New Interactors of the Truncated EBNA-LP Protein Identified by Mass Spectrometry in P3HR1 Burkitt’s Lymphoma Cells

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
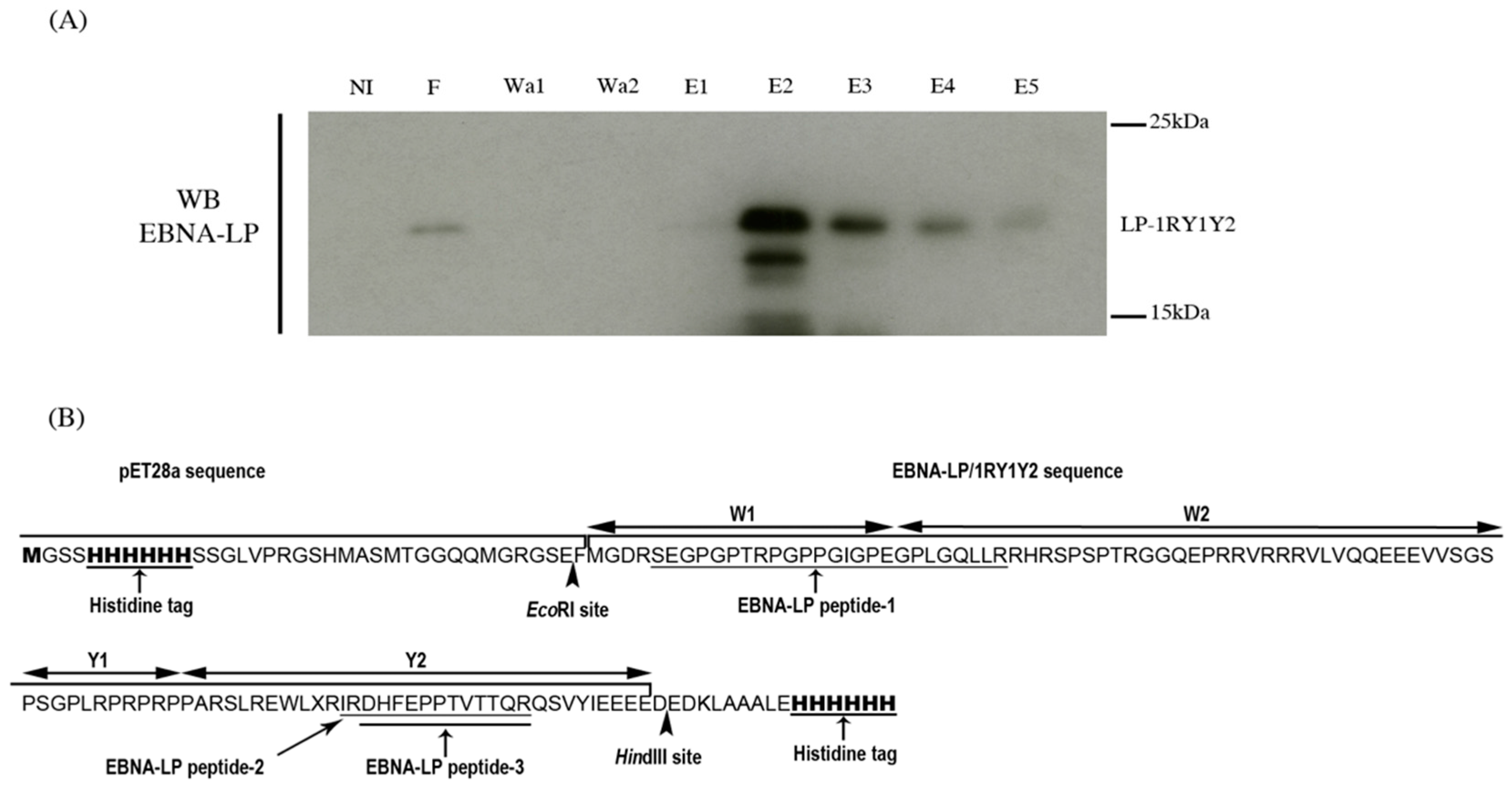
2.1. Production of Recombinant EBNA-LP and Identification of Its Specific Peptides by Mass Spectrometry
2.2. Identification of Truncated EBNA-LP Partners by Mass Spectrometry
2.3. Functional Annotation of tEBNA-LP Partners
2.4. Validation of Novel Protein Interactions Identified by Mass Spectrometry
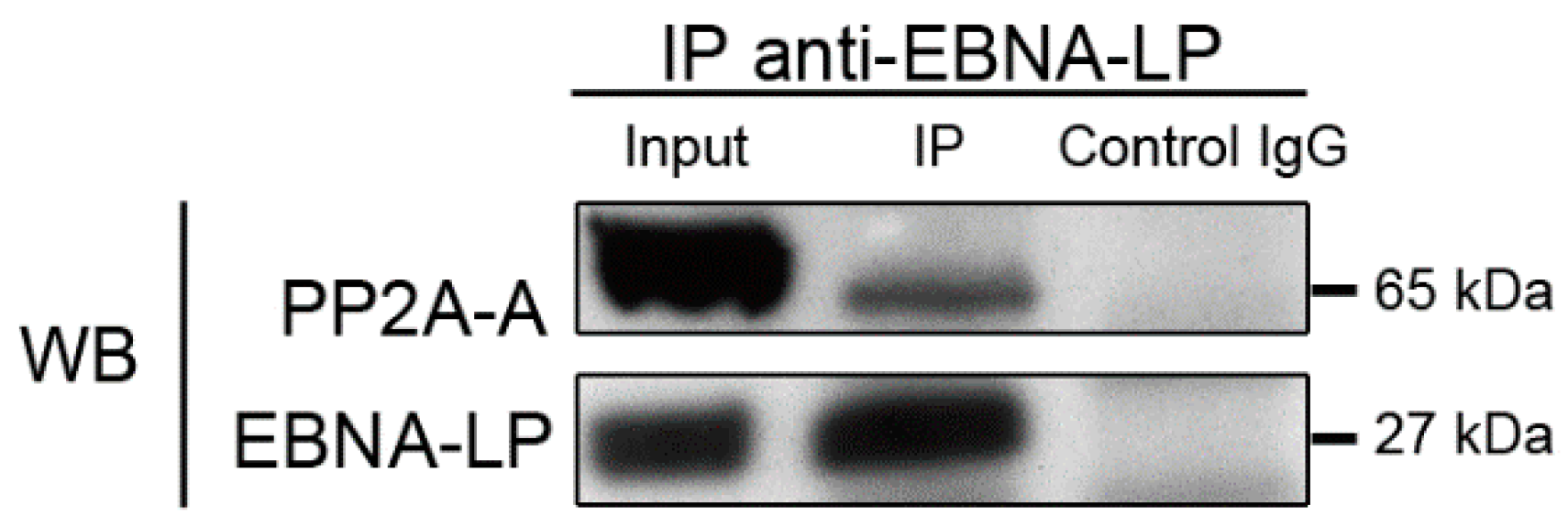
2.4.1. Truncated EBNA-LP interacts with the structural subunit of PP2A
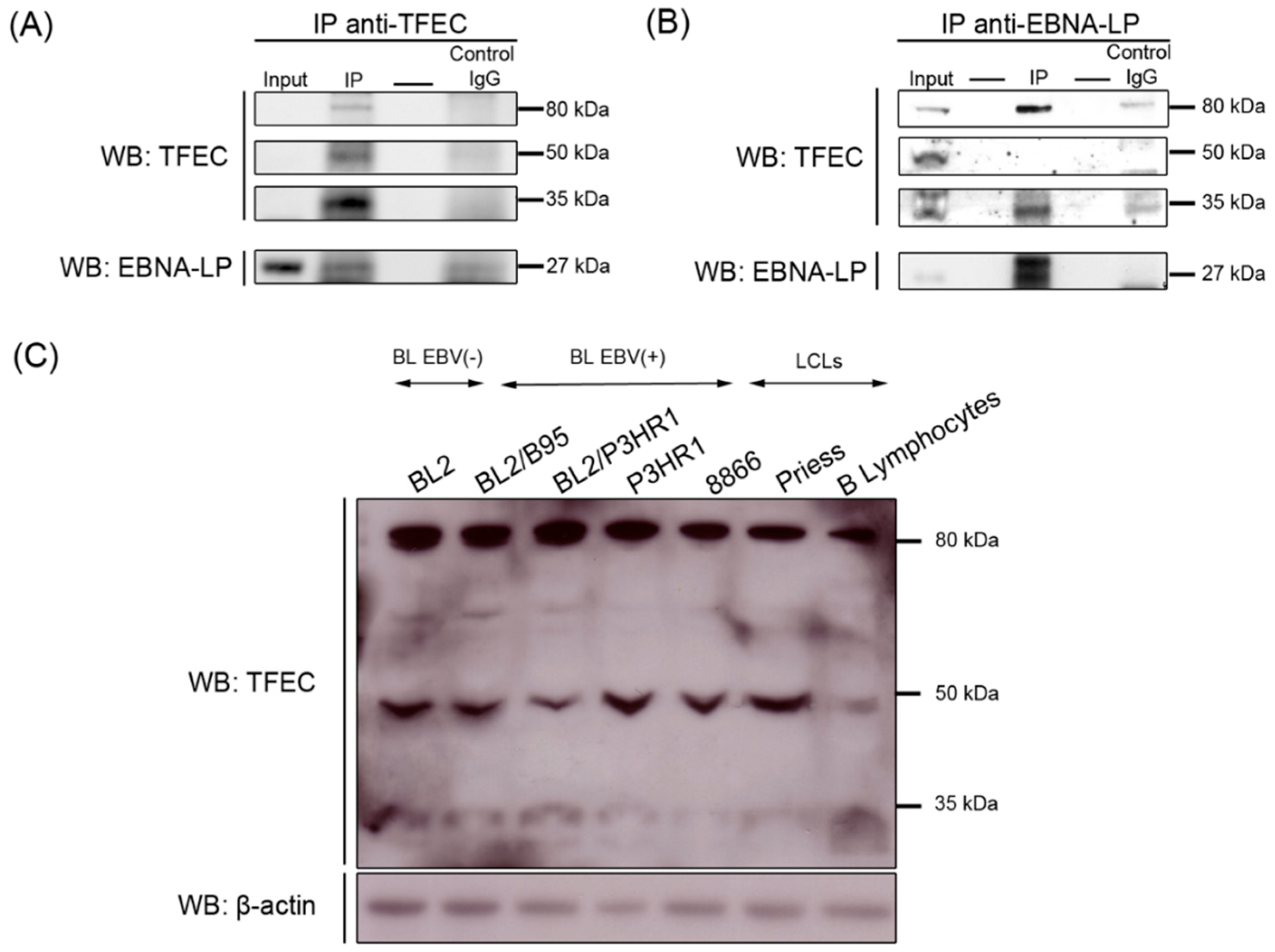
2.4.2. Truncated EBNA-LP Interacts with TFEC
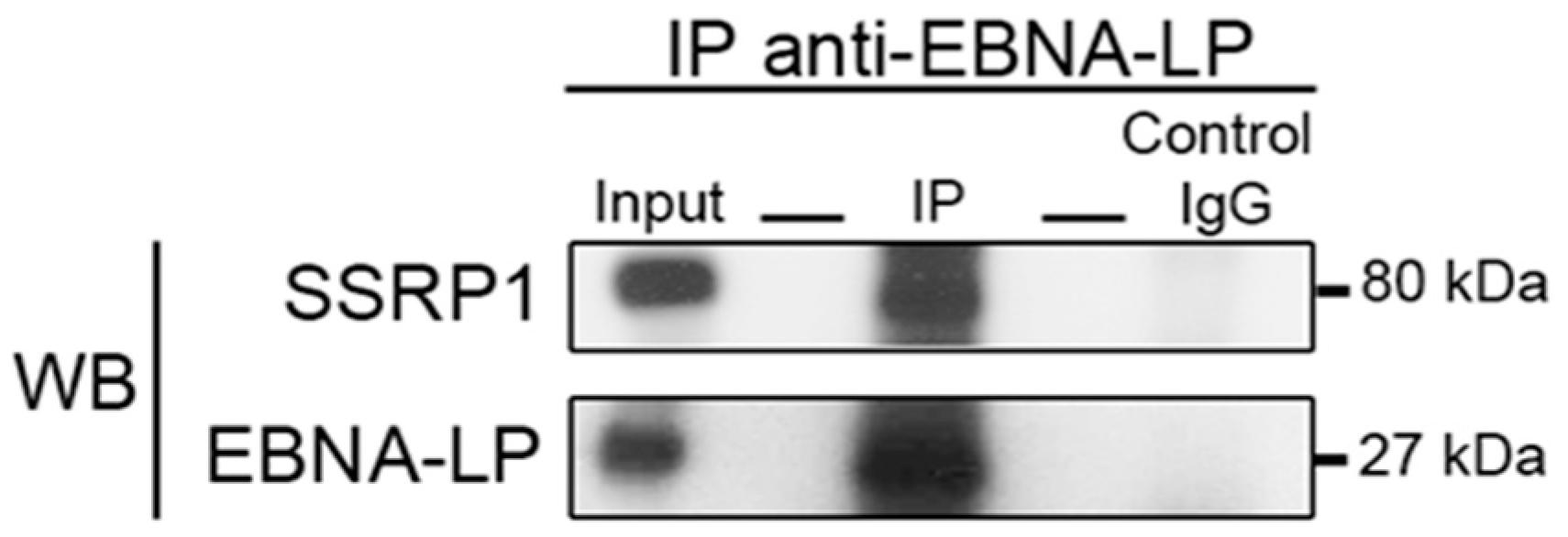
2.4.3. Truncated EBNA-LP Interacts with SSRP1
3. Materials and Methods
3.1. Cell Lines
3.2. Antibodies
3.3. Western Blot Analysis
3.4. Immunoprecipitation
3.5. Production and Purification of 6xHis-EBNA-LP-1RY1Y2
3.6. Identification of EBNA-LP Specific Peptides by Mass Spectrometry
3.7. Identification of tEBNA-LP Partners by Mass Spectrometry
3.8. IPA Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Epstein, M.A.; Achong, B.G.; Barr, Y.M. Virus particles in cultured lymphoblasts from Burkitt’s lymphoma. Lancet 1964, 1, 702–703. [Google Scholar] [CrossRef]
- Schaefer, B.C.; Strominger, J.L.; Speck, S.H. Redefining the Epstein-Barr virus-encoded nuclear antigen EBNA-1 gene promoter and transcription initiation site in group I Burkitt lymphoma cell lines. Proc. Natl. Acad. Sci. USA 1995, 92, 10565–10569. [Google Scholar] [CrossRef] [PubMed]
- Sample, J.; Brooks, L.; Sample, C.; Young, L.; Rowe, M.; Gregory, C.; Rickinson, A.; Kieff, E. Restricted Epstein-Barr virus protein expression in Burkitt lymphoma is due to a different Epstein-Barr nuclear antigen 1 transcriptional initiation site. Proc. Natl. Acad. Sci. USA 1991, 88, 6343–6347. [Google Scholar] [CrossRef] [PubMed]
- Kelly, G.; Bell, A.; Rickinson, A. Epstein-barr virus-associated Burkitt lymphomagenesis selects for downregulation of the nuclear antigen EBNA2. Nat. Med. 2002, 8, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.; Foster, L.; Sheedy, T.; Griffin, B.E. The EB virus genome in daudi Burkitt's lymphoma cells has a deletion similar to that observed in a non-transforming strain (P3HR-1) of the virus. EMBO J. 1984, 3, 813–821. [Google Scholar] [PubMed]
