The Opposing Contribution of SMS1 and SMS2 to Glioma Progression and Their Value in the Therapeutic Response to 2OHOA

 ,
, 
Abstract
:1. Introduction
2. Results
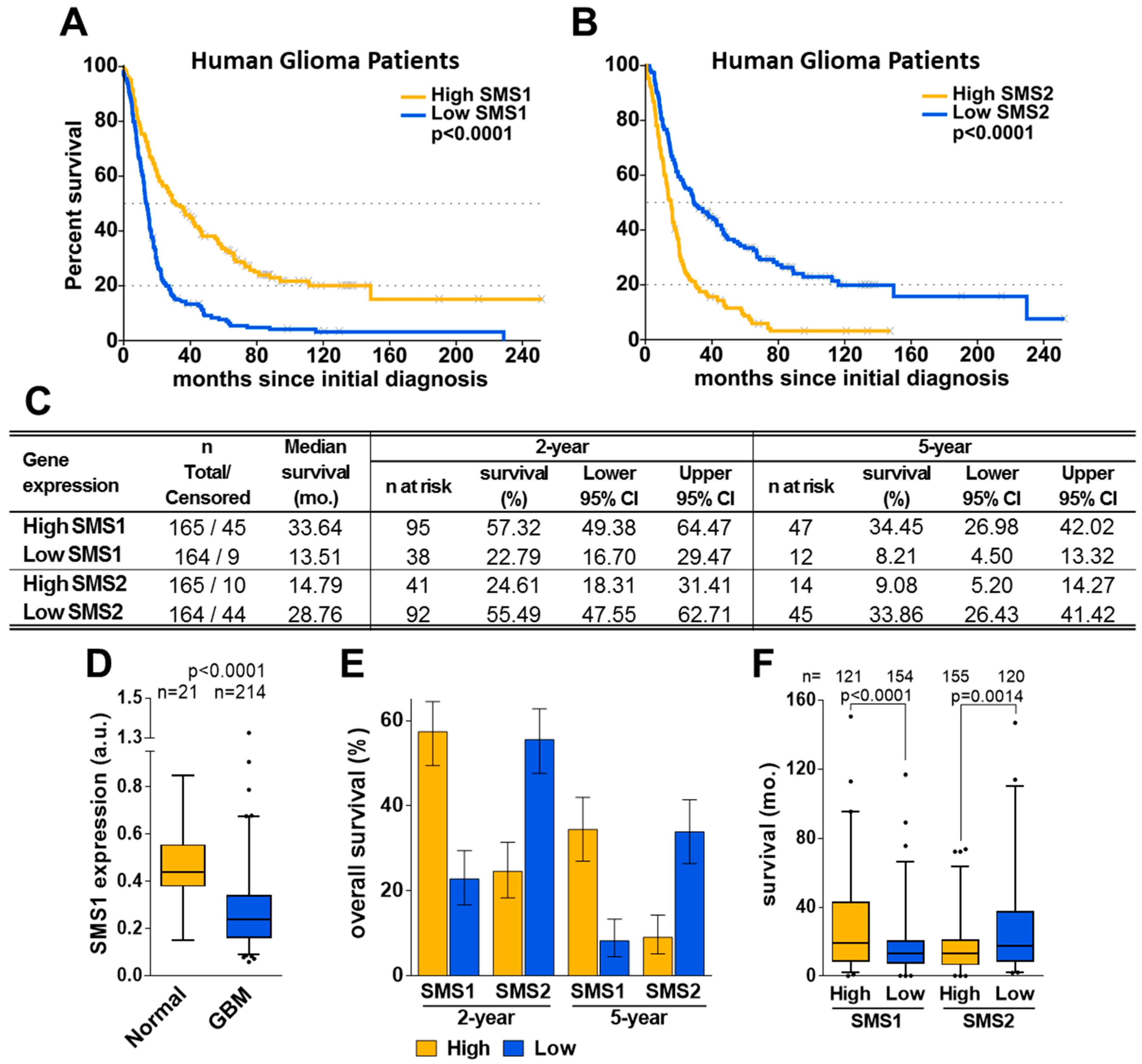
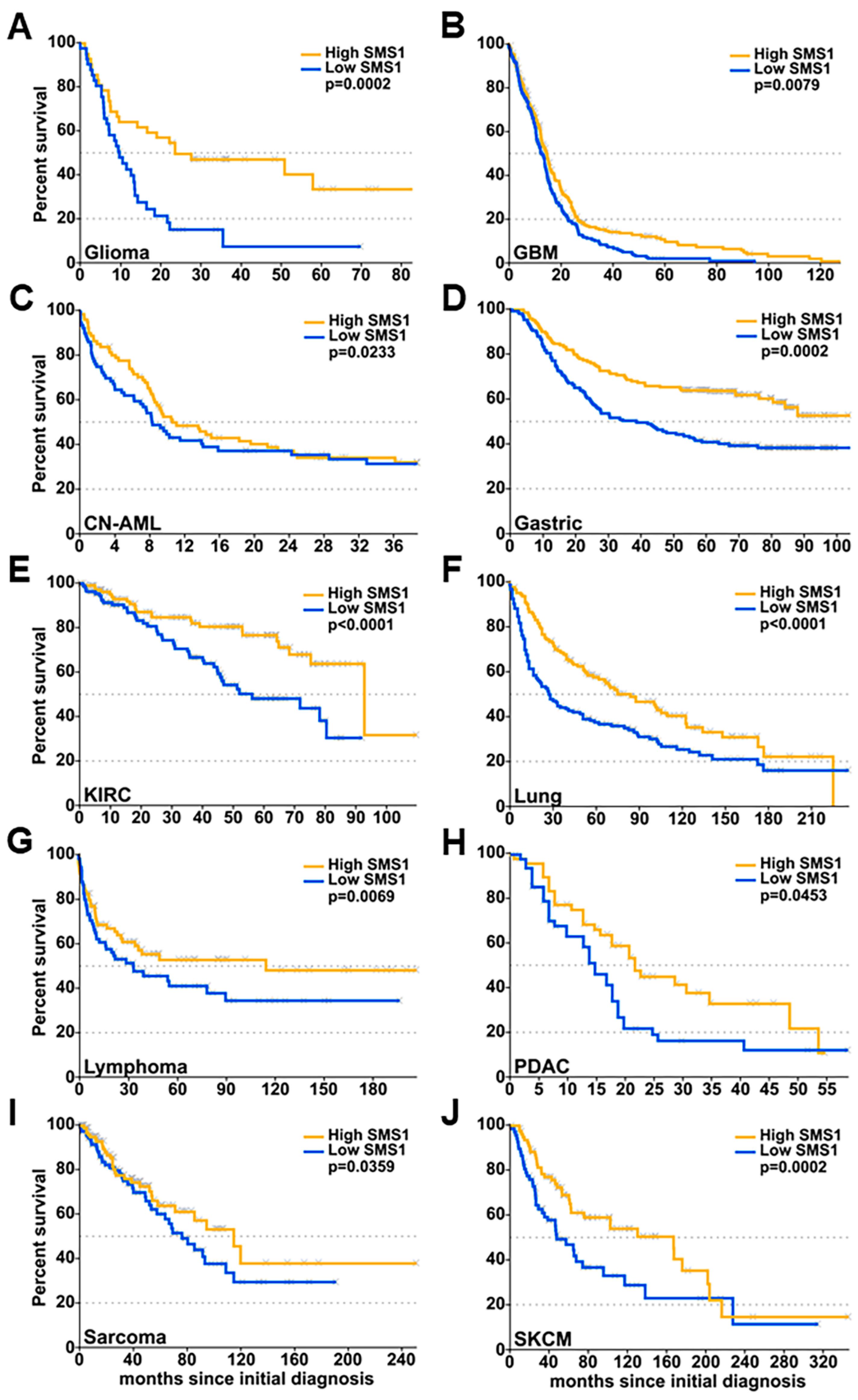
2.1. SMS1 and SMS2 Expression Have Opposite Effects on Glioma Patient Survival
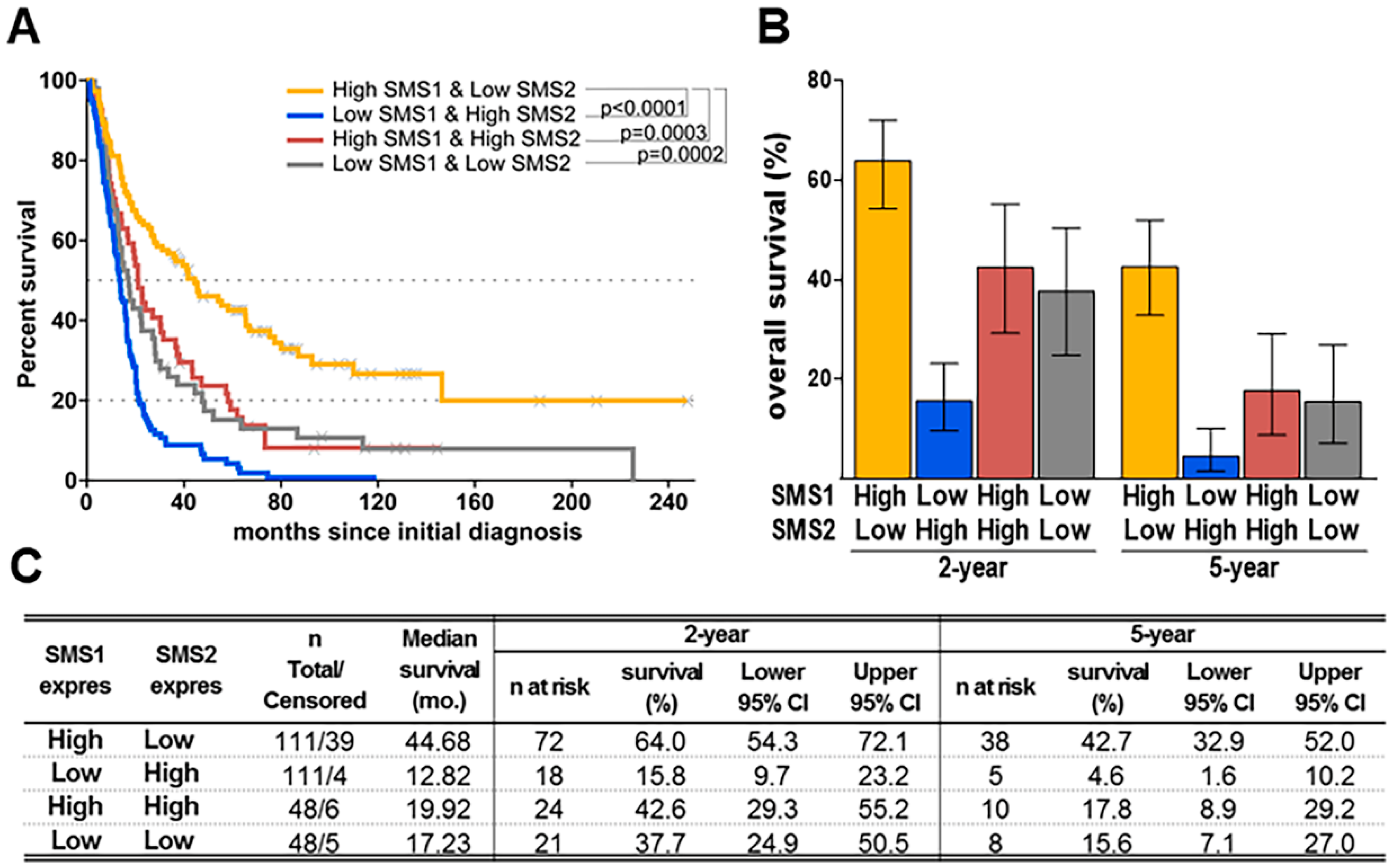
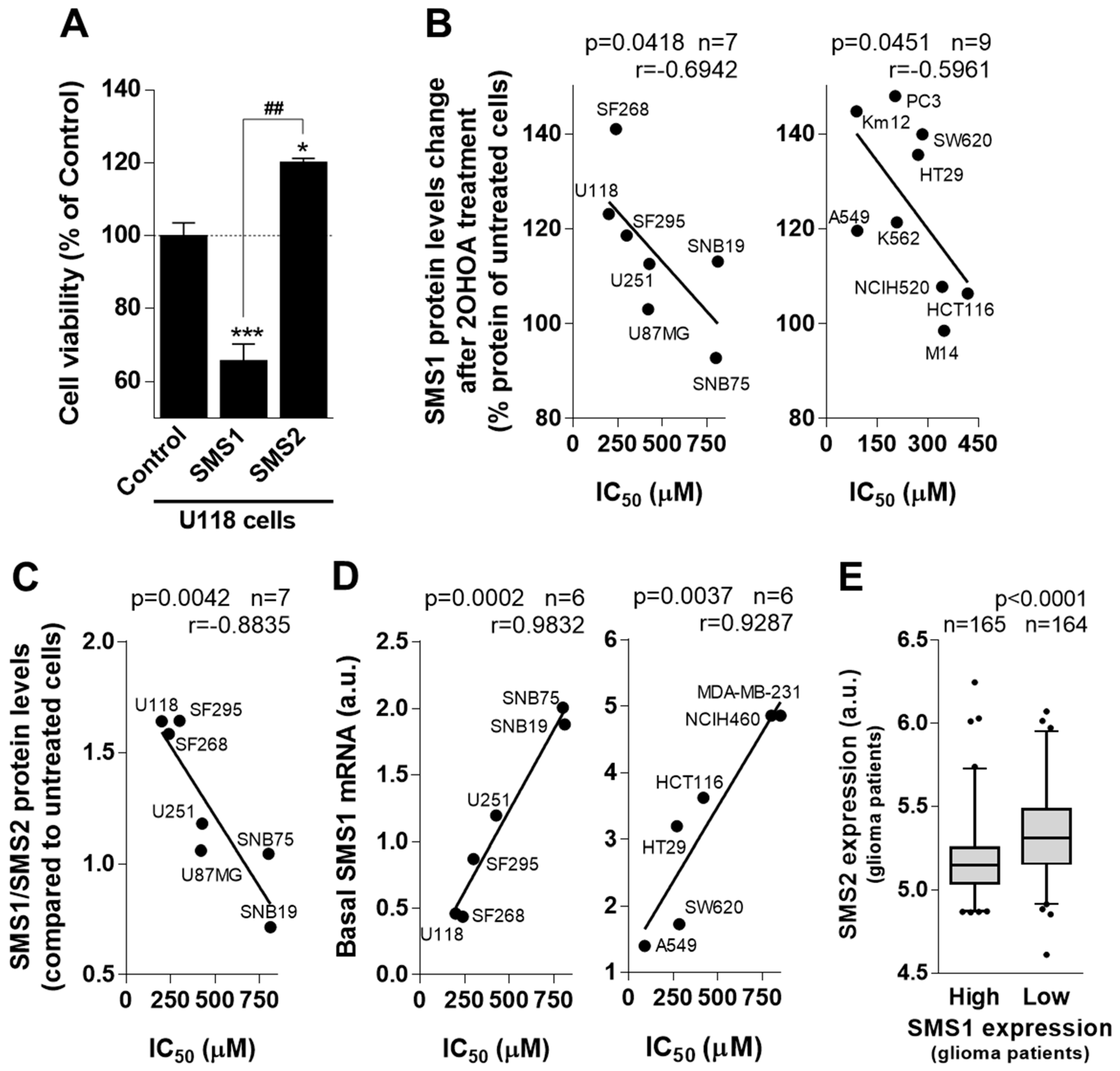
2.2. SMS1/SMS2 Ratio is Relevant for Glioma Patient Survival and Correlates with 2OHOA’s Pharmacological Efficacy
2.3. Basal SMS1 mRNA Levels Anticipate 2OHOA Anti-Cancer Activity against GBM
2.4. Opposing Influence of SMS1 and SMS2 on 2OHOA Regulation of GBM Cell Proliferation and Differentiation
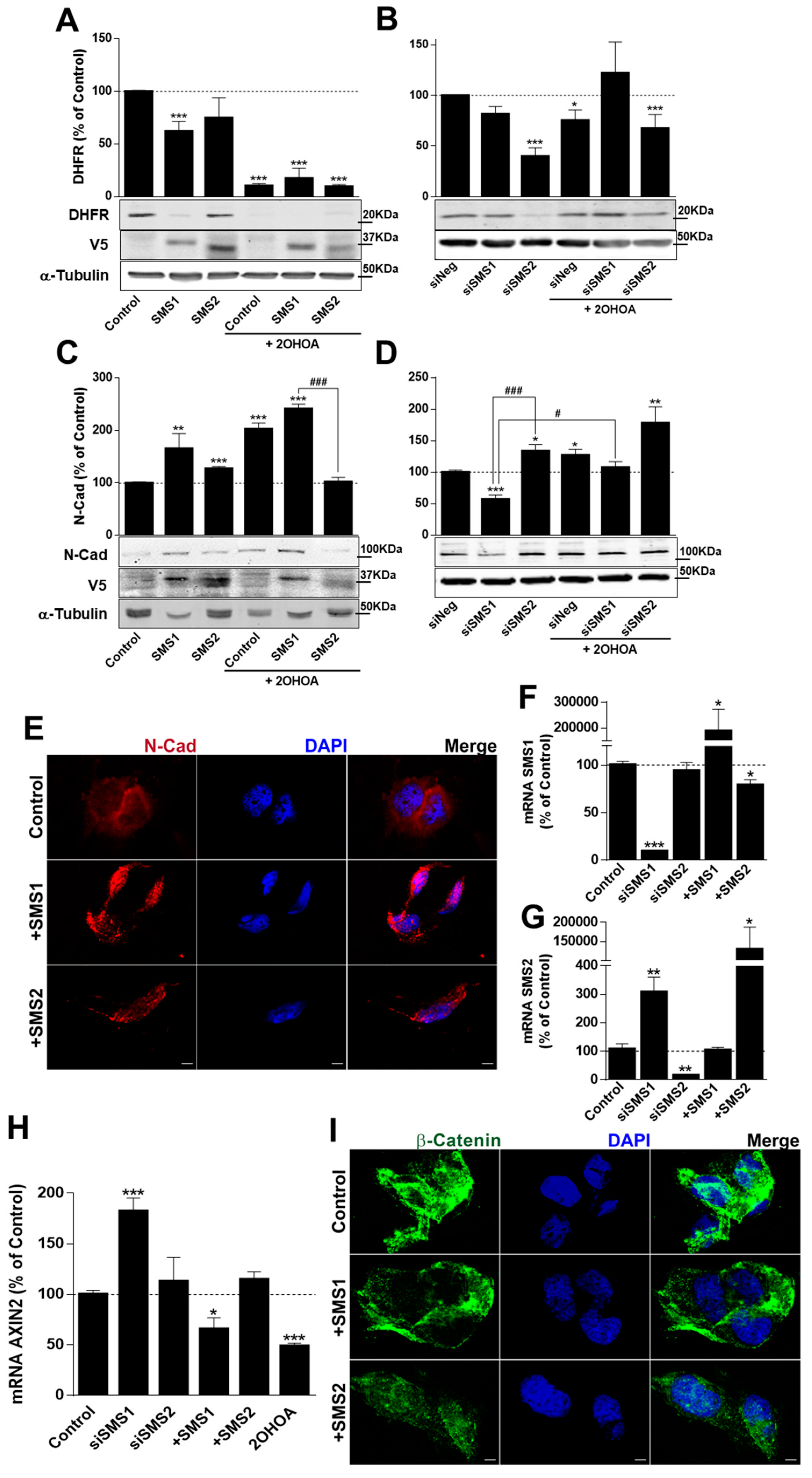
2.5. SMS1 and 2OHOA Modulate the β-Catenin Pathway
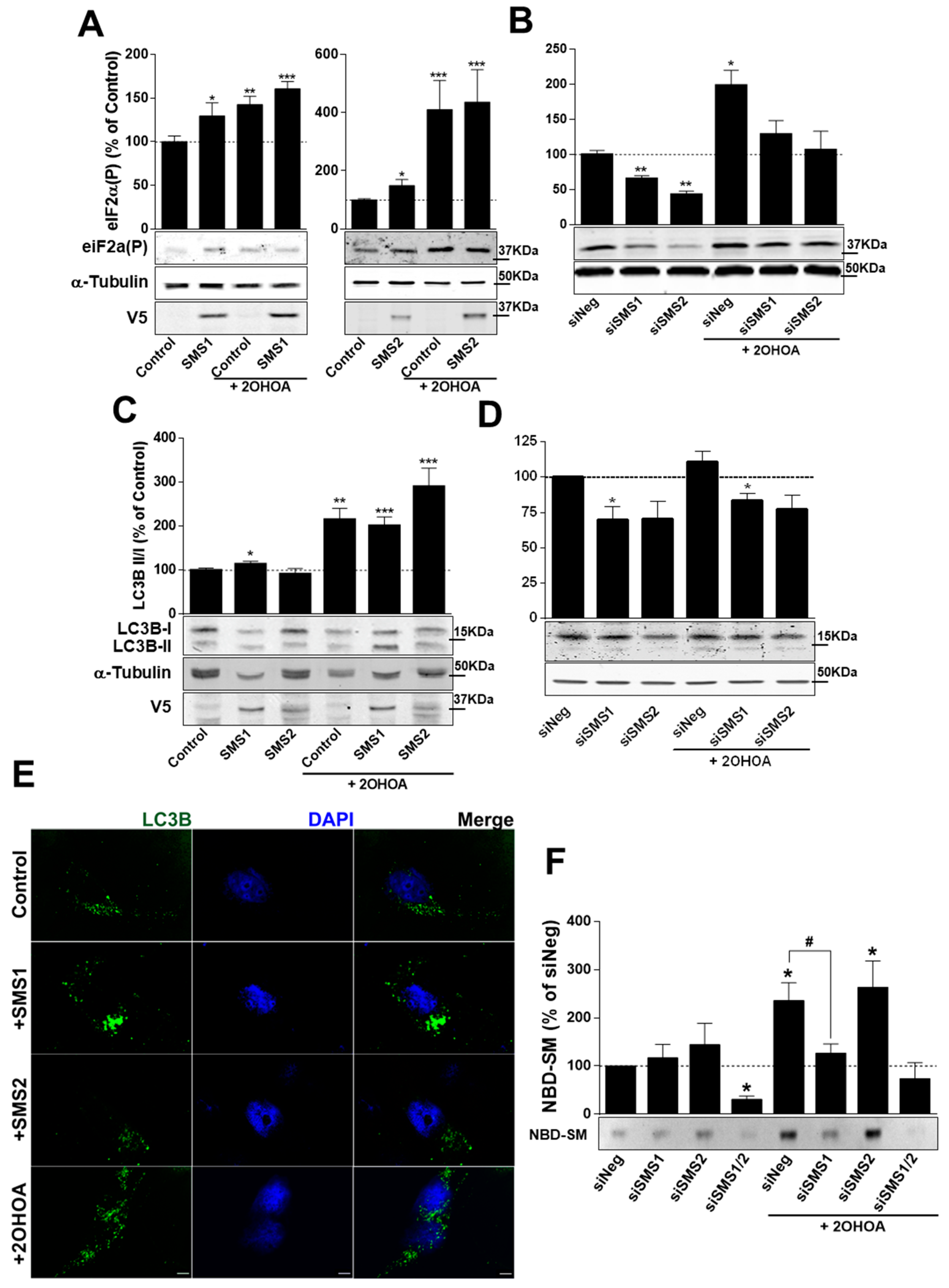
2.6. SMS1 and SMS2 Modulate ER Stress but Only SMS1 Affects Autophagy
2.7. 2OHOA Increases SMS Activity through SMS1 Exclusively
3. Discussion
4. Materials and Methods
4.1. Cancer Patient Data Collection and Processing
4.2. Cell Culture
4.3. Cell Transfection and Protein Overexpression
4.4. Gene Expression Silencing
4.5. Antibody Generation
4.6. Cell Lysis, Electrophoresis, Immunoblotting and Protein Quantification
4.7. Immunofluorescence and Confocal Microscopy
4.8. RNA Isolation
4.9. Quantitative RT-PCR
- SMS1 Forward (Fw): TGACTCCAGTGCAACGTGAC
- SMS1 Reverse (Rv): GTCCACACTCCTTCAGTCGC
- SMS2 Fw: TTAATCTGCTGGCTGCTGAG
- SMS2 Rv: ACCAATCTTCTGAACCCGTG
- AXIN2 Fw: GAGTGGACTTGTGCCGACTTCA
- AXIN2 Rv: GGTGGCGTTGCAAAGACATAG
- HPRT Fw: TGACCTTGATTTATTTTGCATACC
- HPRT Rv: CGAGCAAGACGTTCAGTCCT
4.10. Enzymatic Assay
4.11. Thin Layer Chromatography (TLC)
4.12. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Louis, D.N. Molecular pathology of malignant gliomas. Ann. Rev. Pathol. 