a* are implicated in acquired temozolomide resistance in glioblastoma multiforme cells. Cancer Lett. 2010, 296, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-M.; Huang, W.; Park, R.; Park, P.J.; Johnson, M.D. A developmental taxonomy of glioblastoma defined and maintained by MicroRNAs. Cancer Res. 2011, 71, 3387–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, S.; Patric, I.R.P.; Somasundaram, K. A ten-microRNA expression signature predicts survival in glioblastoma. PLoS ONE 2011, 6, e17438. [Google Scholar] [CrossRef] [Green Version]
- Lakomy, R.; Sana, J.; Hankeova, S.; Fadrus, P.; Kren, L.; Lzicarova, E.; Svoboda, M.; Dolezelova, H.; Smrcka, M.; Vyzula, R.; et al. MiR-195, miR-196b, miR-181c, miR-21 expression levels and O-6-methylguanine-DNA methyltransferase methylation status are associated with clinical outcome in glioblastoma patients. Cancer Sci. 2011, 102, 2186–2190. [Google Scholar] [CrossRef]
- Permuth-Wey, J.; Thompson, R.C.; Nabors, L.B.; Olson, J.J.; Browning, J.E.; Madden, M.H.; Chen, Y.A.; Egan, K.M. A functional polymorphism in the pre-miR-146a gene is associated with risk and prognosis in adult glioma. J. Neuro-Oncol. 2011, 105, 639–646. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Sun, L.; Wang, H.; Yao, J.; Jiang, C.; Xu, W.; Yang, Z. MiR-328 expression is decreased in high-grade gliomas and is associated with worse survival in primary glioblastoma. PLoS ONE 2012, 7, e47270. [Google Scholar] [CrossRef]
- Wei, X.; Chen, D.; Lv, T.; Li, G.; Qu, S. Serum MicroRNA-125b as a potential biomarker for glioma diagnosis. Mol. Neurobiol. 2014, 53, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Ho, H.-L.; Lin, S.-C.; Ho, T.D.-H.; Hsu, C.-Y. Upregulation of miR-125b, miR-181d, and miR-221 predicts poor prognosis in MGMT promoter-unmethylated glioblastoma patients. Am. J. Clin. Pathol. 2018, 149, 412–417. [Google Scholar] [CrossRef]
- D’Alessio, A.; Proietti, G.; Sica, G.; Scicchitano, B.M. Pathological and molecular features of glioblastoma and its peritumoral tissue. Cancers 2019, 11, 469. [Google Scholar] [CrossRef] [Green Version]
- Haehnel, S.; Reiche, K.; Loeffler, D.; Horn, A.; Blumert, C.; Puppel, S.-H.; Kaiser, N.; Rapp, F.; Rade, M.; Horn, F.; et al. Deep sequencing and automated histochemistry of human tissue slice cultures improve their usability as preclinical model for cancer research. Sci. Rep. 2019, 9, 19961. [Google Scholar] [CrossRef]
- Integration of Transcriptome and Proteome Profiles in Glioblastoma: Looking for the Missing Link. Available online: https://0-bmcmolbiol-biomedcentral-com.brum.beds.ac.uk/articles/10.1186/s12867-018-0115-6 (accessed on 30 November 2020).
- Kruthika, B.S.; Jain, R.; Arimappamagan, A.; Bharath, R.D.; Yasha, T.C.; Kondaiah, P.; Santosh, V. Transcriptome profiling reveals PDZ binding kinase as a novel biomarker in peritumoral brain zone of glioblastoma. J. Neuro-Oncol. 2018, 141, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Mangiola, A.; Saulnier, N.; De Bonis, P.; Orteschi, D.; Sica, G.; Lama, G.; Pettorini, B.L.; Sabatino, G.; Zollino, M.; Lauriola, L.; et al. Gene expression profile of glioblastoma peritumoral tissue: An ex vivo study. PLoS ONE 2013, 8, e57145. [Google Scholar] [CrossRef] [PubMed]
- Single-Cell RNA-Seq Analysis of Infiltrating Neoplastic Cells at the Migrating Front of Human Glioblastoma. Available online: https://www.biorxiv.org/content/10.1101/165811v1.full (accessed on 30 November 2020).
