MicroRNA-9-5p-CDX2 Axis: A Useful Prognostic Biomarker for Patients with Stage II/III Colorectal Cancer

 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
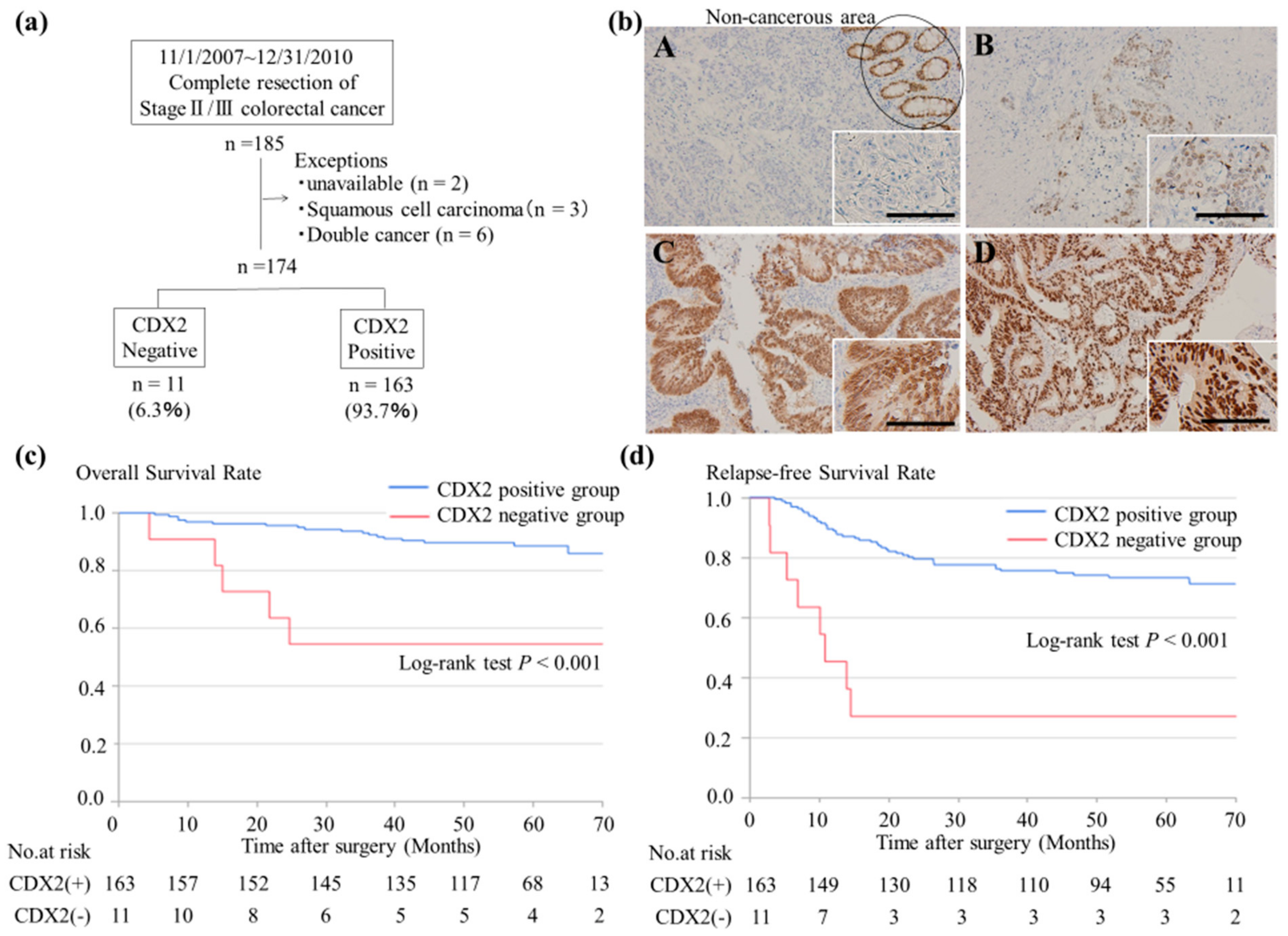
2.1. Lack of CDX2 Expression is Associated with Poor Prognosis of Stage II/ III CRC
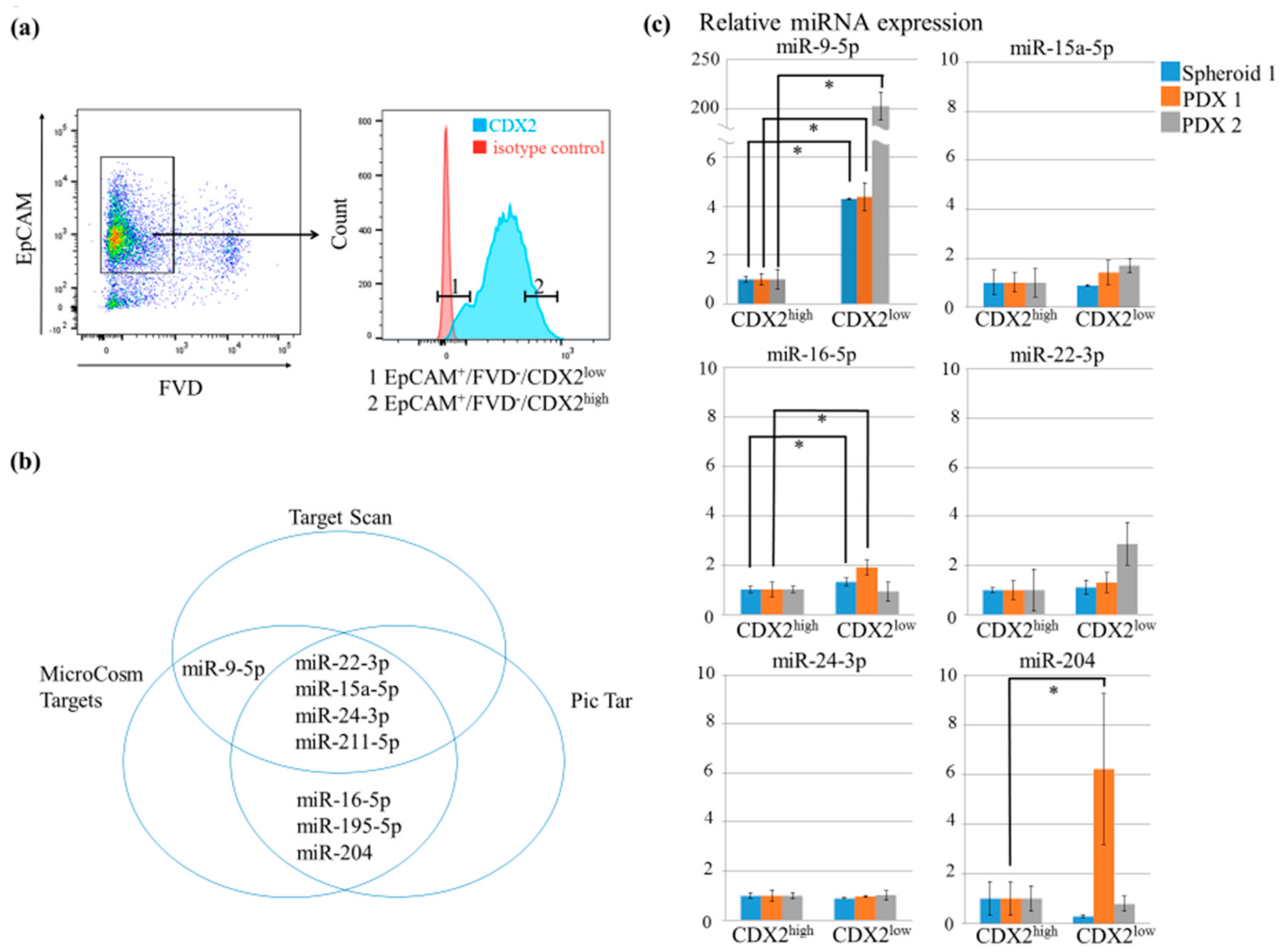
2.2. MiR-9-5p was Upregulated in CDX2low Tumor Samples
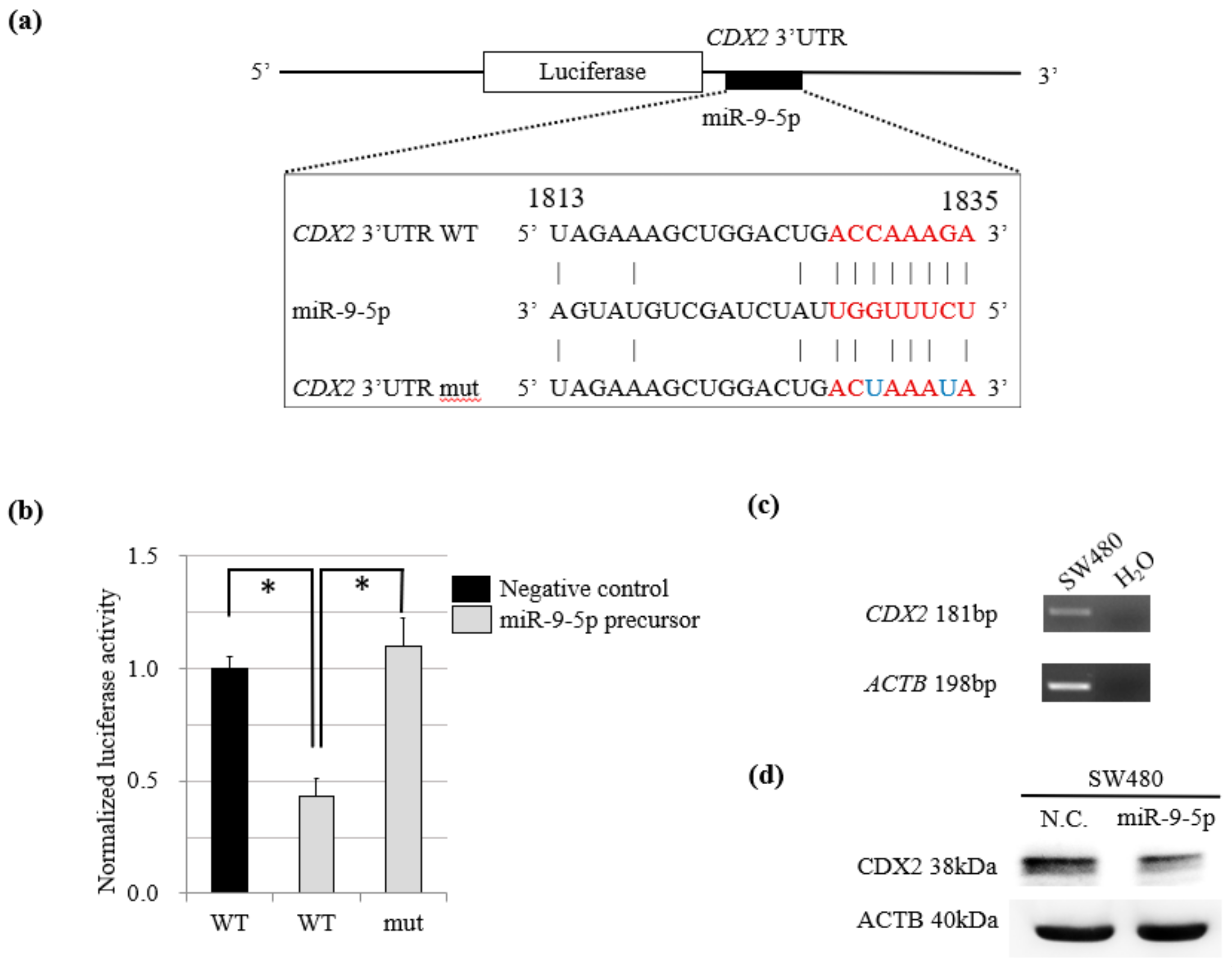
2.3. MiR-9-5p Targets CDX2
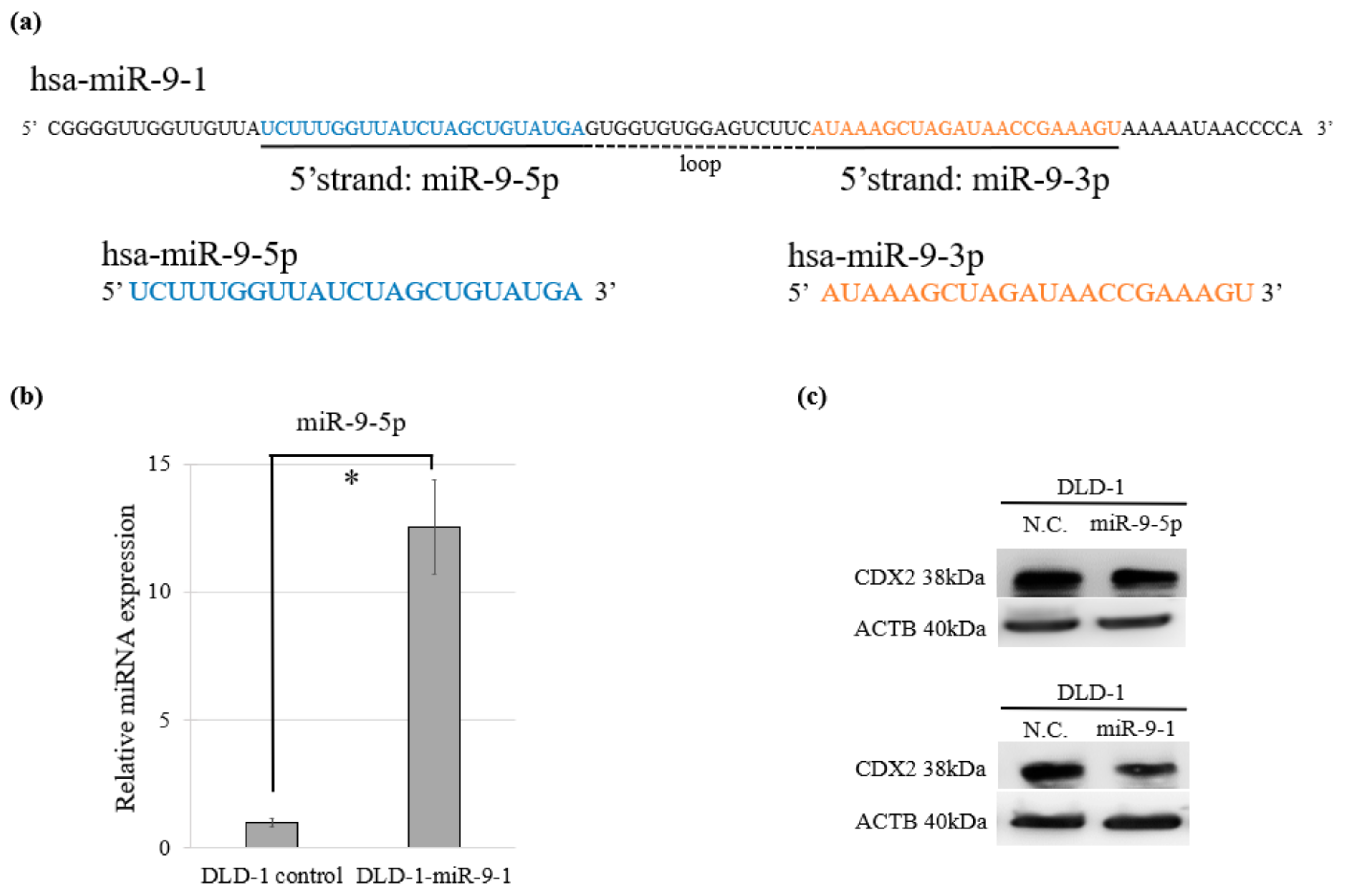
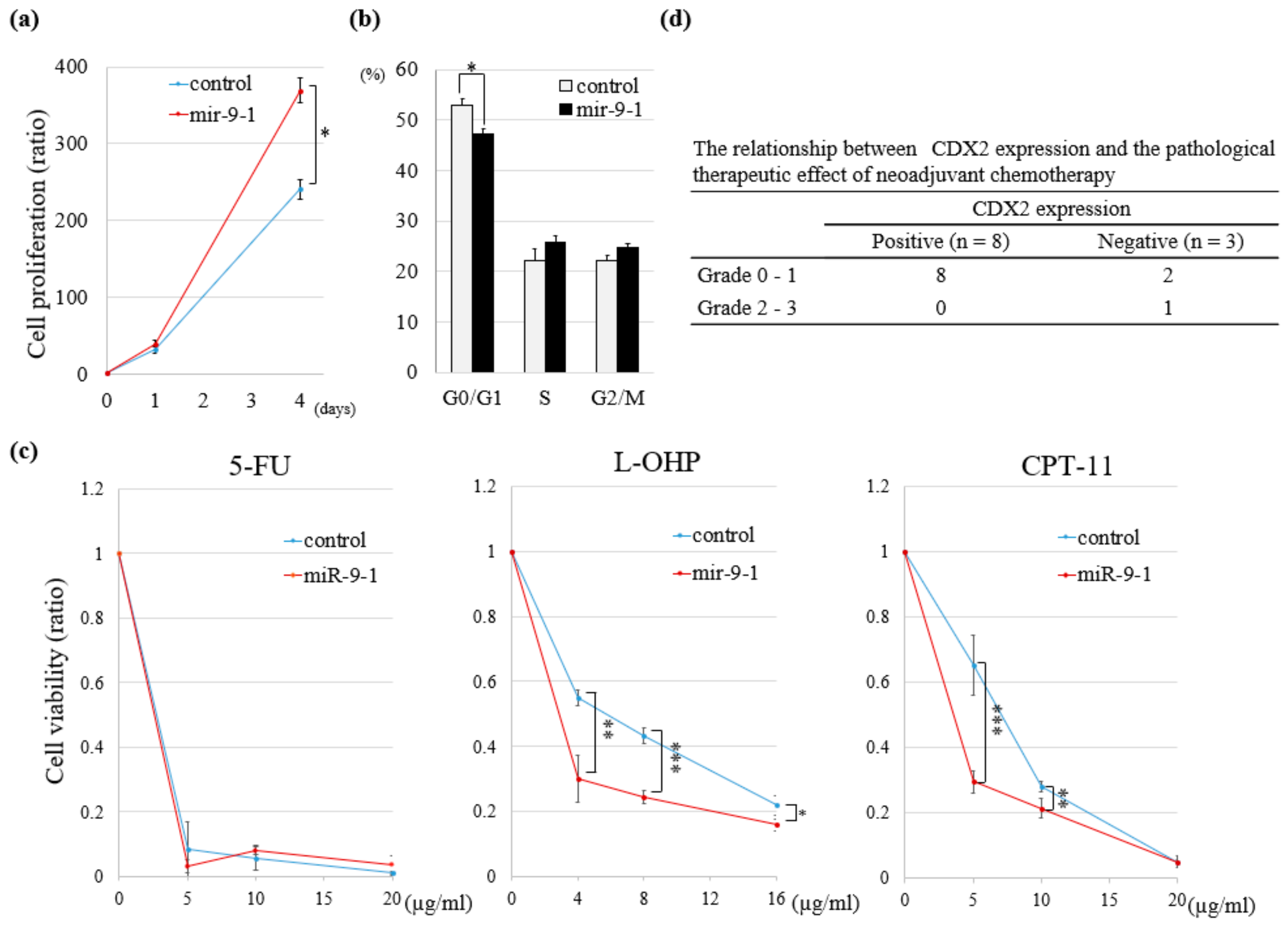
2.4. Overexpression of miR-9-1 Accelerates Cell Cycle and Cell Proliferation and Increases Sensitivity to Anti-Cancer Drugs
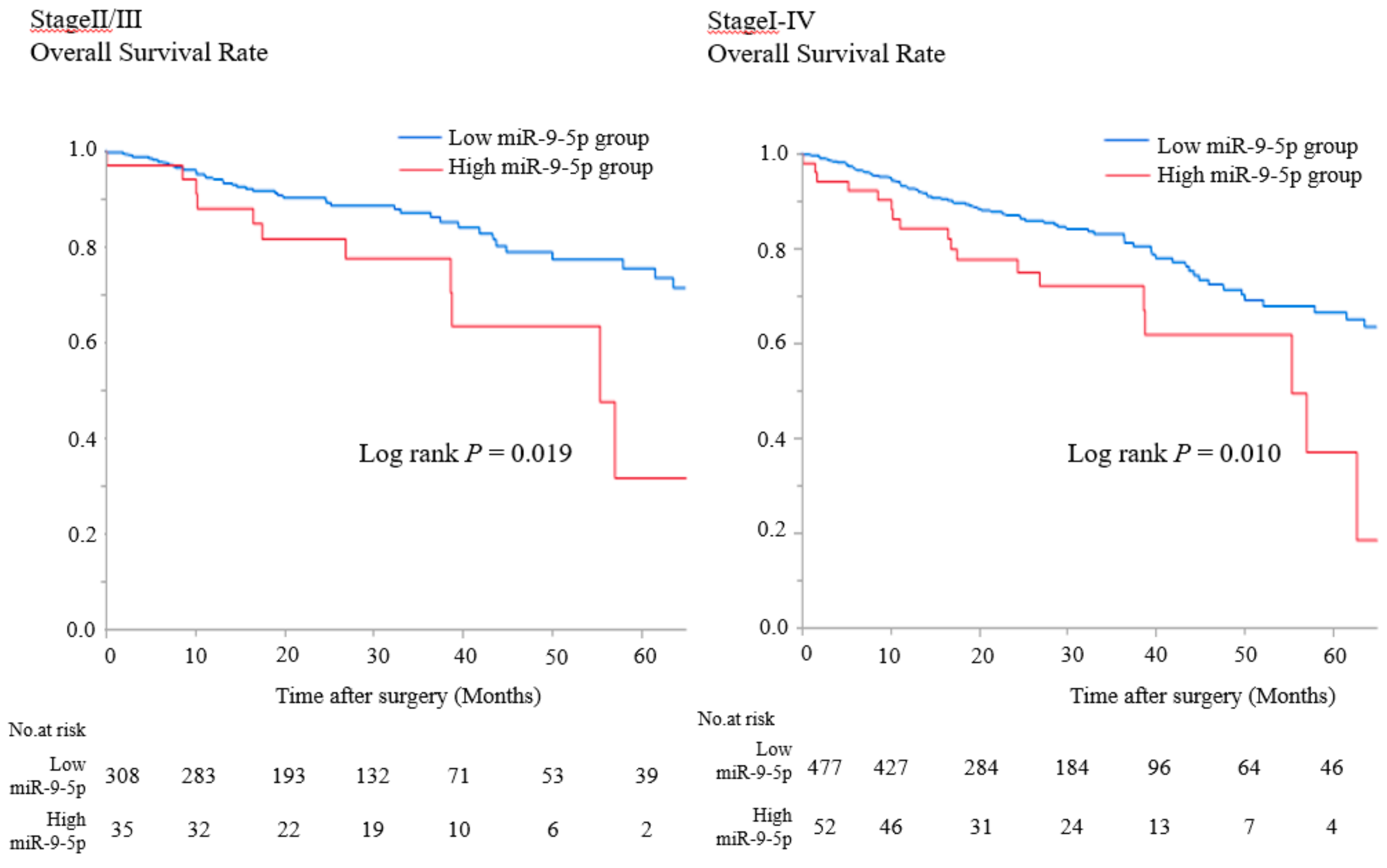
2.5. MiR-9-5p Expression in Patients with CRC Correlates with Poor OS
3. Discussion
4. Materials and Methods
4.1. Patient Population
4.2. Immunohistochemistry (IHC)
4.3. Data Analysis from TCGA-Colorectal Cancer (TCGA-CRC)
4.4. Statistical Analysis
4.5. Cell Lines and Cell Culture
4.6. Patient-Derived Tumor Xenografts (PDX) and Patient-Derived Cancer-Spheroids
4.7. Flow Cytometry
4.8. In-Silico Prediction of MiRs Targeting the CDX2 3′-UTR
4.9. Reverse-Transcriptase Quantitative PCR (RT-qPCR)
4.10. Plasmid Vectors for Luciferase Reporter Assays and Mutagenesis
4.11. Luciferase Reporter Assay
4.12. Western Blotting
4.13. Cell Proliferation, Cell Cycle, and Cytotoxicity Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Boland, G.M.; Chang, G.J.; Haynes, A.B.; Chiang, Y.J.; Chagpar, R.; Xing, Y.; Hu, C.Y.; Feig, B.W.; You, Y.N.; Cormier, J.N. Association between adherence to National Comprehensive Cancer Network Treatment Guidelines and improved survival in patients with colon cancer. Cancer 2013, 119, 1593–1601. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B.; Schrag, D.; Somerfield, M.R.; Cohen, A.M.; Figueredo, A.T.; Flynn, P.J.; Krzyzanowska, M.K.; Maroun, J.; McAllister, P.; Van Cutsem, E.; et al. American Society of Clinical Oncology recommendations on adjuvant chemotherapy for stage II colon cancer. J. Clin. Oncol. 2004, 22, 3408–3419. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B.; Venook, A.P.; Al-Hawary, M.M.; Cederquist, L.; Chen, Y.J.; Ciombor, K.K.; Cohen, S.; Cooper, H.S.; Deming, D.; Engstrom, P.F.; et al. NCCN Guidelines® Insights Colon Cancer, Version 2.2018 Featured Updates to the NCCN Guidelines. J. Natl. Compr. Cancer Netw. 2018, 16, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Schmoll, H.J.; Van Cutsem, E.; Stein, A.; Valentini, V.; Glimelius, B.; Haustermans, K.; Nordlinger, B.; van de Velde, C.J.; Balmana, J.; Regula, J.; et al. Esmo consensus guidelines for management of patients with colon and rectal cancer. A personalized approach to clinical decision making. Ann. Oncol. 2012, 23, 2479–2516. [Google Scholar] [CrossRef] [PubMed]
- Kannarkatt, J.; Joseph, J.; Kurniali, P.C.; Al-Janadi, A.; Hrinczenko, B. Adjuvant chemotherapy for stage II colon cancer: A clinical dilemma. J. Oncol. Pract. 2017, 13, 233–241. [Google Scholar] [CrossRef]
- André, T.; Boni, C.; Mounedji-Boudiaf, L.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Zaninelli, M.; Clingan, P.; Bridgewater, J.; et al. Oxaliplatin, fluorouracil, and leucovorin as adjuvant treatment for colon cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar] [CrossRef]
