EP4 and Class III β-Tubulin Expression in Uterine Smooth Muscle Tumors: Implications for Prognosis and Treatment

 , ,
, ,
Abstract
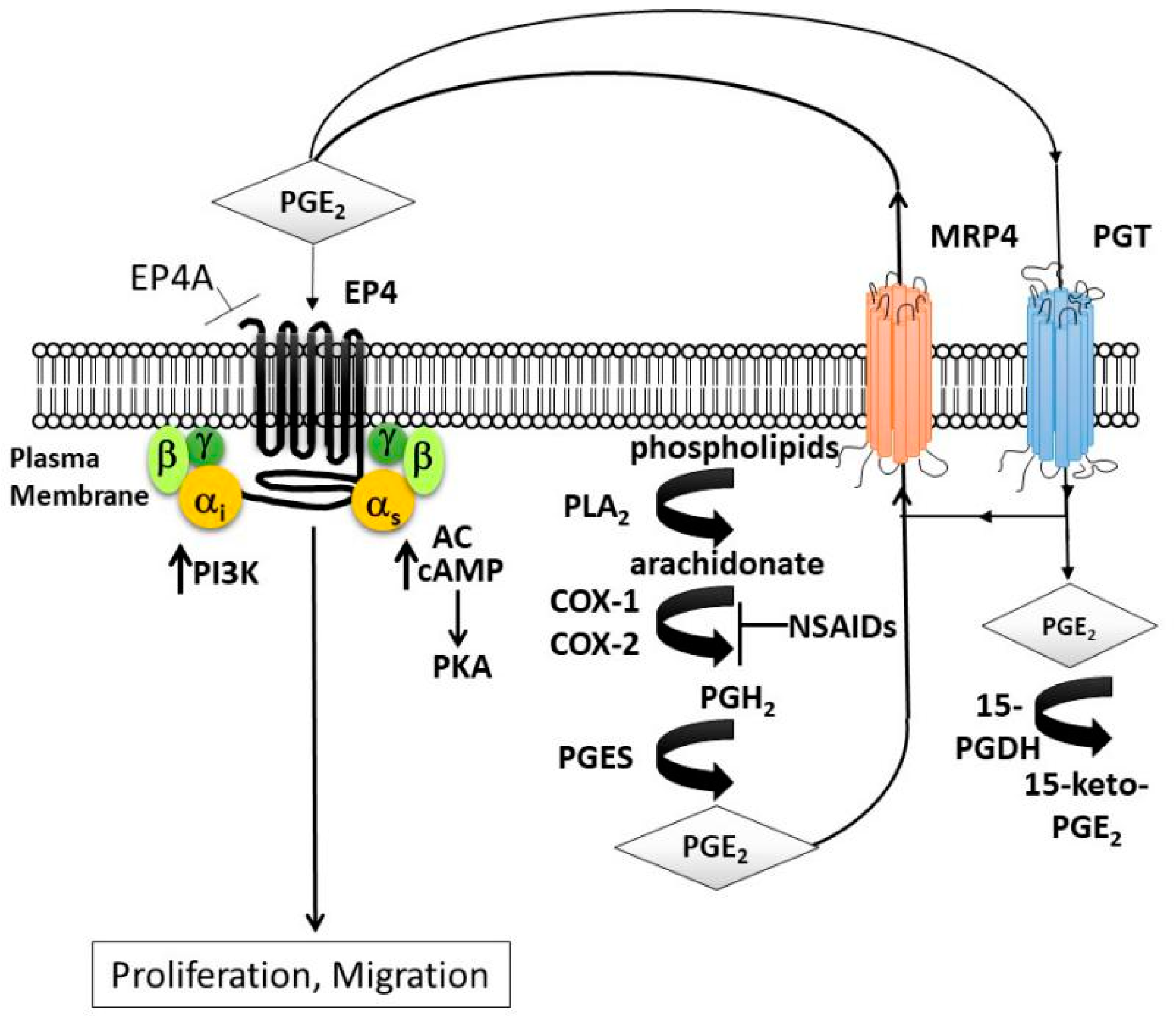
:1. Introduction
2. Results
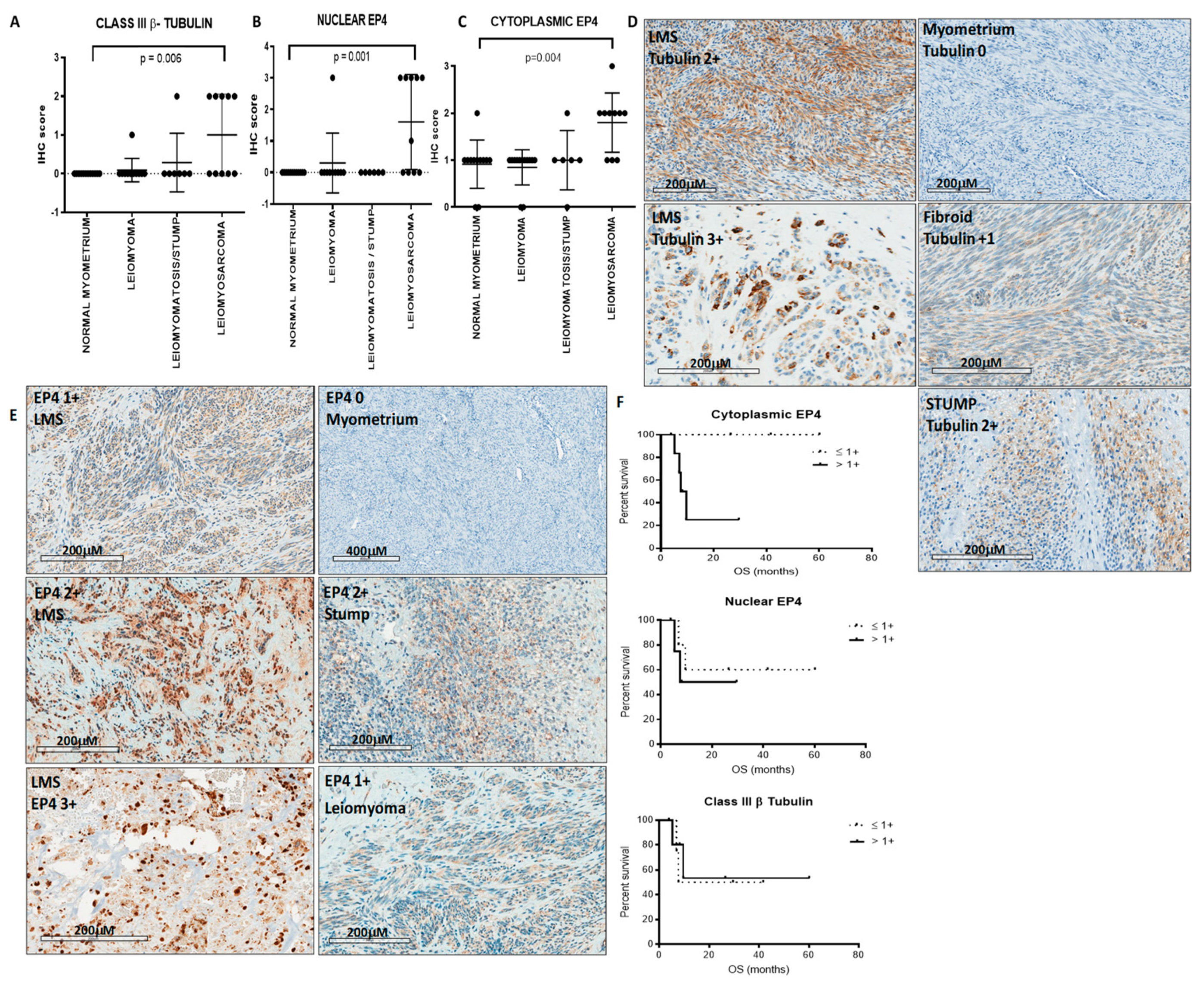
2.1. LMS Expresses Class III β-Tubulin and EP4
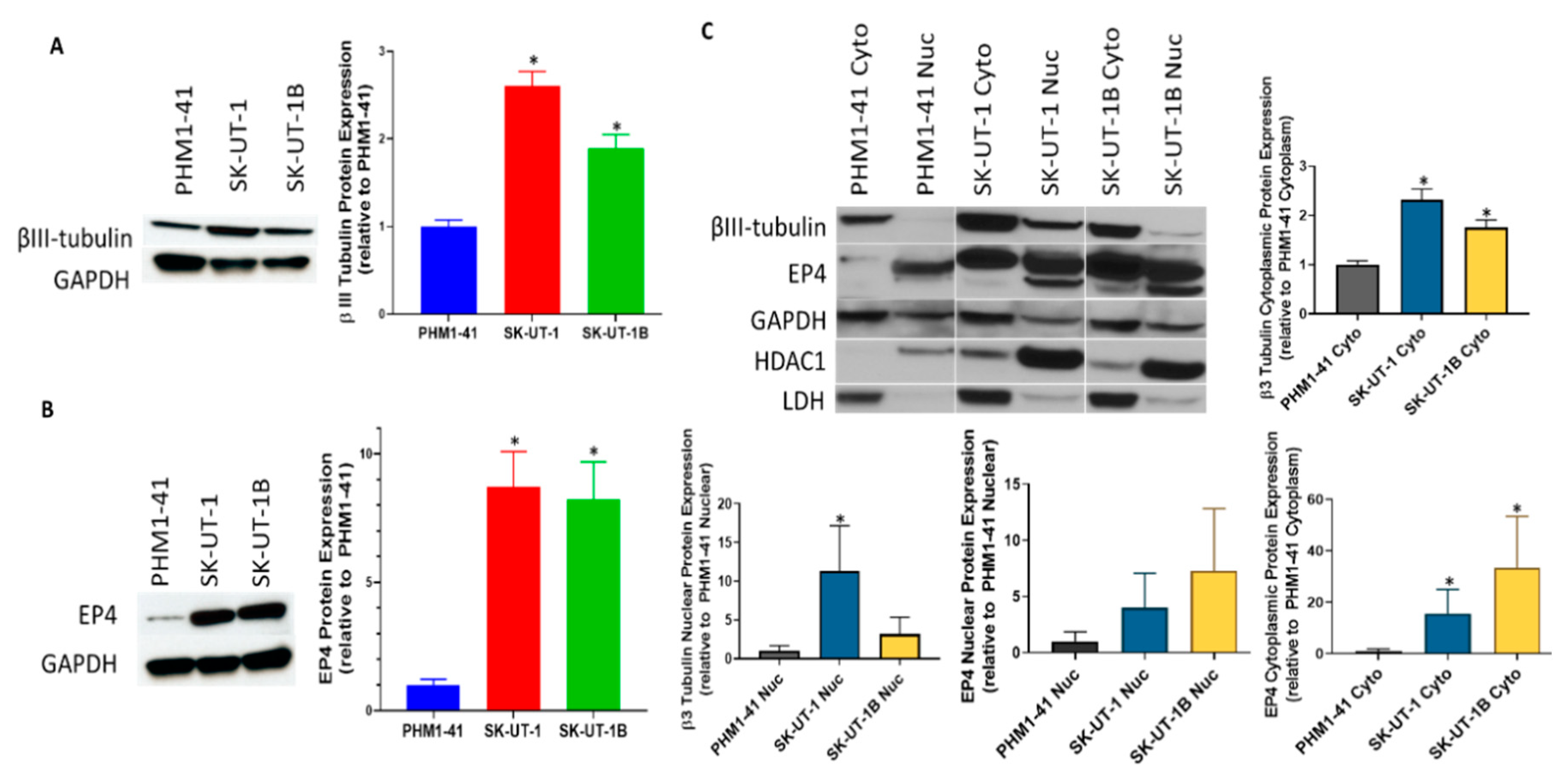
2.2. Carcinosarcoma Cell Lines SK-UT-1 and SK-UT-1B Overexpresses Class III β-Tubulin and EP4 in Comparison to Normal Myometrium
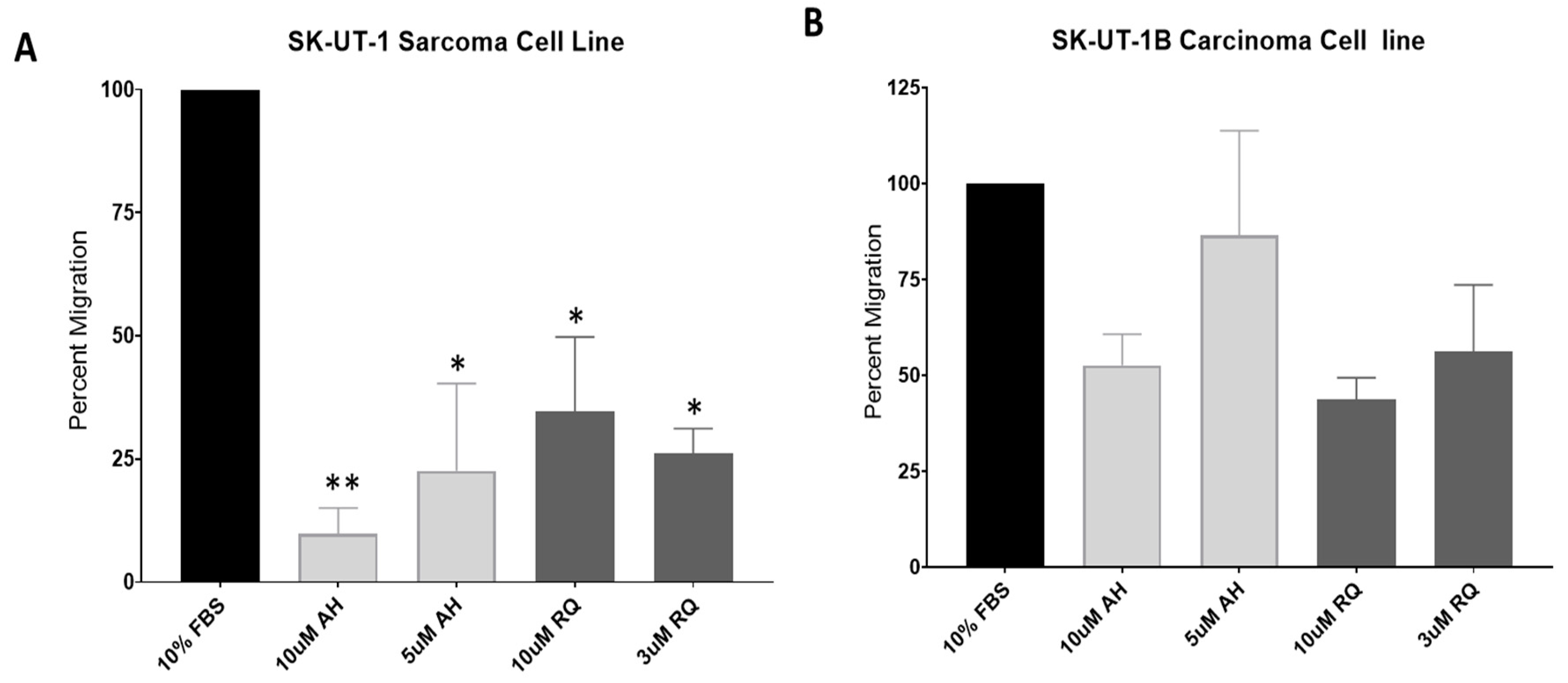
2.3. EP4 Antagonists Inhibit Migration of SK-UT-1 and SK-UT-1B
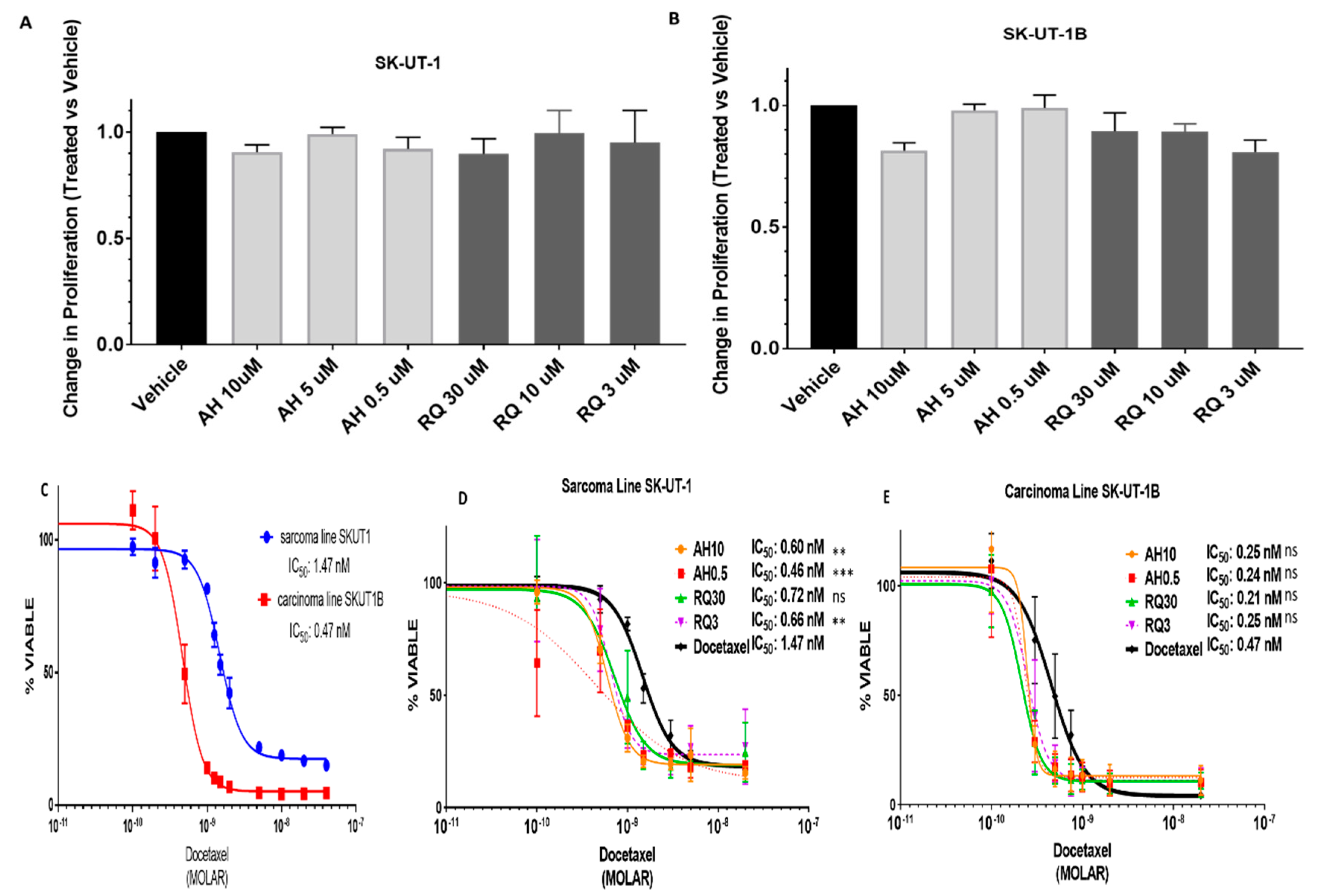
2.4. Pre-Treatment with EP4 Antagonists Enhances Sensitivity to Docetaxel
3. Materials and Methods
3.1. Patients
3.2. Immunohistochemistry
3.3. Cell Lines
3.4. Western Blot Analysis
3.5. Drugs
3.6. Proliferation Assays
3.7. Migration Assays
3.8. Bioinformatics Analysis
3.9. Statistical Analyses
4. Discussion
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Kapp, D.S.; Shin, J.Y.; Chan, J.K. Prognostic factors and survival in 1396 patients with uterine leiomyosarcomas: Emphasis on impact of lymphadenectomy and oophorectomy. Cancer 2008, 112, 820–830. [Google Scholar] [CrossRef]
- English, D.P.; Roque, D.M.; Santin, A.D. Class III b-tubulin overexpression in gynecologic tumors: Implications for the choice of microtubule targeted agents? Expert Rev. Anticancer Ther. 2013, 13, 63–74. [Google Scholar] [CrossRef]
- Conde, C.; Cáceres, A. Microtubule assembly, organization and dynamics in axons and dendrites. Nat. Rev. Neurosci. 2009, 10, 319–332. [Google Scholar] [CrossRef]
- Carrara, L.; Guzzo, F.; Roque, D.M.; Bellone, S.; Emiliano, C.; Sartori, E.; Pecorelli, S.; Schwartz, P.E.; Rutherford, T.J.; Santin, A.D. Differential in vitro sensitivity to patupilone versus paclitaxel in uterine and ovarian carcinosarcoma cell lines is linked to tubulin-beta-III expression. Gynecol. Oncol. 2012, 125, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Terry, S.; Ploussard, G.; Allory, Y.; Nicolaiew, N.; Boissière-Michot, F.; Maillé, P.; Kheuang, L.; Coppolani, E.; Ali, A.; Bibeau, F.; et al. Increased expression of class III beta-tubulin in castration-resistant human prostate cancer. Br. J. Cancer 2009, 101, 951–956. [Google Scholar] [CrossRef]
