Kinome-Wide Screening with Small Interfering RNA Identified Polo-like Kinase 1 as a Key Regulator of Proliferation in Oral Cancer Cells

 , ,
, ,
Abstract
:1. Introduction
2. Experimental Procedure
2.1. Cell Culture, Treatments, and Transfections
2.2. Western Blot for Protein Expression
2.3. Clonogenic Assay
2.4. Cell Viability
2.5. Flow Cytometry for Cell Cycle Analysis
2.6. Statistical Analysis
3. Results
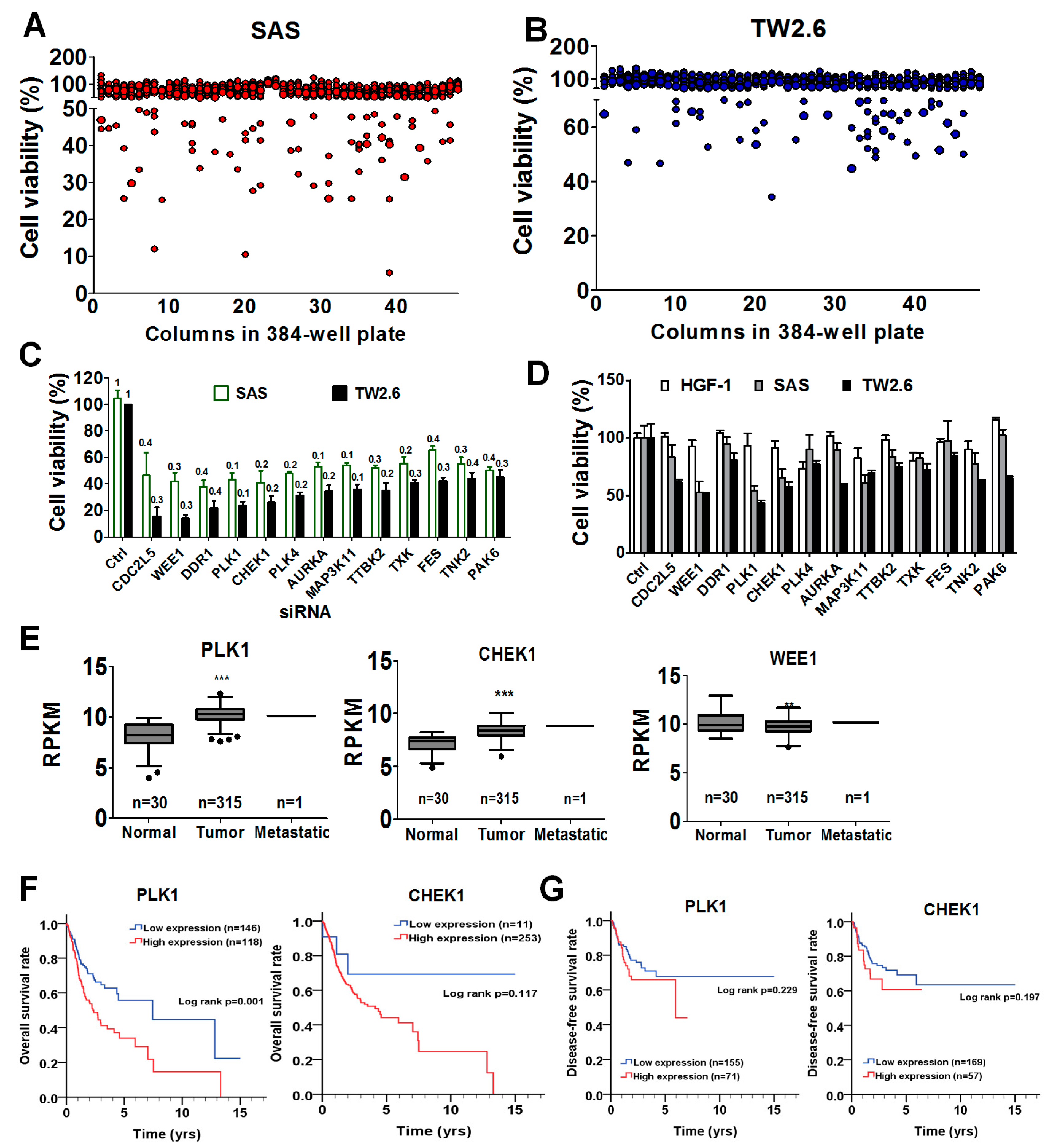
3.1. High-Throughput Screening for Potential Targets in OSCC Cells
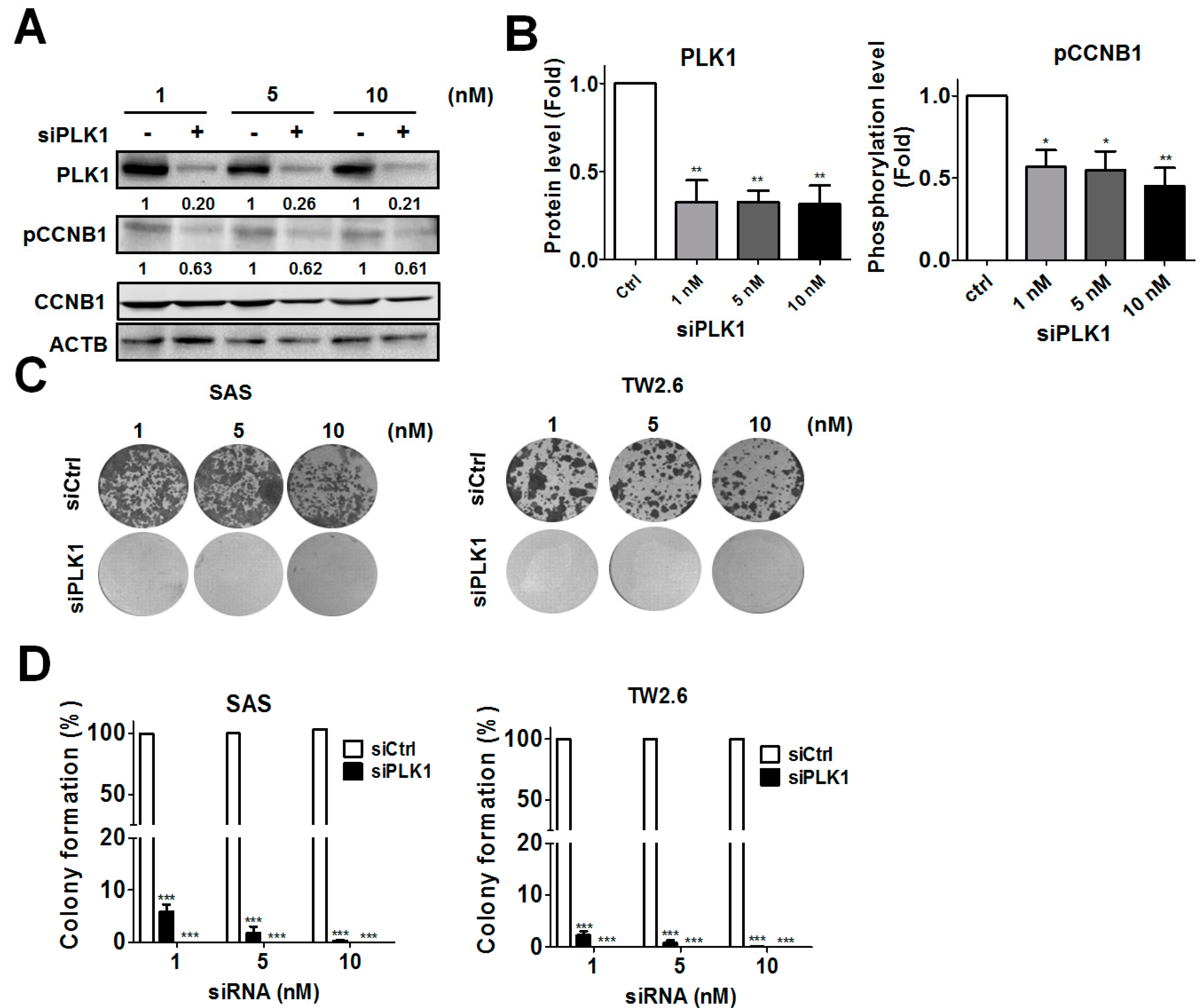
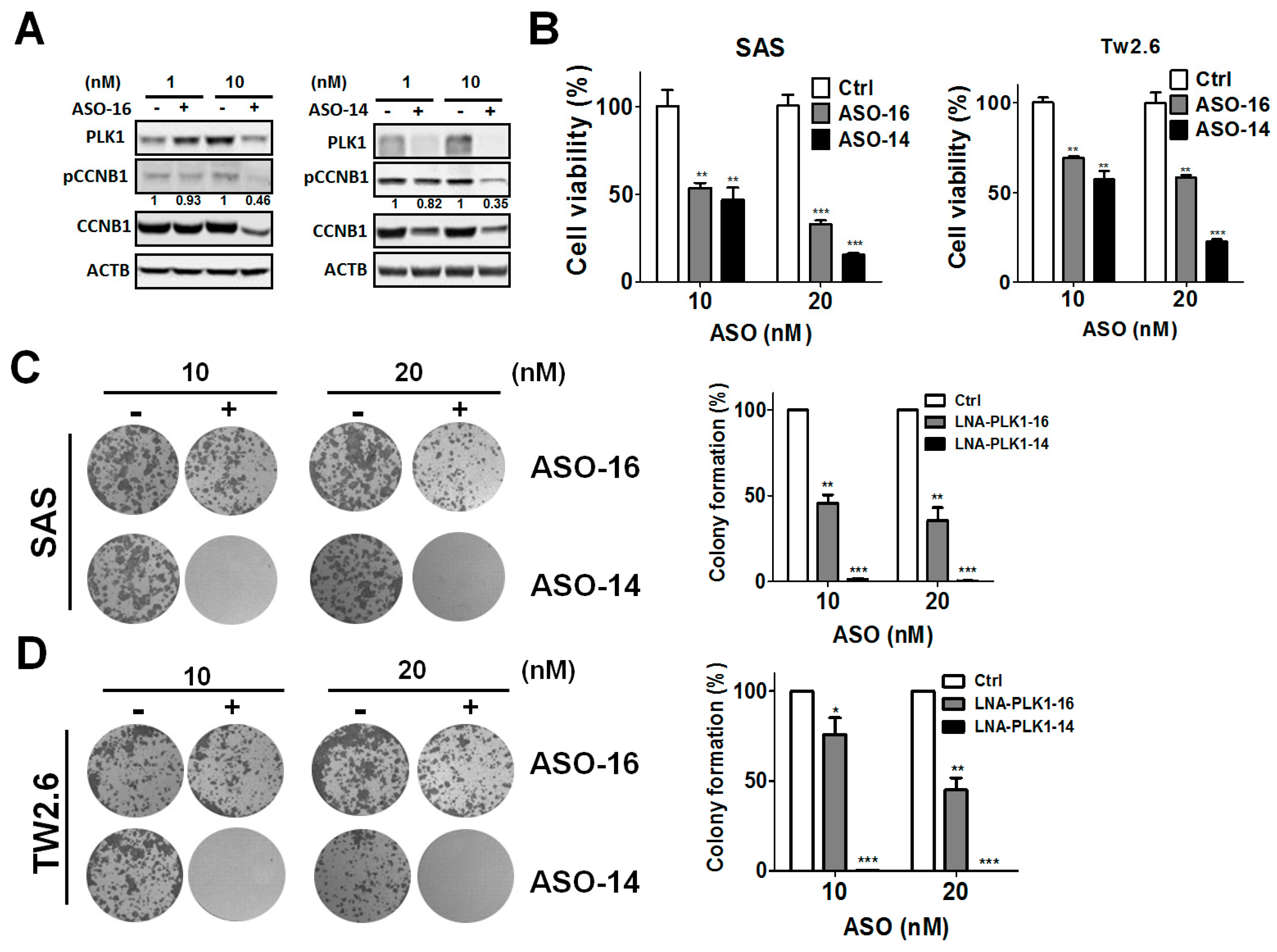
3.2. Genetic Ablation of PLK1-Attenuated Oral Cancer Cell Proliferation
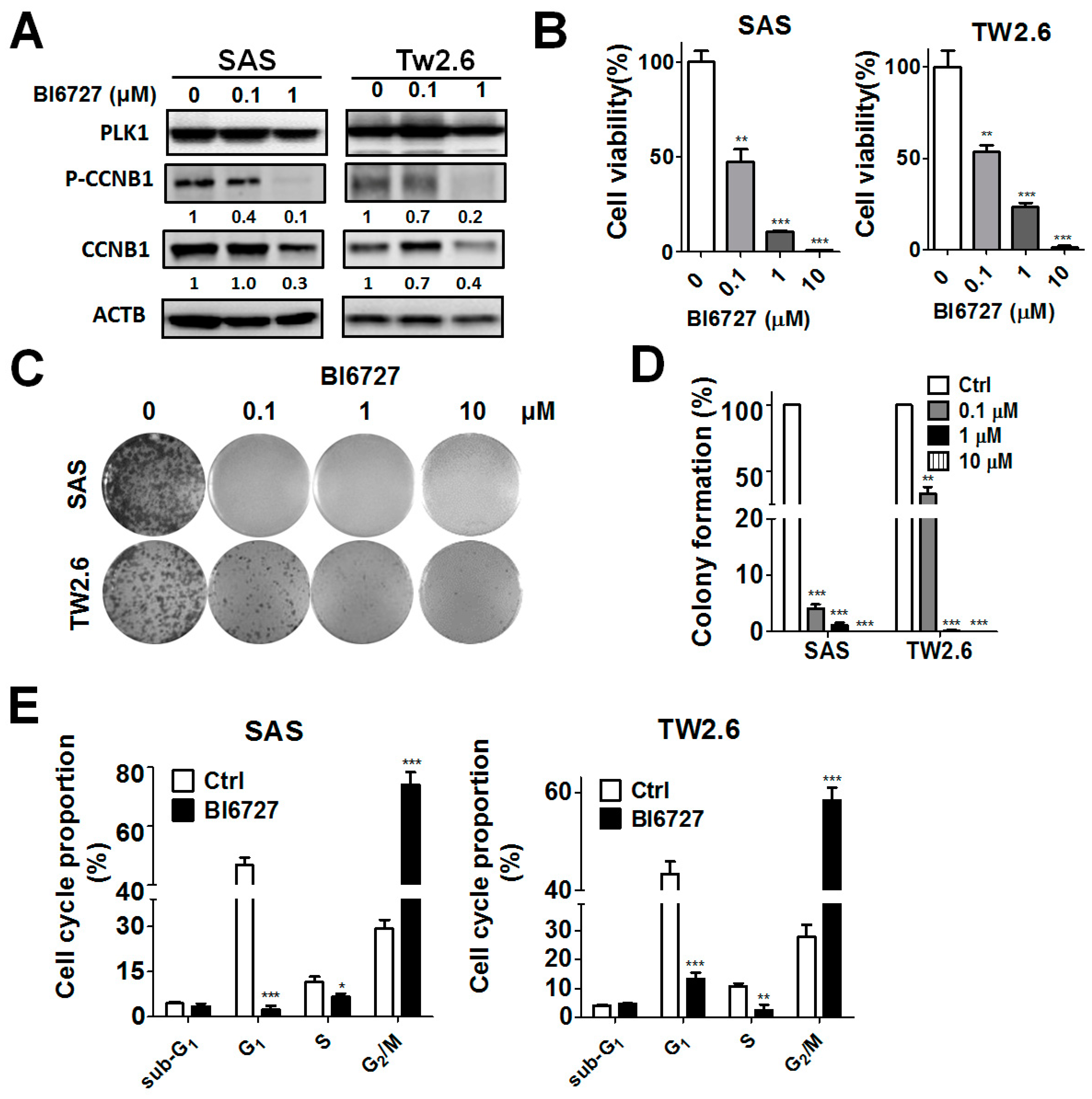
3.3. Pharmacological Ablation of PLK1-Attenuated Oral Cancer Cell Proliferation
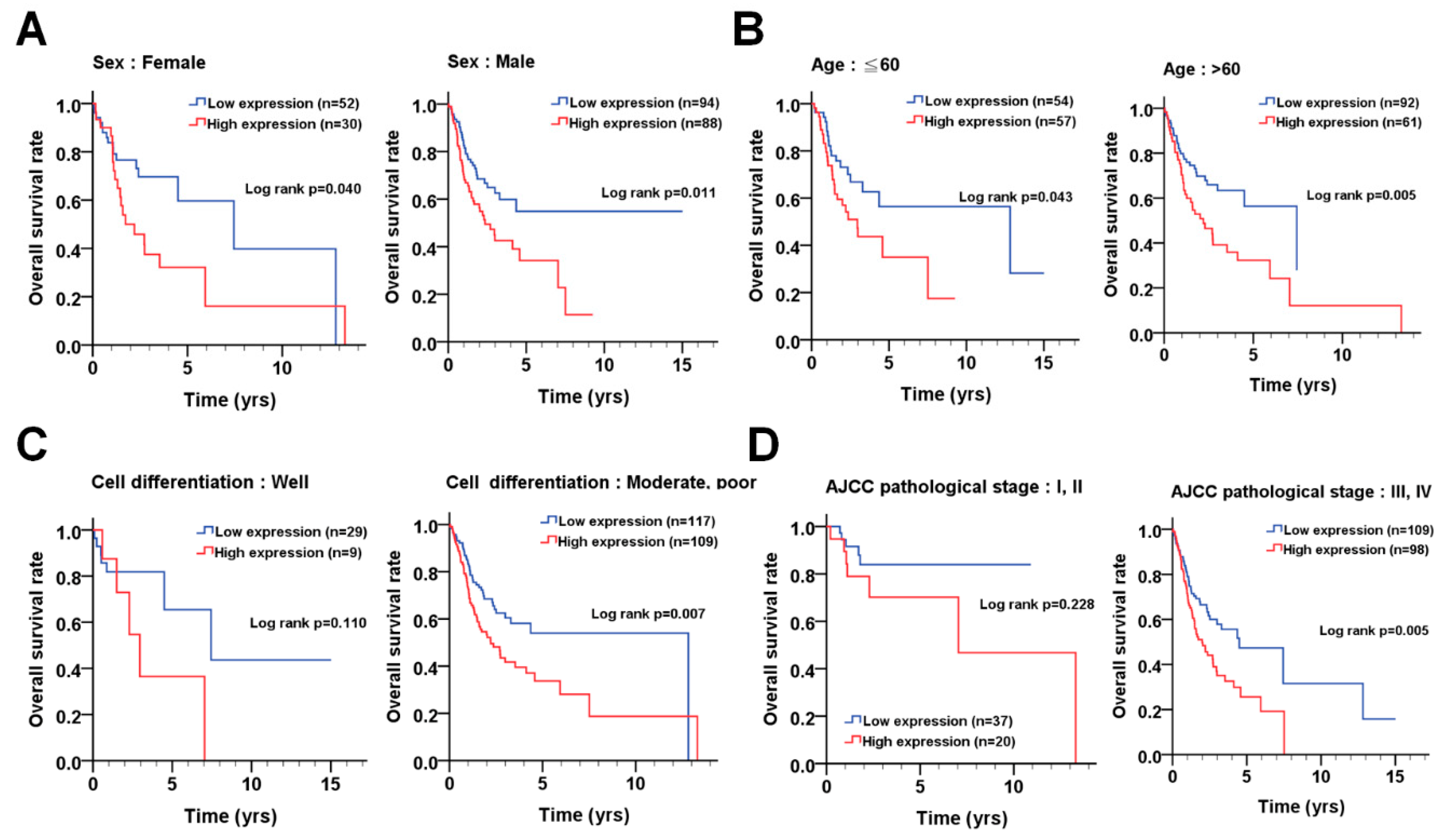
3.4. The Association of PLK1 with Prognosis According to the Demographical and Clinicopathological Features of OSCC Patients
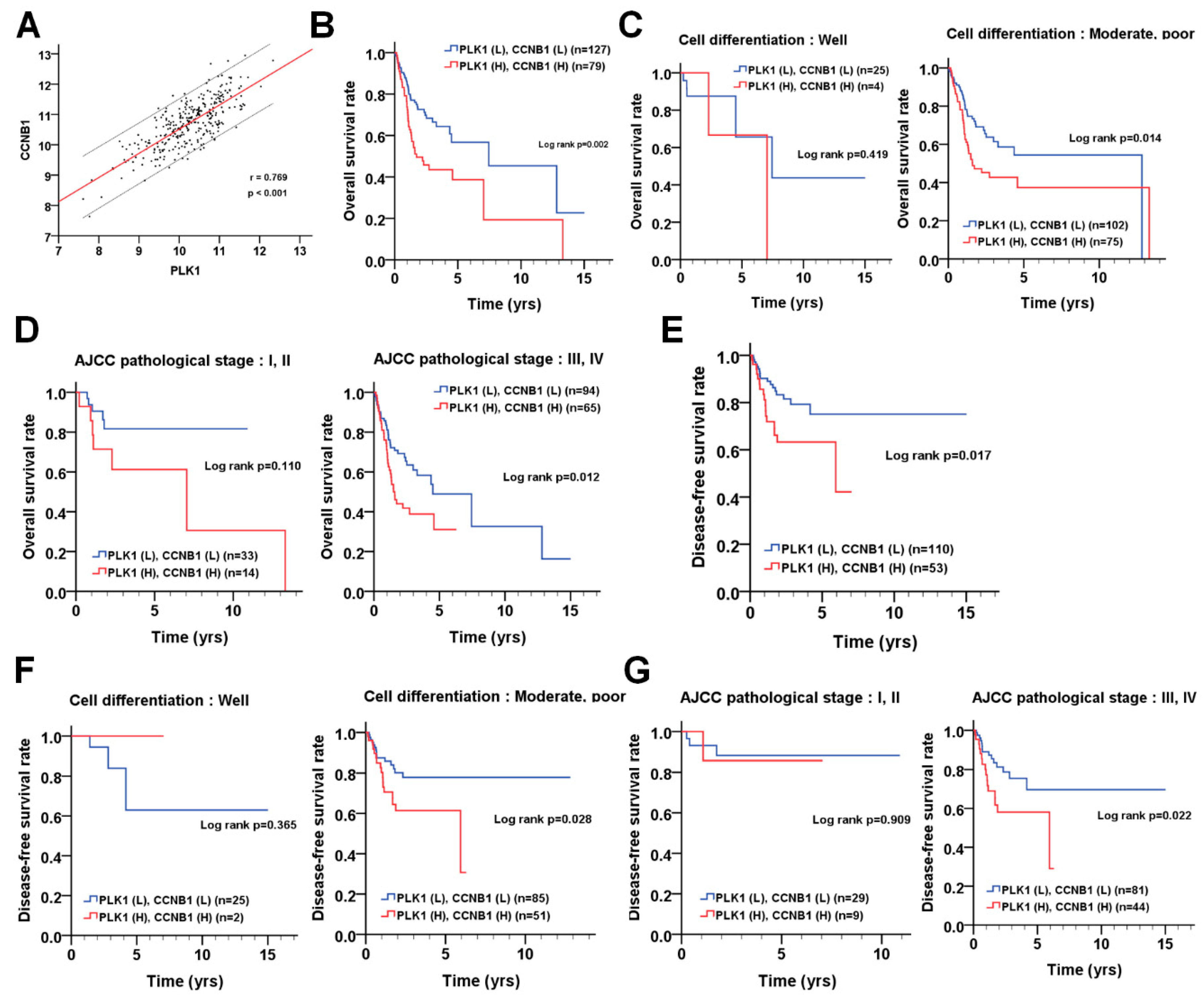
3.5. The Correlation of PLK1 and CCNB1 with Prognosis in Patients with OSCC
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rao, S.V.K.; Mejia, G.; Roberts-Thomson, K.; Logan, R. Epidemiology of oral cancer in Asia in the past decade-an update (2000–2012). Asian Pac. J. Cancer Prev. 2013, 14, 5567–5577. [Google Scholar] [CrossRef] [PubMed]
- Cervino, G.; Fiorillo, L.; Herford, A.S.; Romeo, U.; Bianchi, A.; Crimi, S.; D’Amico, C.; De Stefano, R.; Troiano, G.; Santoro, R.; et al. Molecular Biomarkers Related to Oral Carcinoma: Clinical Trial Outcome Evaluation in a Literature Review. Dis. Markers 2019, 2019, 8040361. [Google Scholar] [CrossRef] [PubMed]