- Haller, D.G.; Tabernero, J.; Maroun, J.; de Braud, F.; Price, T.; Van Cutsem, E.; Hill, M.; Gilberg, F.; Rittweger, K.; Schmoll, H.J. Capecitabine plus oxaliplatin compared with fluorouracil and folinic acid as adjuvant therapy for stage III colon cancer. J. Clin. Oncol. 2011, 29, 1465–1471. [Google Scholar] [CrossRef]
- Yothers, G.; O’Connell, M.J.; Allegra, C.J.; Kuebler, J.P.; Colangelo, L.H.; Petrelli, N.J.; Wolmark, N. Oxaliplatin as adjuvant therapy for colon cancer: Updated results of NSABP C-07 trial, including survival and subset analyses. J. Clin. Oncol. 2011, 29, 3768–3774. [Google Scholar] [CrossRef]
- Tomasello, G.; Barni, S.; Turati, L.; Ghidini, M.; Pezzica, E.; Passalacqua, R.; Petrelli, F. Association of CDX2 expression with survival in early colorectal cancer: A systematic review and meta-analysis. Clin. Colorectal Cancer 2018, 17, 97–103. [Google Scholar] [CrossRef]
- Olsen, J.; Espersen, M.L.M.; Jess, P.; Kirkeby, L.T.; Troelsen, J.T. The clinical perspectives of CDX2 expression in colorectal cancer: A qualitative systematic review. Surg. Oncol. 2014, 23, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Bonhomme, C.; Duluc, I.; Martin, E.; Chawengsaksophak, K.; Chenard, M.P.; Kedinger, M.; Beck, F.; Freund, J.N.; Domon-Dell, C. The Cdx2 homeobox gene has a tumour suppressor function in the distal colon in addition to a homeotic role during gut development. Gut 2003, 52, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Tamai, Y.; Horiike, S.; Oshima, M.; Taketo, M.M. Colonic polyposis caused by mTOR-mediated compound mutant mice. Nat. Genet. 2003, 35, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Beck, F. The role of Cdx genes in the mammalian gut. Gut 2004, 53, 1394–1396. [Google Scholar] [CrossRef]
