Pre- and Post-Treatment with Novel Antiepileptic Drug Oxcarbazepine Exerts Neuroprotective Effect in the Hippocampus in a Gerbil Model of Transient Global Cerebral Ischemia

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Treatment with OXC
2.3. Induction of TGCI
2.4. Tissue Preparation for Histology
2.5. Cresyl Violet (CV) Staining
2.6. Fluoro-Jade B (FJB) Histofluorescence Staining
2.7. Immunohistochemistry
2.8. Data Analysis
2.9. Statistical Analysis
3. Results
3.1. Neuroprotection
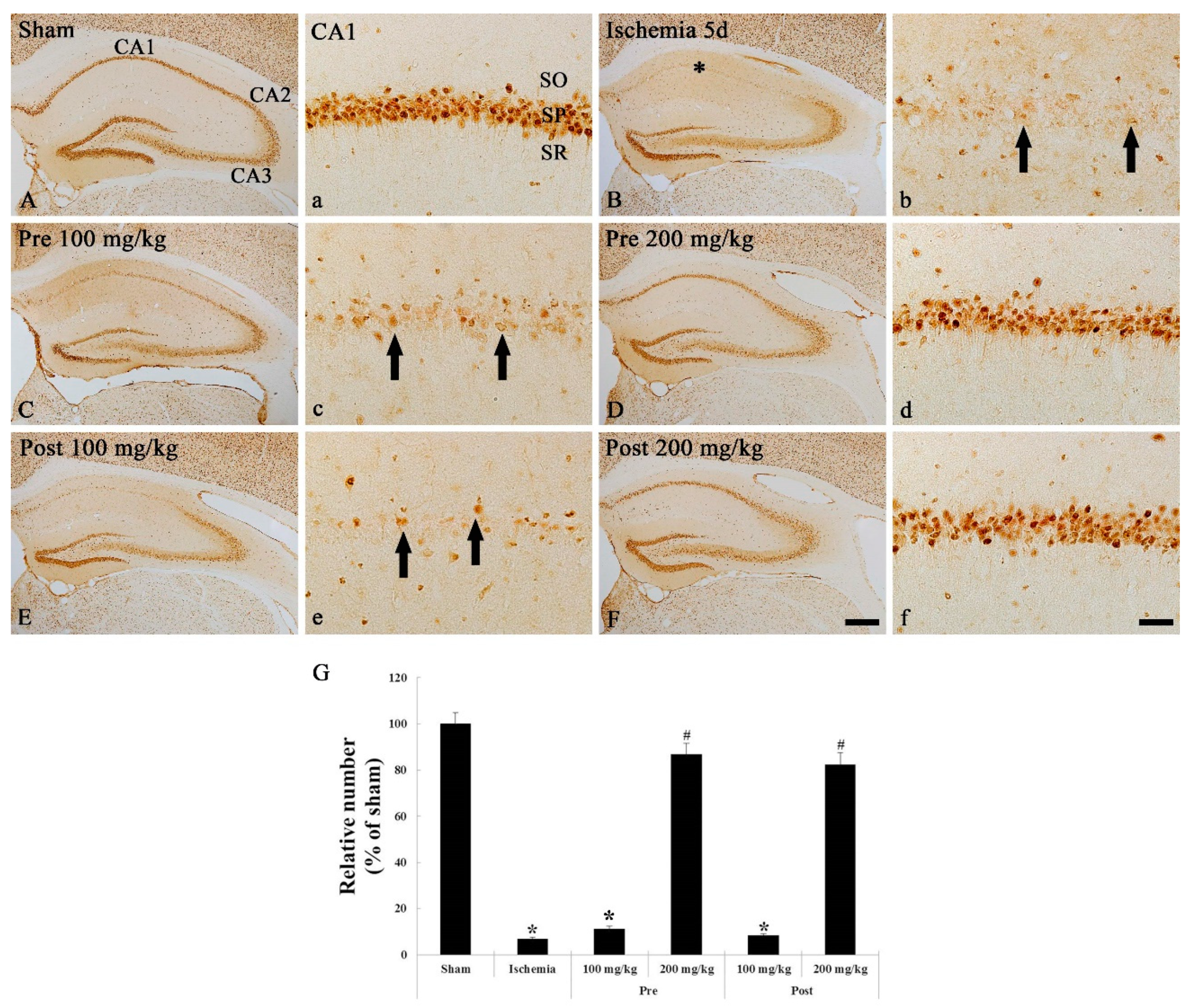
3.1.1. CV Staining
3.1.2. NeuN Immunohistochemistry
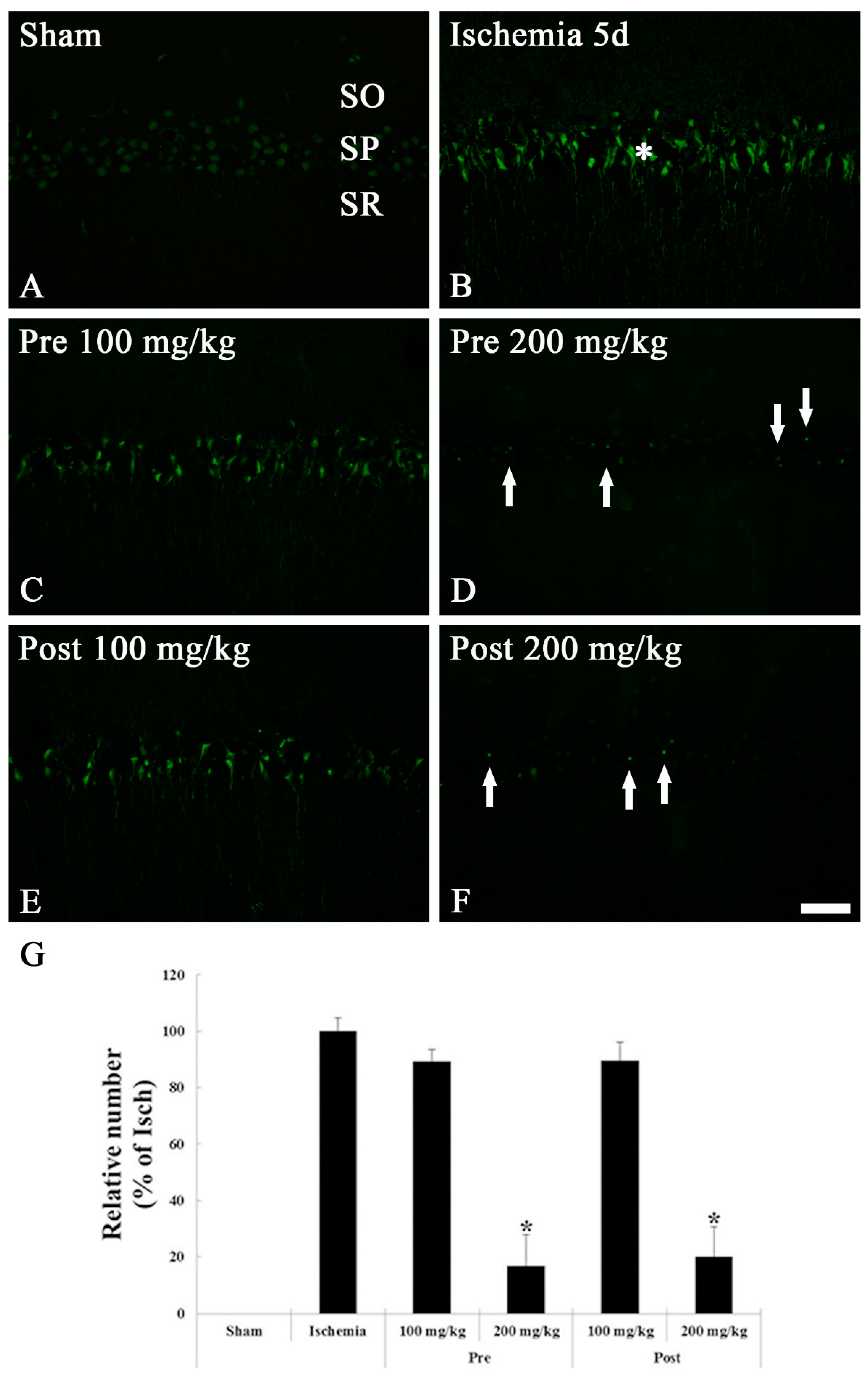
3.1.3. FJB Histofluorescence Staining
3.2. Glial Activation
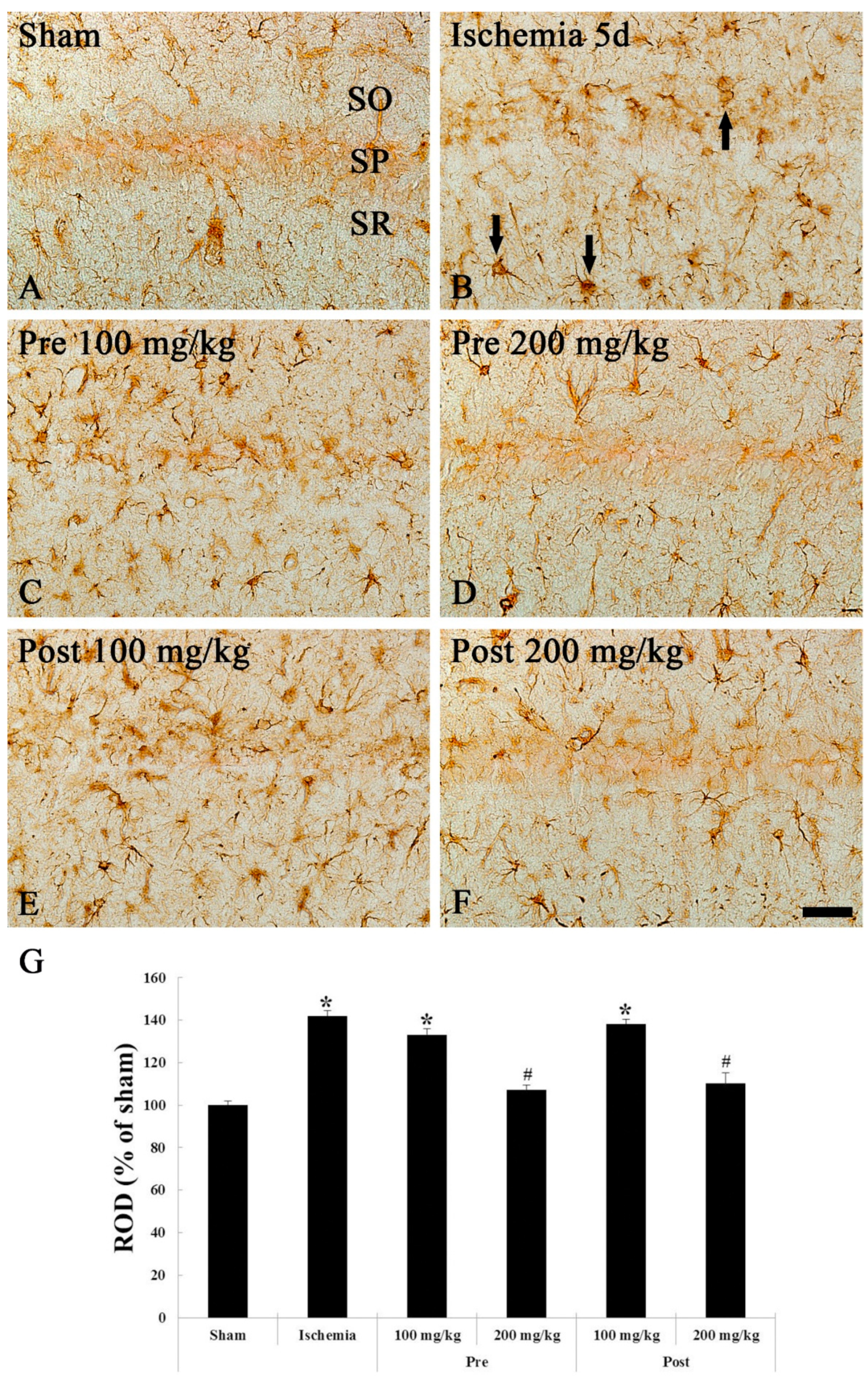
3.2.1. GFAP Immunohistochemistry
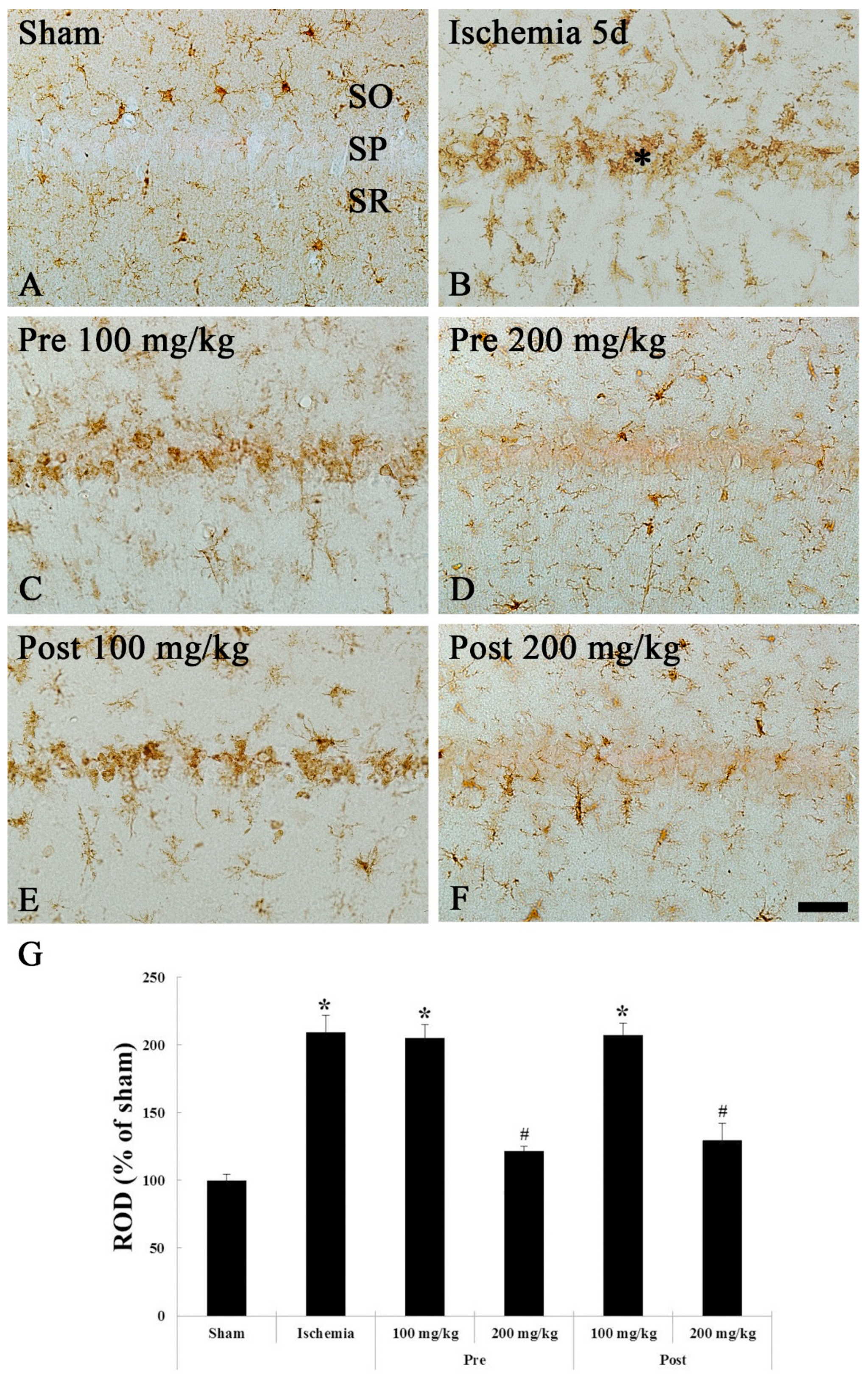
3.2.2. Iba-1 Immunohistochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lee, J.-M.; Grabb, M.C.; Zipfel, G.J.; Choi, D.W. Brain tissue responses to ischemia. J. Clin. Investig. 2000, 106, 723–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endres, M.; Heuschmann, P.U.; Laufs, U.; Hakim, A.M. Primary prevention of stroke: Blood pressure, lipids, and heart failure. Eur. Heart J. 2011, 32, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Petito, C.K.; Pulsinelli, W.A. Delayed Neuronal Recovery and Neuronal Death in Rat Hippocampus following Severe Cerebral Ischemia: Possible Relationship to Abnormalities in Neuronal Processes. Br. J. Pharmacol. 1984, 4, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T.; Sano, K. Selective vulnerability in the gerbil hippocampus following transient ischemia. Acta Neuropathol. 1984, 62, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T. Delayed neuronal death in the gerbil hippocampus following ischemia. Brain Res. 1982, 239, 57–69. [Google Scholar] [CrossRef]