- Targeting of the Bmi-1 Oncogene/Stem Cell Renewal Factor by microRNA-128 Inhibits Glioma Proliferation and Self-Renewal. Available online: https://cancerres-aacrjournals-org.bibliopass.unito.it/content/68/22/9125.long (accessed on 30 November 2020).
- MicroRNA-10b is Overexpressed in Malignant Glioma and Associated with Tumor Invasive Factors, uPAR and RhoC. Available online: https://onlinelibrary-wiley-com.bibliopass.unito.it/doi/full/10.1002/ijc.24522 (accessed on 30 November 2020).
- Fazi, B.; Felsani, A.; Grassi, L.; Moles, A.; D’Andrea, D.; Toschi, N.; Sicari, D.; De Bonis, P.; Anile, C.; Guerrisi, M.G.; et al. The transcriptome and miRNome profiling of glioblastoma tissues and peritumoral regions highlights molecular pathways shared by tumors and surrounding areas and reveals differences between short-term and long-term survivors. Oncotarget 2015, 6, 22526–22552. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, H.-C.; Zhang, Y.; Huang, H. MicroRNA-625-3p inhibits gastric cancer metastasis through modulating EZH2. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1177–1185. [Google Scholar]
- Wang, M.; Li, C.; Nie, H.; Lv, X.; Qu, Y.; Yu, B.; Su, L.; Li, J.; Chen, X.; Ju, J.; et al. Down-regulated miR-625 suppresses invasion and metastasis of gastric cancer by targeting ILK. FEBS Lett. 2012, 586, 2382–2388. [Google Scholar] [CrossRef] [PubMed]
- Piwecka, M.; Rolle, K.; Belter, A.; Barciszewska, A.M.; Żywicki, M.; Michalak, M.; Nowak, S.; Naskręt-Barciszewska, M.Z.; Barciszewski, J. Comprehensive analysis of microRNA expression profile in malignant glioma tissues. Mol. Oncol. 2015, 9, 1324–1340. [Google Scholar] [CrossRef]
- Oligodendrocyte Progenitor Cells and Macrophages/Microglia Produce Glioma Stem Cell Niches at the Tumor Border. Available online: https://www-ncbi-nlm-nih-gov.bibliopass.unito.it/pmc/articles/PMC5952226/ (accessed on 30 November 2020).
- Van Der Vos, K.E.; Abels, E.R.; Zhang, X.; Lai, C.; Carrizosa, E.; Oakley, D.; Prabhakar, S.; Mardini, O.; Crommentuijn, M.H.W.; Skog, J.; et al. Directly visualized glioblastoma-derived extracellular vesicles transfer RNA to microglia/macrophages in the brain. Neuro-Oncology 2016, 18, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Battaglia, R.; Palini, S.; Vento, M.E.; La Ferlita, A.; Faro, M.J.L.; Caroppo, E.; Borzì, P.; Falzone, L.; Barbagallo, D.; Ragusa, M.; et al. Identification of extracellular vesicles and characterization of miRNA expression profiles in human blastocoel fluid. Sci. Rep. 2019, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Abels, E.R.; Maas, S.L.; Nieland, L.; Wei, Z.; Cheah, P.S.; Tai, E.; Kolsteeg, C.-J.; Dusoswa, S.A.; Ting, D.T.; Hickman, S.; et al. Glioblastoma-associated microglia reprogramming is mediated by functional transfer of extracellular miR-21. Cell Rep. 2019, 28, 3105–3119.e7. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-K.; Liu, H.-J.; Wang, Y.; Li, C.; Yang, J.-P.; Yang, L.; Qi, X.-J.; Zhao, Y.-L.; Shi, X.-F.; Li, J.