2006, 1, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Terés, S.; Lladé, V.; Higuera, M.; Barceló-Coblijn, G.; Martin, M.L.; Noguera-Salva, M.A.; Marcilla-Etxenike, A.; García-Verdugo, J.M.; Soriano-Navarro, M.; Saus, C.; et al. 2-Hydroxyoleate, a nontoxic membrane binding anticancer drug, induces glioma cell differentiation and autophagy. Proc. Natl. Acad. Sci. USA 2012, 109, 8489–8494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarguren, M.; López, D.J.; Encinar, J.A.; González-Ros, J.M.; Busquets, X.; Escribá, P.V. Partitioning of liquid-ordered/liquid-disordered membrane microdomains induced by the fluidifying effect of 2-hydroxylated fatty acid derivatives. Biochim. Biophys. Acta 2013, 1828, 2553–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarguren, M.; López, D.J.; Escribá, P.V. The effect of natural and synthetic fatty acids on membrane structure, microdomain organization, cellular functions and human health. Biochim. Biophys. Acta 2014, 1838, 1518–1528. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.L.; Liebisch, G.; Lehneis, S.; Schmitz, G.; Alonso-Sande, M.; Bestard-Escalas, J.; Lopez, D.H.; Garcia-Verdugo, J.M.; Soriano-Navarro, M.; Busquets, X.; et al. Sustained activation of sphingomyelin synthase by 2-hydroxyoleic acid induces sphingolipidosis in tumor cells. J. Lipid Res. 2013, 54, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrera, F.N.; Poveda, J.A.; González-Ros, J.M.; Neira, J.L. Binding of the C-terminal sterile alpha motif (SAM) domain of human p73 to lipid membranes. J. Biol. Chem. 2003, 278, 46878–46885. [Google Scholar] [CrossRef]
- Huitema, K.; van den Dikkenberg, J.; Brouwers, J.F.; Holthuis, J.C. Identification of a family of animal sphingomyelin synthases. EMBO J. 2004, 23, 33–44. [Google Scholar] [CrossRef]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef]
- Tafesse, F.G.; Huitema, K.; Hermansson, M.; van der Poel, S.; van den Dikkenberg, J.; Uphoff, A.; Somerharju, P.; Holthuis, J.C. Both sphingomyelin synthases SMS1 and SMS2 are required for sphingomyelin homeostasis and growth in human HeLa cells. J. Biol. Chem. 2007, 282, 17537–17547. [Google Scholar] [CrossRef]
- Diringer, H.; Marggraf, W.D.; Koch, M.A.; Anderer, F.A. Evidence for a new biosynthetic pathway of sphingomyelin in SV 40 transformed mouse cells. Biochem. Biophys. Res. Commun. 1972, 47, 1345–1352. [Google Scholar] [CrossRef]