- Kuroiwa, T.; Bonnekoh, P.; Hossmann, K.-A. Locomotor hyperactivity and hippocampal CA1 injury after transient forebrain ischemia of gerbils. Neurosci. Lett. 1991, 122, 141–144. [Google Scholar] [CrossRef]
- Andersen, M.B.; Zimmer, J.; Sams-Dodd, F. Postischemic hyperactivity in the Mongolian gerbil correlates with loss of hippocampal neurons. Behav. Neurosci. 1997, 111, 1205–1216. [Google Scholar] [CrossRef]
- Juurlink, B.H.; Sweeney, M.I. Mechanisms that Result in Damage During and Following Cerebral Ischemia. Neurosci. Biobehav. Rev. 1997, 21, 121–128. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Alvarez, D.; Merino, N.; León, O.S. Delayed treatment with nimesulide reduces measures of oxidative stress following global ischemic brain injury in gerbils. Neurosci. Res. 2003, 47, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Matsushima, K.; Schmidt-Kastner, R.; Hogan, M.J.; Hakim, A.M. Cortical spreading depression activates trophic factor expression in neurons and astrocytes and protects against subsequent focal brain ischemia. Brain Res. 1998, 807, 47–60. [Google Scholar] [CrossRef]
- Köhling, R. Voltage-gated sodium channels in epilepsy. Epilepsia 2002, 43, 1278–1295. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. GABAA receptor heterogeneity, function, and implications for epilepsy. Neurology 2007, 68, 612–614. [Google Scholar] [CrossRef] [PubMed]
- Benarroch, E.E. Potassium channels: Brief overview and implications in epilepsy. Neurology 2009, 72, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Rogawski, M.A.; Löscher, W. The neurobiology of antiepileptic drugs. Nat. Rev. Neurosci. 2004, 5, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Calabresi, P.; Cupini, L.M.; Centonze, D.; Pisani, F.; Bernardi, G. Antiepileptic drugs as a possible neuroprotective strategy in brain ischemia. Ann. Neurol. 2003, 53, 693–702. [Google Scholar] [CrossRef]