-C.; et al. Exosomal miR-214-5p released from glioblastoma cells modulates inflammatory response of microglia after lipopolysaccharide stimulation through targeting CXCR5. CNS Neurol. Disord. Drug Targets 2019, 18, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Buonfiglioli, A.; Efe, I.E.; Guneykaya, D.; Ivanov, A.; Huang, Y.; Orlowski, E.; Krüger, C.; Deisz, R.A.; Markovic, D.; Flüh, C.; et al. Let-7 MicroRNAs regulate microglial function and suppress glioma growth through toll-like receptor. Cell Rep. 2019, 29, 3460–3471.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chirshev, E.; Oberg, K.C.; Ioffe, Y.J.; Unternaehrer, J.J. Let-7as biomarker, prognostic indicator, and therapy for precision medicine in cancer. Clin. Transl. Med. 2019, 8, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-R.; Luo, H.; Li, H.-L.; Cao, L.; Wang, X.-F.; Yan, W.; Wang, Y.-Y.; Zhang, J.-X.; Jiang, T.; Kang, C.-S.; et al. Overexpressed let-7a inhibits glioma cell malignancy by directly targeting K-ras, independently of PTEN. Neuro-Oncology 2013, 15, 1491–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthikeyan, A.; Gupta, N.; Tang, C.; Mallilankaraman, K.; Silambarasan, M.; Shi, M.; Lu, L.; Ang, B.T.; Ling, E.-A.; Dheen, S.T. Microglial SMAD4 regulated by microRNA-146a promotes migration of microglia which support tumor progression in a glioma environment. Oncotarget 2018, 9, 24950–24969. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Huang, S.; Yin, Z.; Zhu, J.; Ge, X.; Han, Z.; Tan, J.; Zhang, S.; Zhao, J.; Chen, F.; et al. Increases in miR-124-3p in microglial exosomes confer neuroprotective effects by targeting FIP200-mediated neuronal autophagy following traumatic brain injury. Neurochem. Res. 2019, 44, 1903–1923. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, J.-X.; Chen, W.-K.; Wu, G.-C.; Wang, Y.-Q.; Zhu, K.; Wang, J. miR-124/VAMP3 is a novel therapeutic target for mitigation of surgical trauma-induced microglial activation. Signal Transduct. Target. Ther. 2019, 4, 24. [Google Scholar] [CrossRef]
- MicroRNA-124 Promotes Microglia Quiescence and Suppresses EAE by Deactivating Macrophages via the C/EBP-α-PU.1 Pathway. Available online: https://www-nature-com.bibliopass.unito.it/articles/nm.2266 (accessed on 30 November 2020).
- Li, Z.; Zhang, J.; Zheng, H.; Li, C.; Xiong, J.; Wang, W.; Bao, H.; Jin, H.; Liang, P. Modulating lncRNA SNHG15/CDK6/miR-627 circuit by palbociclib, overcomes temozolomide resistance and reduces M2-polarization of glioma associated microglia in glioblastoma multiforme. J. Exp. Clin. Cancer Res. 2019, 38, 380. [Google Scholar] [CrossRef]
- Bier, A.; Hong, X.; Cazacu, S.; Goldstein, H.; Rand, D.; Xiang, C.; Jiang, W.; Ben-Asher, H.W.; Attia, M.; Brodie, A.; et al. miR-504 modulates the stemness and mesenchymal transition of glioma stem cells and their interaction with microglia via delivery by extracellular vesicles. Cell Death Dis. 2020, 11, 899. [Google Scholar] [CrossRef] [PubMed]
- Roh, T.H.; Kang, S.-G.; Moon, J.H.; Sung, K.S.; Park, H.H.; Kim, S.H.; Kim, E.H.; Hong, C.-K.; Suh, C.-O.; Chang, J.H. Survival benefit of lobectomy over gross-total resection without lobectomy in cases of glioblastoma in the noneloquent area: A retrospective study. J. Neurosurg. 2020, 132, 895–901. [Google Scholar] [CrossRef]
- Schneider, M.; Ilic, I.; Potthoff, A.-L.; Hamed, M.; Schäfer, N.; Velten, M.; Güresir, E.; Herrlinger, U.; Borger, V.; Vatter, H.; et al. Safety metric profiling in surgery for temporal glioblastoma: Lobectomy as a supra-total resection regime preserves perioperative standard quality rates. J. Neuro-Oncol. 2020, 149, 455–461. [Google Scholar] [CrossRef]
