
New Insights into In Vivo Dopamine Physiology and Neurostimulation: A Fiber Photometry Study Highlighting the Impact of Medial Forebrain Bundle Deep Brain Stimulation on the Nucleus Accumbens



 , and
, and
Abstract
:
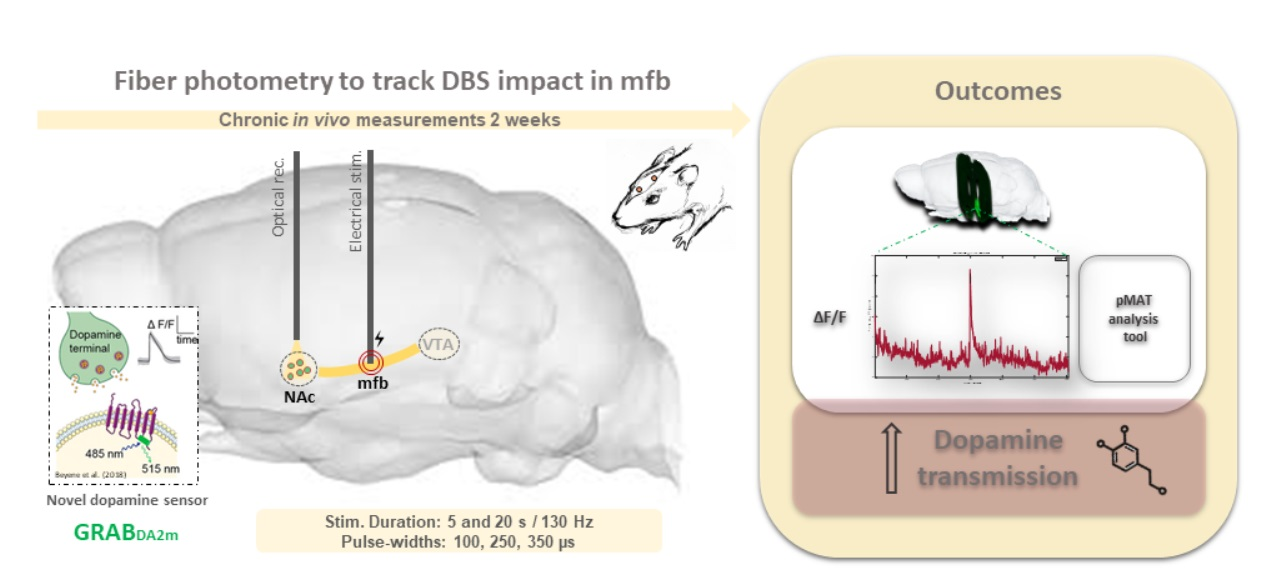{kind=link}
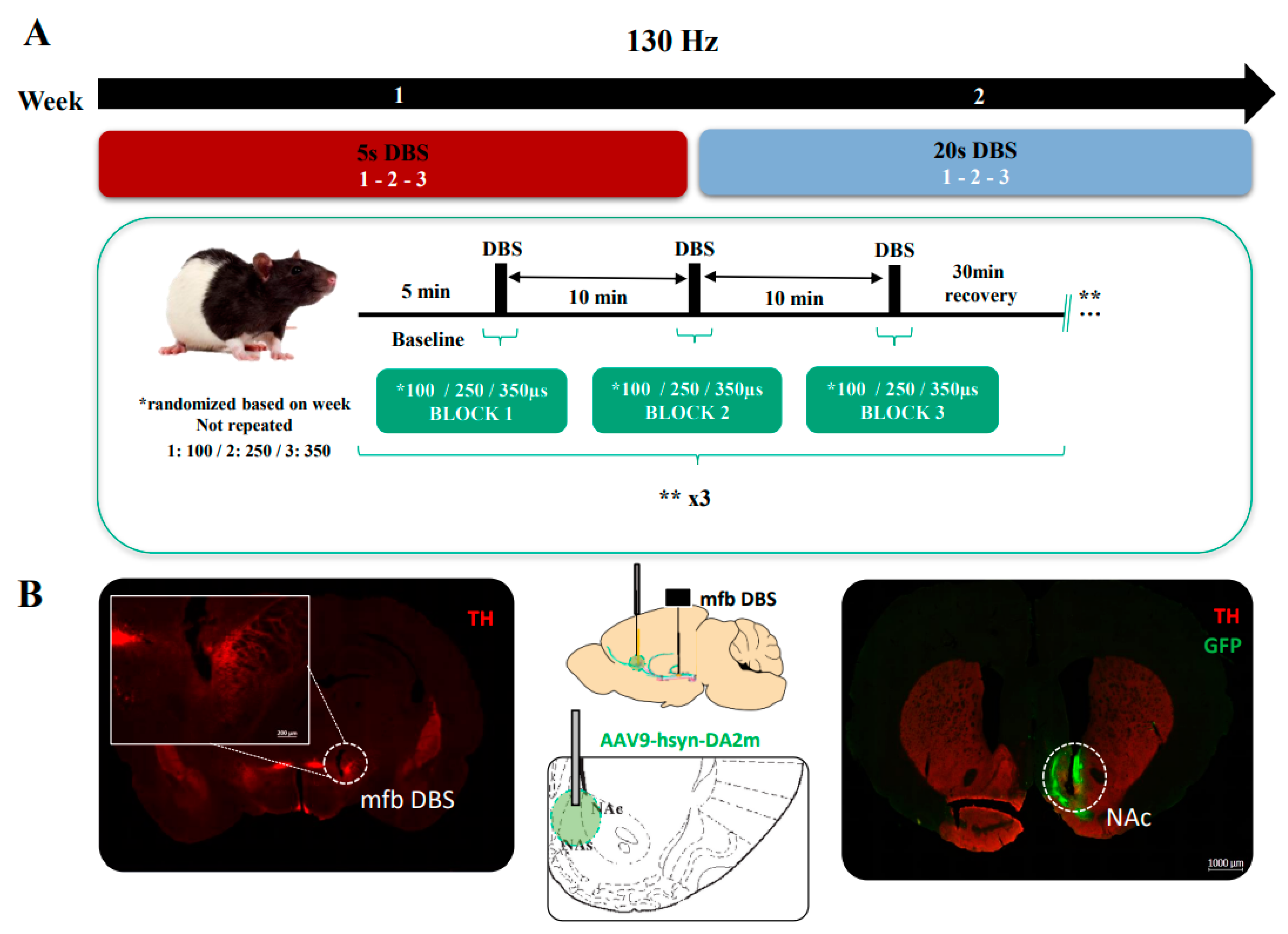{kind=link}
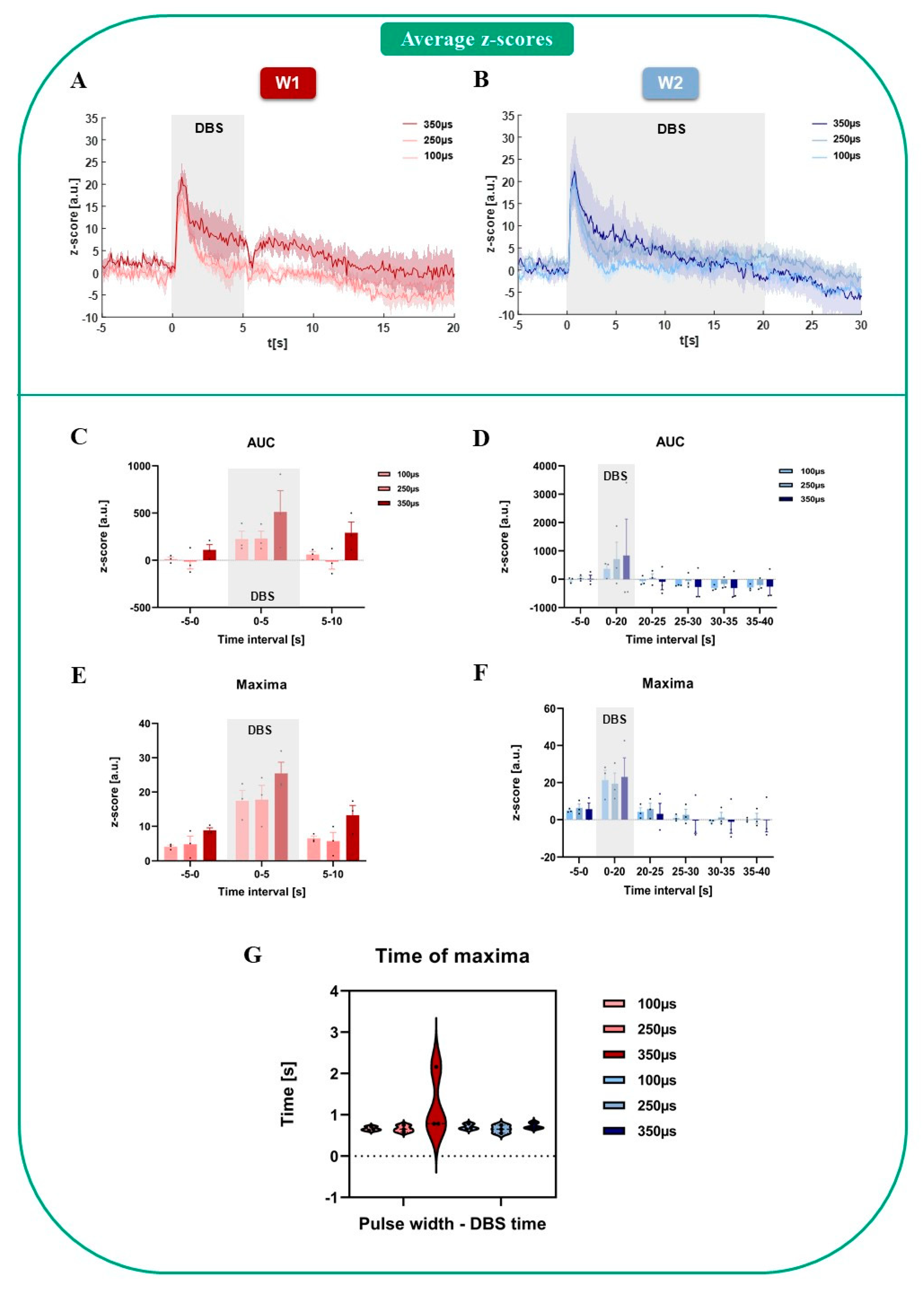{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals—Experimental Design
2.2. Surgery (Viral Construct, Optic Fiber, DBS Electrode)
2.3. Fiber Photometry (FP) Recordings—Data Analysis
2.4. Immunohistochemistry
2.5. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Coenen, V.A.; Bewernick, B.H.; Kayser, S.; Kilian, H.; Boström, J.; Greschus, S.; Hurlemann, R.; Klein, M.E.; Spanier, S.; Sajonz, B.; et al. Superolateral Medial Forebrain Bundle Deep Brain Stimulation in Major Depression: A Gateway Trial. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2019, 44, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Fenoy, A.J.; Schulz, P.; Selvaraj, S.; Burrows, C.; Spiker, D.; Cao, B.; Zunta-Soares, G.; Gajwani, P.; Quevedo, J.; Soares, J. Deep Brain Stimulation of the Medial Forebrain Bundle: Distinctive Responses in Resistant Depression. J. Affect. Disord. 2016, 203, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Fenoy, A.J.; Schulz, P.E.; Selvaraj, S.; Burrows, C.L.; Zunta-Soares, G.; Durkin, K.; Zanotti-Fregonara, P.; Quevedo, J.; Soares, J.C. A Longitudinal Study on Deep Brain Stimulation of the Medial Forebrain Bundle for Treatment-Resistant Depression. Transl. Psychiatry 2018, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Kilian, H.M.; Meyer, D.M.; Bewernick, B.H.; Spanier, S.; Coenen, V.A.; Schlaepfer, T.E. Discontinuation of Superolateral Medial Forebrain Bundle Deep Brain Stimulation for Treatment-Resistant Depression Leads to Critical Relapse. Biol. Psychiatry 2019, 85, e23–e24. [Google Scholar] [CrossRef]
