Rotating and Neurochemical Activity of Rats Lesioned with Quinolinic Acid and Transplanted with Bone Marrow Mononuclear Cells

 , and
, and 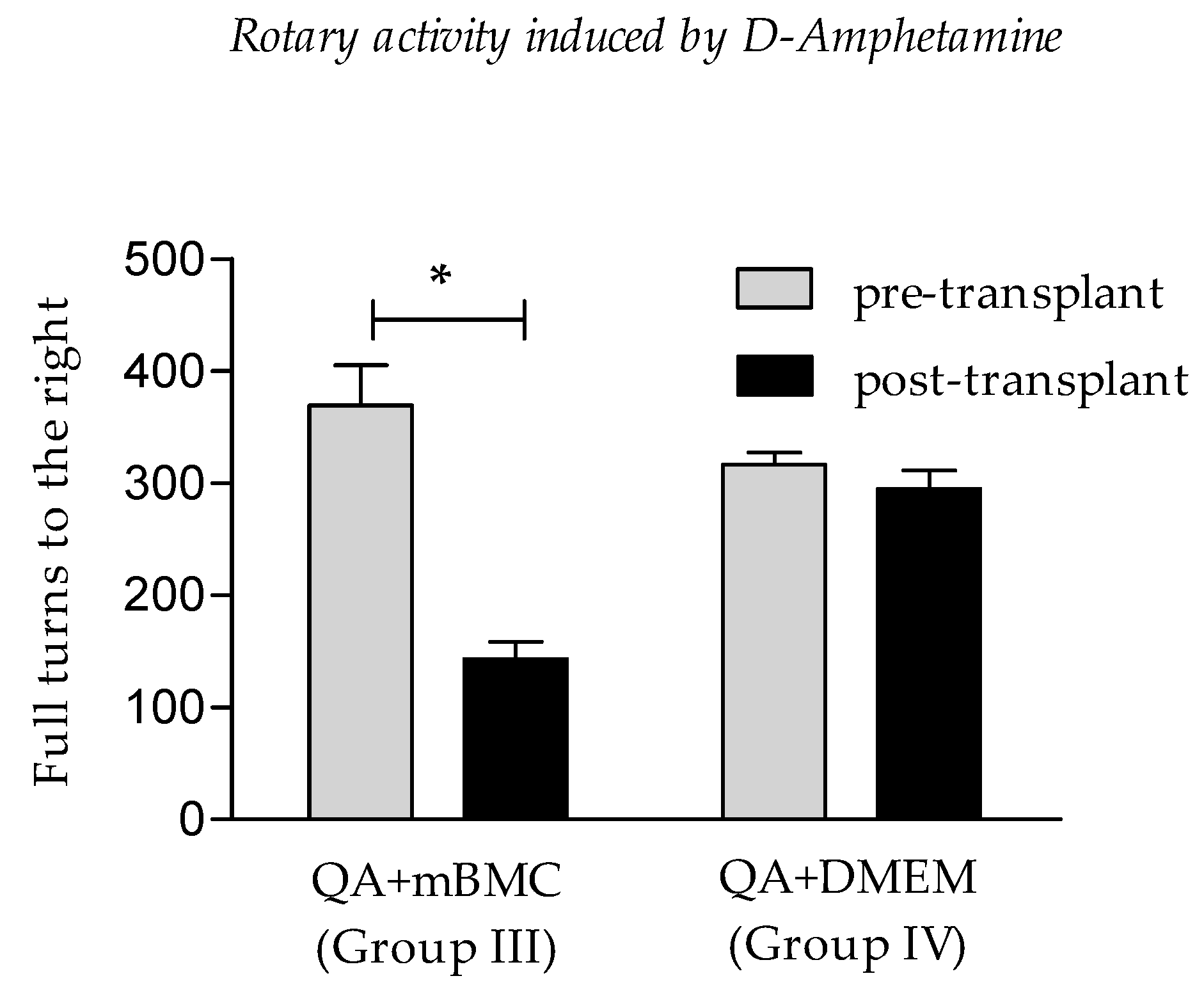{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Striatal QA Lesions
2.3. Obtaining Rat Mononuclear Bone Marrow Cells
2.4. Isolation of Mononuclear Bone Marrow Cells
2.5. Transplantation
2.6. Behavioral Tests (Rotating Activity Induced by D-Amphetamine)
2.7. Obtaining Samples for Neurotransmission Study
2.8. Neurotransmitter Study
2.9. Ethical Considerations
2.10. Statistical Processing
3. Results
3.1. Rotational Activity
3.2. Neurochemical Activity
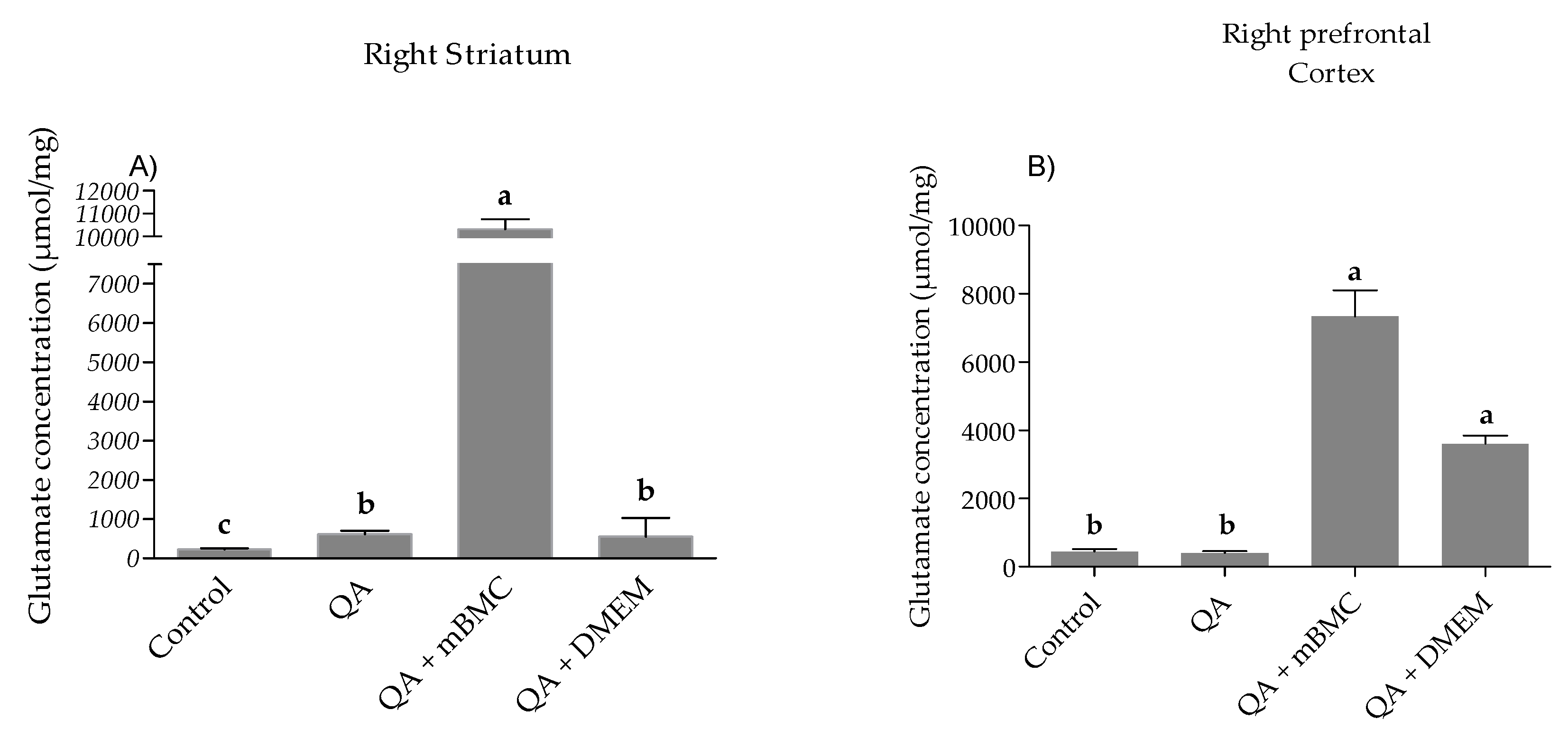
3.2.1. Glutamate Concentration
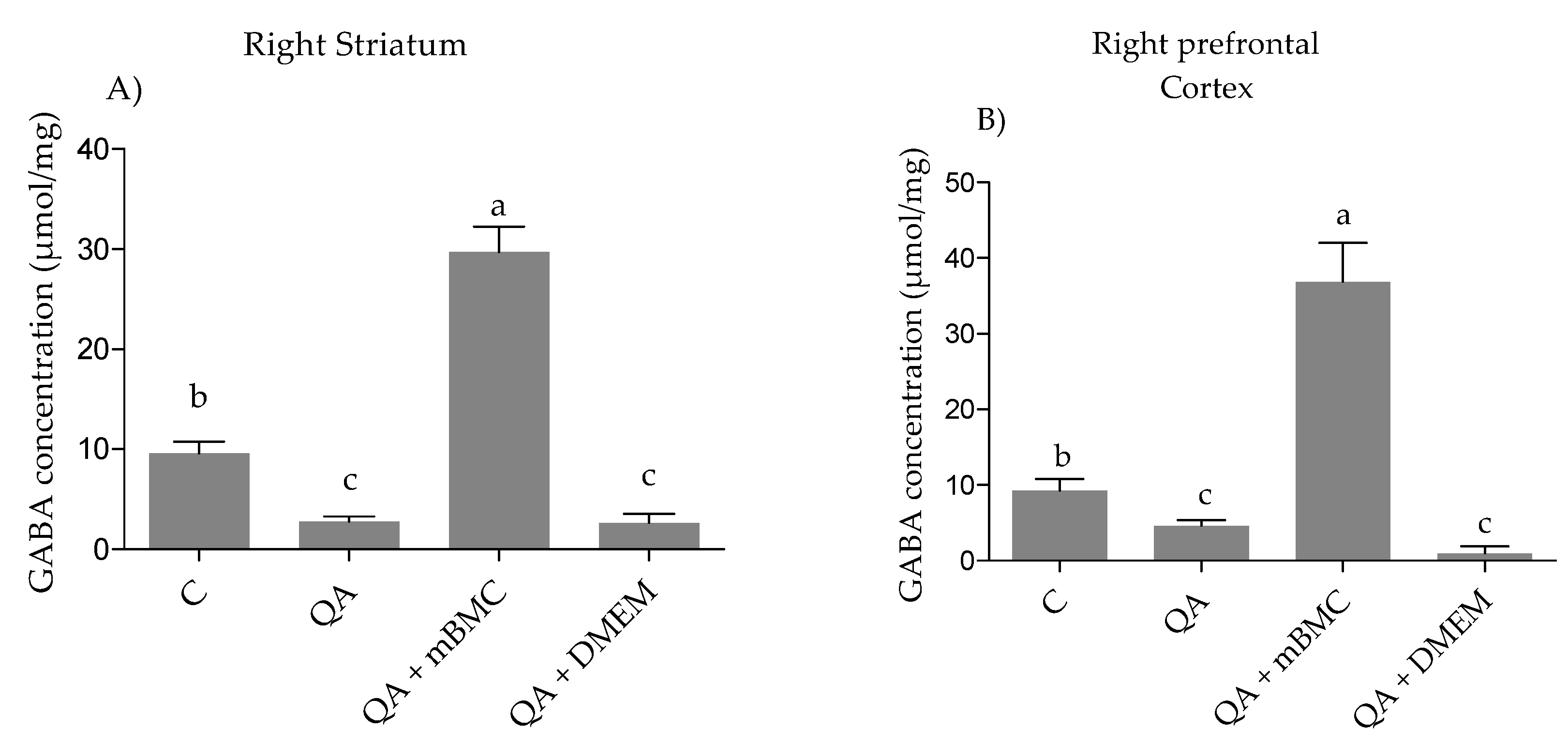
3.2.2. GABA (γ-Aminobutyric Acid) Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dayalu, P.; Albin, R.L. Huntington disease: Pathogenesis and treatment. Neurol. Clin. 2015, 33, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Davies, S.; Ramsden, D.B. Huntington’s disease. Mol. Pathol. 2001, 54, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Araujo, D.M.; Hilt, D.C. Glial cell line-derived neurotrophic factor attenuates the excitotoxin-induced behavioral and neurochemical deficits in a rodent model of Huntington’s disease. Neuroscience 1997, 81, 1099–1110. [Google Scholar] [CrossRef]
- Cox, C.S., Jr.; Hetz, R.A.; Liao, G.P.; Aertker, B.M.; Ewing-Cobbs, L.; Juranek, J.; Savitz, S.I.; Jackson, M.L.; Romanowska-Pawliczek, A.M.; Triolo, F.; et al. Treatment of Severe Adult Traumatic Brain Injury Using Bone Marrow Mononuclear Cells. Stem Cells 2017, 35, 1065–1079. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Deeb, G.; Rahal, R.; Atwi, K.; Mondello, S.; Marei, H.E.; Gali, A.; Sleiman, E. Mesenchymal Stem Cells in the Treatment of Traumatic Brain Injury. Front Neurol. 