Effects of GLP-1 Receptor Activation on a Pentylenetetrazole—Kindling Rat Model

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Study Design
2.3. PTZ-Induced Kindled Rat Model and Behavioral Assessment
2.4. Harvesting of Brain Specimens
2.5. Measurement of Oxidative Stress Markers (MDA, GSH and Catalase Activity) in Brain Tissues
2.6. Measurement of β-Catenin and Caspase-3 Protein Expression by Western Blotting
2.7. Histopathological Examination for Hippocampal Neurons in CA3 Region
2.8. Measurement of Hsp70 and LC3 Expression by Immunohistochemistry
2.9. Statistical Analysis
3. Results
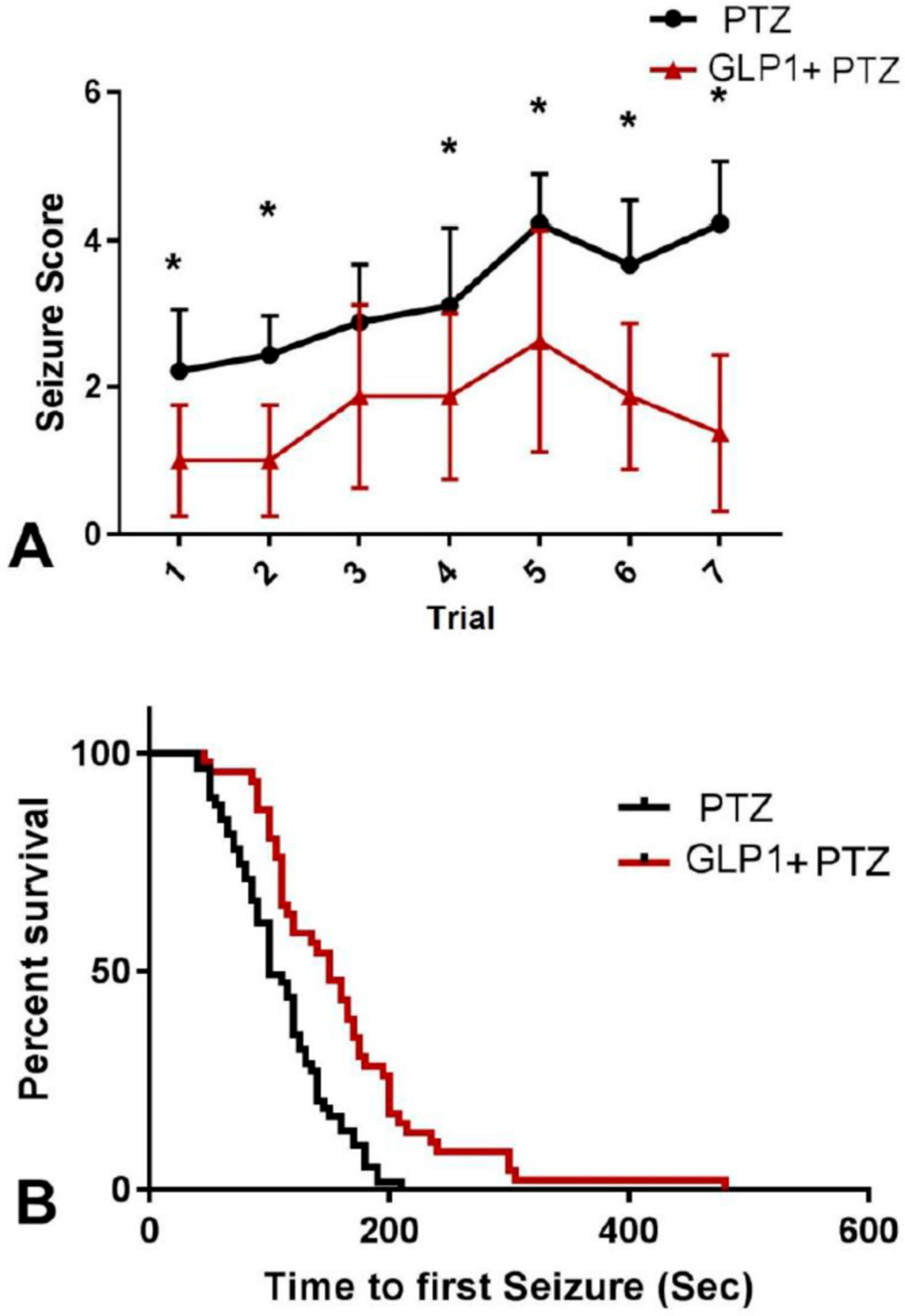
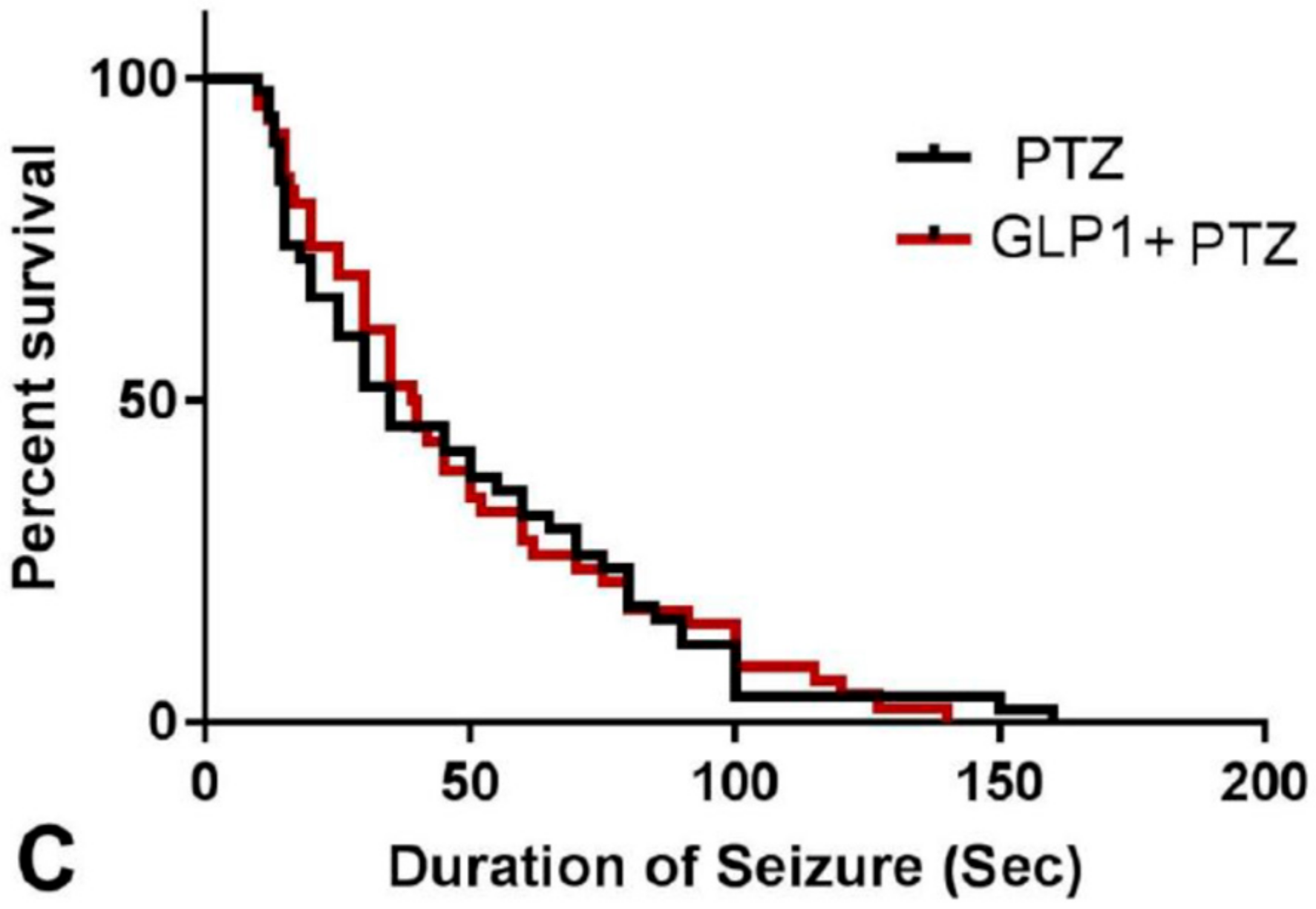
3.1. Animal Survival
3.2. Neurobehavioral Changes
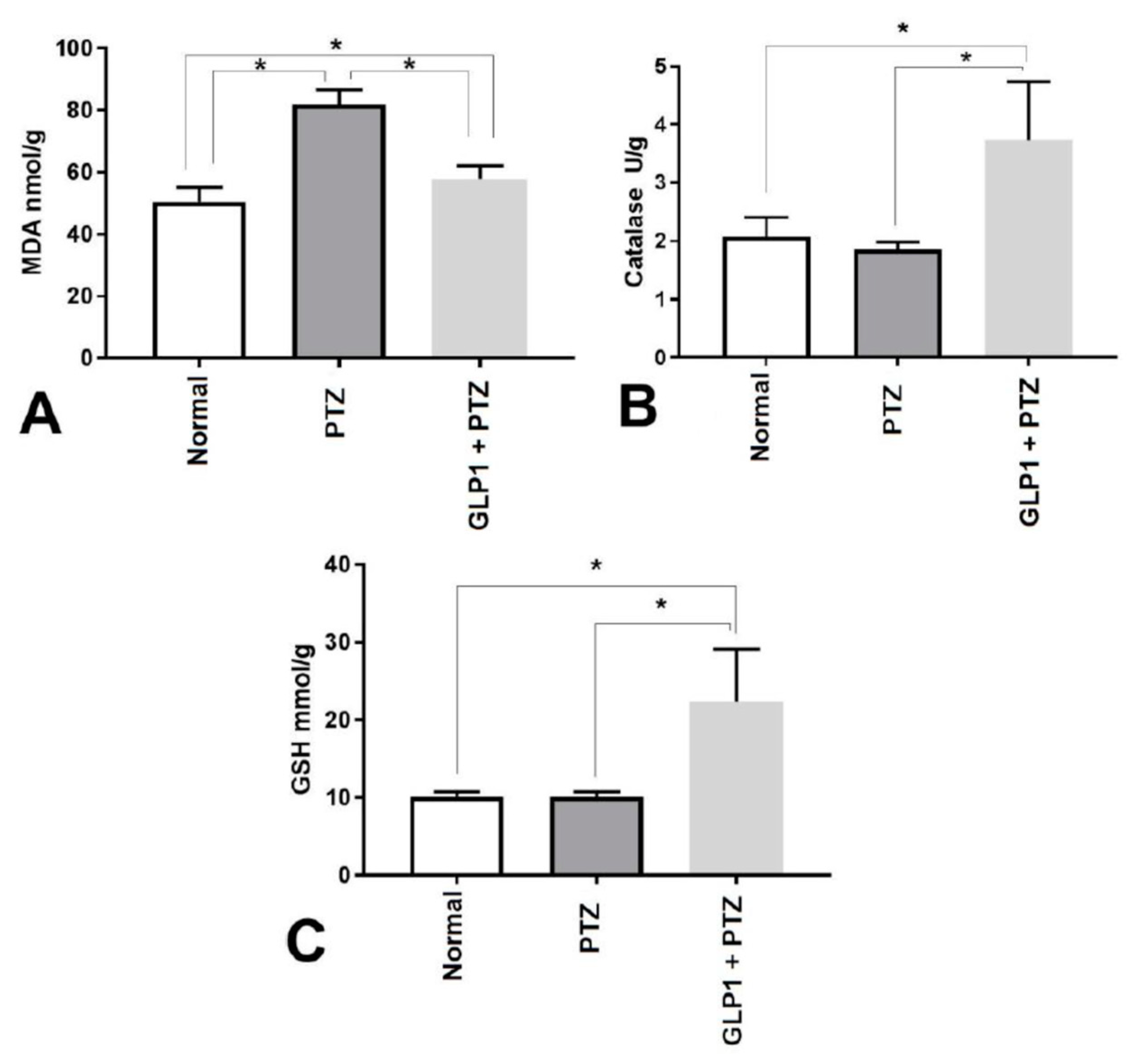
3.3. Markers of oxidative stress (MDA) and antioxidants (CAT and GSH)
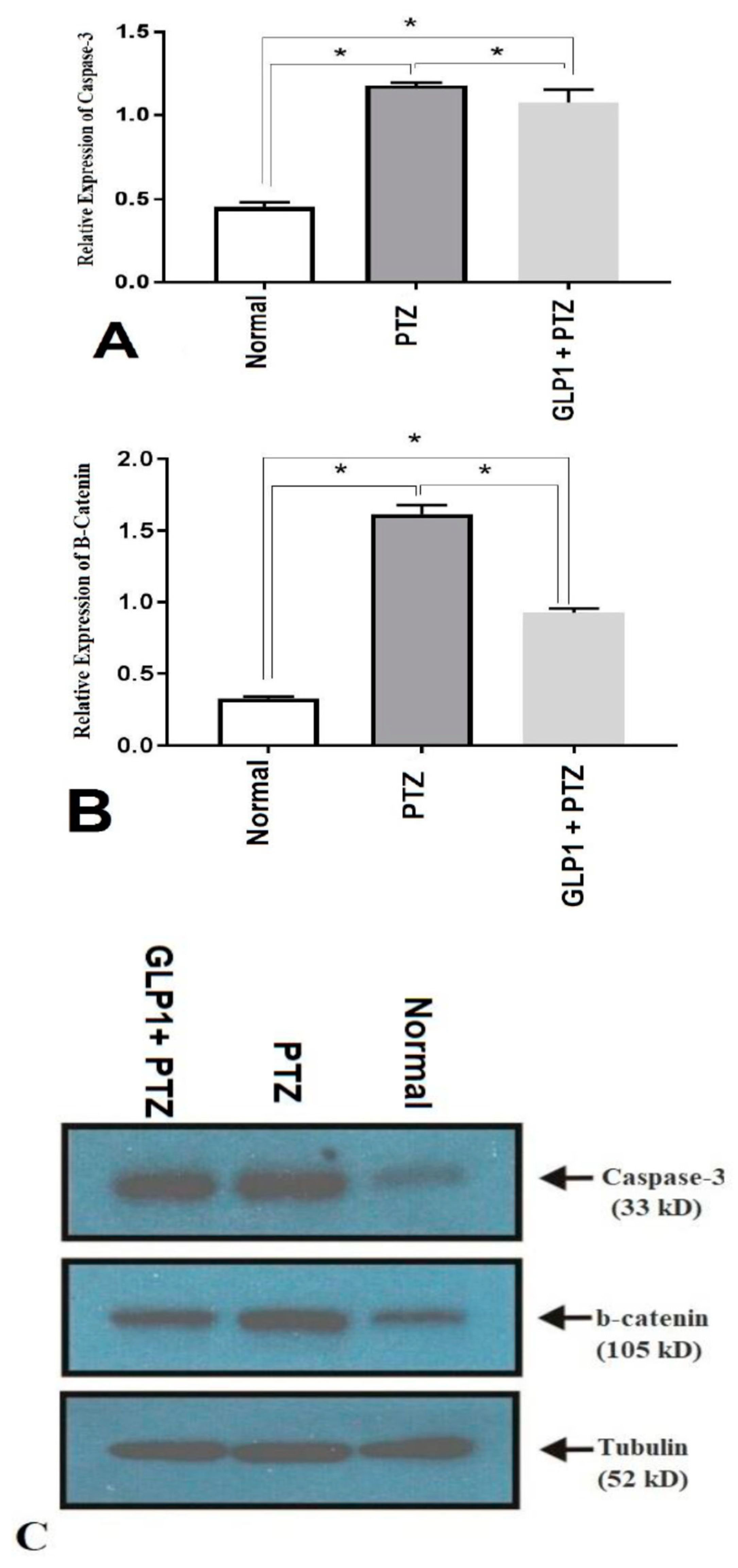
3.4. Expression of Caspase-3 and β-Catenin Proteins by Western Blotting
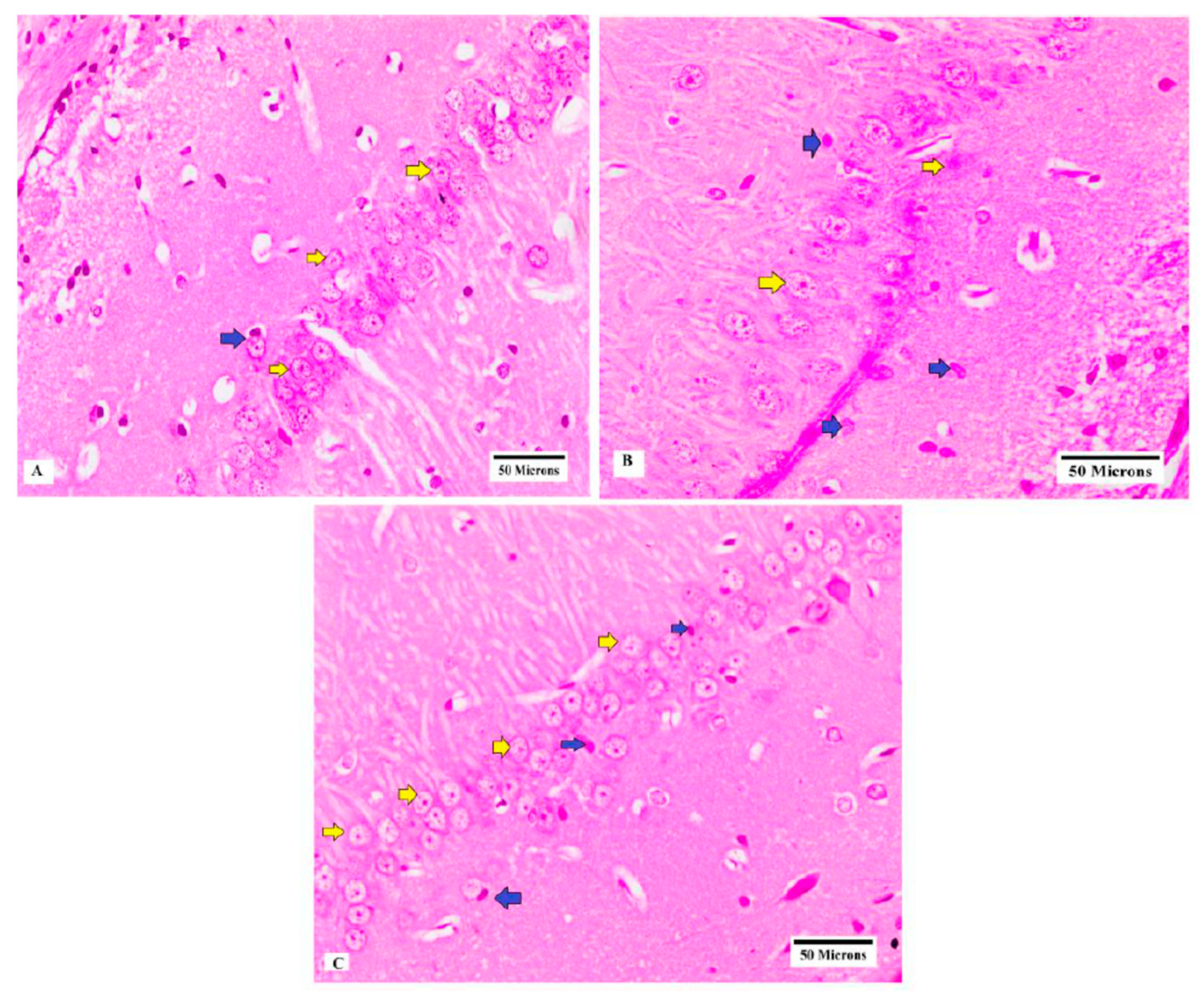
3.5. Histopathological Examination of CA3 Region of Hippocampus
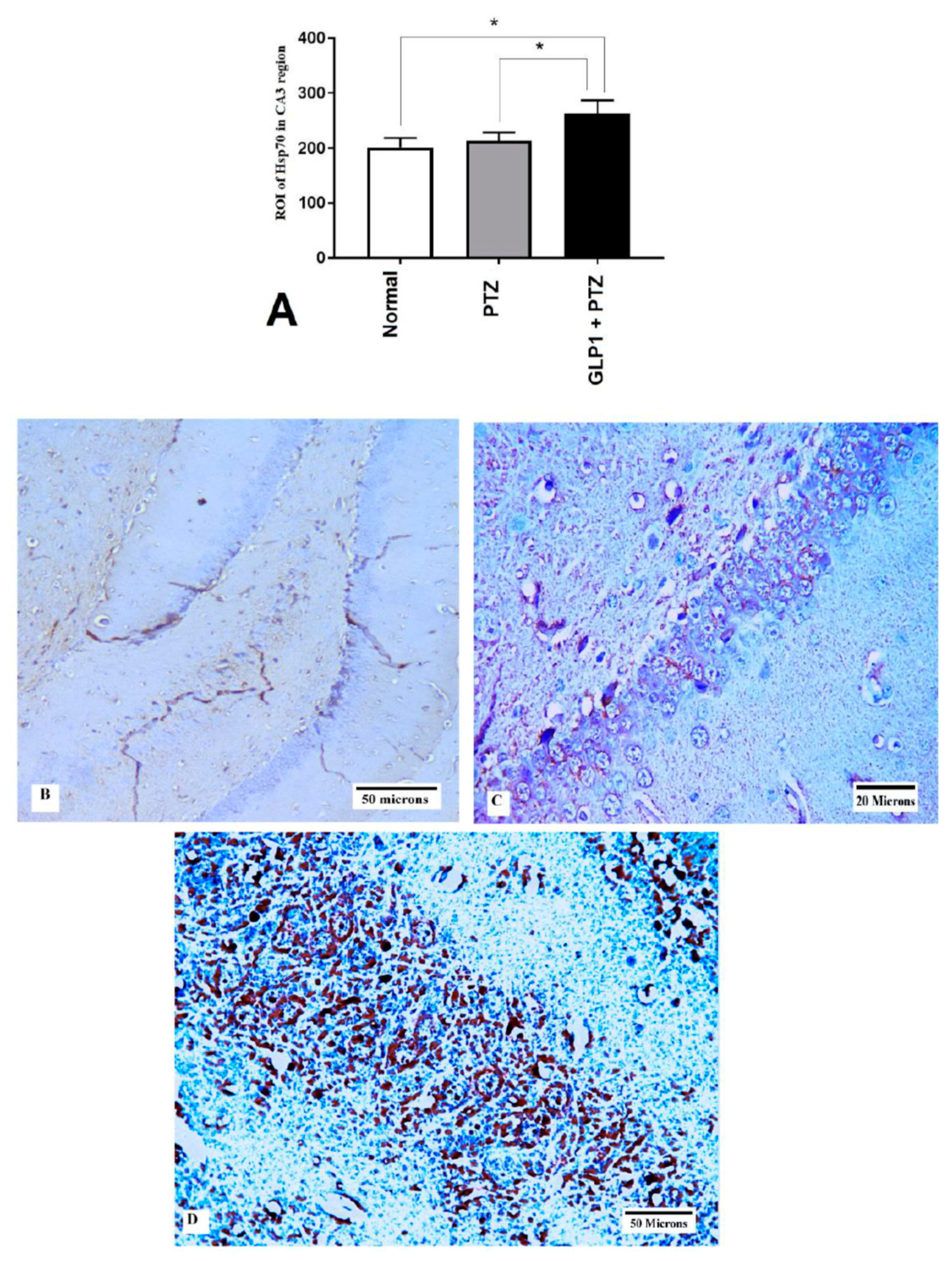
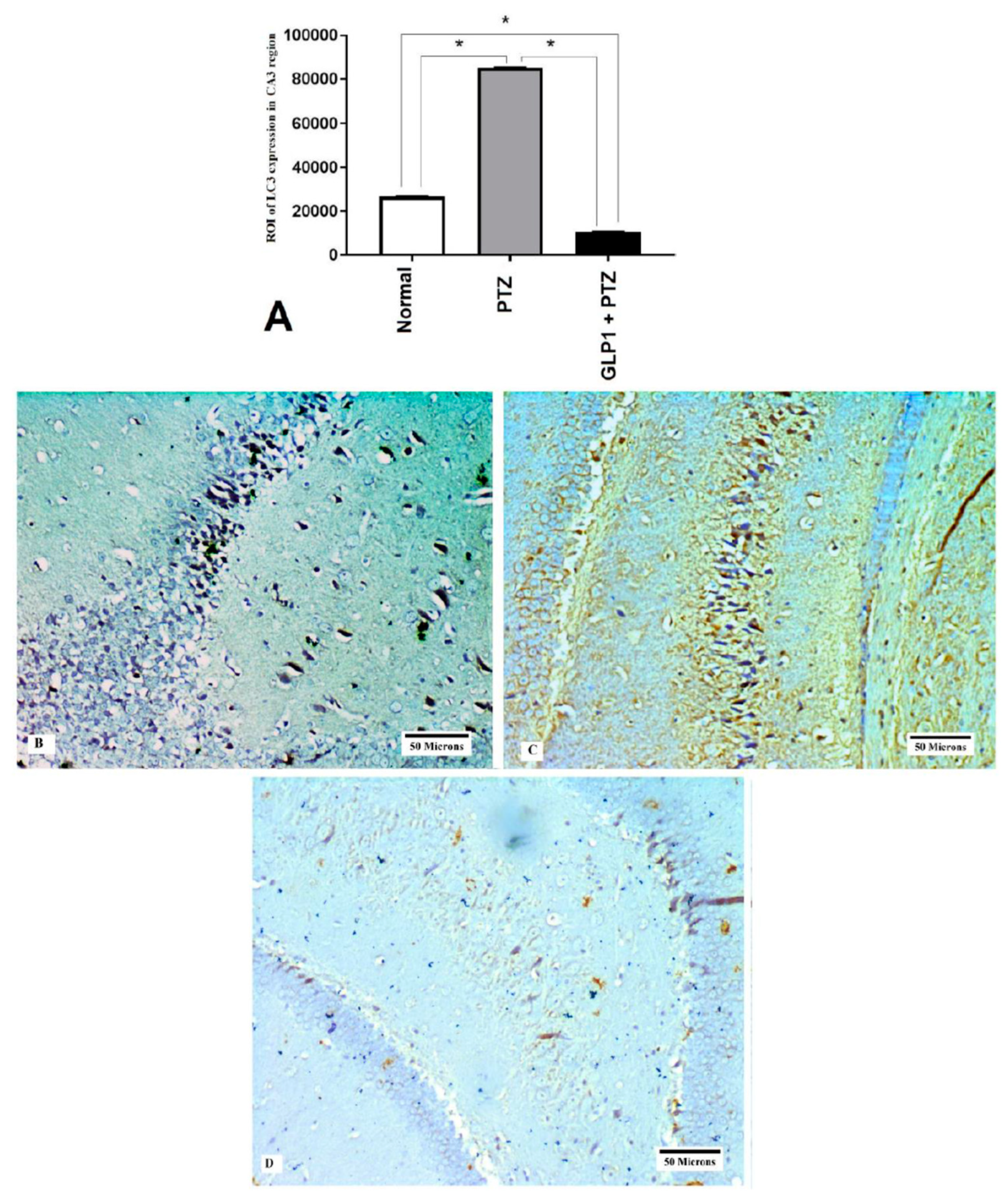
3.6. Expression of Hsp70 and LC3 by Immunohistochemistry
3.7. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Avanzini, G.; Prilipko, L. Epilepsy Care in the World WHO; Epilepsy Atlas, WHO Press, World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Shakirullah, S.; Ali, N.; Khan, A.M.; Nabi, M. The prevalence, incidence and etiology of epilepsy. Int. J. Clin. Exp. Neurol. 2014, 2, 29–39. [Google Scholar]
- El Tallawy, H.N.; Farghaly, W.M.; Metwaly, N.A.; Rageh, T.A.; Shehata, G.A.; Elfetoh, N.A.; Hegazy, A.M.; El-Moselhy, E.A.; Rayan, I.; Al-Fawal, B.M.; et al. Door-to-door survey of major neurological disorders in Al Kharga District, New Valley, Egypt: Methodological aspects. Neuroepidemiology 2010, 35, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gan, J.; Qu, Y.; Li, J.; Zhao, F.; Mu, D. An evaluation of the links between microRNA, autophagy, and epilepsy. Rev. Neurosci. 2015, 26, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Erkec, O.E. Pentylenetetrazol Kindling Epilepsy Model. J. Turk. Epilepsi Soc. 2015, 21, 6–12. [Google Scholar] [CrossRef]