- Roque, D.M.; Bellone, S.; English, D.P.; Buza, N.; Cocco, E.; Gasparrini, S.; Bortolomai, I.; Ratner, E.; Silasi, D.-A.; Azodi, M.; et al. Tubulin-β-III overexpression by uterine serous carcinomas is a marker for poor overall survival after platinum/taxane chemotherapy and sensitivity to epothilones. Cancer 2013, 119, 2582–2592. [Google Scholar] [CrossRef]
- Roque, D.M.; Buza, N.; Glasgow, M.; Bellone, S.; Bortolomai, I.; Gasparrini, S.; Cocco, E.; Ratner, E.; Silasi, D.-A.; Azodi, M.; et al. Class III β-tubulin overexpression within the tumor microenvironment is a prognostic biomarker for poor overall survival in ovarian cancer patients treated with neoadjuvant carboplatin/paclitaxel. Clin. Exp. Metastasis 2014, 31, 101–110. [Google Scholar] [CrossRef]
- Roque, D.M.; Bellone, S.; Buza, N.; Romani, C.; Cocco, E.; Bignotti, E.; Ravaggi, A.; Rutherford, T.J.; Schwartz, P.E.; Pecorelli, S.; et al. Class III β-tubulin overexpression in ovarian clear cell and serous carcinoma as a maker for poor overall survival after platinum/taxane chemotherapy and sensitivity to patupilone. Am. J. Obstet. Gynecol. 2013, 209, e1–e9. [Google Scholar] [CrossRef]
- Rowinsky, E.K. The development and clinical utility of the taxane class of antimicrotubule chemotherapy agents. Ann. Rev. Med. 1997, 48, 353–374. [Google Scholar] [CrossRef]
- Yahiro, K.; Matsumoto, Y.; Fukushi, J.-I.; Kawaguchi, K.-I.; Endo, M.; Setsu, N.; IIda, K.; Fukushima, S.; Nakagawa, M.; Kimura, A.; et al. Class III β-Tubulin Overexpression Induces Chemoresistance to Eribulin in a Leiomyosarcoma Cell Line. Anal. Cell. Pathol. Amst. 2018, 2018, 8987568. [Google Scholar] [CrossRef]
- D’Angelo, E.; Prat, J. Uterine sarcomas: A review. Gynecol. Oncol. 2010, 116, 131–139. [Google Scholar] [CrossRef]
- Hensley, M.L.; Miller, A.; O’Malley, D.M.; Mannel, R.S.; Behbakht, K.; Bakkum-Gamez, J.N.; Michael, H. Randomized phase III trial of gemcitabine plus docetaxel plus bevacizumab or placebo as first-line treatment for metastatic uterine leiomyosarcoma: An NRG Oncology/Gynecologic Oncology Group study. J. Clin. Oncol. 2015, 33, 1180–1185. [Google Scholar] [CrossRef]
- Sutton, G.P.; Blessing, J.A.; Barrett, R.J.; McGehee, R. Phase II trial of ifosfamide and mesna in leiomyosarcoma of the uterus: A Gynecologic Oncology Group study. Am. J. Obstet. Gynecol. 1992, 166, 556–559. [Google Scholar] [CrossRef]