- Czuczwar, S.J.; Ferenc, R.; Błaszczyk, B.; Borowicz, K. Neuroprotective effects of some newer and potential antiepileptic drugs. J. Pre Clin. Clin. Res. 2007, 1, 1–5. [Google Scholar]
- Dam, M. Practical Aspects of Oxcarbazepine Treatment. Epilepsia 1994, 35, S23–S25. [Google Scholar] [CrossRef]
- A Swinyard, E.; Brown, W.C.; Goodman, L.S. Comparative assays of antiepileptic drugs in mice and rats. J. Pharmacol. Exp. Ther. 1952, 106, 319–330. [Google Scholar]
- Wamil, A.W.; Schmutz, M.; Portet, C.; Feldmann, K.F.; McLean, M.J. Effects of oxcarbazepine and 10-hydroxycarbamazepine on action potential firing and generalized seizures. Eur. J. Pharmacol. 1994, 271, 301–308. [Google Scholar] [CrossRef]
- Liu, Z.-B.; Liu, C.; Zeng, B.; Huang, L.-P.; Yao, L.-H. Modulation Effects of Cordycepin on Voltage-Gated Sodium Channels in Rat Hippocampal CA1 Pyramidal Neurons in the Presence/Absence of Oxygen. Neural Plast. 2017, 2017, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.P.; Burke, S.P.; Weber, M.L. Hippocampal slices: Glutamate overflow and cellular damage from ischemia are reduced by sodium-channel blockade. J. Neurosci. Methods 1995, 59, 121–128. [Google Scholar] [CrossRef]
- Ahn, J.Y.; Yan, B.C.; Park, J.H.; Ahn, J.H.; Lee, D.H.; Kim, I.H.; Cho, J.-H.; Chen, B.H.; Lee, J.-C.; Cho, Y.S.; et al. Novel antiepileptic drug lacosamide exerts neuroprotective effects by decreasing glial activation in the hippocampus of a gerbil model of ischemic stroke. Exp. Ther. Med. 2015, 10, 2007–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giustizieri, M.; Armogida, M.; Berretta, N.; Federici, M.; Piccirilli, S.; Mercuri, N.B.; Nistico, R. Differential effect of carbamazepine and oxcarbazepine on excitatory synaptic transmission in rat hippocampus. Synapse 2008, 62, 783–789. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Park, J.H.; Na Shin, B.; Chen, B.H.; Kim, I.H.; Ahn, J.H.; Cho, J.-H.; Tae, H.-J.; Lee, J.-C.; Lee, C.-H.; Kim, Y.-M.; et al. Neuroprotection and reduced gliosis by atomoxetine pretreatment in a gerbil model of transient cerebral ischemia. J. Neurol. Sci. 2015, 359, 373–380. [Google Scholar] [CrossRef]
- Park, J.H.; Shin, B.N.; Ahn, J.H.; Cho, J.H.; Kim, I.H.; Kim, D.W.; Won, M.H.; Hong, S.; Cho, J.H.; Lee, C.H.; et al. Ischemia-Induced Changes of PRAS40 and p-PRAS40 Immunoreactivities in the Gerbil Hippocampal CA1 Region After Transient Cerebral Ischemia. Cell Mol. Neurobiol. 2016, 36, 821–828. [Google Scholar] [CrossRef]
- Ahn, J.H.; Na Shin, B.; Park, J.H.; Kim, I.H.; Cho, J.H.; Chen, B.; Lee, T.-K.; Tae, H.-J.; Lee, J.-C.; Cho, J.H.; et al. Long-term observation of neuronal degeneration and microgliosis in the gerbil dentate gyrus after transient cerebral ischemia. J. Neurol. Sci. 2016, 363, 21–26. [Google Scholar] [CrossRef]
- Yoo, K.-Y.; Kim, I.H.; Cho, J.-H.; Ahn, J.H.; Park, J.H.; Lee, J.-C.; Tae, H.-J.; Kim, D.W.; Kim, J.-D.; Hong, S.; et al. Neuroprotection of Chrysanthemum indicum Linne against cerebral ischemia/reperfusion injury by anti-inflammatory effect in gerbils. Neural Regen. Res. 2016, 11, 270–277. [Google Scholar]
- Radtke-Schuller, S.; Schuller, G.; Angenstein, F.; Grosser, O.S.; Goldschmidt, J.; Budinger, E. Brain atlas of the Mongolian gerbil (Meriones unguiculatus) in CT/MRI-aided stereotaxic coordinates. Brain Struct. Funct. 2016, 221, 1–272. [Google Scholar] [CrossRef]
- Nitatori, T.; Sato, N.; Waguri, S.; Karasawa, Y.; Araki, H.; Shibanai, K.; Kominami, E.; Uchiyama, Y. Delayed neuronal death in the CA1 pyramidal cell layer of the gerbil hippocampus following transient ischemia is apoptosis. J. Neurosci. 1995, 15, 1001–1011. [Google Scholar] [CrossRef] [Green Version]