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Jorge, S.E.; Kobayashi, S.S.; Costa, D.B. Epidermal growth factor receptor (EGFR) mutations in lung cancer: Preclinical and clinical data. Braz. J. Med. Biol. Res. 2014, 47, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Lee, S.H.; Lee, J.S.; Nam, B.; Seong, T.W.; Son, J.; Jang, H.; Hong, K.M.; Lee, C.; Kim, S.Y. Aldehyde dehydrogenase inhibition combined with phenformin treatment reversed NSCLC through ATP depletion. Oncotarget 2016, 7, 49397–49410. [Google Scholar] [CrossRef]
- Lee, S.E.; Choi, S.Y.; Kim, S.H.; Song, H.Y.; Yoo, H.L.; Lee, M.Y.; Kang, K.H.; Hwang, H.J.; Jang, E.J.; Kim, D.W. BCR-ABL1 transcripts (MR(4.5)) at post-transplant 3 months as an early predictor for long-term outcomes in chronic myeloid leukemia. Korean J. Intern. Med. 2017, 32, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.W.; Cheng, J.S.; Tsai, W.L.; Liu, P.F.; Goan, Y.G.; Tseng, H.H.; Lee, C.H. The MAP3K7-mTOR Axis Promotes the Proliferation and Malignancy of Hepatocellular Carcinoma Cells. Front. Oncol. 2019, 9, 474. [Google Scholar] [CrossRef]
- Shu, C.W.; Tzeng, Y.D.; Li, J.Y.; Liu, P.F.; Kuo, S.Y.; Liu, L.F.; Hsieh, C.W.; Lee, C.H.; Wu, C.H.; Hsiao, M.; et al. Kinome-Wide siRNA Screening Identifies Src-Enhanced Resistance of Chemotherapeutic Drugs in Triple-Negative Breast Cancer Cells. Front. Pharmacol. 2018, 9, 1285. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.F.; Tsai, K.L.; Hsu, C.J.; Tsai, W.L.; Cheng, J.S.; Chang, H.W.; Shiau, C.W.; Goan, Y.G.; Tseng, H.H.; Wu, C.H.; et al. Drug Repurposing Screening Identifies Tioconazole as an ATG4 Inhibitor that Suppresses Autophagy and Sensitizes Cancer Cells to Chemotherapy. Theranostics 2018, 8, 830–845. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Q.; Wang, X. PLK1, A Potential Target for Cancer Therapy. Transl. Oncol. 2017, 10, 22–32. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, G.; Kong, C. Targeted inhibition of Polo-like kinase 1 by a novel small-molecule inhibitor induces mitotic catastrophe and apoptosis in human bladder cancer cells. J. Cell Mol. Med. 2017, 21, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.; Wasner, M.; Reinhard, W.; Haugwitz, U.; Lange-zu Dohna, C.; Mössner, J.; Engeland, K. The tumour suppressor protein p53 can repress transcription of cyclin B. Nucleic Acids Res. 2000, 28, 4410–4418. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.K.; Corey, D.R. Silencing disease genes in the laboratory and the clinic. J. Pathol. 2012, 226, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Chery, J. RNA therapeutics: RNAi and antisense mechanisms and clinical applications. Postdr. J. 2016, 4, 35–50. [Google Scholar] [CrossRef]
- Do, K.; Doroshow, J.H.; Kummar, S. Wee1 kinase as a target for cancer therapy. Cell Cycle 2013, 12, 3159–3164. [Google Scholar] [CrossRef] [PubMed]
- McNeely, S.; Beckmann, R.; Bence Lin, A.K. CHEK again: Revisiting the development of CHK1 inhibitors for cancer therapy. Pharmacol. Ther. 2014, 142, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hunter, T. Roles of Chk1 in cell biology and cancer therapy. Int. J. Cancer 2014, 134, 1013–1023. [Google Scholar] [CrossRef] [PubMed]
- Matheson, C.J.; Backos, D.S.; Reigan, P. Targeting WEE1 Kinase in Cancer. Trends Pharm. Sci. 2016, 37, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Mross, K.; Dittrich, C.; Aulitzky, W.E.; Strumberg, D.; Schutte, J.; Schmid, R.M.; Hollerbach, S.; Merger, M.; Munzert, G.; Fleischer, F.; et al. A randomised phase II trial of the Polo-like kinase inhibitor BI 2536 in chemo-naive patients with unresectable exocrine adenocarcinoma of the pancreas-a study within the Central European Society Anticancer Drug Research (CESAR) collaborative network. Br. J. Cancer. 2012, 107, 280–286. [Google Scholar] [CrossRef]