- Sutton, G.; Blessing, J.; Hanjani, P.; Kramer, P.; Gynecologic Oncology Group. Phase II evaluation of liposomal doxorubicin (Doxil) in recurrent or advanced leiomyosarcoma of the uterus: A Gynecologic Oncology Group study. Gynecol. Oncol. 2005, 96, 749–752. [Google Scholar] [CrossRef]
- Gallup, D.G.; Blessing, J.A.; Andersen, W.; Morgan, M.A.; Gynecologic Oncology Group. Study Evaluation of paclitaxel in previously treated leiomyosarcoma of the uterus: A gynecologic oncology group study. Gynecol. Oncol. 2003, 89, 48–51. [Google Scholar] [CrossRef]
- Rose, P.G.; Blessing, J.A.; Soper, J.T.; Barter, J.F. Prolonged oral etoposide in recurrent or advanced leiomyosarcoma of the uterus: A gynecologic oncology group study. Gynecol. Oncol. 1998, 70, 267–271. [Google Scholar] [CrossRef]
- Thigpen, J.T.; Blessing, J.A.; Wilbanks, G.D. Cisplatin as second-line chemotherapy in the treatment of advanced or recurrent leiomyosarcoma of the uterus. A phase II trial of the Gynecologic Oncology Group. Am. J. Clin. Oncol. 1986, 9, 18–20. [Google Scholar] [CrossRef]
- Duska, L.R.; Blessing, J.A.; Rotmensch, J.; Mannel, R.S.; Hanjani, P.; Rose, P.G.; Dizon, D.S. A Phase II evaluation of ixabepilone (IND #59699, NSC #710428) in the treatment of recurrent or persistent leiomyosarcoma of the uterus: An NRG Oncology/Gynecologic Oncology Group Study. Gynecol. Oncol. 2014, 135, 44–48. [Google Scholar] [Green Version]
- Pautier, P.; Floquet, A.; Gladieff, L.; Bompas, E.; Ray-Coquard, I.; Piperno-Neumann, S.; Selle, F.; Guillemet, C.; Weber, B.; Largillier, R.; et al. A randomized clinical trial of adjuvant chemotherapy with doxorubicin, ifosfamide, and cisplatin followed by radiotherapy versus radiotherapy alone in patients with localized uterine sarcomas (SARCGYN study). A study of the French Sarcoma Group. Ann. Oncol. 2013, 24, 1099–1104. [Google Scholar] [CrossRef]
- Greenhough, A.; Smartt, H.J.M.; Moore, A.E.; Roberts, H.R.; Williams, A.C.; Paraskeva, C.; Kaidi, A. The COX-2/PGE2 pathway: Key roles in the hallmarks of cancer and adaptation to the tumour microenvironment. Carcinogenesis 2009, 30, 377–386. [Google Scholar] [CrossRef]
- Kundu, N.; Ma, X.; Kochel, T.; Goloubeva, O.; Staats, P.; Thompson, K.; Martin, S.; Reader, J.; Take, Y.; Collin, P.; et al. Prostaglandin E receptor EP4 is a therapeutic target in breast cancer cells with stem-like properties. Breast Cancer Res. Treat. 2014, 143, 19–31. [Google Scholar] [CrossRef]