- Kelly, G.L.; Milner, A.E.; Tierney, R.J.; Croom-Carter, D.S.; Altmann, M.; Hammerschmidt, W.; Bell, A.I.; Rickinson, A.B. Epstein-barr virus nuclear antigen 2 (EBNA2) gene deletion is consistently linked with EBNA3a, -3b, and -3c expression in Burkitt's lymphoma cells and with increased resistance to apoptosis. J. Virol. 2005, 79, 10709–10717. [Google Scholar] [CrossRef] [PubMed]
- Kelly, G.L.; Long, H.M.; Stylianou, J.; Thomas, W.A.; Leese, A.; Bell, A.I.; Bornkamm, G.W.; Mautner, J.; Rickinson, A.B.; Rowe, M. An Epstein-Barr virus anti-apoptotic protein constitutively expressed in transformed cells and implicated in Burkitt lymphomagenesis: The wp/bhrf1 link. PLoS Pathog. 2009, 5, e1000341. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, A.; Maruo, S.; Ito, T.; Ito, M.; Katsumura, K.R.; Takada, K. Epstein-barr virus-encoded Bcl-2 homologue functions as a survival factor in Wp-restricted Burkitt lymphoma cell line P3HR-1. J. Virol. 2010, 84, 2893–2901. [Google Scholar] [CrossRef] [PubMed]
- Garibal, J.; Hollville, E.; Bell, A.I.; Kelly, G.L.; Renouf, B.; Kawaguchi, Y.; Rickinson, A.B.; Wiels, J. Truncated form of the Epstein-Barr virus protein EBNA-LP protects against caspase-dependent apoptosis by inhibiting protein phosphatase 2a. J. Virol. 2007, 81, 7598–7607. [Google Scholar] [CrossRef] [PubMed]
- Garibal, J.; Hollville, E.; Renouf, B.; Tetaud, C.; Wiels, J. Caspase-8-mediated cleavage of bid and protein phosphatase 2a-mediated activation of bax are necessary for verotoxin-1-induced apoptosis in Burkitt’s lymphoma cells. Cell Signal. 2010, 22, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Petti, L.; Braun, D.; Seung, S.; Kieff, E. A bicistronic Epstein-Barr virus mRNA encodes two nuclear proteins in latently infected, growth-transformed lymphocytes. J. Virol. 1987, 61, 945–954. [Google Scholar] [PubMed]
- Dillner, J.; Kallin, B.; Alexander, H.; Ernberg, I.; Uno, M.; Ono, Y.; Klein, G.; Lerner, R.A. An Epstein-Barr virus (EBV)-determined nuclear antigen (EBNA5) partly encoded by the transformation-associated Bam WYH region of EBV DNA: Preferential expression in lymphoblastoid cell lines. Proc. Natl. Acad. Sci. USA 1986, 83, 6641–6645. [Google Scholar] [CrossRef] [PubMed]
- Sample, J.; Hummel, M.; Braun, D.; Birkenbach, M.; Kieff, E. Nucleotide sequences of mRNAs encoding Epstein-Barr virus nuclear proteins: A probable transcriptional initiation site. Proc. Natl. Acad. Sci. USA 1986, 83, 5096–5100. [Google Scholar] [CrossRef] [PubMed]