- Ullman, M.D.; Radin, N.S. The enzymatic formation of sphingomyelin from ceramide and lecithin in mouse liver. J. Biol. Chem. 1974, 249, 1506–1512. [Google Scholar] [PubMed]
- Thomas, R.L., Jr.; Matsko, C.M.; Lotze, M.T.; Amoscato, A.A. Mass spectrometric identification of increased C16 ceramide levels during apoptosis. J. Biol. Chem. 1999, 274, 30580–30588. [Google Scholar] [CrossRef] [PubMed]
- Hampton, R.Y.; Morand, O.H. Sphingomyelin synthase and PKC activation. Science 1989, 246, 1050. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Toomre, D. Lipid rafts and signal transduction. Nat. Rev. Mol. Cell Biol. 2000, 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Holthuis, J.C.; Pomorski, T.; Raggers, R.J.; Sprong, H.; van Meer, G. The organizing potential of sphingolipids in intracellular membrane transport. Physiol. Rev. 2001, 81, 1689–1723. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, S.; Zenklusen, J.C.; Kotliarov, Y.; Sahni, H.; Fine, H.A.; Buetow, K. Rembrandt: Helping personalized medicine become a reality through integrative translational research. Mol. Cancer Res. MCR 2009, 7, 157–167. [Google Scholar] [CrossRef]
- Lladó, V.; López, D.J.; Ibarguren, M.; Alonso, M.; Soriano, J.B.; Escribá, P.V.; Busquets, X. Regulation of the cancer cell membrane lipid composition by NaCHOleate: Effects on cell signaling and therapeutical relevance in glioma. Biochim. Biophys. Acta 2014, 1838, 1619–1627. [Google Scholar] [CrossRef] [PubMed]
- Barceló-Coblijn, G.; Martin, M.L.; de Almeida, R.F.; Noguera-Salva, M.A.; Marcilla-Etxenike, A.; Guardiola-Serrano, F.; Luth, A.; Kleuser, B.; Halver, J.E.; Escribá, P.V. Sphingomyelin and sphingomyelin synthase (SMS) in the malignant transformation of glioma cells and in 2-hydroxyoleic acid therapy. Proc. Natl. Acad. Sci. USA 2011, 108, 19569–19574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lladó, V.; Terés, S.; Higuera, M.; Alvarez, R.; Noguera-Salva, M.A.; Halver, J.E.; Escriba, P.V.; Busquets, X. Pivotal role of dihydrofolate reductase knockdown in the anticancer activity of 2-hydroxyoleic acid. Proc. Natl. Acad. Sci. USA 2009, 106, 13754–13758. [Google Scholar] [CrossRef] [Green Version]