- Meier, C.; Kraenzlin, M.E. Antiepileptics and bone health. Ther. Adv. Musculoskelet. Dis. 2011, 3, 2435–2443. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Li, X.-H.; Zeng, X.-C.; Li, J.; Zhou, J.; Xiao, B.; Hu, K. Curcumin protects neuronal cells against status-epilepticus-induced hippocampal damage through induction of autophagy and inhibition of necroptosis. Can. J. Physiol. Pharmacol. 2017, 95, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Sudha, K.; Rao, A.V.; Rao, A. Oxidative stress and antioxidants in epilepsy. Clin. Chim. Acta 2001, 303, 19–24. [Google Scholar] [CrossRef]
- Gupta, Y.; Kumar, M.V.; Srivastava, A. Effect of Centella asiatica on pentylenetetrazole-induced kindling, cognition and oxidative stress in rats. Pharmacol. Biochem. 2003, 74, 579–585. [Google Scholar] [CrossRef]
- Simonian, N.; Getz, R.; Leveque, J.-C.; Konradi, C.; Coyle, J. Kainic acid induces apoptosis in neurons. Neuroscience 1996, 75, 1047–1055. [Google Scholar] [CrossRef]
- Naseer, M.I.; Ullah, I.; Ullah, N.; Lee, H.Y.; Cheon, E.W.; Chung, J.; Kim, M.O. Neuroprotective effect of vitamin C against PTZ induced apoptotic neurodegeneration in adult rat brain. J. Pharm. Sci. 2011, 24. [Google Scholar]
- Mylvaganam, S.; Ramani, M.; Krawczyk, M.; Carlen, P.L. Roles of gap junctions, connexins, and pannexins in epilepsy. Front. Physiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dericioglu, N.; Söylemezoglu, F.; Gursoy-Ozdemir, Y.; Akalan, N.; Saygi, S.; Dalkara, T. Cell death and survival mechanisms are concomitantly active in the hippocampus of patients with mesial temporal sclerosis. Neuroscience 2013, 237, 56–65. [Google Scholar] [CrossRef]
- Hussein, A.M.; Sadek, K.M.; Abuoleseod, O. Effects of Ferulic Acid on Oxidative Stress, Heat Shock Protein 70, Connexin 43 and Monoamines in Hippocampus of Pentylenetetrazole-Kindled Rats. J. Physiol. Pharmacol. 2017, 97, 579–585. [Google Scholar] [CrossRef]
- Galluzzi, L.; Pietrocola, F.; Levine, B.; Kroemer, G. Metabolic control of autophagy. Cell 2014, 159, 1263–1276. [Google Scholar] [CrossRef]
- Harris, H.; Rubinsztein, D.C. Control of autophagy as a therapy for neurodegenerative disease. Nat. Rev. Neurol. 2012, 8, 108–117. [Google Scholar] [CrossRef]
- Wong, M. Mammalian target of rapamycin (mTOR) inhibition as potential antiepileptogenic therapy: From tuberous sclerosis to common acquired epilepsies. Epilepsia 2010, 51, 27–36. [Google Scholar] [CrossRef]