- Speck, S.H.; Pfitzner, A.; Strominger, J.L. An Epstein-Barr virus transcript from a latently infected, growth-transformed B-cell line encodes a highly repetitive polypeptide. Proc. Natl. Acad. Sci. USA 1986, 83, 9298–9302. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, C.; Birkenbach, M.; Kieff, E. Early events in Epstein-Barr virus infection of human B lymphocytes. Virology 1991, 181, 595–608. [Google Scholar] [CrossRef]
- Bodescot, M.; Perricaudet, M.; Farrell, P.J. A promoter for the highly spliced EBNA family of RNAs of Epstein-Barr virus. J. Virol. 1987, 61, 3424–3430. [Google Scholar] [PubMed]
- Woisetschlaeger, M.; Yandava, C.N.; Furmanski, L.A.; Strominger, J.L.; Speck, S.H. Promoter switching in Epstein-Barr virus during the initial stages of infection of B lymphocytes. Proc. Natl. Acad. Sci. USA 1990, 87, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Kieff, E. Epstein-barr virus nuclear protein LP stimulates EBNA-2 acidic domain-mediated transcriptional activation. J. Virol. 1997, 71, 6611–6618. [Google Scholar] [PubMed]
- Nitsche, F.; Bell, A.; Rickinson, A. Epstein-barr virus leader protein enhances EBNA-2-mediated transactivation of latent membrane protein 1 expression: A role for the W1W2 repeat domain. J. Virol. 1997, 71, 6619–6628. [Google Scholar] [PubMed]
- Khanna, A.; Pimanda, J.E.; Westermarck, J. Cancerous inhibitor of protein phosphatase 2A, an emerging human oncoprotein and a potential cancer therapy target. Cancer Res. 2013, 73, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Mannick, J.B.; Cohen, J.I.; Birkenbach, M.; Marchini, A.; Kieff, E. The Epstein-Barr virus nuclear protein encoded by the leader of the EBNA RNAs is important in B-lymphocyte transformation. J. Virol. 1991, 65, 6826–6837. [Google Scholar] [PubMed]
- Ling, P.D. Epstein-Barr virus leader protein. In Epstein-barr Virus Latency and Transformation; Robertson, E.S., Ed.; Caister Academic Press: Poole, UK, 2010; pp. 25–36. [Google Scholar]
- Kawaguchi, Y.; Nakajima, K.; Igarashi, M.; Morita, T.; Tanaka, M.; Suzuki, M.; Yokoyama, A.; Matsuda, G.; Kato, K.; Kanamori, M.; et al. Interaction of Epstein-Barr virus nuclear antigen leader protein (EBNA-LP) with HS1-associated protein X-1: Implication of cytoplasmic function of EBNA-LP. J. Virol. 2000, 74, 10104–10111. [Google Scholar] [CrossRef] [PubMed]
- Han, I.; Harada, S.; Weaver, D.; Xue, Y.; Lane, W.; Orstavik, S.; Skalhegg, B.; Kieff, E. EBNA-LP associates with cellular proteins including DNA-Pk and HA95. J. Virol. 2001, 75, 2475–2481. [Google Scholar] [CrossRef] [PubMed]