- Asano, K.; Duntsch, C.D.; Zhou, Q.; Weimar, J.D.; Bordelon, D.; Robertson, J.H.; Pourmotabbed, T. Correlation of N-cadherin expression in high grade gliomas with tissue invasion. J. Neuro-Oncol. 2004, 70, 3–15. [Google Scholar] [CrossRef]
- Perego, C.; Vanoni, C.; Massari, S.; Raimondi, A.; Pola, S.; Cattaneo, M.G.; Francolini, M.; Vicentini, L.M.; Pietrini, G. Invasive behaviour of glioblastoma cell lines is associated with altered organisation of the cadherin-catenin adhesion system. J. Cell Sci. 2002, 115, 3331–3340. [Google Scholar] [PubMed]
- Sudarkina, O.Y.; Filippenkov, I.B.; Brodsky, I.B.; Limborska, S.A.; Dergunova, L.V. Comparative analysis of sphingomyelin synthase 1 gene expression at the transcriptional and translational levels in human tissues. Mol. Cell. Biochem. 2015, 406, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.J.; Nusse, R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science 2004, 303, 1483–1487. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Woodhead, G.J.; Swaminathan, S.K.; Noles, S.R.; McQuinn, E.R.; Pisarek, A.J.; Stocker, A.M.; Mutch, C.A.; Funatsu, N.; Chenn, A. Cortical neural precursors inhibit their own differentiation via N-cadherin maintenance of beta-catenin signaling. Dev. Cell 2010, 18, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Lee, J.K.; Ahn, S.H.; Lee, J.; Nam, D.H. WNT signaling in glioblastoma and therapeutic opportunities. Lab. Investig. J. Tech. Methods Pathol. 2016, 96, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Pu, P.; Zhang, Z.; Kang, C.; Jiang, R.; Jia, Z.; Wang, G.; Jiang, H. Downregulation of Wnt2 and beta-catenin by siRNA suppresses malignant glioma cell growth. Cancer Gene Therapy 2009, 16, 351–361. [Google Scholar] [CrossRef]