- Stadler, W.M.; Vaughn, D.J.; Sonpavde, G.; Vogelzang, N.J.; Tagawa, S.T.; Petrylak, D.P.; Rosen, P.; Lin, C.C.; Mahoney, J.; Modi, S.; et al. An open-label, single-arm, phase 2 trial of the Polo-like kinase inhibitor volasertib (BI 6727) in patients with locally advanced or metastatic urothelial cancer. Cancer 2014, 120, 976–982. [Google Scholar] [CrossRef]
- Bartucci, M.; Svensson, S.; Romania, P.; Dattilo, R.; Patrizii, M.; Signore, M.; Navarra, S.; Lotti, F.; Biffoni, M.; Pilozzi, E.; et al. Therapeutic targeting of Chk1 in NSCLC stem cells during chemotherapy. Cell Death Differ. 2012, 19, 768–778. [Google Scholar] [CrossRef] [PubMed]
- Rundle, S.; Bradbury, A.; Drew, Y.; Curtin, N.J. Targeting the ATR-CHK1 Axis in Cancer Therapy. Cancers 2017, 9, 41. [Google Scholar] [CrossRef] [PubMed]
- Knecht, R.; Elez, R.; Oechler, M.; Solbach, C.; von Ilberg, C.; Strebhardt, K. Prognostic significance of polo-like kinase (PLK) expression in squamous cell carcinomas of the head and neck. Cancer Res. 1999, 59, 2794–2797. [Google Scholar] [PubMed]
- Gutteridge, R.E.; Ndiaye, M.A.; Liu, X.; Ahmad, N. Plk1 Inhibitors in Cancer Therapy: From Laboratory to Clinics. Mol. Cancer Ther. 2016, 15, 1427–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | No. (%) | PLK1 | ||
|---|---|---|---|---|
| Mean ± SD | Median | p-Value | ||
| Sex | ||||
| Female | 83 (31.1) | 10.15 ± 0.76 | 10.19 | 0.123 * |
| Male | 184 (68.9) | 10.30 ± 0.75 | 10.36 | |
| Age, y | ||||
| ≦40 | 10 (3.8) | 10.03 ± 0.85 | 10.20 | 0.211 † |
| 41–50 | 33(12.4) | 10.31 ± 0.60 | 10.38 | |
| 51–60 | 70 (26.3) | 10.39 ± 0.63 | 10.46 | |