- Mannick, J.B.; Tong, X.; Hemnes, A.; Kieff, E. The Epstein-Barr virus nuclear antigen leader protein associates with hsp72/hsc73. J. Virol. 1995, 69, 8169–8172. [Google Scholar] [PubMed]
- Kitay, M.K.; Rowe, D.T. Protein-protein interactions between Epstein-Barr virus nuclear antigen-LP and cellular gene products: Binding of 70-kilodalton heat shock proteins. Virology 1996, 220, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.W.; Zhao, B.; Chen, H.C.; Chou, M.L.; Lai, C.Y.; Lin, S.Z.; Hsu, H.Y.; Kieff, E. Hsp72 up-regulates Epstein-Barr virus EBNALP coactivation with EBNA2. Blood 2007, 109, 5447–5454. [Google Scholar] [CrossRef] [PubMed]
- Forsman, A.; Ruetschi, U.; Ekholm, J.; Rymo, L. Identification of intracellular proteins associated with the EBV-encoded nuclear antigen 5 using an efficient TAP procedure and FT-ICR mass spectrometry. J. Proteome Res. 2008, 7, 2309–2319. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Tan, J.; Ling, P.D. Conserved regions in the Epstein-Barr virus leader protein define distinct domains required for nuclear localization and transcriptional cooperation with EBNA2. J. Virol. 2000, 74, 9953–9963. [Google Scholar] [CrossRef] [PubMed]
- Ling, P.D.; Tan, J.; Peng, R. Nuclear-cytoplasmic shuttling is not required for the Epstein-Barr virus EBNA-LP transcriptional coactivation function. J. Virol. 2009, 83, 7109–7116. [Google Scholar] [CrossRef] [PubMed]
- Rabouille, C.; Malhotra, V.; Nickel, W. Diversity in unconventional protein secretion. J. Cell Sci. 2012, 125, 5251–5255. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Yokoyama, A.; Tohya, Y.; Akashi, H.; Nishiyama, Y.; Kawaguchi, Y. Identification of protein kinases responsible for phosphorylation of Epstein-Barr virus nuclear antigen leader protein at serine-35, which regulates its coactivator function. J. Gen. Virol. 2003, 84, 3381–3392. [Google Scholar] [CrossRef] [PubMed]
- Burgess, A.; Buck, M.; Krauer, K.; Sculley, T. Nuclear localization of the Epstein-Barr virus EBNA3b protein. J. Gen. Virol. 2006, 87, 789–793. [Google Scholar] [CrossRef] [PubMed]
- White, R.E.; Groves, I.J.; Turro, E.; Yee, J.; Kremmer, E.; Allday, M.J. Extensive co-operation between the Epstein-Barr virus EBNA3 proteins in the manipulation of host gene expression and epigenetic chromatin modification. PLoS ONE 2010, 5, e13979. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, C.; Haesen, D.; Sents, W.; Ivanova, E.; Janssens, V. Structure, regulation, and pharmacological modulation of PP2A phosphatases. Methods Mol. Biol. 2013, 1053, 283–305. [Google Scholar]