- Wong, M. Cleaning Up Epilepsy and Neurodegeneration: The Role of Autophagy in Epileptogenesis. Epilepsy Curr. 2013, 13, 177–178. [Google Scholar] [CrossRef] [Green Version]
- Knauf, C.; Cani, P.D.; Kim, D.H.; Iglesias, M.A.; Chabo, C.; Waget, A.; Colom, A.; Rastrelli, S.; Delzenne, N.M.; Drucker, D.J.; et al. Role of central nervous system glucagon-like peptide- 1 receptors in enteric glucose sensing. Diabetes 2008, 57, 2603–2612. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Ishisaka, M.; Tsujii, S.; Shimazawa, M.; Hara, H. Glucagon-like peptide-1 protects the murine hippocampus against stressors via Akt and ERK1/2 signaling. Biochem. Biophys. Commun. 2015, 458, 274–279. [Google Scholar] [CrossRef]
- Harkavyi, A.; Whitton, P.S. Glucagon-like peptide 1 receptor stimulation as a means of neuroprotection. Br. J. Pharmacol. 2010, 159, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Abbas, T.; Faivre, E.; Holscher, C. Impairment of synaptic plasticity and memory formation in GLP-1 receptor KO mice: Interaction between type 2 diabetes and Alzheimer’s disease. Behav. Brain Res. 2009, 205, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.H.; Fabricius, K.; Barkholt, P.; Niehoff, M.L.; Morley, J.E.; Jelsing, J.; Pyke, C.; Knudsen, L.B.; Farr, S.A.; Vrang, N. The GLP-1 Receptor Agonist Liraglutide Improves Memory Function and Increases Hippocampal CA1 Neuronal Numbers in a Senescence-Accelerated Mouse Model of Alzheimer’s Disease. J. Alzheimer’s 2015, 46, 877–888. [Google Scholar] [CrossRef] [Green Version]
- Koshal, P.; Kumar, P. Effect of Liraglutide on Corneal Kindling Epilepsy Induced Depression and Cognitive Impairment in Mice. Neurochem. Res. 2016, 41, 1741–1750. [Google Scholar] [CrossRef]
- Koshal, P.; Kumar, P. Neurochemical modulation involved in the beneficial effect of liraglutide, GLP-1 agonist on PTZ kindling epilepsy-induced comorbidities in mice. Mol. Cell Biochem. 2016, 415, 77–87. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogram. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Hussein, A.M.; Adel, M.; El-Mesery, M.; Abbas, K.M.; Ali, A.N.; Abulseoud, O.A. l-Carnitine Modulates Epileptic Seizures in Pentylenetetrazole-Kindled Rats via Suppression of Apoptosis and Autophagy and Upregulation of Hsp70. Brain Sci. 2018, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Hunter, K.; Hölscher, C. Drugs developed to treat diabetes, liraglutide and lixisenatide, cross the blood brain barrier and enhance neurogenesis. BMC Neurosci. 