- Ee, H.C.; Erler, T.; Bhathal, P.S.; Young, G.P.; James, R.J. Cdx-2 Homeodomain protein expression in human and rat colorectal adenoma and carcinoma. Am. J. Pathol. 1995, 147, 586–592. [Google Scholar]
- Bou, G.; Liu, S.; Sun, M.; Zhu, J.; Xue, B.; Guo, J.; Zhao, Y.; Qu, B.; Weng, X.; Wei, Y.; et al. CDX2 is essential for cell proliferation and polarity in porcine blastocysts. Development 2017, 144, 1296–1306. [Google Scholar] [CrossRef]
- Woodford-Richens, K.L.; Halford, S.; Rowan, A.; Bevan, S.; Aaltonen, L.A.; Wasan, H.; Bicknell, D.; Bodmer, W.F.; Houlston, R.S.; Tomlinson, I.P. CDX2 mutations do not account for juvenile polyposis or Peutz-Jeghers syndrome and occur infrequently in sporadic colorectal cancers. Br. J. Cancer 2001, 84, 1314–1316. [Google Scholar] [CrossRef]
- Da Costa, L.T.; He, T.C.; Yu, J.; Sparks, A.B.; Morin, P.J.; Polyak, K.; Laken, S.; Vogelstein, B.; Kinzler, K.W. CDX2 is mutated in a colorectal cancer with normal APC/β-catenin signaling. Oncogene 1999, 18, 5010–5014. [Google Scholar] [CrossRef]
- Yagi, O.; Akiyama, Y.; Yuasa, Y. Genomic structure and alterations of homeobox gene CDX2 in colorectal carcinomas. Br. J. Cancer 1999, 79, 440–444. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNA target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Tsuchiya, N.; Izumiya, M.; Ogata-Kawata, H.; Okamoto, K.; Fujiwara, Y.; Nakai, M.; Okabe, A.; Schetter, A.J.; Bowman, E.D.; Midorikawa, Y.; et al. Tumor suppressor miR-22 determines p53-dependent cellular fate through post-transcriptional regulation of p21. Cancer Res. 2011, 71, 4628–4639. [Google Scholar] [CrossRef] [PubMed]