- Tamura, Y.; Simizu, S.; Osada, H. The phosphorylation status and anti-apoptotic activity of Bcl-2 are regulated by ERK and protein phosphatase 2A on the mitochondria. FEBS Lett. 2004, 569, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Deng, X. Protein phosphatase 2A enhances the proapoptotic function of Bax through dephosphorylation. J. Biol. Chem. 2006, 281, 18859–18867. [Google Scholar] [CrossRef] [PubMed]
- Junttila, M.R.; Puustinen, P.; Niemela, M.; Ahola, R.; Arnold, H.; Bottzauw, T.; Ala-aho, R.; Nielsen, C.; Ivaska, J.; Taya, Y.; et al. CIP2A inhibits PP2A in human malignancies. Cell 2007, 130, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Perrotti, D.; Neviani, P. Protein phosphatase 2A: A target for anticancer therapy. Lancet Oncol. 2013, 14, e229–e238. [Google Scholar] [CrossRef]
- Pallas, D.C.; Shahrik, L.K.; Martin, B.L.; Jaspers, S.; Miller, T.B.; Brautigan, D.L.; Roberts, T.M. Polyoma small and middle T antigens and SV40 small t antigen form stable complexes with protein phosphatase 2A. Cell 1990, 60, 167–176. [Google Scholar] [CrossRef]
- Shtrichman, R.; Sharf, R.; Barr, H.; Dobner, T.; Kleinberger, T. Induction of apoptosis by adenovirus E4orf4 protein is specific to transformed cells and requires an interaction with protein phosphatase 2A. Proc. Natl. Acad. Sci. USA 1999, 96, 10080–10085. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, Y.; Bao, Q.; Xing, Y.; Li, Z.; Lin, Z.; Stock, J.B.; Jeffrey, P.D.; Shi, Y. Structural and biochemical insights into the regulation of protein phosphatase 2A by small t antigen of SV40. Nat. Struct. Mol. Biol. 2007, 14, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Cho, U.S.; Morrone, S.; Sablina, A.A.; Arroyo, J.D.; Hahn, W.C.; Xu, W. Structural basis of PP2A inhibition by small t antigen. PLoS Biol. 2007, 5, e202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemesath, T.J.; Steingrimsson, E.; McGill, G.; Hansen, M.J.; Vaught, J.; Hodgkinson, C.A.; Arnheiter, H.; Copeland, N.G.; Jenkins, N.A.; Fisher, D.E. Microphthalmia, a critical factor in melanocyte development, defines a discrete transcription factor family. Genes Dev. 1994, 8, 2770–2780. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, K.; Shibahara, S. Molecular cloning of cDNA encoding a human TFEC isoform, a newly identified transcriptional regulator. Biochim. Biophys. Acta 1997, 1353, 23–31. [Google Scholar] [CrossRef]