- Jiang, H.; Cui, Y.; Liu, X.; Ren, X.; Li, M.; Lin, S. Proliferation-dominant high-grade astrocytoma: Survival benefit associated with extensive resection of FLAIR abnormality region. J. Neurosurg. 2020, 132, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Zigiotto, L.; Annicchiarico, L.; Corsini, F.; Vitali, L.; Falchi, R.; Dalpiaz, C.; Rozzanigo, U.; Barbareschi, M.; Avesani, P.; Papagno, C.; et al. Effects of supra-total resection in neurocognitive and oncological outcome of high-grade gliomas comparing asleep and awake surgery. J. Neuro-Oncol. 2020, 148, 97–108. [Google Scholar] [CrossRef]
- Saavedra-López, E.; Roig-Martínez, M.; Cribaro, G.P.; Casanova, P.V.; Gallego, J.M.; Pérez-Vallés, A.; Barcia, C. Phagocytic glioblastoma-associated microglia and macrophages populate invading pseudopalisades. Brain Commun. 2020, 2, fcz043. [Google Scholar] [CrossRef] [Green Version]
- La Rocca, G.; Della Pepa, G.M.; Menna, G.; Altieri, R.; Ius, T.; Rapisarda, A.; Olivi, A.; Sabatino, G. State of the art of fluorescence guided techniques in neurosurgery. J. Neurosurg. Sci. 2020, 63, 619–624. [Google Scholar] [CrossRef] [PubMed]
- Stummer, W.; Stocker, S.; Wagner, S.; Stepp, H.; Fritsch, C.; Goetz, C.; Goetz, A.E.; Kiefmann, R.; Reulen, H.J. Intraoperative detection of malignant gliomas by 5-aminolevulinic acid-induced porphyrin fluorescence. Neurosurgery 1998, 42, 518–525, discussion 525–526. [Google Scholar]
- Lau, D.; Hervey-Jumper, S.L.; Chang, S.; Molinaro, A.M.; McDermott, M.W.; Phillips, J.J.; Berger, M.S. A prospective phase II clinical trial of 5-aminolevulinic acid to assess the correlation of intraoperative fluorescence intensity and degree of histologic cellularity during resection of high-grade gliomas. J. Neurosurg. 2016, 124, 1300–1309. [Google Scholar] [CrossRef] [Green Version]
- Eyüpoglu, I.Y.; Hore, N.; Merkel, A.; Buslei, R.; Buchfelder, M.; Savaskan, N. Supra-complete surgery via dual intraoperative visualization approach (DiVA) prolongs patient survival in glioblastoma. Oncotarget 2016, 7, 25755–25768. [Google Scholar] [CrossRef] [PubMed]
- La Rocca, G.; Sabatino, G.; Menna, G.; Altieri, R.; Ius, T.; Marchese, E.; Olivi, A.; Barresi, V.; Della Pepa, G.M. 5-Aminolevulinic acid false positives in cerebral neuro-oncology: Not all that is fluorescent is tumor. A case-based update and literature review. World Neurosurg. 2020, 137, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhang, X.; Wei, R.; Li, G.; Sun, B.; Zhang, H.; Liu, D.; Wang, C.; Feng, M. Engineering microglia as intraoperative optical imaging agent vehicles potentially for fluorescence-guided surgery in gliomas. Biomater. Sci. 2020, 8, 1117–1126. [Google Scholar] [CrossRef]
- Gutmann, D.H.; Kettenmann, H. Microglia/brain macrophages as central drivers of brain tumor pathobiology. Neuron 2019, 104, 442–449. [Google Scholar] [CrossRef]
- Markovic, D.; Vinnakota, K.; Van Rooijen, N.; Kiwit, J.; Synowitz, M.; Glass, R.; Kettenmann, H. Minocycline reduces glioma expansion and invasion by attenuating microglial MT1-MMP expression. Brain Behav. Immun. 2011, 25, 624–628. [Google Scholar] [CrossRef]