- Zhan, L.; Lu, Z.; Zhu, X.; Xu, W.; Li, L.; Li, X.; Chen, S.; Sun, W.; Xu, E. Hypoxic preconditioning attenuates necroptotic neuronal death induced by global cerebral ischemia via Drp1-dependent signaling pathway mediated by CaMKIIalpha inactivation in adult rats. FASEB J. 2019, 33, 1313–1329. [Google Scholar] [CrossRef]
- Pagnussat, A.S.; Faccioni-Heuser, M.C.; Netto, C.A.; Achaval, M. An ultrastructural study of cell death in the CA1 pyramidal field of the hippocapmus in rats submitted to transient global ischemia followed by reperfusion. J. Anat. 2007, 211, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.J.; Sieber, F.E.; Traystman, R.J. Apoptosis and Necrosis Occur in Separate Neuronal Populations in Hippocampus and Cerebellum after Ischemia and Are Associated with Differential Alterations in Metabotropic Glutamate Receptor Signaling Pathways. Br. J. Pharmacol. 2000, 20, 153–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Gao, Y.; Barrett, J.; Hu, B. Autophagy and protein aggregation after brain ischemia. J. Neurochem. 2010, 115, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festing, M.F.W.; Altman, D.G. Guidelines for the design and statistical analysis of experiments using laboratory animals. ILAR J. 2002, 43, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Holland-Letz, T.; Kopp-Schneider, A. Optimal experimental designs for dose-response studies with continuous endpoints. Arch. Toxicol. 2015, 89, 2059–2068. [Google Scholar] [CrossRef] [PubMed]
- Iadecola, C.; Alexander, M. Cerebral ischemia and inflammation. Curr. Opin. Neurol. 2001, 14, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Stoll, G.; Jander, S.; Schroeter, M. Inflammation and glial responses in ischemic brain lesions. Prog. Neurobiol. 1998, 56, 149–171. [Google Scholar] [CrossRef]