- Rappl, A.; Piontek, G.; Schlegel, J. EGFR-dependent migration of glial cells is mediated by reorganisation of N-cadherin. J. Cell Sci. 2008, 121, 4089–4097. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Wiesmann, M.; Rohan, M.; Chan, V.; Jefferson, A.B.; Guo, L.; Sakamoto, D.; Caothien, R.H.; Fuller, J.H.; Reinhard, C.; et al. Elevated expression of axin2 and hnkd mRNA provides evidence that Wnt/beta -catenin signaling is activated in human colon tumors. Proc. Natl. Acad. Sci. USA 2001, 98, 14973–14978. [Google Scholar] [CrossRef]
- Shtutman, M.; Zhurinsky, J.; Simcha, I.; Albanese, C.; D’Amico, M.; Pestell, R.; Ben-Ze’ev, A. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc. Natl. Acad. Sci. USA 1999, 96, 5522–5527. [Google Scholar] [CrossRef]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Marcilla-Etxenike, A.; Martin, M.L.; Noguera-Salva, M.A.; García-Verdugo, J.M.; Soriano-Navarro, M.; Dey, I.; Escribá, P.V.; Busquets, X. 2-Hydroxyoleic acid induces ER stress and autophagy in various human glioma cell lines. PLoS ONE 2012, 7, e48235. [Google Scholar] [CrossRef]
- Terés, S.; Lladó, V.; Higuera, M.; Barceló-Coblijn, G.; Martin, M.L.; Noguera-Salva, M.A.; Marcilla-Etxenike, A.; García-Verdugo, J.M.; Soriano-Navarro, M.; Saus, C.; et al. Normalization of sphingomyelin levels by 2-hydroxyoleic acid induces autophagic cell death of SF767 cancer cells. Autophagy 2012, 8, 1542–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holcik, M.; Sonenberg, N. Translational control in stress and apoptosis. Nat. Rev. Mol. Cell Biol. 2005, 6, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Kouroku, Y.; Fujita, E.; Tanida, I.; Ueno, T.; Isoai, A.; Kumagai, H.; Ogawa, S.; Kaufman, R.J.; Kominami, E.; Momoi, T. ER stress (PERK/eIF2alpha phosphorylation) mediates the polyglutamine-induced LC3 conversion, an essential step for autophagy formation. Cell Death Differ. 2007, 14, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, A.; Grimm, M.O.; Scherthan, H.; Horsch, M.; Beckers, J.; Fuchs, H.; Gailus-Durner, V.; Hrabe de Angelis, M.; Ford, S.J.; Burton, N.C.; et al. Sphingomyelin Synthase 1 Is Essential for Male Fertility in Mice. PLoS ONE 2016, 11, e0164298. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Watanabe, K.; Itoh, M.; Huan, C.R.; Tong, X.P.; Nakamura, T.; Miki, M.; Iwao, H.; Nakajima, A.; Sakai, T.; et al. CD4+ T-cell dysfunctions through the impaired lipid rafts ameliorate concanavalin A-induced hepatitis in sphingomyelin synthase 1-knockout mice. Int. Immunol. 2012, 24, 327–337. [Google Scholar] [CrossRef] [Green Version]