- Chung, M.C.; Kim, H.K.; Kawamoto, S. TFEC can function as a transcriptional activator of the nonmuscle myosin II heavy chain-A gene in transfected cells. Biochemistry 2001, 40, 8887–8897. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Levy, C.; Davis, I.J.; Razin, E.; Fisher, D.E. Sumoylation of MITF and its related family members TFE3 and TFEB. J. Biol. Chem. 2005, 280, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Murakami, H.; Arnheiter, H. Sumoylation modulates transcriptional activity of MITF in a promoter-specific manner. Pigment Cell Melanoma Res. 2005, 18, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Bertolotto, C.; Lesueur, F.; Giuliano, S.; Strub, T.; de Lichy, M.; Bille, K.; Dessen, P.; d’Hayer, B.; Mohamdi, H.; Remenieras, A.; et al. A sumoylation-defective MITF germline mutation predisposes to melanoma and renal carcinoma. Nature 2011, 480, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xie, Y.; Zheng, Y.; Jiang, S.; Liu, W.; Mu, W.; Zhao, Y.; Xue, Y.; Ren, Y. GPS-SUMO: A tool for the prediction of sumoylation sites and SUMO-interaction motifs. Nucleic Acids Res. 2014, 42, W325–W330. [Google Scholar] [CrossRef] [PubMed]
- Orphanides, G.; Wu, W.H.; Lane, W.S.; Hampsey, M.; Reinberg, D. The chromatin-specific transcription elongation factor FACT comprises human SPT16 and SSRP1 proteins. Nature 1999, 400, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Chu, G. Cellular responses to cisplatin. The roles of DNA-binding proteins and DNA repair. J. Biol. Chem. 1994, 269, 787–790. [Google Scholar] [PubMed]
- Yarnell, A.T.; Oh, S.; Reinberg, D.; Lippard, S.J. Interaction of FACT, SSRP1, and the high mobility group (HMG) domain of SSRP1 with DNA damaged by the anticancer drug cisplatin. J. Biol. Chem. 2001, 276, 25736–25741. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Liu, E.; Renne, R. Involvement of SSRP1 in latent replication of Kaposi's sarcoma-associated Herpesvirus. J. Virol. 2009, 83, 11051–11063. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, S.X.; Landais, I.; Lu, H. Human SSRP1 has spt16-dependent and -independent roles in gene transcription. J. Biol. Chem. 2007, 282, 6936–6945. [Google Scholar] [CrossRef] [PubMed]
- Fox, H.L.; Dembowski, J.A.; DeLuca, N.A. A Herpesviral immediate early protein promotes transcription elongation of viral transcripts. mBio 2017, 8, e00745-17. [Google Scholar] [CrossRef] [PubMed]