| >60 | 153 (57.6) | 10.18 ± 0.82 | 10.21 | |
| Cell differentiation | ||||
| Well | 38 (14.2) | 9.89 ± 0.74ab | 9.97 | 0.004† |
| Moderate | 177 (66.3) | 10.30 ± 0.73a | 10.29 | |
| Poor | 52 (19.5) | 10.36 ± 0.78b | 10.45 | |
| AJCC pathological stage | ||||
| I | 18 (6.7) | 9.89 ± 0.71 | 9.91 | 0.168 † |
| II | 40(15.0) | 10.21 ± 0.79 | 10.25 | |
| III | 55 (20.6) | 10.24 ± 0.78 | 10.26 | |
| IV | 154(57.7) | 10.31 ± 0.73 | 10.33 | |
| T classification | ||||
| T1 | 27 (10.1) | 9.95 ± 0.71 | 9.92 | 0.093 † |
| T2 | 80 (30.0) | 10.23 ± 0.77 | 10.25 | |
| T3 | 57 (21.3) | 10.24 ± 0.70 | 10.26 | |
| T4 | 103 (38.6) | 10.36 ± 0.76 | 10.41 | |
| N classification | ||||
| N0 | 119 (44.6) | 10.20 ± 0.73 | 10.24 | 0.647 § |
| N1 | 46 (17.2) | 10.27 ± 0.97 | 10.33 | |
| N2 | 99 (37.1) | 10.30 ± 0.66 | 10.31 | |
| N3 | 3 (1.1) | 10.67 ± 0.87 | 11.06 | |
| Variable | PLK1 | No. (%) | CHR (95% CI) | p-Value * | AHR (95% CI) | p-Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 52 (63.4) | 1.00 | 1.00 | ||
| High | 30 (36.6) | 1.98 (1.02–3.87) | 0.044 | 2.06 (0.97–4.39) | 0.060 a | |
| Male | Low | 94 (51.6) | 1.00 | 1.00 | ||
| High | 88 (48.4) | 1.78 (1.13–2.79) | 0.012 | 1.64 (1.04–2.59) | 0.034 a | |
| Age, yrs | ||||||
| ≦60 | Low | 54 (48.6) | 1.00 | 1.00 | ||
| High | 57 (51.4) | 1.85 (1.01–3.38) | 0.046 | 1.76 (0.96–3.22) | 0.069 a | |
| >60 | Low | 92 (60.1) | 1.00 | 1.00 | ||
| High | 61 (39.9) | 1.97 (1.22–3.18) | 0.006 | 1.86 (1.12–3.11) | 0.017 a | |
| Cell differentiation | ||||||
| Well | Low | 29 (76.3) | 1.00 | 1.00 | ||
| High | 9 (23.7) | 2.54 (0.78–8.35) | 0.124 | 4.32 (1.19–15.70) | 0.026 b | |
| Moderate, poor | Low | 117 (51.8) | 1.00 | 1.00 | ||
| High | 109 (48.2) | 1.71 (1.15–2.55) | 0.008 | 1.57 (1.06–2.34) | 0.026 b | |