- Majumder, M.; Xin, X.; Liu, L.; Girish, G.V.; Lala, P.K. Prostaglandin E2 receptor EP4 as the common target on cancer cells and macrophages to abolish angiogenesis, lymphangiogenesis, metastasis, and stem-like cell functions. Cancer Sci. 2014, 105, 1142–1151. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, Z.; Ogawa, O.; Yoshikawa, T.; Sakamoto, H.; Shibasaki, N.; Goto, T.; Wang, L.; Terada, N. An EP4 antagonist ONO-AE3-208 suppresses cell invasion, migration, and metastasis of prostate cancer. Cell Biochem. Biophys. 2014, 70, 521–527. [Google Scholar] [CrossRef]
- Ma, X.; Kundu, N.; Rifat, S.; Walser, T.; Fulton, A.M. Prostaglandin E receptor EP4 antagonism inhibits breast cancer metastasis. Cancer Res. 2006, 66, 2923–2927. [Google Scholar] [CrossRef]
- George Paul, A.; Sharma-Walia, N.; Kerur, N.; White, C.; Chandran, B. Piracy of prostaglandin E2/EP receptor-mediated signaling by Kaposi’s sarcoma-associated herpes virus (HHV-8) for latency gene expression: Strategy of a successful pathogen. Cancer Res. 2010, 70, 3697–3708. [Google Scholar] [CrossRef]
- Wang, Z.; He, M.; Xiao, Z.; Wu, H.; Wu, Y. Quantitative assessment of the association of COX-2 (Cyclooxygenase-2) immunoexpression with prognosis in human osteosarcoma: A meta-analysis. PLoS ONE 2013, 8, e82907. [Google Scholar] [CrossRef]
- Menczer, J.; Schreiber, L.; Berger, E.; Levy, T. The Effect of Cyclooxygenase-2 Expression in Uterine Carcinosarcoma on Survival: A Reassessment Based on Mature Data. Int. J. Gynecol. Cancer 2015, 25, 1415–1417. [Google Scholar] [CrossRef]
- Guo, X.; Jo, V.Y.; Mills, A.M.; Zhu, S.X.; Lee, C.-H.; Espinosa, I.; Nucci, M.R.; Varma, S.; Forgó, E.; Hastie, T.; et al. Clinically Relevant Molecular Subtypes in Leiomyosarcoma. Clin. Cancer Res. 2015, 21, 3501–3511. [Google Scholar] [CrossRef]
- An, Y.; Wang, S.; Li, S.; Zhang, L.; Wang, D.; Wang, H.; Zhu, S.; Zhu, W.; Li, Y.; Chen, W.; et al. Distinct molecular subtypes of uterine leiomyosarcoma respond differently to chemotherapy treatment. BMC Cancer 2017, 17, 639. [Google Scholar] [CrossRef]