2012, 13, 33. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Candeias, E.; Correia, S.; Santos, R.; Carvalho, C.; Cardoso, S.; Plácido, A.; Santos, M.S.; Oliveira, C.R.; Moreira, P.I.; et al. Crosstalk between diabetes and brain: glucagonlike peptide-1 mimetics as a promising therapy against neurodegeneration. Biochim. Biophys. Acta 2013, 1832, 527–541. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, S.; Zhang, T.; Zhao, X.; Liu, X.; Cao, L.; Chi, Z. Ascorbic acid ameliorates seizures and brain damage in rats through inhibiting autophagy. Brain Res. 2013, 1535, 115–123. [Google Scholar] [CrossRef]
- Saha, L.; Bhandari, S.; Bhatia, A.; Banerjee, D.; Chakrabarti, A. Anti-kindling Effect of Bezafibrate, a Peroxisome Proliferator-activated Receptors Alpha Agonist, in Pentylenetetrazole Induced Kindling Seizure Model. J. Epilepsy Res. 2014, 4, 45–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shehata, A.M. Neurophysiological Studies on the Effect of Acetone on Pentylenetetrazole-Induced Seizure in Rats. Bull. Egypt. Soc. Physiol. Sci. 2011, 31, 135–146. [Google Scholar]
- Zhao, X.; Liu, G.; Shen, H.; Gao, B.; Li, X.; Fu, J.; Zhou, J.; Ji, Q. Liraglutide inhibits autophagy and apoptosis induced by high glucose through GLP-1R in renal tubular epithelial cells. Int. J. Mol. Med. 2015, 35, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Han, R.; Liang, Z.Q.; Wu, J.C.; Zhang, X.D.; Gu, Z.L.; Qin, Z.H. An autophagic mechanism is involved in apoptotic death of rat striatal neurons induced by the non-N-methyl-Daspartate receptor agonist kainic acid. Autophagy 2008, 4, 214–226. [Google Scholar] [CrossRef]
- Shacka, J.J.; Lu, J.; Xie, Z.-L.; Uchiyama, Y.; Roth, K.A.; Zhang, J. Kainic acid induces early and transient autophagic stress in mouse hippocampus. Neurosci. Lett. 2007, 414, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Xu, J.; Lin, Y.; Zhao, X.; Liu, X.; Chi, Z. Autophagy is upregulated in rats with status epilepticus and partly inhibited by vitamin E. Biochem. Biophys. Res. Commun. 2009, 379, 949–995. [Google Scholar] [CrossRef]
- Scherz-Shouval, R.; Elazar, Z. ROS, mitochondria and the regulation of autophagy. Trends Cell Boil. 2007, 17, 422–427. [Google Scholar] [CrossRef]