- Lou, B.; Liu, Q.; Hou, J.; Kabir, I.; Liu, P.; Ding, T.; Dong, J.; Mo, M.; Ye, D.; Chen, Y.; et al. 2-Hydroxy-oleic acid does not activate sphingomyelin synthase activity. J. Biol. Chem. 2018, 293, 18328–18336. [Google Scholar] [CrossRef]
- Sui, X.; Chen, R.; Wang, Z.; Huang, Z.; Kong, N.; Zhang, M.; Han, W.; Lou, F.; Yang, J.; Zhang, Q.; et al. Autophagy and chemotherapy resistance: A promising therapeutic target for cancer treatment. Cell Death Dis. 2013, 4, e838. [Google Scholar] [CrossRef]
- Chang, H.W.; Lee, Y.S.; Nam, H.Y.; Han, M.W.; Kim, H.J.; Moon, S.Y.; Jeon, H.; Park, J.J.; Carey, T.E.; Chang, S.E.; et al. Knockdown of beta-catenin controls both apoptotic and autophagic cell death through LKB1/AMPK signaling in head and neck squamous cell carcinoma cell lines. Cell. Signal. 2013, 25, 839–847. [Google Scholar] [CrossRef]
- Cho, K.J.; van der Hoeven, D.; Zhou, Y.; Maekawa, M.; Ma, X.; Chen, W.; Fairn, G.D.; Hancock, J.F. Inhibition of Acid Sphingomyelinase Depletes Cellular Phosphatidylserine and Mislocalizes K-Ras from the Plasma Membrane. Mol. Cell. Biol. 2015, 36, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Van der Hoeven, D.; Cho, K.J.; Zhou, Y.; Ma, X.; Chen, W.; Naji, A.; Montufar-Solis, D.; Zuo, Y.; Kovar, S.E.; Levental, K.R.; et al. Sphingomyelin metabolism is a regulator of KRAS function. Mol. Cell. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rozhkova, A.V.; Dmitrieva, V.G.; Zhapparova, O.N.; Sudarkina, O.Y.; Nadezhdina, E.S.; Limborska, S.A.; Dergunova, L.V. Human sphingomyelin synthase 1 gene (SMS1): Organization, multiple mRNA splice variants and expression in adult tissues. Gene 2011, 481, 65–75. [Google Scholar] [CrossRef]