- Goldman, M.; Craft, B.; Hastie, M.; Repecka, K.; Kamath, A.; McDade, F.; Rogers, D.; Brooks, A.; Zhu, J.; Haussler, D. The UCSC Xena platform for public and private cancer genomics data visualization and interpretation. Biorxiv 2019, 326470. [Google Scholar] [CrossRef]
- Hosh, M.; Antar, S.; Nazzal, A.; Warda, M.; Gibreel, A.; Refky, B. Uterine Sarcoma: Analysis of 13,089 Cases Based on Surveillance, Epidemiology, and End Results Database. Int. J. Gynecol. Cancer 2016, 26, 1098–1104. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.-H.; Kim, J.-Y.; Kim, Y.-S.; Song, H.-J.; Song, K.-J.; Song, J.-W.; Baek, L.-J.; Seo, E.-Y.; Kim, C.-D.; Lee, J.-H.; et al. Upregulation of Class II β-Tubulin Expression in Differentiating Keratinocytes. J. Investig. Dermatol. 2005, 124, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, E.; Inoue, S.; Mizushima, T.; Chen, J.; Ide, H.; Kawahara, T.; Reis, L.O.; Baras, A.S.; Netto, G.J.; Miyamoto, H. Prostaglandin receptors induce urothelial tumourigenesis as well as bladder cancer progression and cisplatin resistance presumably via modulating PTEN expression. Br. J. Cancer 2018, 118, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.; Matos, T.; Charytonowicz, E.; Hricik, T.; Castillo-Martin, M.; Remotti, F.; Lee, F.Y.; Matushansky, I. Characterization and comparison of the properties of sarcoma cell lines in vitro and in vivo. Hum. Cell 2009, 22, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Şahin, H.; Karatas, F.; Coban, G.; Özen, Ö.; Erdem, Ö.; Onan, M.A.; Ayhan, A. Uterine smooth muscle tumor of uncertain malignant potential: Fertility and clinical outcomes. J. Gynecol. Oncol. 2019, 19, 288–295. [Google Scholar] [CrossRef]
- Devereaux, K.A.; Schoolmeester, J.K. Smooth Muscle Tumors of the Female Genital Tract. Surg. Pathol. Clin. 2019, 12, 397–455. [Google Scholar] [CrossRef]
- In, G.K.; Hu, J.S.; Tseng, W.W. Treatment of advanced, metastatic soft tissue sarcoma: Latest evidence and clinical considerations. Ther. Adv. Med. Oncol. 2017, 9, 533–550. [Google Scholar] [CrossRef]
- Baird, D.D.; Dunson, D.B.; Hill, M.C.; Cousins, D.; Schectman, J.M. High cumulative incidence of uterine leiomyoma in black and white women: Ultrasound evidence. Am. J. Obstet. Gynecol. 2003, 188, 100–107. [Google Scholar] [CrossRef]
- Lin, J.; Song, X.; Liu, C. Pelvic intravascular leiomyomatosis associated with benign pulmonary metastasizing leiomyoma: Clinicopathologic, clonality, and copy number variance analysis. Int. J. Gynecol. Pathol. 2014, 33, 140–145. [Google Scholar] [CrossRef]