- Günther, A.; Schneider, P.-M.; Kunert, I.; Berrouschot, J.; Schneider, D.; Roßner, S.; Küppers-Tiedt, L.; Küppers-Tiedt, L. Reduced infarct volume and differential effects on glial cell activation after hyperbaric oxygen treatment in rat permanent focal cerebral ischaemia. Eur. J. Neurosci. 2005, 21, 3189–3194. [Google Scholar] [CrossRef]
- Wang, Z.-Q.; Wu, D.-C.; Huang, F.-P.; Yang, G.-Y. Inhibition of MEK/ERK 1/2 pathway reduces pro-inflammatory cytokine interleukin-1 expression in focal cerebral ischemia. Brain Res. 2004, 996, 55–66. [Google Scholar] [CrossRef]
- Yan, B.C.; Kim, S.K.; Park, J.H.; Ahn, J.H.; Lee, C.H.; Yoo, K.-Y.; Choi, J.H.; Lee, D.-S.; Kim, M.J.; Kim, Y.-M.; et al. Comparison of inflammatory cytokines changes in the hippocampal CA1 region between the young and adult gerbil after transient cerebral ischemia. Brain Res. 2012, 1461, 64–75. [Google Scholar] [CrossRef]
- Al-Izki, S.; Pryce, G.; Hankey, D.J.; Lidster, K.; von Kutzleben, S.M.; Browne, L.; Clutterbuck, L.; Posada, C.; Edith Chan, A.W.; Amor, S.; et al. Lesional-targeting of neuroprotection to the inflammatory penumbra in experimental multiple sclerosis. Brain 2014, 137, 92–108. [Google Scholar] [CrossRef] [PubMed]
- Gómez, C.D.; Buijs, R.M.; Sitges, M. The anti-seizure drugs vinpocetine and carbamazepine, but not valproic acid, reduce inflammatory IL-1β and TNF-α expression in rat hippocampus. J. Neurochem. 2014, 130, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, P.E.; Zhu, L.; Verret, L.; Vossel, K.A.; Orr, A.G.; Cirrito, J.R.; Devidze, N.; Ho, K.; Yu, G.Q.; Palop, J.J.; et al. Levetiracetam suppresses neuronal network dysfunction and reverses synaptic and cognitive deficits in an Alzheimer’s disease model. Proc. Natl. Acad. Sci. USA 2012, 109, E2895–E2903. [Google Scholar] [CrossRef] [PubMed]