- Prionisti, I.; Bühler, L.H.; Walker, P.R.; Jolivet, R.B. Harnessing microglia and macrophages for the treatment of glioblastoma. Front. Pharmacol. 2019, 10, 506. [Google Scholar] [CrossRef] [PubMed]
- Lisi, L.; Laudati, E.; Navarra, P.; Russo, C.D. The mTOR kinase inhibitors polarize glioma-activated microglia to express a M1 phenotype. J. Neuroinflammation 2014, 11, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altieri, R.; Fontanella, M.; Agnoletti, A.; Panciani, P.P.; Spena, G.; Crobeddu, E.; Pilloni, G.; Tardico, V.; Lanotte, M.; Zenga, F.; et al. Role of nitric oxide in glioblastoma therapy: Another step to resolve the terrible puzzle? Transl. Med. UniSa 2014, 12, 54–59. [Google Scholar] [PubMed]
- Martins, T.A.; Schmassmann, P.; Shekarian, T.; Boulay, J.-L.; Ritz, M.-F.; Zanganeh, S.; Berg, J.V.; Hutter, G. Microglia-centered combinatorial strategies against glioblastoma. Front. Immunol. 2020, 11, 571951. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhang, B.; Zhou, L.; Shi, Y.; Li, Z.; Xia, Y.; Tian, J. Imaging dendrimer-grafted graphene oxide mediated Anti-miR-21 delivery with an activatable luciferase reporter. ACS Appl. Mater. Interfaces 2016, 8, 9014–9021. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.; Kavishwar, A.; Ghosh, S.K.; Barteneva, N.; Yigit, M.V.; Moore, A.; Medarova, Z. Detection of miRNA expression in intact cells using activatable sensor oligonucleotides. Chem. Biol. 2014, 21, 199–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, B.; Kavishwar, A.; Ross, A.; Pantazopoulos, P.; Moore, A.; Medarova, Z. In vivo detection of miRNA expression in tumors using an activatable nanosensor. Mol. Imaging Biol. 2015, 18, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| DE miRNA (GBM Core vs. Peritumoral Area) | Expression (GBM Core vs. Peritumoral Area) | Year of Publication | Technique Used to Assay miRNA Expression | Reference (PMID) |
|---|---|---|---|---|
| miR-10b; miR-130a; miR-221; miR-125b-1; miR-125b-2; miR9-2; miR-21; miR-25; miR-123 | Upregulated | 2005 | Microarray | 16039986 |
| miR-128a; miR-181c; miR-181a; miR-181b | Downregulated | 2005 | Microarray | 16039986 |
| miR-21; miR-10b | Upregulated | 2008; 2009 | Microarray | 16039986; 19536818 |
| miR-21-3p; miR-196b-5p; miR-135b-5p; miR-183-3p | Upregulated | 2015 | SAGE sequencing and qRT-PCR | 26188123 |
| miR-219a; miR-338-3p; miR-338-5p; miR-34b; miR-34c | Downregulated | 2015 | SAGE sequencing and qRT-PCR | 26188123 |
| miR-625 | Downregulated in peritumoral area vs healthy white matter | 2015 | Microarray; small RNA deep sequencing | 25864039 |
| miR-219-5p; miR-219-2-3p; miR-338-3p; miR-27b; miR-23b | Downregulated | 2018 | Microarray | 29559295 |
| miR-630; miR-1246; miR-642b; miR-1181; hsv-miR-H18; miR-3195; miR-3663-3p | Upregulated | 2018 | Microarray | 29559295 |
| miRNA Involved in the Cross-Talk between GBM Cells and Microglia | Functional Involvement of miRNA in the Cross-Talk | Year of Publication | Validated Target (Official Gene Symbol) | Technique Used to Assay miRNA Expression | Reference (PMID) |
|---|---|---|---|---|---|
| miR-21 and miR-451 | Internalization of miR-21 and miR-451, two known oncogenic miRNAs enriched within GBM extracellular vesicles (EVs), into microglial cells. The consequence is a switch of microglial cells versus a tumor-supportive phenotype through the secretion of immunosuppressive cytokines | 2016 | MYC | qRT-PCR | 26433199 |
