
Investigating the Intervention Parameters of Endogenous Paired Associative Stimulation (ePAS)

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
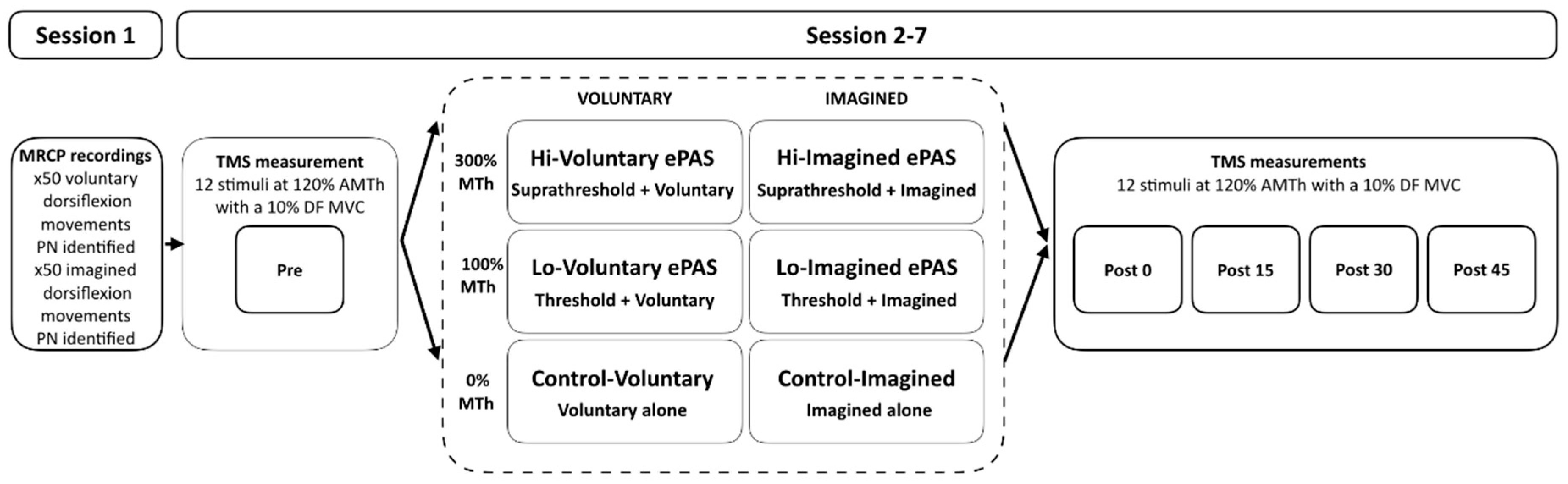
2.1. Study Design
2.2. Sample Size Calculation
2.3. Participants
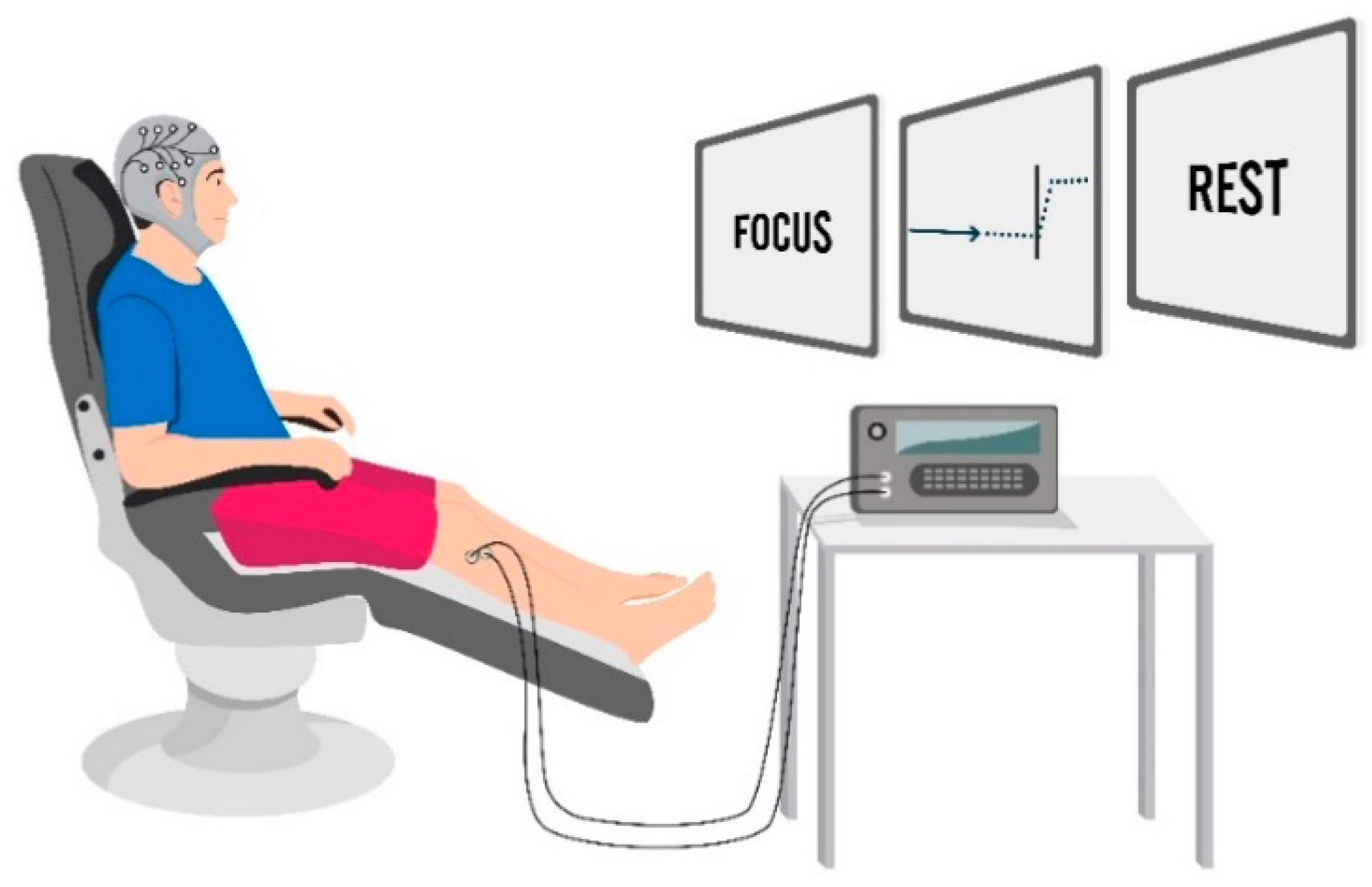
2.4. Experimental Set-Up and Procedures
2.4.1. Session 1
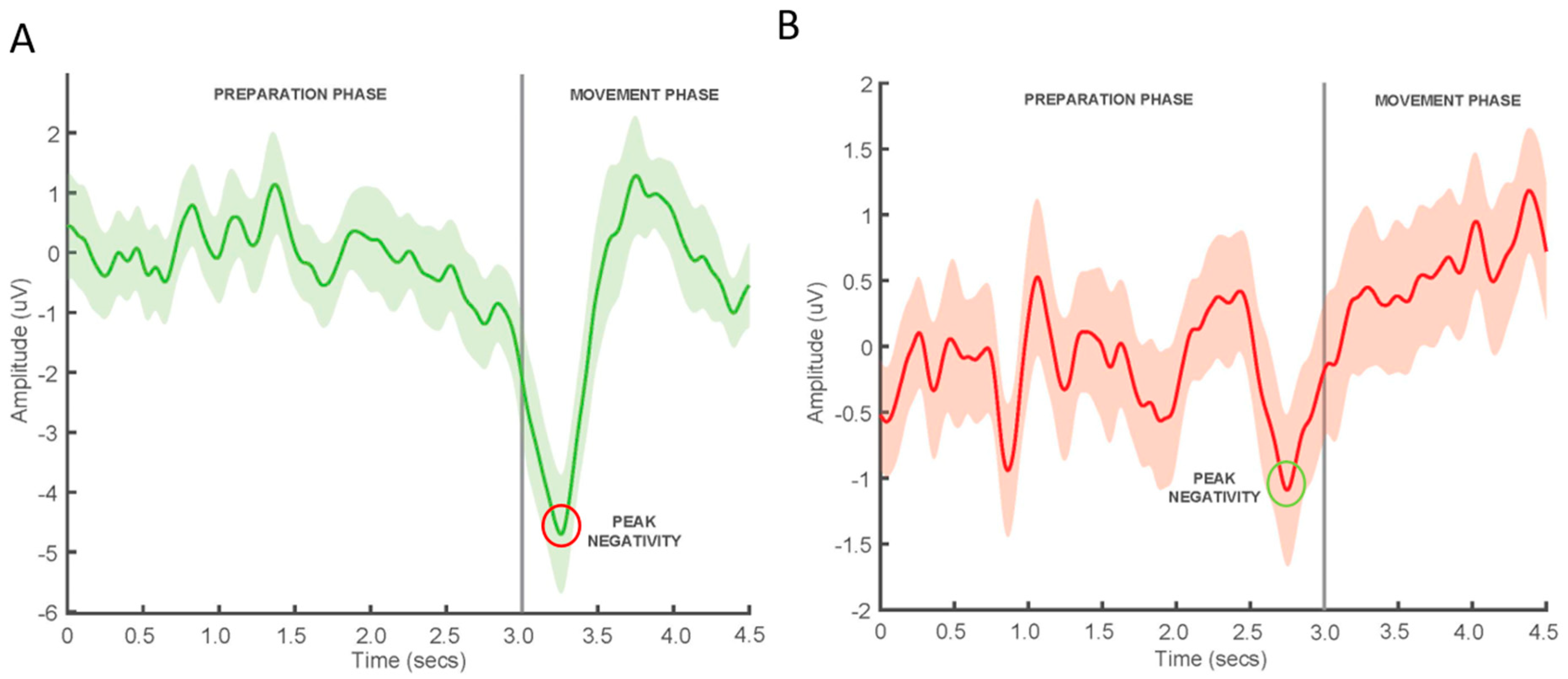
MRCP Recordings
MRCP Feature Extraction
2.4.2. Sessions 2–7