- Filippenkov, I.B.; Sudarkina, O.Y.; Limborska, S.A.; Dergunova, L.V. Circular RNA of the human sphingomyelin synthase 1 gene: Multiple splice variants, evolutionary conservatism and expression in different tissues. RNA Biol. 2015, 12, 1030–1042. [Google Scholar] [CrossRef] [Green Version]
- Vladychenskaya, I.P.; Dergunova, L.V.; Dmitrieva, V.G.; Limborska, S.A. Human gene MOB: Structure specification and aspects of transcriptional activity. Gene 2004, 338, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Filippenkov, I.B.; Sudarkina, O.Y.; Limborska, S.A.; Dergunova, L.V. Multi-step splicing of sphingomyelin synthase linear and circular RNAs. Gene 2018, 654, 14–22. [Google Scholar] [CrossRef]
- Mitsutake, S.; Zama, K.; Yokota, H.; Yoshida, T.; Tanaka, M.; Mitsui, M.; Ikawa, M.; Okabe, M.; Tanaka, Y.; Yamashita, T.; et al. Dynamic modification of sphingomyelin in lipid microdomains controls development of obesity, fatty liver, and type 2 diabetes. J. Biol. Chem. 2011, 286, 28544–28555. [Google Scholar] [CrossRef] [PubMed]
- Goswami, C.P.; Nakshatri, H. PROGgeneV2: enhancements on the existing database. BMC Cancer 2014, 14, 970. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Data Set | SMS1 Express | n Total/Censored | Median Survival (mo.) | 2-Year | 5-Year | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n at Risk | Survival (%) | Lower 95%CI | Upper 95%CI | n at Risk | Survival (%) | Lower 95%CI | Upper 95%CI | |||||
| Glioma | GSE4412 | High | 42/18 | 23.87 | 22 | 49.8 | 36.7 | 67.5 | N/A | N/A | N/A | N/A |
| Low | 41/8 | 9.93 | 6 | 15.4 | 7.0 | 33.8 | N/A | N/A | N/A | N/A | ||
| GMB | TCGA-GMB | High | 271/60 | 14.83 | 58 | 27.9 | 22.6 | 34.6 | 13 | 9.9 | 6.3 | 15.6 |
| Low | 271/56 | 12.95 | 36 | 19.4 | 14.7 | 25.7 | 5 | 2.3 | 0.9 | 6.0 | ||
| CN-AML | GSE12417 | High | 82/30 | 11.18 | 26 | 37.5 | 28 | 50.1 | N/A | N/A | N/A | N/A |
| Low | 81/30 | 8.42 | 26 | 35.9 | 26.4 | 48.8 | N/A | N/A | N/A | N/A | ||
| Gastric | GSE62254 | High | 150/90 | N/A | 116 | 76.7 | 70.2 | 83.7 | 97 | 64.0 | 56.8 | 72.2 |
| Low | 149/58 | 38.99 | 91 | 60.4 | 53 | 68.8 | 59 | 40.9 | 33.7 | 49.6 | ||
| KIRC | TCGA-KIRC | High | 117/93 | 93.04 | 71 | 84.9 | 77.8 | 92.6 | 41 | 76.8 | 67.87 | 87.0 |