- Ip, P.P.C.; Tse, K.Y.; Tam, K.F. Uterine smooth muscle tumors other than the ordinary leiomyomas and leiomyosarcomas: A review of selected variants with emphasis on recent advances and unusual morphology that may cause concern for malignancy. Adv. Anat. Pathol. 2010, 17, 91–112. [Google Scholar] [CrossRef] [PubMed]
- Ganjoo, K.N. Uterine sarcomas. Curr. Probl. Cancer 2019, 116, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Hensley, M.L.; Blessing, J.A.; Mannel, R.; Rose, P.G. Fixed-dose rate gemcitabine plus docetaxel as first-line therapy for metastatic uterine leiomyosarcoma: A Gynecologic Oncology Group phase II trial. Gynecol. Oncol. 2008, 109, 329–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensley, M.L.; Ishill, N.; Soslow, R.; Larkin, J.; Abu-Rustum, N.; Sabbatini, P.; Konner, J.; Tew, W.; Spriggs, D.; Aghajanian, C.A. Adjuvant gemcitabine plus docetaxel for completely resected stages I-IV high grade uterine leiomyosarcoma: Results of a prospective study. Gynecol. Oncol. 2009, 112, 563–567. [Google Scholar] [CrossRef]
- Reyners, A.K.L.; de Munck, L.; Erdkamp, F.L.G.; Smit, W.M.; Hoekman, K.; Lalisang, R.I.; de Graaf, H.; Wymenga, A.N.M.; Polee, M.; Hollema, H.; et al. A randomized phase II study investigating the addition of the specific COX-2 inhibitor celecoxib to docetaxel plus carboplatin as first-line chemotherapy for stage IC to IV epithelial ovarian cancer, Fallopian tube or primary peritoneal carcinomas: The DoCaCel study. Ann. Oncol. ESMO 2012, 23, 2896–2902. [Google Scholar]
- Ma, X.; Kundu, N.; Ioffe, O.B.; Goloubeva, O.; Konger, R.; Baquet, C.; Gimotty, P.; Reader, J.; Fulton, A.M. Prostaglandin E receptor EP1 suppresses breast cancer metastasis and is linked to survival differences and cancer disparities. Mol. Cancer Res. MCR 2010, 8, 1310–1318. [Google Scholar] [CrossRef]
- Bresalier, R.S.; Sandler, R.S.; Quan, H.; Bolognese, J.A.; Oxenius, B.; Horgan, K.; Lines, C.; Riddell, R.; Morton, D.; Lanas, A.; et al. Cardiovascular events associated with rofecoxib in a colorectal adenoma chemoprevention trial. N. Engl. J. Med. 2005, 352, 1092–1102. [Google Scholar] [CrossRef]