| miR-21 | GBM EV-mediated transport of miR-21 exerted the downregulation of the mRNA transcribed by the BTG anti-proliferation factor 2 (Btg2) gene, thus stimulating microglial cell proliferation | 2019 | BTG | qRT-PCR; ddPCR | 31533034 |
| miR-214-5p | MiR-214-5p, aberrantly upregulated in GBM cells, can be transferred to microglia, through exosomes, contributing to the suppression of microglial C-X-C motif chemokine receptor 5 (CXCR5), and, consequently, increasing the microglial secretion of inflammatory cytokines IL-6, IL-8 and TNF-α | 2019 | CXCR5 | qRT-PCR | 30394221 |
| let-7 UUGU motif | Let-7 family members containing UUGU motif determined the activation of an M1-like microglial phenotype, through the interaction with the Toll-like receptor 7 (TLR7), which can trigger an anti-tumoral microenvironment at the periphery of the GBM cell mass | 2019 | Physical interaction and activation of TLR7 in microglial cells | qRT-PCR | 31825829 |
| miR-146a | Microglial cells exposed to GBM conditioned-medium exhibited a greater ability to migrate. This was linked to downregulation of miR-146a and upregulation of its target SMAD family member 4 (SMAD4) | 2018 | SMAD4 | qRT-PCR | 29861845 |
| miR-124-3p | MiR-124-3p is a brain-specific miRNA, downregulated both in activated microglia and in GBM cells. MiR-124-3p was demonstrated to contribute to communication between microglial and neuronal cells via microglial exosomes | 2019 | N/A | qRT-PCR | 31190315 |
| miR-627 | Temozolomide-resistant GBM cells trigger the M2-polarization of microglial cells thanks to the long-noncoding RNA SNHG15 (upregulated in GBM cells) and its associated molecular axis made up of miR-627 (tumor suppressor, normally downregulated in GBM) and CDK6 (oncoprotein, directly targeted by miR-627) | 2019 | CDK6 | qRT-PCR | 31462285 |
| miR-504 | MiR-504, normally downregulated in both GBM and glioma stem cells, compared with healthy white matter, may be transferred to microglial cells, allowing their M1 polarization, thanks to EV cargo | 2020 | N/A | Microarray; qRT-PCR | 33093452 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altieri, R.; Barbagallo, D.; Certo, F.; Broggi, G.; Ragusa, M.; Di Pietro, C.; Caltabiano, R.; Magro, G.; Peschillo, S.; Purrello, M.; et al. Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia–Glioma Cross-Talk. Brain Sci. 2021, 11, 200. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020200
Altieri R, Barbagallo D, Certo F, Broggi G, Ragusa M, Di Pietro C, Caltabiano R, Magro G, Peschillo S, Purrello M, et al. Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia–Glioma Cross-Talk. Brain Sciences. 2021; 11(2):200. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020200
Chicago/Turabian StyleAltieri, Roberto, Davide Barbagallo, Francesco Certo, Giuseppe Broggi, Marco Ragusa, Cinzia Di Pietro, Rosario Caltabiano, Gaetano Magro, Simone Peschillo, Michele Purrello, and et al. 2021. "Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia–Glioma Cross-Talk" Brain Sciences 11, no. 2: 200. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020200