2017, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Y.; Shen, L.; Ding, W.; Chen, X.; Wu, E.; Cai, K.; Wang, G. Hypoxic Preconditioning Augments the Therapeutic Efficacy of Bone Marrow Stromal Cells in a Rat Ischemic Stroke Model. Cell. Mol. Neurobiol. 2017, 37, 1115–1129. [Google Scholar] [CrossRef] [PubMed]
- Lescaudron, L.; Unni, D.; Dunbar, G.L. Autologous adult bone marrow stem cell transplantation in an animal model of huntington’s disease: Behavioral and morphological outcomes. Int. J. Neurosci. 2003, 113, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Kerkis, I.; Haddad, M.S.; Valverde, C.W.; Glosman, S. Neural and mesenchymal stem cells in animal models of Huntington’s disease: Past experiences and future challenges. Stem Cell Res. Ther. 2015, 6, 232. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.; Fink, K.D.; Crane, A.T.; Davis, K.K.; Bombard, M.C.; Clerc, S.; Bavar, A.M.; Lowrance, S.A.; Song, C.; Witte, S.; et al. Reductions in behavioral deficits and neuropathology in the R6/2 mouse model of Huntington’s disease following transplantation of bone-marrow-derived mesenchymal stem cells is dependent on passage number. Stem Cell Res. Ther. 2015, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodbury, D.; Schwarz, E.J.; Prockop, D.J.; Black, I.B. Adult rat and human bone marrow stromal cells differentiate into neurons. J. Neurosci. Res. 2000, 61, 364–370. [Google Scholar] [CrossRef] [Green Version]
- Ungerstedt, U.; Arbuthnott, G.W. Quantitative recording of rotational behavior in rats after 6-hydroxy-dopamine lesions of the nigrostriatal dopamine system. Brain Res. 1970, 24, 485–493. [Google Scholar] [CrossRef]
- National Research Council (US). National Research Council (US) Committee on Guidelines for the Care and Use of Animals in Neuroscience and Behavioral Research; National Academies Press: Washington, DC, USA, 2003; pp. 1–224. [Google Scholar]