Outcome Measure—Corticomotor Excitability (CME)
ePAS Interventions
Control Interventions
2.5. Data Processing and Analysis
2.6. Statistical Analysis
3. Results
3.1. Participant Characteristics
3.1.1. MRCP
3.1.2. PES Intensities (mA)
3.1.3. Baseline Corticomotor Excitability (CME)
3.2. Study Findings
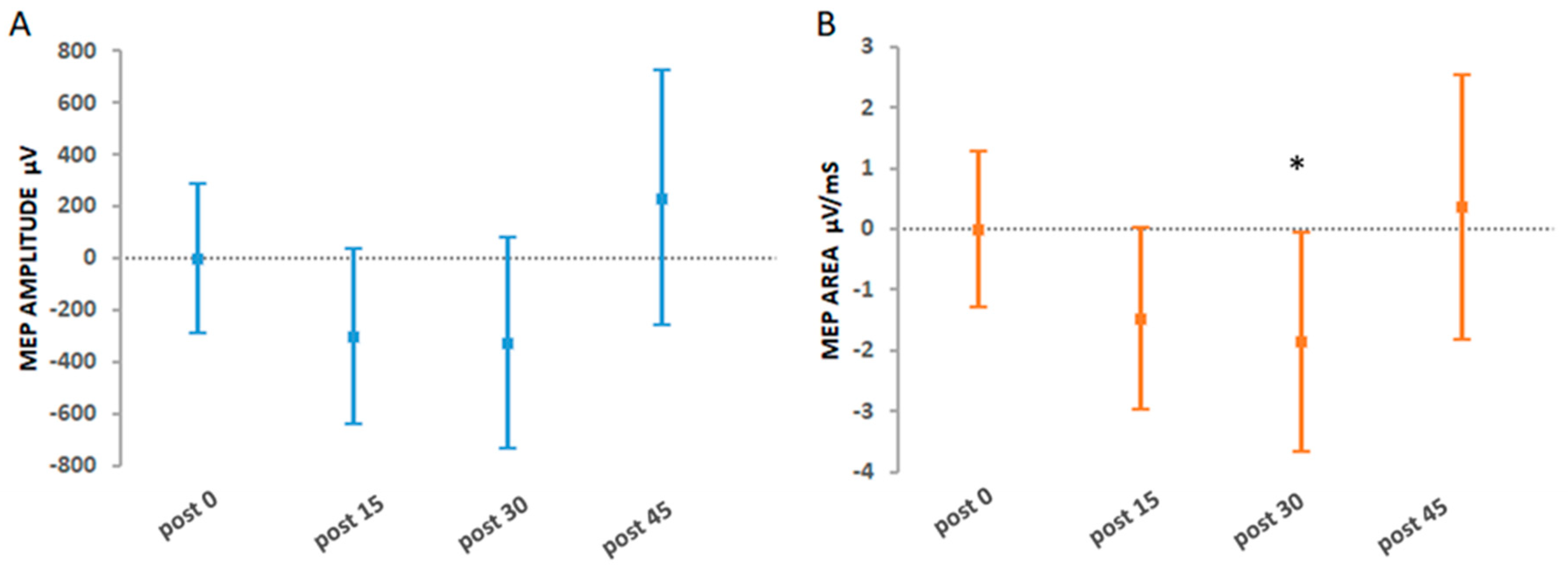
3.2.1. Primary Findings
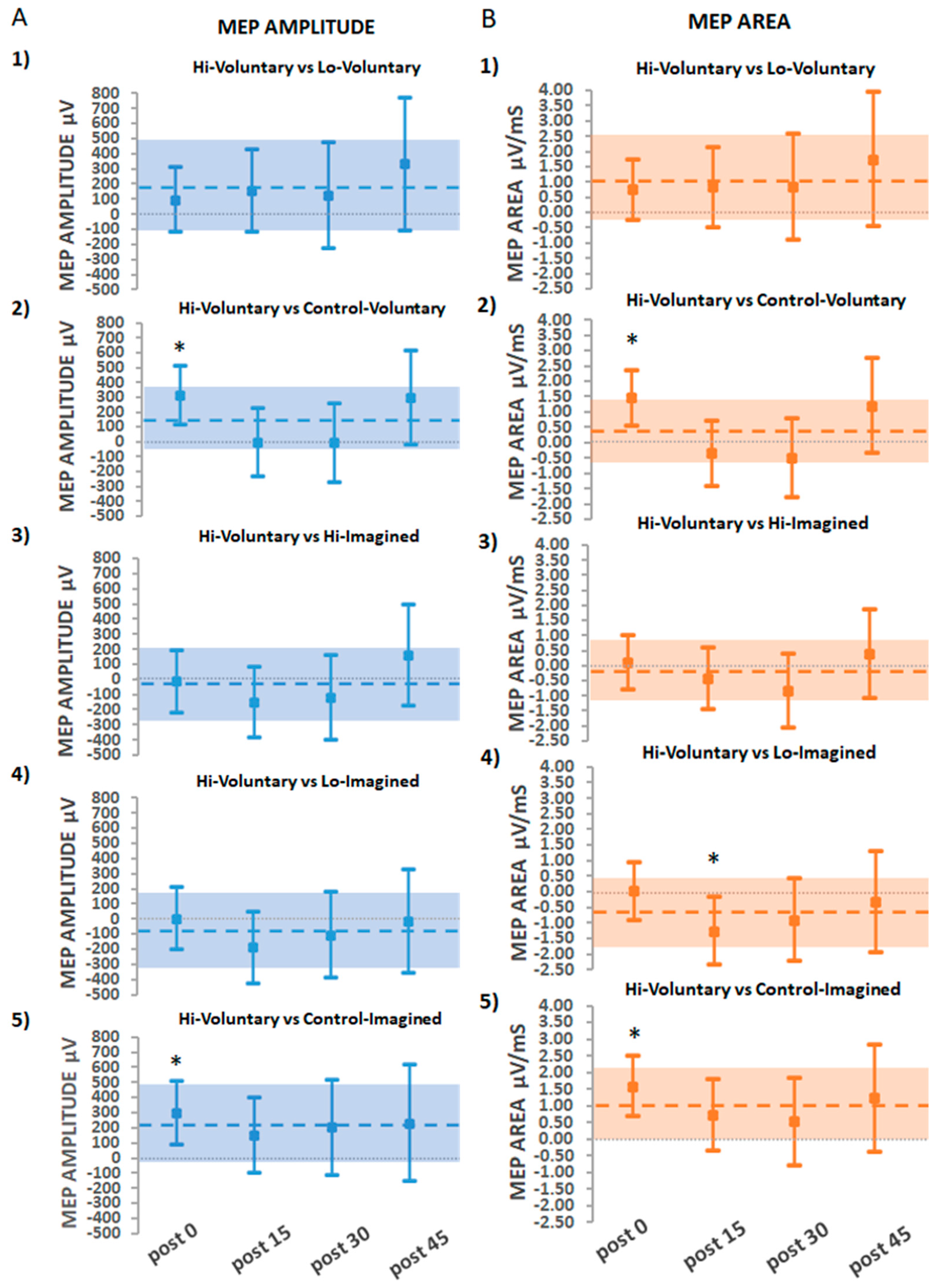
3.2.2. Secondary Findings: The Effects of Hi-Voluntary Compared to All Other Interventions