| AJCC pathological stage | ||||||
| I, II | Low | 37 (64.9) | 1.00 | 1.00 | ||
| High | 20 (35.1) | 2.06 (0.62–6.84) | 0.237 | 2.09 (0.63–6.96) | 0.229 c | |
| III, IV | Low | 109 (52.7) | 1.00 | 1.00 | ||
| High | 98 (47.3) | 1.75 (1.18–2.60) | 0.005 | 1.67 (1.11–2.51) | 0.014 c | |
| T classification | ||||||
| T1, T2 | Low | 65 (62.5) | 1.00 | 1.00 | ||
| High | 39 (37.5) | 2.48 (1.14–5.43) | 0.023 | 2.51 (1.14–5.50) | 0.022 d | |
| T3, T4 | Low | 81 (50.6) | 1.00 | 1.00 | ||
| High | 79 (49.4) | 1.50 (0.98–2.29) | 0.063 | 1.42 (0.91–2.21) | 0.118 d | |
| N classification | ||||||
| N0 | Low | 70 (59.3) | 1.00 | 1.00 | ||
| High | 48 (40.7) | 2.32 (1.21–4.45) | 0.011 | 2.24 (1.12–4.49) | 0.023 e | |
| N1, N2, N3 | Low | 76 (52.1) | 1.00 | 1.00 | ||
| High | 70 (47.9) | 1.60 (1.01–2.53) | 0.045 | 1.37 (0.86–2.19) | 0.187 e | |
| Postoperative RT | ||||||
| No | Low | 56 (56.6) | 1.00 | 1.00 | ||
| High | 43 (43.4) | 2.54 (1.40–4.64) | 0.002 | 2.12 (1.14–3.92) | 0.017 a | |
| Yes | Low | 73 (52.5) | 1.00 | 1.00 | ||
| High | 66 (47.5) | 2.08 (1.17–3.73) | 0.013 | 1.95 (1.07–3.54) | 0.029 a |
| Variable | PLK1 | No. (%) | CHR (95% CI) | p-Value * | AHR (95% CI) | p-Value † |
|---|---|---|---|---|---|---|
| Sex | ||||||
| Female | Low | 56 (76.7) | 1.00 | 1.00 | ||
| High | 17 (23.3) | 1.87 (0.85–4.13) | 0.120 | 1.63 (0.70–3.82) | 0.257 a | |
| Male | Low | 99 (64.7) | 1.00 | 1.00 | ||
| High | 54 (35.3) | 1.24 (0.73–2.12) | 0.426 | 1.09 (0.63–1.87) | 0.765 a | |
| Age, yrs | ||||||
| ≦60 | Low | 61 (64.9) | 1.00 | 1.00 | ||
| High | 33 (35.1) | 1.22 (0.61–2.46) | 0.571 | 1.11 (0.55–2.25) | 0.766 a | |
| >60 | Low | 94 (71.2) | 1.00 | 1.00 | ||
| High | 38 (28.8) | 1.54 (0.87–2.73) | 0.137 | 1.31 (0.72–2.39) | 0.374 a | |
| Cell differentiation | ||||||