- Vajda, F.J.E. Neuroprotection and neurodegenerative disease. In Alzheimer’s Disease; Richter, R.W., Richter, B.Z., Eds.; Humana Press: Totowa, NJ, USA, 2004; pp. 235–243. [Google Scholar]
- Kapoor, R.; Furby, J.; Hayton, T.; Smith, K.J.; Altmann, D.R.; Brenner, R.; Chataway, J.; Hughes, R.A.; Miller, D.H. Lamotrigine for neuroprotection in secondary progressive multiple sclerosis: A randomised, double-blind, placebo-controlled, parallel-group trial. Lancet Neurol. 2010, 9, 681–688. [Google Scholar] [CrossRef]
- Schapira, A.H.V.; Olanow, C.W.; Greenamyre, J.T.; Bezard, E. Slowing of neurodegeneration in Parkinson’s disease and Huntington’s disease: Future therapeutic perspectives. Lancet 2014, 384, 545–555. [Google Scholar] [CrossRef]
- Meldrum, B.S.; Rogawski, M.A. Molecular targets for antiepileptic drug development. Neurotherapeutics 2007, 4, 18–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepień, K.; Tomaszewski, M.; Czuczwar, S.J. Profile of anticonvulsant activity and neuroprotective effects of novel and potential antiepileptic drugs—An update. Pharmacol. Rep. 2005, 57, 719–733. [Google Scholar]
- Wang, S.-J.; Sihra, T.S.; Gean, P.-W. Lamotrigine inhibition of glutamate release from isolated cerebrocortical nerve terminals (synaptosomes) by suppression of voltage-activated calcium channel activity. Neuroreport 2001, 12, 2255–2258. [Google Scholar] [CrossRef]
- Rekling, J.C. Neuroprotective effects of anticonvulsants in rat hippocampal slice cultures exposed to oxygen/glucose deprivation. Neurosci. Lett. 2003, 335, 167–170. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, S.K.; Chung, J.H.; Kim, J.W. Protective effect of carbamazepine on kainic acid-induced neuronal cell death through activation of signal transducer and activator of transcription. J. Mol. Neurosci. 2013, 49, 172–181. [Google Scholar] [CrossRef]