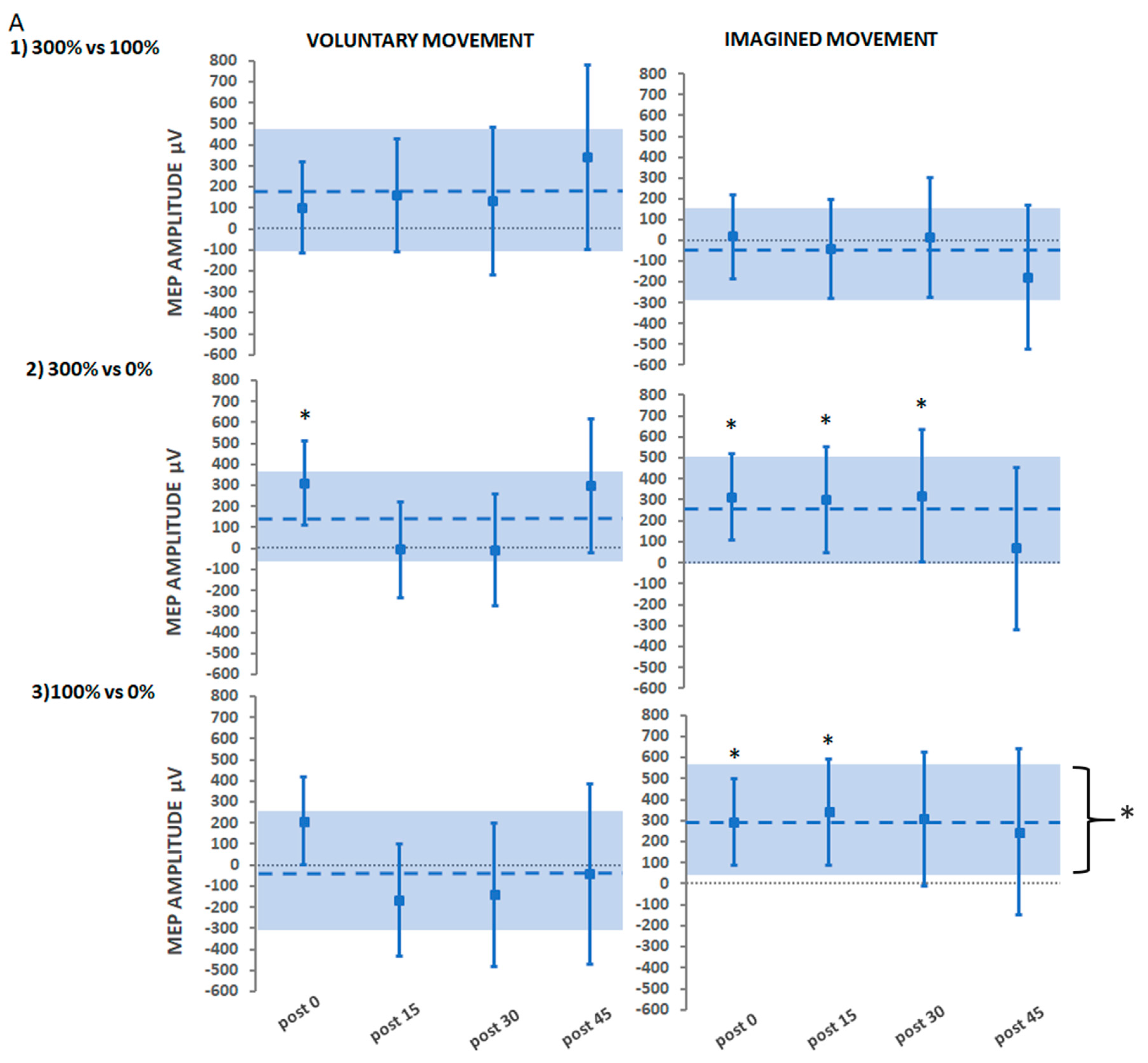
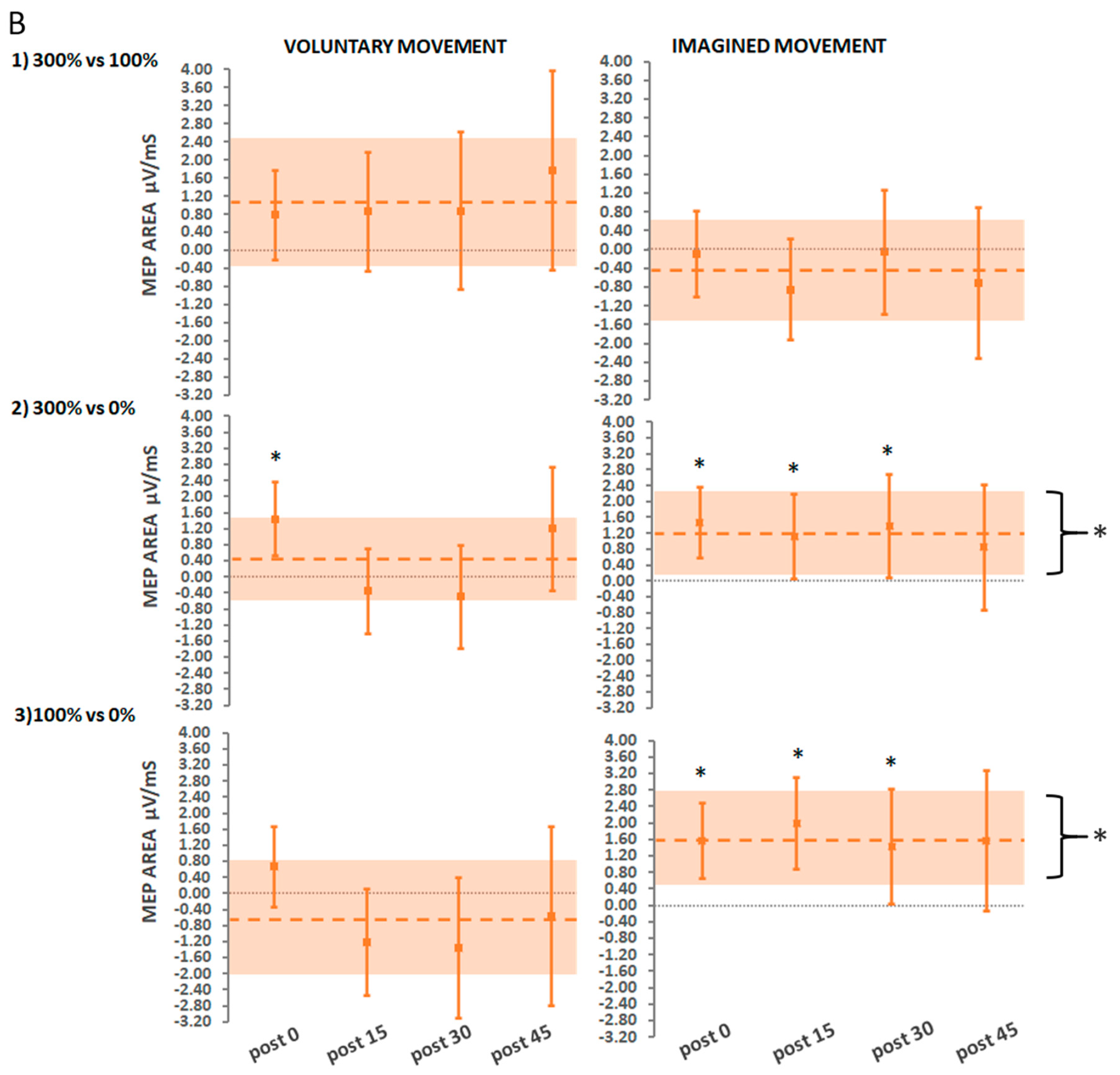
3.2.3. Secondary Findings: Stimulation Intensity
Suprathreshold Stimulation (Hi) vs. Threshold Stimulation (Lo)
Suprathreshold Stimulation vs. No Stimulation
Threshold Stimulation vs. No Stimulation
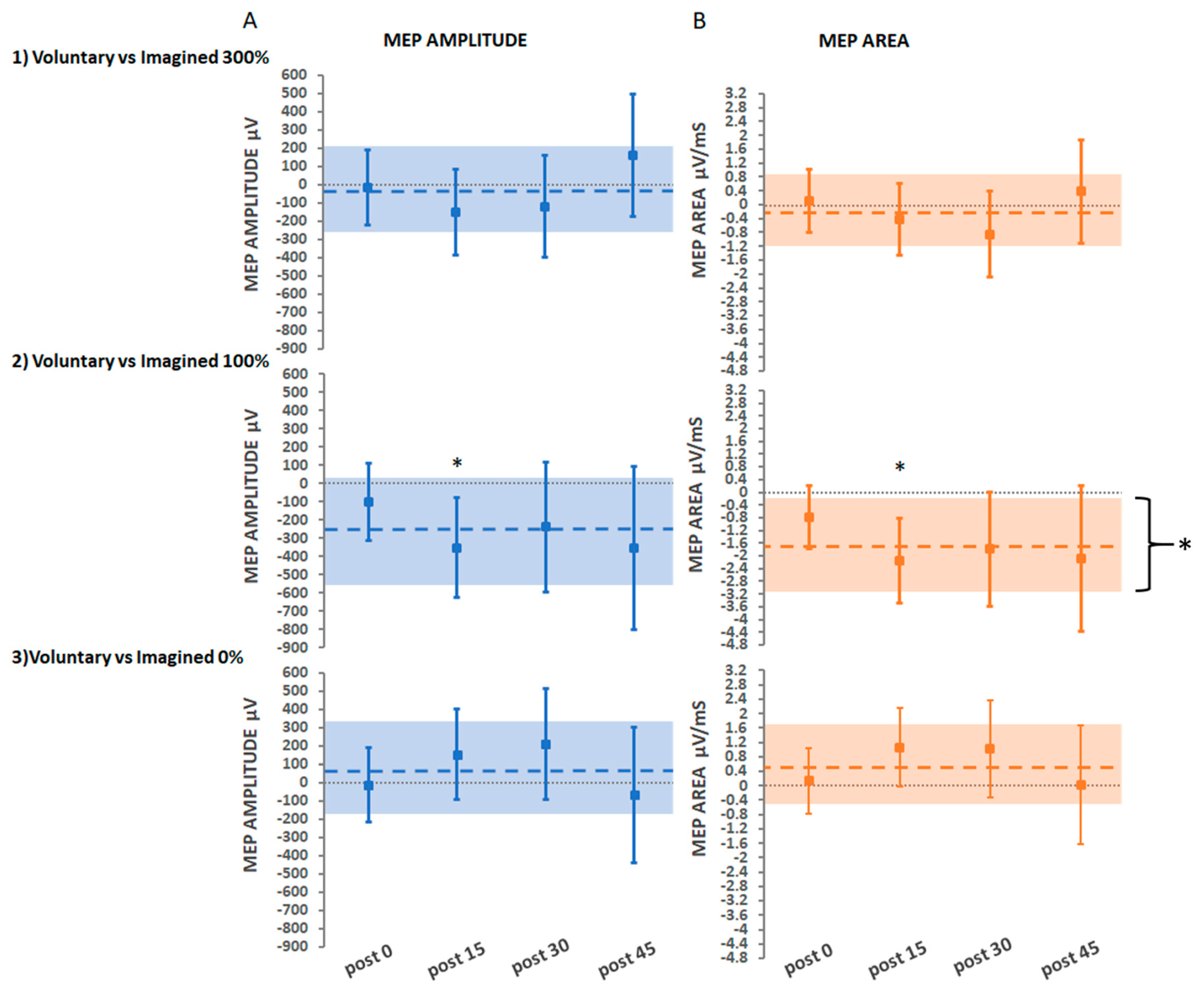
3.2.4. Secondary Findings: Movement Type
Voluntary vs. Imagined Movement at Suprathreshold Stimulation
Voluntary vs. Imagined Movement at Threshold Stimulation
Voluntary vs. Imagined Movement with No Stimulation
4. Discussion
4.1. ePAS Intervention Efficacy
4.2. Hi-Voluntary ePAS
4.3. Stimulation Intensity
4.4. Movement Type
4.5. Strength and Limitations
4.6. Future Recommendations
- Factorial designs should be used to explore the interaction effects of different intervention parameters in neuromodulatory interventions; parameters could include stimulation intensity, movement type, ISI, and the number of stimuli.