| Well | Low | 32 (91.4) | 1.00 | 1.00 | ||
| High | 3 (8.6) | 1.33 (0.27–6.65) | 0.731 | 3.56 (0.63–20.25) | 0.153 b | |
| Moderate, poor | Low | 123 (64.4) | 1.00 | 1.00 | ||
| High | 68 (35.6) | 1.34 (0.84–2.13) | 0.222 | 1.15 (0.72–1.85) | 0.557 b | |
| AJCC pathological stage | ||||||
| I, II | Low | 41 (78.8) | 1.00 | 1.00 | ||
| High | 11 (21.2) | 1.98 (0.49–7.95) | 0.334 | 2.24 (0.55–9.17) | 0.262 c | |
| III, IV | Low | 114 65.5) | 1.00 | 1.00 | ||
| High | 60 (34.5) | 1.20 (0.76–1.92) | 0.435 | 1.14 (0.71–1.85) | 0.582 c | |
| T classification | ||||||
| T1, T2 | Low | 71 (74.0) | 1.00 | 1.00 | ||
| High | 25 (26.0) | 1.61 (0.64–4.04) | 0.310 | 1.40 (0.54–3.60) | 0.489 d | |
| T3, T4 | Low | 84 (64.6) | 1.00 | 1.00 | ||
| High | 46 (35.4) | 1.20 (0.72–1.99) | 0.480 | 1.16 (0.69–1.95) | 0.580 d | |
| N classification | ||||||
| N0 | Low | 80 (74.8) | 1.00 | 1.00 | ||
| High | 27 (25.2) | 1.35 (0.64–2.86) | 0.428 | 1.11 (0.52–2.40) | 0.785 e | |
| N1, N2 | Low | 75 (63.0) | 1.00 | 1.00 | ||
| High | 44 (37.0) | 1.31 (0.75–2.28) | 0.342 | 1.23 (0.70–2.17) | 0.466 e | |
| Postoperative RT | ||||||
| No | Low | 62 (70.5) | 1.00 | 1.00 | ||
| High | 26 (29.5) | 1.75 (0.93–3.29) | 0.084 | 1.28 (0.66–2.46) | 0.467 a | |
| Yes | Low | 85 (66.4) | 1.00 | 1.00 | ||
| High | 43 (33.6) | 1.51 (0.79–2.88) | 0.214 | 1.43 (0.74–2.78) | 0.288 a |
| Variable | No. (%) | CHR (95% CI) | p-Value | AHR (95% CI) | p-Value | |
|---|---|---|---|---|---|---|
| Overall survival | ||||||
| PLK1 | Low | 146 (55.3) | 1.00 | 1.00 | ||
| High | 118 (44.7) | 1.87 (1.29–2.71) | 0.001 * | 1.73 (1.18–2.53) | 0.005 a | |
| CCNB1 | Low | 166 (62.9) | 1.00 | 1.00 | ||
| High | 98 (37.1) | 1.53 (1.06–2.22) | 0.024 * | 1.50 (1.03–2.18) | 0.035 a | |