| Low | 117/74 | 56.35 | 66 | 80.9 | 73.3 | 89.4 | 24 | 48.4 | 37.8 | 62.0 | ||
| Lung | GSE30219 | High | 141/60 | 83.84 | 111 | 77.0 | 70.3 | 84.3 | 69 | 58.1 | 50.3 | 67.1 |
| Low | 141/37 | 28.60 | 77 | 53.8 | 46.2 | 62.7 | 49 | 37.8 | 30.52 | 46.8 | ||
| Lymphoma | GSE4475 | High | 79/46 | 115.00 | 44 | 66.0 | 56.0 | 77.9 | 23 | 53.4 | 42.5 | 67.3 |
| Low | 79/37 | 33.44 | 35 | 53.4 | 43.0 | 66.3 | 19 | 41.4 | 30.7 | 55.9 | ||
| PDAC | GSE21501 | High | 51/22 | 21.70 | 17 | 45.3 | 32.3 | 63.5 | N/A | N/A | N/A | N/A |
| Low | 51/14 | 14.79 | 11 | 22.0 | 12.4 | 38.9 | N/A | N/A | N/A | N/A | ||
| Sarcoma | TCGA-SACR | High | 117/86 | 81.01 | 59 | 77.7 | 69.2 | 87.2 | 16 | 57.5 | 45.2 | 73.1 |
| Low | 117/76 | 53.49 | 50 | 75.0 | 66.2 | 84.9 | 18 | 43.9 | 32.3 | 59.7 | ||
| SKCM | TCGA-SKMC | High | 82/46 | 167.87 | 67 | 88.3 | 81.4 | 95.8 | 41 | 69.0 | 58.9 | 80.8 |
| Low | 81/41 | 48.59 | 47 | 72.9 | 63 | 84.5 | 21 | 47.1 | 35.7 | 62.1 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-García, P.; Rosselló, C.A.; Rodríguez-Lorca, R.; Beteta-Göbel, R.; Fernández-Díaz, J.; Lladó, V.; Busquets, X.; Escribá, P.V. The Opposing Contribution of SMS1 and SMS2 to Glioma Progression and Their Value in the Therapeutic Response to 2OHOA. Cancers 2019, 11, 88. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010088
Fernández-García P, Rosselló CA, Rodríguez-Lorca R, Beteta-Göbel R, Fernández-Díaz J, Lladó V, Busquets X, Escribá PV. The Opposing Contribution of SMS1 and SMS2 to Glioma Progression and Their Value in the Therapeutic Response to 2OHOA. Cancers. 2019; 11(1):88. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010088
Chicago/Turabian StyleFernández-García, Paula, Catalina A. Rosselló, Raquel Rodríguez-Lorca, Roberto Beteta-Göbel, Javier Fernández-Díaz, Victoria Lladó, Xavier Busquets, and Pablo V. Escribá. 2019. "The Opposing Contribution of SMS1 and SMS2 to Glioma Progression and Their Value in the Therapeutic Response to 2OHOA" Cancers 11, no. 1: 88. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11010088