- Oldani, L.; Benatti, B.; Macellaro, M.; Porta, M.; Servello, D.; Zekaj, E.; Dell’Osso, B. A Case of Treatment-Resistant Bipolar Depression and Comorbid OCD Treated with Deep Brain Stimulation of the Medial Forebrain Bundle: 5 Years Follow-up Results. J. Clin. Neurosci. Off. J. Neurosurg. Soc. Australas. 2021, 89, 103–105. [Google Scholar] [CrossRef]
- Schlaepfer, T.E.; Bewernick, B.H.; Kayser, S.; Mädler, B.; Coenen, V.A. Rapid Effects of Deep Brain Stimulation for Treatment-Resistant Major Depression. Biol. Psychiatry 2013, 73, 1204–1212. [Google Scholar] [CrossRef]
- Schlaepfer, T.E.; Bewernick, B.H.; Kayser, S.; Hurlemann, R.; Coenen, V.A. Deep Brain Stimulation of the Human Reward System for Major Depression--Rationale, Outcomes and Outlook. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2014, 39, 1303–1314. [Google Scholar] [CrossRef]
- Bregman, T.; Reznikov, R.; Diwan, M.; Raymond, R.; Butson, C.R.; Nobrega, J.N.; Hamani, C. Antidepressant-like Effects of Medial Forebrain Bundle Deep Brain Stimulation in Rats Are Not Associated With Accumbens Dopamine Release. Brain Stimulat. 2015, 8, 708–713. [Google Scholar] [CrossRef]
- Bühning, F.; Miguel Telega, L.; Tong, Y.; Pereira, J.; Coenen, V.A.; Döbrössy, M.D. Electrophysiological and Molecular Effects of Bilateral Deep Brain Stimulation of the Medial Forebrain Bundle in a Rodent Model of Depression. Exp. Neurol. 2022, 355, 114122. [Google Scholar] [CrossRef]
- Dandekar, M.P.; Saxena, A.; Scaini, G.; Shin, J.H.; Migut, A.; Giridharan, V.V.; Zhou, Y.; Barichello, T.; Soares, J.C.; Quevedo, J.; et al. Medial Forebrain Bundle Deep Brain Stimulation Reverses Anhedonic-Like Behavior in a Chronic Model of Depression: Importance of BDNF and Inflammatory Cytokines. Mol. Neurobiol. 2019, 56, 4364–4380. [Google Scholar] [CrossRef]
- Döbrössy, M.D.; Furlanetti, L.L.; Coenen, V.A. Electrical Stimulation of the Medial Forebrain Bundle in Pre-Clinical Studies of Psychiatric Disorders. Neurosci. Biobehav. Rev. 2015, 49, 32–42. [Google Scholar] [CrossRef]