- To support translation into clinical practice, this work and similar factorial designs should be undertaken in people with stroke. These studies should not only explore the neurophysiological effects of the intervention but also assess changes in impairment and functional outcomes.
- Using TMS measurements from both resting and active muscle would allow comparison to previous work and enable the researcher to consider the impact of measurement error on the findings.
- The baseline covariates AMTh and MVC were considered and suitably adjusted for in the factorial models for the primary analysis of this study. Future research should also consider interactions with baseline covariates as part of a pre-planned blind model selection process. This may shed light on how baseline covariates modulate responses to neuromodulatory interventions.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ayache, S.S.; Farhat, W.H.; Zouari, H.G.; Hosseini, H.; Mylius, V.; Lefaucheur, J.-P. Stroke rehabilitation using noninvasive cortical stimulation: Motor deficit. Exp. Rev. Neurother. 2012, 12, 949–972. [Google Scholar] [CrossRef] [PubMed]
- Knotkova, H.; Rasche, D. Textbook of Neuromodulation; Spinger-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Hebb, D.O. The organization of behavior: A neuropsycholocigal theory. In A Wiley Book in Clinical Psychology; Wiley: New York, NY, USA, 1949; pp. 62–78. [Google Scholar]
- Thickbroom, G.W. Transcranial magnetic stimulation and synaptic plasticity: Experimental framework and human models. Exp. Brain Res. 2007, 180, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Stefan, K.; Kunesch, E.; Cohen, L.G.; Benecke, R.; Classen, J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain J. Neurol. 2000, 123, 572–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolters, A.; Sandbrink, F.; Schlottmann, A.; Kunesch, E.; Stefan, K.; Cohen, L.G.; Benecke, R.; Classen, J. A temporally asymmetric hebbian rule governing plasticity in the human motor cortex. J. Neurophysiol. 2003, 89, 2339–2345. [Google Scholar] [CrossRef] [PubMed]
- Carson, R.G.; Kennedy, N.C. Modulation of human corticospinal excitability by paired associative stimulation. Front. Hum. Neurosci. 2013, 7, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suppa, A.; Quartarone, A.; Siebner, H.; Chen, R.; Di Lazzaro, V.; Del Giudice, P.; Paulus, W.; Rothwell, J.; Ziemann, U.; Classen, J. The associative brain at work: Evidence from paired associative stimulation studies in humans. Clin. Neurophysiol. 2017, 128, 2140–2164. [Google Scholar] [CrossRef]
- Alder, G.; Signal, N.; Olsen, S.; Taylor, D. A Systematic Review of Paired Associative Stimulation (PAS) to Modulate Lower Limb Corticomotor Excitability: Implications for Stimulation Parameter Selection and Experimental Design. Front. Neurosci. 2019, 13, 895. [Google Scholar] [CrossRef] [Green Version]
- Bi, G.-Q.; Poo, M.-M. Synaptic modifications in cultured hippocampal neurons: Dependence on spike timing, synaptic strength, and postsynaptic cell type. J. Neurosci. 1998, 18, 10464–10472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.I.; Tao, H.W.; Holt, C.E.; Harris, W.A.; Poo, M.-M. A critical window for cooperation and competition among developing retinotectal synapses. Nature 1998, 395, 37. [Google Scholar] [CrossRef] [PubMed]
- Jacob, V.; Brasier, D.J.; Erchova, I.; Feldman, D.; Shulz, D.E. Spike timing-dependent synaptic depression in the in vivo barrel cortex of the rat. J. Neurosci. 2007, 27, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Magee, J.C.; Johnston, D. A synaptically controlled, associative signal for Hebbian plasticity in hippocampal neurons. Science 1997, 275, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Markram, H.; Lübke, J.; Frotscher, M.; Sakmann, B. Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 1997, 275, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, H.; Hallett, M. What is the Bereitschaftspotential? Clin. Neurophysiol. 2006, 117, 2341–2356. [Google Scholar] [CrossRef] [PubMed]
- Jankelowitz, S.K.; Colebatch, J.G. Movement-related potentials associated with self-paced, cued and imagined arm movements. Exp. Brain Res. 2002, 147, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Walter, W.; Cooper, R.; Aldridge, V.; McCallum, W.; Winter, A. Contingent negative variation: An electric sign of sensori-motor association and expectancy in the human brain. Nature 1964, 203, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Kornhuber, H.H.; Deecke, L. [Changes in the brain potential in voluntary movements and passive movements in man: Readiness potential and reafferent potentials]. Pflug. Arch. Gesamte Physiol Menschen Tiere 1965, 284, 1–17. [Google Scholar] [CrossRef]
- Brunia, C.H.M.; van Boxtel, G.J.M.; Speelman, J.D. The Bilateral Origin of Movement-Related Potentials Preceding Unilateral Actions. J. Psychophysiol. 2004, 18, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Toma, K.; Matsuoka, T.; Immisch, I.; Mima, T.; Waldvogel, D.; Koshy, B.; Hanakawa, T.; Shill, H.; Hallett, M. Generators of movement-related cortical potentials: fMRI-constrained EEG dipole source analysis. Neuroimage 2002, 17, 161–173. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.K.; Arai, N.; Tsai, C.H.; Ziemann, U. Movement related cortical potentials of cued versus self-initiated movements: Double dissociated modulation by dorsal premotor cortex versus supplementary motor area rTMS. Hum. Brain Mapp. 2012, 33, 824–839. [Google Scholar] [CrossRef] [PubMed]