- Nosho, K.; Igarashi, H.; Nojima, M.; Ito, M.; Maruyama, R.; Yoshii, S.; Naito, T.; Sukawa, Y.; Mikami, M.; Sumioka, W.; et al. Association of MicroRNA-31 with BRAF mutation, colorectal cancer survival and serrated pathway. Carcinogenesis 2014, 35, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Hisamori, S.; Oshima, N.; Sato, F.; Shimono, Y. miR-137 regulates the tumorigenicity of colon cancer stem cells through the inhibition of DCLK1. Mol. Cancer Res. 2016, 137, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Nowek, K.; Wiemer, E.A.C.; Jongen-Lavrencic, M. The versatile nature of miR-9/9* in human cancer. Oncotarget 2018, 9, 20838–20854. [Google Scholar] [CrossRef]
- Dalerba, P.; Sahoo, D.; Paik, S.; Guo, X.; Yothers, G.; Song, N.; Wilcox-Fogel, N.; Forgó, E.; Rajendran, P.S.; Miranda, S.P.; et al. CDX2 as a prognostic biomarker in stage II and stage III colon cancer. N. Engl. J. Med. 2016, 374, 211–222. [Google Scholar] [CrossRef]
- Baba, Y.; Nosho, K.; Shima, K.; Freed, E.; Irahara, N.; Philips, J.; Meyerhardt, J.A.; Hornick, J.L.; Shivdasani, R.A.; Fuchs, C.S.; et al. Relationship of CDX2 loss with molecular features and prognosis in colorectal cancer. Clin. Cancer Res. 2009, 15, 4665–4673. [Google Scholar] [CrossRef]
- Zlobec, I.; Bihl, M.P.; Schwarb, H.; Terracciano, L.; Lugli, A. Clinicopathological and protein characterization of BRAF- and K-RAS-mutated colorectal cancer and implications for prognosis. Int. J. Cancer 2010, 127, 367–380. [Google Scholar]
- Dawson, H.; Galván, J.A.; Helbling, M.; Muller, D.E.; Karamitopoulou, E.; Koelzer, V.H.; Economou, M.; Hammer, C.; Lugli, A.; Zlobec, I. Possible role of Cdx2 in the serrated pathway of colorectal cancer characterized by BRAF mutation, high-level CpG island methylator phenotype and mismatch repair-deficiency. Int. J. Cancer 2014, 134, 2342–2351. [Google Scholar] [CrossRef]