- Scherz-Shouval, R.; Shvets, E.; Fass, E.; Shorer, H.; Gil, L.; Elazar, Z. Reactive oxygen species are essential for autophagy and specifically regulate the activity of Atg4. EMBO J. 2007, 26, 1749–1760. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.N.; Baehrecke, E.H. Caspases function in autophagic programmed cell death in Drosophila. Development 2004, 131, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Crighton, D.; Wilkinson, S.; O’Prey, J.; Syed, N.; Smith, P.; Harrison, P.R.; Gasco, M.; Garrone, O.; Crook, T.; Ryan, K.M. DRAM, a p53-Induced Modulator of Autophagy, Is Critical for Apoptosis. Cell 2006, 126, 121–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espert, L.; Denizot, M.; Grimaldi, M.; Robert-Hebmann, V.; Gay, B.; Varbanov, M.; Codogno, P.; Biard-Piechaczyk, M. Autophagy is involved in T cell death after binding of HIV-1 envelope proteins to CXCR4. J. Clin. Investig. 2006, 116, 2161–2172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, M.; Shibata, M.; Tadakoshi, M.; Gotoh, K.; Komatsu, M.; Waguri, S.; Kawahara, N.; Kuida, K.; Nagata, S.; Kominami, E.; et al. Inhibition of Autophagy Prevents Hippocampal Pyramidal Neuron Death after Hypoxic-Ischemic Injury. Am. J. Pathol. 2008, 172, 454–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Xu, J.; Lin, Y.; Zhao, X.; Liu, X.; Chi, Z. Vitamin E inhibits activated chaperone-mediated autophagy in rats with status epilepticus. Neuroscience 2009, 161, 73–77. [Google Scholar] [CrossRef]
- Hosseinzadeh, M.; Nikseresht, S.; Khodagholi, F.; Naderi, N.; Maghsoudi, N. Cannabidiol Post-Treatment Alleviates Rat Epileptic-Related Behaviors and Activates Hippocampal Cell Autophagy Pathway Along with Antioxidant Defense in Chronic Phase of Pilocarpine-Induced Seizure. J. Mol. Neurosci. 2016, 58, 432–440. [Google Scholar] [CrossRef]
- Fornai, F.; Longone, P.; Cafaro, L.; Kastsiuchenka, O.; Ferrucci, M.; Manca, M.L.; Lazzeri, G.; Spalloni, A.; Bellio, N.; Lenzi, P.; et al. Lithium delays progression of amyotrophic lateral sclerosis. Proc. Natl. Acad. Sci. USA 2008, 105, 2052–2057. [Google Scholar] [CrossRef] [Green Version]
- Calderó, J.; Brunet, N.; Tarabal, O.; Piedrafita, L.; Hereu, M.; Ayala, V.; E Esquerda, J. Lithium prevents excitotoxic cell death of motoneurons in organotypic slice cultures of spinal cord. Neuroscience 2010, 165, 1353–1369. [Google Scholar] [CrossRef]