- Chell, S.D.; Witherden, I.R.; Dobson, R.R.; Moorghen, M.; Herman, A.A.; Qualtrough, D.; Williams, A.C.; Paraskeva, C. Increased EP4 receptor expression in colorectal cancer progression promotes cell growth and anchorage independence. Cancer Res. 2006, 66, 3106–3113. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Frilot, N.; Kim, J.I.; Kim, W.-J.; Daaka, Y. Prostaglandin E2 regulates renal cell carcinoma invasion through the EP4 receptor-Rap GTPase signal transduction pathway. J. Biol. Chem. 2011, 286, 33954–33962. [Google Scholar] [CrossRef]
- Terada, N.; Shimizu, Y.; Kamba, T.; Inoue, T.; Maeno, A.; Kobayashi, T.; Nakamura, E.; Kamoto, T.; Kanaji, T.; Maruyama, T.; et al. Identification of EP4 as a potential target for the treatment of castration-resistant prostate cancer using a novel xenograft model. Cancer Res. 2010, 70, 1606–1615. [Google Scholar] [CrossRef]
- Reader, J.; Holt, D.; Fulton, A. Prostaglandin E2 EP receptors as therapeutic targets in breast cancer. Cancer Metastasis Rev. 2011, 30, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, M.; Peri, K.; Ribeiro-da-Silva, A.; Almazan, G.; Shichi, H.; Hou, X.; Varma, D.R.; Chemtob, S. Localization of functional prostaglandin E2 receptors EP3 and EP4 in the nuclear envelope. J. Biol. Chem. 1999, 274, 15719–15724. [Google Scholar] [CrossRef] [PubMed]
- Bhooshan, N.; Staats, P.N.; Fulton, A.M.; Feliciano, J.L.; Edelman, M.J. Prostaglandin E Receptor EP4 expression, survival and pattern of recurrence in locally advanced NSCLC. Lung Cancer Amst. Neth. 2016, 101, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Von der Emde, L.; Goltz, D.; Latz, S.; Müller, S.C.; Kristiansen, G.; Ellinger, J.; Syring, I. Prostaglandin receptors EP1-4 as a potential marker for clinical outcome in urothelial bladder cancer. Am. J. Cancer Res. 2014, 4, 952–962. [Google Scholar] [PubMed]
- Gemcitabine Hydrochloride and Docetaxel Followed by Doxorubicin Hydrochloride or Observation in Treating Patients with High-Risk Uterine Leiomyosarcoma Previously Removed by Surgery; Gynecologic Oncology Group, 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT01533207 (accessed on 13 October 2018).
- Lin, M.-C.; Chen, S.-Y.; He, P.-L.; Herschman, H.; Li, H.-J. PGE2 /EP4 antagonism enhances tumor chemosensitivity by inducing extracellular vesicle-mediated clearance of cancer stem cells. Int. J. Cancer 2018, 143, 1440–1455. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Poorly Differentiated LMS Subtype I: n = 22 | Conventional LMS Subtype II: n = 32 | Tests *, p-Value | |