- Bae, J.M.; Lee, T.H.; Cho, N.; Kim, T.; Kang, G.H. Loss of CDX2 expression is associated with poor prognosis in colorectal cancer patients. World J. Gastroenterol. 2015, 21, 1457–1467. [Google Scholar] [CrossRef]
- Coolen, M.; Katz, S.; Bally-Cuif, L. miR-9: A versatile regulator of neurogenesis. Front. Cell Neurosci. 2013, 7, 1–11. [Google Scholar] [CrossRef]
- Leucci, E.; Zriwil, A.; Gregersen, L.H.; Jensen, K.T.; Obad, S.; Bellan, C.; Leoncini, L.; Kauppinen, S.; Lund, A.H. Inhibition of miR-9 de-represses HuR and DICER1 and impairs Hodgkin lymphoma tumour outgrowth in vivo. Oncogene 2012, 31, 5081–5089. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Young, J.; Prabhala, H.; Pan, E.; Mestdagh, P.; Muth, D.; Teruya-Feldstein, J.; Reinhardt, F.; Onder, T.T.; Valastyan, S.; et al. miR-9, a MYC/MYCN-activated microRNA, regulates E-cadherin and cancer metastasis. Nat. Cell. Biol. 2010, 12, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Wilting, S.M.; Snijders, P.J.F.; Verlaat, W.; Jaspers, A.; van de Wiel, M.A.; van Wieringen, W.N.; Meijer, G.A.; Kenter, G.G.; Yi, Y.; le Sage, C.; et al. Altered microRNA expression associated with chromosomal changes contributes to cervical carcinogenesis. Oncogene 2013, 32, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.H.; Huang, C.C.; Pan, M.R.; Chen, H.H.; Hung, W.C. Prospero homeobox 1 promotes epithelial-mesenchymal transition in colon cancer cells by inhibiting e-cadherin via miR-9. Clin. Cancer Res. 2012, 18, 6416–6425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotkrua, P.; Akiyama, Y.; Hashimoto, Y.; Otsubo, T.; Yuasa, Y. MiR-9 Downregulates CDX2 expression in gastric cancer cells. Int. J. Cancer 2011, 129, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Kerschner, J.L.; Harris, A. Transcriptional