- DeLong, M.; Wichmann, T. Changing views of basal ganglia circuits and circuit disorders. Clin. EEG Neurosci. 2010, 41, 61–67. [Google Scholar] [CrossRef] [PubMed]
- DeLong, M.R.; Wichmann, T. Basal Ganglia Circuits as Targets for Neuromodulation in Parkinson Disease. JAMA Neurol. 2015, 72, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Ribak, C.E.; Vaughn, J.E.; Roberts, E. The GABA neurons and their axon terminals in rat corpus striatum as demonstrated by GAD immunocytochemistry. J. Comp. Neurol. 1979, 187, 261–283. [Google Scholar] [CrossRef] [PubMed]
- Bergman, H.; Feingold, A.; Nini, A.; Raz, A.; Slovin, H.; Abeles, M.; Vaadia, E. Physiological aspects of information processing in the basal ganglia of normal and parkinsonian primates. Trends Neurosci. 1998, 21, 32–38. [Google Scholar] [CrossRef]
- Kelly, R.M.; Strick, P.L. Macro-architecture of basal ganglia loops with the cerebral cortex: Use of rabies virus to reveal multisynaptic circuits. Prog. Brain Res. 2004, 143, 449–459. [Google Scholar] [PubMed]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Rodriguez, M.; Lanciego, J.L.; Artieda, J.; Gonzalo, N.; Olanow, C.W. Pathophysiology of the basal ganglia in Parkinson’s disease. Trends Neurosci. 2000, 23, S8–S19. [Google Scholar] [CrossRef]
- Hantraye, P.; Riche, D.; Maziere, M.; Isacson, O. A primate model of Huntington’s disease: Behavioral and anatomical studies of unilateral excitotoxic lesions of the caudate-putamen in the baboon. Exp. Neurol. 1990, 108, 91–104. [Google Scholar] [CrossRef]
- Schwarcz, R.; Fuxe, K.; Agnati, L.F.; Hokfelt, T.; Coyle, J.T. Rotational behaviour in rats with unilateral striatal kainic acid lesions: A behavioural model for studies on intact dopamine receptors. Brain Res. 1979, 170, 485–495. [Google Scholar] [CrossRef]