- Edemann-Callesen, H.; Voget, M.; Empl, L.; Vogel, M.; Wieske, F.; Rummel, J.; Heinz, A.; Mathé, A.A.; Hadar, R.; Winter, C. Medial Forebrain Bundle Deep Brain Stimulation Has Symptom-Specific Anti-Depressant Effects in Rats and as Opposed to Ventromedial Prefrontal Cortex Stimulation Interacts With the Reward System. Brain Stimulat. 2015, 8, 714–723. [Google Scholar] [CrossRef]
- Thiele, S.; Furlanetti, L.; Pfeiffer, L.-M.; Coenen, V.A.; Döbrössy, M.D. The Effects of Bilateral, Continuous, and Chronic Deep Brain Stimulation of the Medial Forebrain Bundle in a Rodent Model of Depression. Exp. Neurol. 2018, 303, 153–161. [Google Scholar] [CrossRef]
- Thiele, S.; Sörensen, A.; Weis, J.; Braun, F.; Meyer, P.T.; Coenen, V.A.; Döbrössy, M.D. Deep Brain Stimulation of the Medial Forebrain Bundle in a Rodent Model of Depression: Exploring Dopaminergic Mechanisms with Raclopride and Micro-PET. Stereotact. Funct. Neurosurg. 2020, 98, 8–20. [Google Scholar] [CrossRef]
- Tong, Y.; Pfeiffer, L.; Serchov, T.; Coenen, V.A.; Döbrössy, M.D. Optogenetic Stimulation of Ventral Tegmental Area Dopaminergic Neurons in a Female Rodent Model of Depression: The Effect of Different Stimulation Patterns. J. Neurosci. Res. 2022, 100, 897–911. [Google Scholar] [CrossRef]
- Pallikaras, V.; Shizgal, P. The Convergence Model of Brain Reward Circuitry: Implications for Relief of Treatment-Resistant Depression by Deep-Brain Stimulation of the Medial Forebrain Bundle. Front. Behav. Neurosci. 2022, 16, 851067. [Google Scholar] [CrossRef]
- Mohebi, A.; Pettibone, J.R.; Hamid, A.A.; Wong, J.-M.T.; Vinson, L.T.; Patriarchi, T.; Tian, L.; Kennedy, R.T.; Berke, J.D. Dissociable Dopamine Dynamics for Learning and Motivation. Nature 2019, 570, 65–70. [Google Scholar] [CrossRef]
- Saddoris, M.P.; Sugam, J.A.; Cacciapaglia, F.; Carelli, R.M. Rapid Dopamine Dynamics in the Accumbens Core and Shell: Learning and Action. Front. Biosci. Elite Ed. 2013, 5, 273–288. [Google Scholar] [CrossRef]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorders. Int. J. Neuropsychopharmacol. 2017, 20, 1036–1046. [Google Scholar] [CrossRef]