networks driving enhancer function in the CFTR gene. Biochem. J. 2012, 446, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Koh, I.; Hinoi, T.; Sentani, K.; Hirata, E.; Nosaka, S.; Niitsu, H.; Miguchi, M.; Adachi, T.; Yasui, W.; Ohdan, H.; et al. Regulation of multidrug resistance 1 expression by CDX2 in ovarian mucinous adenocarcinoma. Cancer Med. 2016, 5, 1546–1555. [Google Scholar] [CrossRef]
- Bruun, J.; Sveen, A.; Barros, R.; Eide, P.W.; Eilertsen, I.; Kolberg, M.; Pellinen, T.; David, L.; Svindland, A.; Kallioniemi, O.; et al. Prognostic, predictive, and pharmacogenomic assessments of CDX2 refine stratification of colorectal cancer. Mol. Oncol. 2018, 12, 1639–1655. [Google Scholar] [CrossRef]
- Japanese Society for Cancer of the Colon and Rectum. Japanese Classification of Colorectal Carcinoma, 8th ed.; Kanehara & Co., Ltd.: Tokyo, Japan, 2013; p. 34. [Google Scholar]
- The R Project for Statistical Computing. Available online: http://r-project.org/ (accessed on 20 September 2018).
- Hothorn, T.; Hornik, K.; Zeileis, A. Unbiased recursive partitioning: A conditional inference framework. J. Comput. Graph. Stat. 2006, 15, 651–674. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. stringr: Simple, Consistent Wrappers for Common String Operations. Available online: https://stringr.tidyverse.org/ (accessed on 20 September 2018).
- Therneau, T.M. A Package for Survival Analysis in S. Available online: https://www.mayo.edu/research/documents/tr53pdf/doc-10027379 (accessed on 20 September 2018).
- Wickham, H. tidyverse: Easily Install and Load the “Tidyverse”. Available online: https://tidyverse.tidyverse.org/ (accessed on 20 September 2018).