| PLK1 (L) CCNB1 (L) | 127 (48.1) | 1.00 | 1.00 | |||
| either | 58 (22.0) | 1.33 (0.88–2.01) | 0.172 * | 1.79 (1.12–2.85) | 0.015 † | |
| PLK1 (H) CCNB1 (H) | 79(29.9) | 1.60 (1.09–2.33) | 0.016 * | 1.98 (1.29–3.05) | 0.002 † | |
| Disease-free survival | ||||||
| PLK1 | Low | 155 (68.6) | 1.00 | 1.00 | ||
| High | 71 (31.4) | 1.39 (0.81–2.40) | 0.231 * | 1.19 (0.68–2.08) | 0.540 a | |
| CCNB1 | Low | 128 (56.6) | 1.00 | 1.00 | ||
| High | 98 (43.4) | 2.30 (1.34–3.95) | 0.003 * | 2.22 (1.28–3.86) | 0.005 a | |
| PLK1 (L) CCNB1 (L) | 110 (48.7) | 1.00 | 1.00 | |||
| either | 63 (27.9) | 1.69 (0.97–2.93) | 0.063 * | 2.30 (1.21–4.36) | 0.011 † | |
| PLK1 (H) CCNB1 (H) | 53 (23.5) | 1.55 (0.87–2.74) | 0.136 * | 2.20 (1.13–4.27) | 0.020 † |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goan, Y.-G.; Liu, P.-F.; Chang, H.-W.; Chen, H.-C.; Chen, W.-C.; Kuo, S.-M.; Lee, C.-H.; Shu, C.-W. Kinome-Wide Screening with Small Interfering RNA Identified Polo-like Kinase 1 as a Key Regulator of Proliferation in Oral Cancer Cells. Cancers 2019, 11, 1117. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11081117
Goan Y-G, Liu P-F, Chang H-W, Chen H-C, Chen W-C, Kuo S-M, Lee C-H, Shu C-W. Kinome-Wide Screening with Small Interfering RNA Identified Polo-like Kinase 1 as a Key Regulator of Proliferation in Oral Cancer Cells. Cancers. 2019; 11(8):1117. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11081117
Chicago/Turabian StyleGoan, Yih-Gang, Pei-Feng Liu, Hsueh-Wei Chang, Hung-Chih Chen, Wen-Chi Chen, Shyh-Ming Kuo, Cheng-Hsin Lee, and Chih-Wen Shu. 2019. "Kinome-Wide Screening with Small Interfering RNA Identified Polo-like Kinase 1 as a Key Regulator of Proliferation in Oral Cancer Cells" Cancers 11, no. 8: 1117. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11081117