- Jalewa, J.; Sharma, M.K.; Hölscher, C. Novel Incretin analogues improve autophagy and protect from mitochondrial stress induced by rotenone in SH-SY5Y cells. J. Neurochem. 2016, 139, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.-W.; Zhang, M.; Zhu, M. Liraglutide reduces lipid accumulation in steatotic L-02 cells by enhancing autophagy. Mol. Med. Rep. 2014, 10, 2351–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-F.; Li, Y.-B.; Han, J.-Y.; Yin, J.-J.; Wang, Y.; Zhu, L.-B.; Xie, G.-Y. Liraglutide prevents high glucose level induced insulinoma cells apoptosis by targeting autophagy. Chin. Med. J. 2013, 126, 937–941. [Google Scholar]
- Green, D.R.; Reed, J.C. Mitochondria and apoptosis. Science 1998, 281, 1309–1312. [Google Scholar] [CrossRef]
- Nassar, N.N.; Al-Shorbagy, M.Y.; Arab, H.H.; Abdallah, D.M. Saxagliptin: A novel antiparkinsonian approach. Neuropharmacology 2015, 89, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, Y.; Li, L.; Hölscher, C. Neuroprotective effects of geniposide in the MPTP mouse model of Parkinson’s disease. Eur. J. Pharmacol. 2015, 768, 21–27. [Google Scholar] [CrossRef]
- Soti, C.; Nagy, E.; Giricz, Z.; Vígh, L.; Csermely, P.; Ferdinandy, P. Heat shock proteins as emerging therapeutic targets. Br. J. Pharmacol. 2015, 146, 769–780. [Google Scholar] [CrossRef] [PubMed]
- A Foster, J.; Brown, I.R. Differential induction of heat shock mRNA in oligodendrocytes, microglia, and astrocytes following hyperthermia. Mol. Brain 1997, 45, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Gass, P.; Prior, P.; Kiessling, M. Correlation between seizure intensity and stress protein expression after limbic epilepsy in the rat brain. Neuroscience 1995, 65, 27–36. [Google Scholar] [CrossRef]
- Thom, M.; Seetah, S.; Sisodiya, S.; Koepp, M.; Scaravilli, F. Sudden and unexpected death in epilepsy (SUDEP): evidence of acute neuronal injury using HSP-70 and c-Jun immunohistochemistry. Neuropathol. Appl. Neurobiol. 2003, 29, 132–143. [Google Scholar] [CrossRef]
- Li, W.X.; Chen, C.H.; Ling, C.C.; Li, G.C. Apoptosis in heat induced cell killing: the protective role of hsp-70 and the sensitization eVect of the c-myc gene. Radiat. Res. 1996, 145, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.-G.; Ma, Q.-Z.; Xu, C.-X. Abrogation of heat-shock protein (HSP)70 expression induced cell growth inhibition and apoptosis in human androgen-independent prostate cancer cell line PC-3m. Asian J. Androl. 2004, 6, 319–324. [Google Scholar] [PubMed]