- Mrachacz-Kersting, N.; Kristensen, S.R.; Niazi, I.K.; Farina, D. Precise temporal association between cortical potentials evoked by motor imagination and afference induces cortical plasticity. J. Physiol. 2012, 590, 1669–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niazi, I.K.; Mrachacz-Kersting, N.; Jiang, N.; Dremstrup, K.; Farina, D. Peripheral electrical stimulation triggered by self-paced detection of motor intention enhances motor evoked potentials. IEEE Trans Neural Syst. Rehabil. Eng. 2012, 20, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Mrachacz-Kersting, N.; Voigt, M.; Stevenson, A.J.T.; Aliakbaryhosseinabadi, S.; Jiang, N.; Dremstrup, K.; Farina, D. The effect of type of afferent feedback timed with motor imagery on the induction of cortical plasticity. Brain Res. 2017, 1674, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, S.; Signal, N.; Niazi, I.K.; Christensen, T.; Jochumsen, M.; Taylor, D. Paired Associative Stimulation Delivered by Pairing Movement-Related Cortical Potentials with Peripheral Electrical Stimulation: An Investigation of the Duration of Neuromodulatory Effects. Neuromodulation J. Int. Neuromodulation Soc. 2018, 21, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Mrachacz-Kersting, N.; Aliakbaryhosseinabadi, S. Comparison of the Efficacy of a Real-Time and Offline Associative Brain-Computer-Interface. Front. Neurosci. 2018, 12, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochumsen, M.; Navid, M.S.; Rashid, U.; Haavik, H.; Niazi, I.K. EMG-versus EEG-Triggered Electrical Stimulation for Inducing Corticospinal Plasticity. IEEE Trans. Neural Syst. Rehabil. Eng. 2019, 27, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Mrachacz-Kersting, N.; Stevenson, A.J.T.; Jorgensen, H.R.M.; Severinsen, K.E.; Aliakbaryhosseinabadi, S.; Jiang, N.; Farina, D. Brain state-dependent stimulation boosts functional recovery following stroke. Ann. Neurol. 2019, 85, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Mrachacz-Kersting, N.; Jiang, N.; Stevenson, A.J.T.; Niazi, I.K.; Kostic, V.; Pavlovic, A.; Radovanovic, S.; Djuric-Jovicic, M.; Agosta, F.; Dremstrup, K. Efficient neuroplasticity induction in chronic stroke patients by an associative brain-computer interface. J. Neurophysiol. 2016, 115, 1410–1421. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.; Signal, N.; Niazi, I.K.; Rashid, U.; Alder, G.; Mawston, G.; Nedergaard, R.B.; Jochumsen, M.; Taylor, D. Peripheral Electrical Stimulation Paired with Movement-Related Cortical Potentials Improves Isometric Muscle Strength and Voluntary Activation Following Stroke. Front. Hum. Neurosci. 2020, 14, 156. [Google Scholar] [CrossRef] [PubMed]
- Jochumsen, M.; Signal, N.; Nedergaard, R.W.; Taylor, D.; Haavik, H.; Niazi, I.K. Induction of Long-term Depression-like Plasticity by Pairings of Motor Imagination and Peripheral Electrical Stimulation. Front. Hum. Neurosci. 2015, 9, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochumsen, M.; Niazi, I.K.; Signal, N.; Nedergaard, R.W.; Holt, K.; Haavik, H.; Taylor, D. Pairing voluntary movement and muscle-located electrical stimulation increases cortical excitability. Front. Hum. Neurosci. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochumsen, M.; Cremoux, S.; Robinault, L.; Lauber, J.; Arceo, J.C.; Navid, M.S.; Nedergaard, R.W.; Rashid, U.; Haavik, H.; Niazi, I.K. Investigation of Optimal Afferent Feedback Modality for Inducing Neural Plasticity with A Self-Paced Brain-Computer Interface. Sensors (BaselSwitz.) 2018, 18, 3761. [Google Scholar] [CrossRef] [Green Version]
- Chipchase, L.S.; Schabrun, S.M.; Hodges, P.W. Peripheral electrical stimulation to induce cortical plasticity: A systematic review of stimulus parameters. Clin. Neurophysiol. 2011, 122, 456–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.V.; Alon, G.; Roys, S.R.; Gullapalli, R.P. Functional MRI determination of a dose-response relationship to lower extremity neuromuscular electrical stimulation in healthy subjects. Exp. Brain Res. 2003, 150, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, K.; Tanabe, S.; Higashi, T.; Tsurumi, T.; Kasai, T. Changes of excitability in M1 induced by neuromuscular electrical stimulation differ between presence and absence of voluntary drive. Int. J. Rehabil. Res. 2011, 34, 100–109. [Google Scholar] [CrossRef]
- Chipchase, L.S.; Schabrun, S.M.; Hodges, P.W. Corticospinal excitability is dependent on the parameters of peripheral electric stimulation: A preliminary study. Arch. Phys. Med. Rehabil. 2011, 92, 1423–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardwick, R.M.; Caspers, S.; Eickhoff, S.B.; Swinnen, S.P. Neural correlates of action: Comparing meta-analyses of imagery, observation, and execution. Neurosci. Biobehav. Rev. 2018, 94, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Eagles, J.S.; Carlsen, A.N.; MacKinnon, C.D. Neural processes mediating the preparation and release of focal motor output are suppressed or absent during imagined movement. Exp. Brain Res. 2015, 233, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Jeannerod, M.; Decety, J. Mental motor imagery: A window into the representational stages of action. Curr. Opin. Neurobiol. 1995, 5, 727–732. [Google Scholar] [CrossRef]
- Angelini, M.; Calbi, M.; Ferrari, A.; Sbriscia-Fioretti, B.; Franca, M.; Gallese, V.; Umilta, M.A. Motor Inhibition during Overt and Covert Actions: An Electrical Neuroimaging Study. PLoS ONE 2015, 10, e0126800. [Google Scholar] [CrossRef]
- Yilmaz, O.; Cho, W.; Braun, C.; Birbaumer, N.; Ramos-Murguialday, A. Movement related cortical potentials in severe chronic stroke. In Proceedings of the 35th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Osaka, Japan, 3–7 July 2013; pp. 2216–2219. [Google Scholar]
- Do Nascimento, O.F.; Nielsen, K.D.; Voigt, M. Movement-related parameters modulate cortical activity during imaginary isometric plantar-flexions. Exp. Brain Res. 2006, 171, 78–90. [Google Scholar] [CrossRef]
- Hanakawa, T.; Dimyan, M.A.; Hallett, M. Motor Planning, Imagery, and Execution in the Distributed Motor Network: A Time-Course Study with Functional MRI. Cereb. Cortex 2008, 18, 2775–2788. [Google Scholar] [CrossRef] [Green Version]
- Gerardin, E.; Sirigu, A.; Lehéricy, S.; Poline, J.B.; Gaymard, B.; Marsault, C.; Agid, Y.; Le Bihan, D. Partially overlapping neural networks for real and imagined hand movements. Cereb. Cortex (New York, N.Y. 1991) 2000, 10, 1093–1104. [Google Scholar] [CrossRef]
- Dechent, P.; Merboldt, K.D.; Frahm, J. Is the human primary motor cortex involved in motor imagery? Cogn. Brain Res. 2004, 19, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Hanakawa, T.; Immisch, I.; Toma, K.; Dimyan, M.A.; Van Gelderen, P.; Hallett, M. Functional properties of brain areas associated with motor execution and imagery. J. Neurophysiol. 2003, 89, 989–1002. [Google Scholar] [CrossRef] [Green Version]