- Fuxe, K.; Janson, A.M.; Jansson, A.; Andersson, K.; Eneroth, P.; Agnati, L.F. Chronic nicotine treatment increases dopamine levels and reduces dopamine utilization in substantia nigra and in surviving forebrain dopamine nerve terminal systems after a partial di-mesencephalic hemitransection. Naunyn Schmiedebergs Arch. Pharmacol. 1990, 341, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.T.; Alberti, A.E.; Lorigados, P.L.; Blanco, L.L.; Diaz, A.I.; Bergado, J.A. BDNF in quinolinic acid lesioned rats after bone marrow cells transplant. Neurosci. Lett. 2014, 559, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Rossignol, J.; Fink, K.; Davis, K.; Clerc, S.; Crane, A.; Matchynski, J.; Lowrance, S.; Bombard, M.; Dekorver, N.; Lescaudron, L.; et al. Transplants of adult mesenchymal and neural stem cells provide neuroprotection and behavioral sparing in a transgenic rat model of Huntington’s disease. Stem Cells 2014, 32, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Tattersfield, A.S.; Croon, R.J.; Liu, Y.W.; Kells, A.P.; Faull, R.L.; Connor, B. Neurogenesis in the striatum of the quinolinic acid lesion model of Huntington’s disease. Neuroscience 2004, 127, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Del Barco, D.G.; Berlanga, J.; Penton, E.; Hardiman, O.; Montero, E. Boosting controlled autoimmunity: A new therapeutic target for CNS disorders. Expert Rev. Neurother. 2008, 8, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N. Neurotransmitters in the human and nonhuman primate basal ganglia. Hum. Neurobiol. 1986, 5, 159–168. [Google Scholar] [PubMed]