- Lee, C.S.; Montebello, J.; Rush, M.; Georgiou, T.; Wawryk, S.; Rode, J. Overexpression of heat shock protein (hsp) 70 associated with abnormal p53 expression in cancer of the pancreas. Zentralblatt fur Pathol. 1994, 140, 259–264. [Google Scholar]
- Merrick, B.A.; He, C.; Witcher, L.L.; Patterson, R.M.; Reid, J.J.; Pence-Pawlowski, P.M.; Selkirka, J.K. HSP binding and mitochondrial localization of p53 protein in human HT1080 and mouse C3H10T1/2 cell lines. Biochim. Biophys. Acta 1996, 1297, 57–68. [Google Scholar] [CrossRef]
- Kanitkar, M.; Bhonde, R.R. Curcumin treatment enhances islet recovery by induction of heat shock response proteins, Hsp70 and heme oxygenase-1, during cryopreservation. Life Sci. 2008, 82, 182–189. [Google Scholar] [CrossRef]
- Wisniewska, M.B. Physiological role of beta-catenin/TCF signaling in neurons of the adult brain. Neurochem. Res. 2013, 38, 1144–1155. [Google Scholar] [CrossRef]
- De Ferrari, G.V.; Papassotiropoulos, A.; Biechele, T.; Wavrant De-Vrieze, F.; Avila, M.E.; Major, M.B.; Myers, A.; Sáez, K.; Henríquez, J.P.; Zhao, A.; et al. Common genetic variation within the low-density lipoprotein receptor-related protein 6 and late-onset Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 9434–9439. [Google Scholar] [CrossRef]
- Lovestone, S.; Güntert, A.; Hye, A.; Lynham, S.; Thambisetty, M.; Ward, M. Proteomics of Alzheimer’s disease: understanding mechanisms and seeking biomarkers. Expert Rev. Proteom. 2007, 4, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, X.; Wang, Z.; Li, R. The Canonical Wnt Signaling Pathway in Autism. CNS Neurol. Disord. Drug Targets 2014, 13, 765–770. [Google Scholar] [CrossRef]
- Fasen, K.; Beck, H.; Elger, C.E.; Lie, A.A. Differential Regulation of Cadherins and Catenins During Axonal Reorganization in the Adult Rat CNS. J. Neuropathol. Exp. Neurol. 2002, 61, 903–913. [Google Scholar] [CrossRef] [Green Version]
- Madsen, T.M.; Newton, S.S.; E Eaton, M.; Russell, D.S.; Duman, R.S. Chronic electroconvulsive seizure up-regulates beta-catenin expression in rat hippocampus: role in adult neurogenesis. Biol. Psychiatry 2003, 54, 1006–1014. [Google Scholar] [CrossRef]