- Avanzino, L.; Gueugneau, N.; Bisio, A.; Ruggeri, P.; Papaxanthis, C.; Bove, M. Motor cortical plasticity induced by motor learning through mental practice. Front. Behav Neurosci. 2015, 9, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonassi, G.; Biggio, M.; Bisio, A.; Ruggeri, P.; Bove, M.; Avanzino, L. Provision of somatosensory inputs during motor imagery enhances learning-induced plasticity in human motor cortex. Sci. Rep. 2017, 7, 9300. [Google Scholar] [CrossRef] [PubMed]
- Dobkin, B.H. Training and exercise to drive poststroke recovery. Nat. Clin. Pract. Neurol. 2008, 4, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Mrachacz-Kersting, N.; Fong, M.; Murphy, B.A.; Sinkjaer, T. Changes in excitability of the cortical projections to the human tibialis anterior after paired associative stimulation. J. Neurophysiol 2007, 97, 1951–1958. [Google Scholar] [CrossRef] [PubMed]
- Barsi, G.I.; Popovic, D.B.; Tarkka, I.M.; Sinkjaer, T.; Grey, M.J. Cortical excitability changes following grasping exercise augmented with electrical stimulation. Exp. Brain Res. 2008, 191, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Khaslavskaia, S.; Sinkjaer, T. Motor cortex excitability following repetitive electrical stimulation of the common peroneal nerve depends on the voluntary drive. Exp. Brain Res. 2005, 162, 497–502. [Google Scholar] [CrossRef]
- Thompson, A.K.; Stein, R.B. Short-term effects of functional electrical stimulation on motor-evoked potentials in ankle flexor and extensor muscles. Exp. Brain Res. 2004, 159, 491–500. [Google Scholar] [CrossRef]
- Baker, T.B.; Smith, S.S.; Bolt, D.M.; Loh, W.-Y.; Mermelstein, R.; Fiore, M.C.; Piper, M.E.; Collins, L.M. Implementing Clinical Research Using Factorial Designs: A Primer. Behav. Ther. 2017, 48, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A.; Safety of TMS Consensus Group. Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochumsen, M.; Niazi, I.K.; Mrachacz-Kersting, N.; Jiang, N.; Farina, D.; Dremstrup, K. Comparison of spatial filters and features for the detection and classification of movement-related cortical potentials in healthy individuals and stroke patients. J. Neural Eng. 2015, 12, 056003. [Google Scholar] [CrossRef]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.; George, M. Non-invasive electrical and magnetic stimulation of the brain, spinal cord, roots and peripheral nerves: Basic principles and procedures for routine clinical and research application. An updated report from an IFCN Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef]
- Hermens, H.J.; Freriks, B.; Disselhorst-Klug, C.; Rau, G. Development of recommendations for SEMG sensors and sensor placement procedures. J. Electromyogr. Kinesiol. 2000, 10, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.N.; Signal, N.; Taylor, D. Reliability of lower limb motor evoked potentials in stroke and healthy populations: How many responses are needed? Clin. Neurophysiol. 2014, 125, 748–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.R.; Kim, J.H.; Lim, J.Y. Optimization of facilitation related to threshold in transcranial magnetic stimulation. Clin. Neurophysiol. 2001, 112, 593–599. [Google Scholar] [CrossRef]
- Groppa, S.; Oliviero, A.; Eisen, A.; Quartarone, A.; Cohen, L.G.; Mall, V.; Kaelin-Lang, A.; Mima, T.; Rossi, S.; Thickbroom, G.W.; et al. A practical guide to diagnostic transcranial magnetic stimulation: Report of an IFCN committee. Clin. Neurophysiol. 2012, 123, 858–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cacchio, A.; Cimini, N.; Alosi, P.; Santilli, V.; Marrelli, A. Reliability of transcranial magnetic stimulation-related measurements of tibialis anterior muscle in healthy subjects. Clin. Neurophysiol. 2009, 120, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Cacchio, A.; Paoloni, M.; Cimini, N.; Mangone, M.; Liris, G.; Aloisi, P.; Santilli, V.; Marrelli, A. Reliability of TMS-related measures of tibialis anterior muscle in patients with chronic stroke and healthy subjects. J. Neurol. Sci. 2011, 303, 90–94. [Google Scholar] [CrossRef]
- Chipchase, L.; Schabrun, S.; Cohen, L.; Hodges, P.; Ridding, M.; Rothwell, J.; Taylor, J.; Ziemann, U. A checklist for assessing the methodological quality of studies using transcranial magnetic stimulation to study the motor system: An international consensus study. Clin. Neurophysiol. 2012, 123, 1698–1704. [Google Scholar] [CrossRef] [Green Version]
- Bergquist, A.; Clair, J.; Lagerquist, O.; Mang, C.; Okuma, Y.; Collins, D. Neuromuscular electrical stimulation: Implications of the electrically evoked sensory volley. Eur. J. Appl. Physiol. 2011, 111, 2409–2426. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, M.N.; Ridding, M.C.; Miles, T.S. Do alternate methods of analysing motor evoked potentials give comparable results? J. Neurosci. Methods 2004, 136, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Yamaguchi, T.; Yoshida, N.; Tanabe, S.; Kondo, K.; Sugawara, K. Combined effect of motor imagery and peripheral nerve electrical stimulation on the motor cortex. Exp. Brain Res. 2013, 227, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Boisgontier, M.P.; Cheval, B. The anova to mixed model transition. Neurosci. Biobehav. Rev. 2016, 68, 1004–1005. [Google Scholar] [CrossRef] [PubMed]
- Lumley, T.; Diehr, P.; Emerson, S.; Chen, L. The importance of the normality assumption in large public health data sets. Annu. Rev. Public Health 2002, 23, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw.e 2015, 67. [Google Scholar]
- Hurvich, C.M.; Tsai, C.-L. Bias of the corrected AIC criterion for underfitted regression and time series models. Biometrika 1991, 78, 499–509. [Google Scholar] [CrossRef]
- Carpenter, M.K. Missing Data in Randomised Controlled Trials: A Practical Guide; London School of Hygiene: London, UK, 2007; Available online: http://missingdata.lshtm.ac.uk/downloads/rm04_jh17_mk.pdf (accessed on 9 February 2021).