- Moberly, A.H.; Czarnecki, L.A.; Pottackal, J.; Rubinstein, T.; Turkel, D.J.; Kass, M.D.; McGann, J.P. Intranasal exposure to manganese disrupts neurotransmitter release from glutamatergic synapses in the central nervous system in vivo. Neurotoxicology 2012, 33, 996–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van, D.E.; Reedeker, N.; Giltay, E.J.; Eindhoven, D.; Roos, R.A.; van der Mast, R.C. Course of irritability, depression and apathy in Huntington’s disease in relation to motor symptoms during a two-year follow-up period. Neurodegener. Dis. 2014, 13, 9–16. [Google Scholar]
- Lau, A.; Tymianski, M. Glutamate receptors, neurotoxicity and neurodegeneration. Pflugers Arch. 2010, 460, 525–542. [Google Scholar] [CrossRef] [PubMed]
- Ahmado, A.; Carr, A.J.; Vugler, A.A.; Semo, M.; Gias, C.; Lawrence, J.M.; Chen, L.L.; Chen, F.K.; Turowski, P.; da Cruz, L.; Coffey, P.J. Induction of differentiation by pyruvate and DMEM in the human retinal pigment epithelium cell line ARPE-19. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7148–7159. [Google Scholar] [CrossRef] [PubMed]
- Braidy, N.; Grant, R.; Adams, S.; Brew, B.J.; Guillemin, G.J. Mechanism for quinolinic acid cytotoxicity in human astrocytes and neurons. Neurotox. Res. 2009, 16, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J. Quinolinic acid, the inescapable neurotoxin. FEBS J. 2012, 279, 1356–1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St’astny, F.; Lisy, V.; Mares, V.; Lisa, V.; Balcar, V.J.; Santamaria, A. Quinolinic acid induces NMDA receptor-mediated lipid peroxidation in rat brain microvessels. Redox. Rep. 2004, 9, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Marin, O. Origin of cortical interneurons: Basic concepts and clinical implications. Rev. Neurol. 2002, 35, 743–751. [Google Scholar] [PubMed]
- Alberti, E.; Los, M.; Garcia, R.; Fraga, J.L.; Serrano, T.; Hernandez, E.; Klonisch, T.; Macias, R.; Martinez, L.; Castillo, L.; de la Cuétara, K. Prolonged survival and expression of neural markers by bone marrow-derived stem cells transplanted into brain lesions. Med. Sci. Monit. 2009, 15, BR47–BR54. [Google Scholar] [PubMed]