- Kempkes, B.; Ling, P.D. EBNA2 and its coactivator EBNA-LP. Curr. Top. Microbiol. Immunol. 2015, 391, 35–59. [Google Scholar] [PubMed]
- Shaku, F.; Matsuda, G.; Furuya, R.; Kamagata, C.; Igarashi, M.; Tanaka, M.; Kanamori, M.; Nishiyama, Y.; Yamamoto, N.; Kawaguchi, Y. Development of a monoclonal antibody against Epstein-Barr virus nuclear antigen leader protein (EBNA-LP) that can detect EBNA-LP expressed in P3HR1 cells. Microbiol. Immunol. 2005, 49, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.J.; Palmero, I.; Peters, G.; Farrell, P.J. EBNA-2 and EBNA-LP cooperate to cause G0 to G1 transition during immortalization of resting human B lymphocytes by Epstein-Barr virus. EMBO J. 1994, 13, 3321–3328. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Proteins | Symbols | ID | Distinct Peptides | MS/MS Best Search Score (Identification Search) |
| Transcription factor EC | TFEC | O14948 | 1 | 12.50 |
| FACT complex subunit SSRP1 | SSRP1 | Q08945 | 6 | 89.72 |
| Serine/threonine-protein phosphatase 2A | PP2A A | P30153 | 3 | * |
| Tyrosine-protein phosphatase non-receptor type 13 | PTPN13/FAP1 | Q12923 | 2 | 23.39 |
| Calcium-calmodulin independent nitric oxide synthase | iNOS | P35228 | 1 | 9.26 |
| Cytoplasmic linker associated protein 1 | CLASP1 | Q7Z460 | 4 | 25.50 |
| Fibronectin | FN1 | P02751 | 14 | 165.44 |
| Profilin-1 | PFN1 | P07737 | 10 | 137.87 |
| α-2-macroglobulin | A2M | P01023 | 1 | 9.12 |
| L-lactate dehydrogenase A chain | LDHA | P00338 | 19 | 301.06 |
| Ceruloplasmin precursor | CP | P00450 | 2 | 33.22 |
| Oxysterol-binding protein-related protein 3 | OSBPL3 | Q9H4L5 | 1 | 11.06 |
| RING finger protein unkempt homolog | UNK | Q9C0B0 | 2 | 15.79 |
| H/ACA ribonucleoprotein complex subunit 4 | DKC1 | O60832 | 1 | 7.55 |
| Haptoglobin precursor | HP | P00738 | 1 | 13.57 |
| 10 kDa heat shock protein, mitochondrial | HSPE1 | P61604 | 6 | 96.96 |
| Fibrinogen gamma chain precursor | FGG | P02679 | 4 | 53.50 |
| Fibrinogen beta chain precursor | FGB | P02675 | 10 | 15.81 |
| Putative heat shock protein HSP 90-β-3 | HSP90 | Q58FF7 | 19 | 202.04 |
| Ezrin | EZR | P15311 | 24 | 199.31 |
| L-lactate dehydrogenase B chain | LDHB | P07195 | 18 | 285.01 |
| Exosome complex exonuclease RRP44 | DIS3 | Q9Y2L1 | 14 | 160.93 |
| Glycogen debranching enzyme | AGL | P35573 | 9 | 112.33 |
| Pre-mRNA branch site protein p14 | SF3B14 | Q9Y3B4 | 2 | 41.10 |
| Tight junction protein ZO-2 | TJP2 | Q9UDY2 | 3 | 14.13 |