- Döbrössy, M.D.; Ramanathan, C.; Ashouri Vajari, D.; Tong, Y.; Schlaepfer, T.; Coenen, V.A. Neuromodulation in Psychiatric Disorders: Experimental and Clinical Evidence for Reward and Motivation Network Deep Brain Stimulation: Focus on the Medial Forebrain Bundle. Eur. J. Neurosci. 2021, 53, 89–113. [Google Scholar] [CrossRef]
- Ramasubbu, R.; Lang, S.; Kiss, Z.H.T. Dosing of Electrical Parameters in Deep Brain Stimulation (DBS) for Intractable Depression: A Review of Clinical Studies. Front. Psychiatry 2018, 9, 302. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, J.S. Two Substrates for Medial Forebrain Bundle Self-Stimulation: Myelinated Axons and Dopamine Axons. Neurosci. Biobehav. Rev. 1989, 13, 91–98. [Google Scholar] [CrossRef]
- Torres-Sanchez, S.; Perez-Caballero, L.; Berrocoso, E. Cellular and Molecular Mechanisms Triggered by Deep Brain Stimulation in Depression: A Preclinical and Clinical Approach. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bielajew, C.; Jurgens, S.; Fouriezos, G. The Effect of Pulse Duration of Refractory Periods of Neurons Mediating Brain-Stimulation Reward. Behav. Brain Res. 1987, 24, 233–241. [Google Scholar] [CrossRef]
- Shizgal, P.; Conover, K.; Schindler, D. Medial Forebrain Bundle Units in the Rat: Dependence of Refractory Period Estimates on Pulse Duration. Behav. Brain Res. 1991, 42, 151–160. [Google Scholar] [CrossRef]
- Klanker, M.; Feenstra, M.; Willuhn, I.; Denys, D. Deep Brain Stimulation of the Medial Forebrain Bundle Elevates Striatal Dopamine Concentration without Affecting Spontaneous or Reward-Induced Phasic Release. Neuroscience 2017, 364, 82–92. [Google Scholar] [CrossRef]
- Yavich, L.; Tanila, H. Mechanics of Self-Stimulation and Dopamine Release in the Nucleus Accumbens. Neuroreport 2007, 18, 1271–1274. [Google Scholar] [CrossRef]