|---|---|---|---|---|
| LMOD1 | 0.0002 ** | |||
| Median | 3.6 | 6.5 | ||
| Range | 8.3 | 10.4 | ||
| Mean | 3.3 | 5.9 | ||
| SD | 2.4 | 2.2 | ||
| ARL4C | 0.01 ** | |||
| Median | −0.003 | −0.7 | ||
| Range | 6.1 | 5.8 | ||
| Mean | 0.4 | −0.6 | ||
| SD | 1.6 | 1.3 | ||
| TUBB3 (β-3 Tubulin) | 0.0001 ** | |||
| Median | 1 | −0.6 | ||
| Range | 4.7 | 4.3 | ||
| Mean | 0.9 | −0.6 | ||
| SD | 1.4 | 1.2 | ||
| PTGER4 (EP4) | 0.14 | |||
| Median | 1.7 | 2.8 | ||
| Range | 5.6 | 6.6 | ||
| Mean | 1.5 | 2.2 | ||
| SD | 1.3 | 1.9 | ||
| Average Age (Years ± SD) | All | 52.18 ± 13.69 | |
|---|---|---|---|
| Leiomyosarcoma | 59.2 ± 13.46 | ||
| n | % | ||
| Race | White | 12 | 41.4 |
| Black | 16 | 55.2 | |
| Hispanic | 1 | 3.4 | |
| Smooth Muscle Type (n = 29) | Leiomyoma | 11 | 37.9 |
| Leiomyomatosis | 3 | 10.3 | |
| STUMP | 5 | 17.2 | |
| Leiomyosarcoma | 10 | 34.5 | |
| Leiomyosarcoma (n = 10) | Stage | ||
| IA | 1 | 10 | |
| IIIA | 1 | 10 | |
| IIIC | 2 | 20 | |
| IVB | 6 | 60 | |
| Surgery | |||
| TAH/BSO ± LND | 6 | 60 | |
| Other | 4 | 40 | |
| Residual Disease | 2 | 20 | |
| Adjuvant Therapy | 6 | 60 | |
| Dead of Disease | 4 | 40 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reader, J.; Harper, A.K.; Legesse, T.; Staats, P.N.; Goloubeva, O.; Rao, G.G.; Fulton, A.; Roque, D.M. EP4 and Class III β-Tubulin Expression in Uterine Smooth Muscle Tumors: Implications for Prognosis and Treatment. Cancers 2019, 11, 1590. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101590
Reader J, Harper AK, Legesse T, Staats PN, Goloubeva O, Rao GG, Fulton A, Roque DM. EP4 and Class III β-Tubulin Expression in Uterine Smooth Muscle Tumors: Implications for Prognosis and Treatment. Cancers. 2019; 11(10):1590. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101590
Chicago/Turabian StyleReader, Jocelyn, Amy K. Harper, Teklu Legesse, Paul N. Staats, Olga Goloubeva, Gautam G. Rao, Amy Fulton, and Dana M. Roque. 2019. "EP4 and Class III β-Tubulin Expression in Uterine Smooth Muscle Tumors: Implications for Prognosis and Treatment" Cancers 11, no. 10: 1590. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11101590