- Busceti, C.L.; Biagioni, F.; Aronica, E.; Riozzi, B.; Storto, M.; Battaglia, G.; Giorgi, F.S.; Gradini, R.; Fornai, F.; Caricasole, A.; et al. Induction of the Wnt inhibitor, Dickkopf-1, is associated with neurodegeneration related to temporal lobe epilepsy. Epilepsia 2007, 48, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, S.; Schleusner, D.; Pietrzik, C.; Skutella, T.; Behl, C. Glycogen synthase kinase 3beta links neuroprotection by 17 beta-estradiol to key Alzheimer processes. Neuroscience 2005, 132, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.E.; Du, M.; Li, Y. Increased seizure susceptibility and cortical malformation in beta-catenin mutant mice. Biochem. Biophys. Res. Commun. 2004, 320, 606–614. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage of Seizure | MDA | GSH | CAT | Caspase-3 | β-Catenin | Hsp70 | LC3 | ||
|---|---|---|---|---|---|---|---|---|---|
| Seizure stage | r | 0.77 | −0.73 | −0.35 | 0.64 | 0.64 | −0.62 | 0.66 | |
| p | 0.001 | 0.021 | 0.090 | 0.004 | 0.031 | 0.041 | 0.012 | ||
| MDA | r | −0.810 | −0.45 | 0.87 | 0.65 | −0.73 | 0.74 | ||
| p | 0.010 | 0.066 | 0.004 | 0.012 | 0.009 | 0.01 | |||
| GSH | r | 0.31 | −0.81 | 0.84 | 0.66 | −0.72 | |||
| p | 0.22 | 0.006 | 0.002 | 0.003 | 0.004 | ||||
| CAT | r | −0.14 | −0.31 | 0.62 | −0.66 | ||||
| p | 0.61 | 0.22 | 0.01 | 0.008 | |||||
| Caspase-3 | r | 0.67 | −0.61 | 0.67 | |||||
| p | 0.009 | 0.041 | 0.032 | ||||||
| β-catenin | r | 0.61 | 0.60 | ||||||
| p | 0.009 | 0.009 | |||||||
| Hsp70 | r | −0.33 | |||||||
| p | 0.076 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, A.M.; Eldosoky, M.; El-Shafey, M.; El-Mesery, M.; Abbas, K.M.; Ali, A.N.; Helal, G.M.; Abulseoud, O.A. Effects of GLP-1 Receptor Activation on a Pentylenetetrazole—Kindling Rat Model. Brain Sci. 2019, 9, 108. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9050108
Hussein AM, Eldosoky M, El-Shafey M, El-Mesery M, Abbas KM, Ali AN, Helal GM, Abulseoud OA. Effects of GLP-1 Receptor Activation on a Pentylenetetrazole—Kindling Rat Model. Brain Sciences. 2019; 9(5):108. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9050108
Chicago/Turabian StyleHussein, Abdelaziz M., Mohamed Eldosoky, Mohamed El-Shafey, Mohamed El-Mesery, Khaled M. Abbas, Amr N. Ali, Ghada M. Helal, and Osama A. Abulseoud. 2019. "Effects of GLP-1 Receptor Activation on a Pentylenetetrazole—Kindling Rat Model" Brain Sciences 9, no. 5: 108. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9050108