| Histone-lysine N-methyltransferase | MLL | Q03164 | 2 | 17.19 |
| GTPase-activating Rap/Ran-GAP domain-like protein 3 | GARNL3 | Q5VVW2 | 1 | 14.75 |
| Laminin subunit α-4 | LAMA4 | Q16363 | 1 | 12.71 |
| Viral Proteins | Symbol | ID | Distinct Peptides | Best Score |
| Epstein-Barr nuclear antigen leader protein | EBNA-LP | Q1HVI8 | 2 | 42.68 |
| Portal protein | BBRF1 | Q1HVF2 | 2 | 7.74 |
| Ribonucleoside-diphosphate reductase large subunit | BORF2 | P0C702 | 4 | 13.19 |
| Immediate-early phosphoprotein Serine/threonine-protein kinase | BGLF4 | P0C6Z8 | 1 | 7.68 |
| Epstein-Barr nuclear antigen 4 | EBNA-3B | Q1HVG4 | 1 | 49.95 |
| Cellular Functions | Molecules | Numbers of Molecules |
|---|---|---|
| Cell Death | EZR, FN1, iNOS, PP2A A, PTPN13/FAP1, LAMA4, HSPE1, SF3B14, A2M, SSRP1, LDHA, MLL. | 12 |
| Cellular Growth and Proliferation | FN1, LDHA, iNOS, PP2A A, PTPN13/FAP1, PFN1, TJP2, LAMA4, EZR, DKC1, A2M, MLL. | 12 |
| Cellular Development | EZR, FN1, PFN1, PTPN13/FAP1, PP2A A, LAMA4, iNOS, A2M, LDHA, MLL. | 10 |
| Cell Signaling | FGB, FGG, FN1, iNOS, HP, PP2A A, PFN1, A2M, MLL. | 9 |
| Cellular Movement | EZR, FN1, iNOS, HP, PFN1, FGB, SSRP1, A2M. | 8 |
| Cell-To-Cell Signaling and Interaction | FGG, FN1, TJP2, CP, iNOS, A2M. | 6 |
| Gene Expression | SSRP1, TFEC, FN1, iNOS, A2M. | 5 |
| Post-Translational Modification | HSPE1, PP2A A, PTPN13/FAP1, CP, MLL. | 5 |
| Cell Morphology | LAMA4, FN1, PFN1, EZR, A2M. | 5 |
| Amino Acid Metabolism | CP, PP2A A, PTPN13/FAP1, iNOS. | 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chelouah, S.; Cochet, E.; Couvé, S.; Balkaran, S.; Robert, A.; May, E.; Ogryzko, V.; Wiels, J. New Interactors of the Truncated EBNA-LP Protein Identified by Mass Spectrometry in P3HR1 Burkitt’s Lymphoma Cells. Cancers 2018, 10, 12. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10010012
Chelouah S, Cochet E, Couvé S, Balkaran S, Robert A, May E, Ogryzko V, Wiels J. New Interactors of the Truncated EBNA-LP Protein Identified by Mass Spectrometry in P3HR1 Burkitt’s Lymphoma Cells. Cancers. 2018; 10(1):12. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10010012
Chicago/Turabian StyleChelouah, Sonia, Emilie Cochet, Sophie Couvé, Sandy Balkaran, Aude Robert, Evelyne May, Vasily Ogryzko, and Joëlle Wiels. 2018. "New Interactors of the Truncated EBNA-LP Protein Identified by Mass Spectrometry in P3HR1 Burkitt’s Lymphoma Cells" Cancers 10, no. 1: 12. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers10010012