- Rothman, K.J. No adjustments are needed for multiple comparisons. Epidemiology (Camb. Mass.) 1990, 1, 43–46. [Google Scholar] [CrossRef] [Green Version]
- Mang, C.; Lagerquist, O.; Collins, D. Changes in corticospinal excitability evoked by common peroneal nerve stimulation depend on stimulation frequency. Exp. Brain Res. 2010, 203, 11–20. [Google Scholar] [CrossRef]
- Place, N.; Casartelli, N.; Glatthorn, J.F.; Maffiuletti, N.A. Comparison of quadriceps inactivation between nerve and muscle stimulation. Muscle Nerve 2010, 42, 894–900. [Google Scholar] [CrossRef]
- Gregory, C.M.; Bickel, C.S. Recruitment Patterns in Human Skeletal Muscle During Electrical Stimulation. Phys. Ther. 2005, 85, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Koski, L.; Lin, J.C.-H.; Wu, A.D.; Winstein, C.J. Reliability of intracortical and corticomotor excitability estimates obtained from the upper extremities in chronic stroke. Neurosci. Res. 2007, 58, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Wheaton, L.A.; Villagra, F.; Hanley, D.F.; Macko, R.F.; Forrester, L.W. Reliability of TMS motor evoked potentials in quadriceps of subjects with chronic hemiparesis after stroke. J. Neurol. Sci. 2009, 276, 115–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, C.I.; Pandyan, A.D. Electrical stimulation for preventing and treating post-stroke shoulder pain: A systematic Cochrane review. Clin. Rehabil. 2001, 15, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Ada, L.; Foongchomcheay, A. Efficacy of electrical stimulation in preventing or reducing subluxation of the shoulder after stroke: A meta-analysis. Aust. J. Physiother. 2002, 48, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Sujith, O.K. Functional electrical stimulation in neurological disorders. Eur. J. Neurol. 2008, 15, 437–444. [Google Scholar] [CrossRef]
- Stefan, K.; Kunesch, E.; Benecke, R.; Cohen, L.G.; Classen, J. Mechanisms of enhancement of human motor cortex excitability induced by interventional paired associative stimulation. J. Physiol. 2002, 543, 699–708. [Google Scholar] [CrossRef] [PubMed]
- de Vries, S.; Mulder, T. Motor imagery and stroke rehabilitation: A critical discussion. J. Rehabil. Med. 2007, 39, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto, O.; Valls-Sole, J.; Shanahan, P.; Rothwell, J. Reduction of intracortical inhibition in soleus muscle during postural activity. J. Neurophysiol. 2006, 96, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Ngomo, S.; Leonard, G.; Moffet, H.; Mercier, C. Comparison of transcranial magnetic stimulation measures obtained at rest and under active conditions and their reliability. J. Neurosci. Methods 2012, 205, 65–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, S.; Bathe-Peters, R.; Fleischmann, R.; Rönnefarth, M.; Scholz, M.; Brandt, S.A. Nonphysiological factors in navigated TMS studies; confounding covariates and valid intracortical estimates. Hum. Brain Mapp. 2015, 36, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Gugino, L.D.; Romero, J.R.; Aglio, L.; Titone, D.; Ramirez, M.; Pascual-Leone, A.; Grimson, E.; Weisenfeld, N.; Kikinis, R.; Shenton, M.E. Transcranial magnetic stimulation coregistered with MRI: A comparison of a guided versus blind stimulation technique and its effect on evoked compound muscle action potentials. Clin. Neurophysiol. 2001, 112, 1781–1792. [Google Scholar] [CrossRef] [Green Version]
- Schönfeldt-Lecuona, C.; Thielscher, A.; Freudenmann, R.W.; Kron, M.; Spitzer, M.; Herwig, U. Accuracy of stereotaxic positioning of transcranial magnetic stimulation. Brain Topogr. 2005, 17, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Julkunen, P.; Säisänen, L.; Danner, N.; Niskanen, E.; Hukkanen, T.; Mervaala, E.; Könönen, M. Comparison of navigated and non-navigated transcranial magnetic stimulation for motor cortex mapping, motor threshold and motor evoked potentials. Neuroimage 2009, 44, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Cuypers, K.; Thijs, H.; Meesen, R.L. Optimization of the transcranial magnetic stimulation protocol by defining a reliable estimate for corticospinal excitability. PLoS ONE 2014, 9, e86380. [Google Scholar] [CrossRef] [PubMed]
- Jochumsen, M.; Navid, M.S.; Nedergaard, R.W.; Signal, N.; Rashid, U.; Hassan, A.; Haavik, H.; Taylor, D.; Niazi, I.K. Self-Paced Online vs. Cue-Based Offline Brain–Computer Interfaces for Inducing Neural Plasticity. Brain Sci. 2019, 9, 127. [Google Scholar] [CrossRef] [Green Version]
- Jayaram, G.; Santos, L.; Stinear, J.W. Spike-timing-dependent plasticity induced in resting lower limb cortex persists during subsequent walking. Brain Res. 2007, 1153, 92–97. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MEP Amplitude (µV) | MEP Area (µV/ms) | |
|---|---|---|
| Mean | 1730 | 7.2 |
| Within SD | 1060 | 4.6 |
| Between SD | 839 | 3.8 |
| MEP Amplitude (µV) | MEP Area (µV/ms) | ||
|---|---|---|---|
| Numerator df | p Value | p Value | |
| Baseline covariates | |||
| AMTh | 1 | 0.0001 * | 0.0001 * |
| MVC | 1 | 0.06 | 0.04 * |
| Main effects and interactions | |||
| Stimulation intensity | 16 | <0.00005 * | <0.00005 * |
| Movement type | 12 | 0.002 * | 0.001 * |
| Time | 18 | 0.009 * | 0.0004 * |
| Stimulation intensity × Movement type | 8 | 0.0005 * | 0.0001 * |
| Stimulation intensity × Time | 12 | 0.002 * | <0.00005 * |
| Movement type × Time | 9 | 0.001 * | 0.002 * |
| Stimulation intensity × Movement type × Time | 6 | 0.0004 * | 0.0003 * |
| Hi-Voluntary intervention | |||
| Super-/sub-additivity of suprathreshold stimulation overall, voluntary vs. imagined | 8 | 0.0005 * | 0.0001 * |
| Super-/sub-additivity of suprathreshold stimulation vs. no stimulation and voluntary vs. imagined | 4 | 0.0001 * | <0.00005 * |
| Super-/sub-additivity of suprathreshold stimulation vs. threshold and voluntary vs. imagined | 4 | 0.139 | 0.108 |
| Lo-Voluntary intervention | |||
| Super-/sub-additivity of threshold stimulation vs. no stimulation and voluntary vs. imagined | 4 | 0.0013 * | 0.0004 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alder, G.; Signal, N.; Vandal, A.C.; Olsen, S.; Jochumsen, M.; Niazi, I.K.; Taylor, D. Investigating the Intervention Parameters of Endogenous Paired Associative Stimulation (ePAS). Brain Sci. 2021, 11, 224. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020224
Alder G, Signal N, Vandal AC, Olsen S, Jochumsen M, Niazi IK, Taylor D. Investigating the Intervention Parameters of Endogenous Paired Associative Stimulation (ePAS). Brain Sciences. 2021; 11(2):224. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020224
Chicago/Turabian StyleAlder, Gemma, Nada Signal, Alain C. Vandal, Sharon Olsen, Mads Jochumsen, Imran Khan Niazi, and Denise Taylor. 2021. "Investigating the Intervention Parameters of Endogenous Paired Associative Stimulation (ePAS)" Brain Sciences 11, no. 2: 224. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11020224