- Ashouri Vajari, D.; Ramanathan, C.; Tong, Y.; Stieglitz, T.; Coenen, V.A.; Döbrössy, M.D. Medial Forebrain Bundle DBS Differentially Modulates Dopamine Release in the Nucleus Accumbens in a Rodent Model of Depression. Exp. Neurol. 2020, 327, 113224. [Google Scholar] [CrossRef]
- Sabatini, B.L.; Tian, L. Imaging Neurotransmitter and Neuromodulator Dynamics In Vivo with Genetically Encoded Indicators. Neuron 2020, 108, 17–32. [Google Scholar] [CrossRef]
- Wang, Y.; DeMarco, E.M.; Witzel, L.S.; Keighron, J.D. A Selected Review of Recent Advances in the Study of Neuronal Circuits Using Fiber Photometry. Pharmacol. Biochem. Behav. 2021, 201, 173113. [Google Scholar] [CrossRef]
- Wu, Z.; Lin, D.; Li, Y. Pushing the Frontiers: Tools for Monitoring Neurotransmitters and Neuromodulators. Nat. Rev. Neurosci. 2022, 23, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhou, J.; Dai, B.; Qian, T.; Zeng, J.; Li, X.; Zhuo, Y.; Zhang, Y.; Wang, Y.; Qian, C.; et al. Next-Generation GRAB Sensors for Monitoring Dopaminergic Activity in Vivo. Nat. Methods 2020, 17, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Beck, C.; Gong, Y. Genetically Encoded Fluorescent Indicators for Imaging Brain Chemistry. Biosensors 2021, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Bruno, C.A.; O’Brien, C.; Bryant, S.; Mejaes, J.I.; Estrin, D.J.; Pizzano, C.; Barker, D.J. PMAT: An Open-Source Software Suite for the Analysis of Fiber Photometry Data. Pharmacol. Biochem. Behav. 2021, 201, 173093. [Google Scholar] [CrossRef] [PubMed]
- Ikemoto, S.; Panksepp, J. The Role of Nucleus Accumbens Dopamine in Motivated Behavior: A Unifying Interpretation with Special Reference to Reward-Seeking. Brain Res. Brain Res. Rev. 1999, 31, 6–41. [Google Scholar] [CrossRef]
- Alcaro, A.; Panksepp, J. The SEEKING Mind: Primal Neuro-Affective Substrates for Appetitive Incentive States and Their Pathological Dynamics in Addictions and Depression. Neurosci. Biobehav. Rev. 2011, 35, 1805–1820. [Google Scholar] [CrossRef]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: New York, NY, USA, 1998; ISBN 0-19-509673-8. [Google Scholar]