- Serrano, T.; Pierozan, P.; Alberti, E.; Blanco, L.; de la Cuétara Bernal, K.; González, M.E.; Pavón, N.; Lorigados, L.; Robinson-Agramonte, M.A.; Bergado, J.A. Transplantation of mononuclear cells from bone marrow in a rat model of Huntington’s disease. Neurorestoratology 2016, 4, 95–105. [Google Scholar] [CrossRef]
- Teresa, S.-S. Havana: International Center for Neurological Restoration (CIREN). Ph.D. Thesis, Cuba University, Havana, Cuba, 2017. [Google Scholar]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano Sánchez, T.; González Fraguela, M.E.; Blanco Lezcano, L.; Alberti Amador, E.; Caballero Fernández, B.; Robinson Agramonte, M.D.l.Á.; Lorigados Pedre, L.; Bergado Rosado, J.A. Rotating and Neurochemical Activity of Rats Lesioned with Quinolinic Acid and Transplanted with Bone Marrow Mononuclear Cells. Behav. Sci. 2018, 8, 87. https://0-doi-org.brum.beds.ac.uk/10.3390/bs8100087
Serrano Sánchez T, González Fraguela ME, Blanco Lezcano L, Alberti Amador E, Caballero Fernández B, Robinson Agramonte MDlÁ, Lorigados Pedre L, Bergado Rosado JA. Rotating and Neurochemical Activity of Rats Lesioned with Quinolinic Acid and Transplanted with Bone Marrow Mononuclear Cells. Behavioral Sciences. 2018; 8(10):87. https://0-doi-org.brum.beds.ac.uk/10.3390/bs8100087
Chicago/Turabian StyleSerrano Sánchez, Teresa, María Elena González Fraguela, Lisette Blanco Lezcano, Esteban Alberti Amador, Beatriz Caballero Fernández, María De los Ángeles Robinson Agramonte, Lourdes Lorigados Pedre, and Jorge A Bergado Rosado. 2018. "Rotating and Neurochemical Activity of Rats Lesioned with Quinolinic Acid and Transplanted with Bone Marrow Mononuclear Cells" Behavioral Sciences 8, no. 10: 87. https://0-doi-org.brum.beds.ac.uk/10.3390/bs8100087