- Wasterlain, C.G.; Adams, L.M.; Wichmann, J.K.; Sofia, R.D.; MacManus, J. Felbamate Protects CA1 Neurons from Apoptosis in a Gerbil Model of Global Ischemia. Stroke 1996, 27, 1236–1240. [Google Scholar] [CrossRef] [PubMed]
- Wasterlain, C.G.; Adams, L.M.; Schwartz, P.H.; Hattori, H.; Sofia, R.D.; Wichmann, J.K. Posthypoxic treatment with felbamate is neuroprotective in a rat model of hypoxia-ischemia. Neurology 1993, 43, 2303. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, J.H.; Shin, B.N.; Park, J.H.; Lee, T.-K.; Park, Y.E.; Lee, J.-C.; Yang, G.E.; Shin, M.C.; Cho, J.H.; Lee, K.C.; et al. Pre- and Post-Treatment with Novel Antiepileptic Drug Oxcarbazepine Exerts Neuroprotective Effect in the Hippocampus in a Gerbil Model of Transient Global Cerebral Ischemia. Brain Sci. 2019, 9, 279. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9100279
Ahn JH, Shin BN, Park JH, Lee T-K, Park YE, Lee J-C, Yang GE, Shin MC, Cho JH, Lee KC, et al. Pre- and Post-Treatment with Novel Antiepileptic Drug Oxcarbazepine Exerts Neuroprotective Effect in the Hippocampus in a Gerbil Model of Transient Global Cerebral Ischemia. Brain Sciences. 2019; 9(10):279. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9100279
Chicago/Turabian StyleAhn, Ji Hyeon, Bich Na Shin, Joon Ha Park, Tae-Kyeong Lee, Young Eun Park, Jae-Chul Lee, Go Eun Yang, Myoung Cheol Shin, Jun Hwi Cho, Kyu Chang Lee, and et al. 2019. "Pre- and Post-Treatment with Novel Antiepileptic Drug Oxcarbazepine Exerts Neuroprotective Effect in the Hippocampus in a Gerbil Model of Transient Global Cerebral Ischemia" Brain Sciences 9, no. 10: 279. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9100279