- Wise, R.A.; McDevitt, R.A. Drive and Reinforcement Circuitry in the Brain: Origins, Neurotransmitters, and Projection Fields. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2018, 43, 680–689. [Google Scholar] [CrossRef]
- Howe, M.W.; Tierney, P.L.; Sandberg, S.G.; Phillips, P.E.M.; Graybiel, A.M. Prolonged Dopamine Signalling in Striatum Signals Proximity and Value of Distant Rewards. Nature 2013, 500, 575–579. [Google Scholar] [CrossRef]
- Berridge, K.C.; Kringelbach, M.L. Pleasure Systems in the Brain. Neuron 2015, 86, 646–664. [Google Scholar] [CrossRef]
- Berridge, K.C. Affective Valence in the Brain: Modules or Modes? Nat. Rev. Neurosci. 2019, 20, 225–234. [Google Scholar] [CrossRef]
- Wu, H.; Kakusa, B.; Neuner, S.; Christoffel, D.J.; Heifets, B.D.; Malenka, R.C.; Halpern, C.H. Local Accumbens in Vivo Imaging during Deep Brain Stimulation Reveals a Strategy-Dependent Amelioration of Hedonic Feeding. Proc. Natl. Acad. Sci. USA 2022, 119, e2109269118. [Google Scholar] [CrossRef] [PubMed]
- Patriarchi, T.; Mohebi, A.; Sun, J.; Marley, A.; Liang, R.; Dong, C.; Puhger, K.; Mizuno, G.O.; Davis, C.M.; Wiltgen, B.; et al. An Expanded Palette of Dopamine Sensors for Multiplex Imaging in Vivo. Nat. Methods 2020, 17, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, M.P.; Luse, D.; Hoffmann, C.; Cotton, P.; Peery, T.; Ruiz, C.; Hussey, C.; Giridharan, V.V.; Soares, J.C.; Quevedo, J.; et al. Increased Dopamine Receptor Expression and Anti-Depressant Response Following Deep Brain Stimulation of the Medial Forebrain Bundle. J. Affect. Disord. 2017, 217, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Gallo, E.F.; Meszaros, J.; Sherman, J.D.; Chohan, M.O.; Teboul, E.; Choi, C.S.; Moore, H.; Javitch, J.A.; Kellendonk, C. Accumbens Dopamine D2 Receptors Increase Motivation by Decreasing Inhibitory Transmission to the Ventral Pallidum. Nat. Commun. 2018, 9, 1086. [Google Scholar] [CrossRef]
- Cossette, M.-P.; Conover, K.; Shizgal, P. The Neural Substrates for the Rewarding and Dopamine-Releasing Effects of Medial Forebrain Bundle Stimulation Have Partially Discrepant Frequency Responses. Behav. Brain Res. 2016, 297, 345–358. [Google Scholar] [CrossRef]
- Nakahara, D.; Fuchikami, K.; Ozaki, N.; Iwasaki, T.; Nagatsu, T. Differential Effect of Self-Stimulation on Dopamine Release and Metabolism in the Rat Medial Frontal Cortex, Nucleus Accumbens and Striatum Studied by in Vivo Microdialysis. Brain Res. 1992, 574, 164–170. [Google Scholar] [CrossRef]
- de Jong, J.W.; Fraser, K.M.; Lammel, S. Mesoaccumbal Dopamine Heterogeneity: What Do Dopamine Firing and Release Have to Do with It? Annu. Rev. Neurosci. 2022, 45, 109–129. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miguel Telega, L.; Ashouri Vajari, D.; Stieglitz, T.; Coenen, V.A.; Döbrössy, M.D. New Insights into In Vivo Dopamine Physiology and Neurostimulation: A Fiber Photometry Study Highlighting the Impact of Medial Forebrain Bundle Deep Brain Stimulation on the Nucleus Accumbens. Brain Sci. 2022, 12, 1105. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12081105
Miguel Telega L, Ashouri Vajari D, Stieglitz T, Coenen VA, Döbrössy MD. New Insights into In Vivo Dopamine Physiology and Neurostimulation: A Fiber Photometry Study Highlighting the Impact of Medial Forebrain Bundle Deep Brain Stimulation on the Nucleus Accumbens. Brain Sciences. 2022; 12(8):1105. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12081105
Chicago/Turabian StyleMiguel Telega, Lidia, Danesh Ashouri Vajari, Thomas Stieglitz, Volker A. Coenen, and Máté D. Döbrössy. 2022. "New Insights into In Vivo Dopamine Physiology and Neurostimulation: A Fiber Photometry Study Highlighting the Impact of Medial Forebrain Bundle Deep Brain Stimulation on the Nucleus Accumbens" Brain Sciences 12, no. 8: 1105. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12081105