- Miyoshi, H.; Maekawa, H.; Kakizaki, F.; Yamaura, T.; Kawada, K.; Sakai, Y.; Taketo, M.M. An improved method for culturing patient-derived colorectal cancer spheroids. Oncotarget 2018, 9, 21950–21964. [Google Scholar] [CrossRef] [Green Version]
- Peltier, H.J.; Latham, G.J. Normalization of microRNA expression levels in quantitative RT-PCR assays: Identification of suitable reference RNA targets in normal and cancerous human solid tissues. RNA 2008, 14, 844–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimono, Y.; Zabala, M.; Cho, R.W.; Lobo, N.; Dalerba, P.; Qian, D.; Diehn, M.; Liu, H.; Panula, S.P.; Chiao, E.; et al. Downregulation of miRNA-200c links breast cancer stem cells with normal stem cells. Cell 2009, 138, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | No. of Cases | CDX2 Expression | p-Value | ||
|---|---|---|---|---|---|
| Positive (n = 163) | Negative (n = 11) | ||||
| n (%) | n (%) | ||||
| Sex | Male | 102 | 97 (55.8) | 5 (2.9) | 0.364 |
| Female | 72 | 66 (37.9) | 6 (3.5) | ||
| Age (years) | <70 | 80 | 77(44.3) | 3(1.7) | 0.189 |
| ≥70 | 94 | 86(49.4) | 8(4.6) | ||
| Location | Right colon | 52 | 44 (25.3) | 8 (4.6) | <0.001 |
| Left colon | 73 | 73 (42.0) | 0 (0.0) | ||
| Rectum | 49 | 46 (26.4) | 3 (1.7) | ||
| T-factor | T 1–3 | 119 | 111 (63.8) | 8 (4.6) | 0.746 |
| T 4 | 55 | 52 (29.9) | 3 (1.7) | ||
| N-factor | N 0 | 95 | 91 (52.3) | 4 (2.3) | 0.201 |
| N + | 79 | 72 (41.4) | 7 (4.2) | ||
| Histology | Wel/Mod | 163 | 158 (90.8) | 5 (2.9) | <0.001 |
| Por/Sig/Muc | 11 | 5 (2.9) | 6 (3.5) | ||
| Lymphatic invasion | Ly 0 | 114 | 109 (62.6) | 5 (2.9) | 0.159 |
| Ly + | 60 | 54 (31.0) | 6 (3.5) | ||
| Venous invasion | V 0 | 68 | 65 (37.4) | 3 (1.7) | 0.396 |
| V + | 106 | 98 (56.3) | 8 (4.6) | ||
| Adjuvant chemotherapy | Yes | 89 | 82 (47.1) | 7 (4.0) | 0.389 |
| No | 85 | 81 (46.6) | 4 (2.3) | ||
| Factor | Category | No. of Cases | Univariate Analysis | Multivariate Analysis | ||||
|---|---|---|---|---|---|---|---|---|
| HR | 95 % CI | p-Value | HR | 95 % CI | p-Value | |||
| CDX2 expression | Positive | 163 | Ref | <0.001 | Ref | 0.014 | ||
| Negative | 11 | 5.17 | 2.24–10.47 | 4.33 | 1.37–12.3 | |||
| Sex | Male | 102 | Ref | 0.488 | Ref | 0.439 | ||
| Female | 72 | 0.82 | 0.46–1.43 | 0.79 | 0.43–1.42 | |||
| Age (years) | <70 | 80 | Ref | 0.954 | Ref | 0.705 | ||
| ≥70 | 94 | 1.02 | 0.59–1.77 | 1.12 | 0.70–2.33 | |||
| Location | Right colon | 52 | Ref | Ref | ||||
| Left colon | 73 | 0.91 | 0.47–1.81 | 0.784 | 1.37 | 0.65–3.08 | 0.416 | |
| Rectum | 49 | 1.07 | 0.53–2.19 | 0.850 | 1.60 | 0.71–3.75 | 0.260 | |
| T-factor | T 1–3 | 119 | Ref | 0.064 | Ref | 0.037 | ||
| T 4 | 55 | 1.70 | 0.96–2.95 | 1.87 | 1.04–3.30 | |||
| N-factor | N 0 | 95 | Ref | 0.091 | Ref | 0.109 | ||
| N + | 79 | 1.61 | 0.93–2.81 | 1.58 | 0.90–2.80 | |||
| Histology | Wel/Mod | 163 | Ref | <0.001 | Ref | 0.078 | ||
| Por/Sig/Muc | 11 | 4.95 | 2.15–10.05 | 2.61 | 0.89–7.01 | |||
| Lymphatic invasion | Ly 0 | 114 | Ref | 0.561 | ||||
| Ly + | 60 | 0.84 | 0.45–1.49 | |||||
| Venous invasion | V 0 | 68 | Ref | 0.199 | ||||
| V + | 106 | 1.46 | 0.82–2.71 | |||||
| Adjuvant chemotherapy | Yes | 89 | Ref | 0.395 | ||||
| No | 85 | 0.787 | 0.45–1.36 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishiuchi, A.; Hisamori, S.; Sakaguchi, M.; Fukuyama, K.; Hoshino, N.; Itatani, Y.; Honma, S.; Maekawa, H.; Nishigori, T.; Tsunoda, S.; et al. MicroRNA-9-5p-CDX2 Axis: A Useful Prognostic Biomarker for Patients with Stage II/III Colorectal Cancer. Cancers 2019, 11, 1891. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11121891
Nishiuchi A, Hisamori S, Sakaguchi M, Fukuyama K, Hoshino N, Itatani Y, Honma S, Maekawa H, Nishigori T, Tsunoda S, et al. MicroRNA-9-5p-CDX2 Axis: A Useful Prognostic Biomarker for Patients with Stage II/III Colorectal Cancer. Cancers. 2019; 11(12):1891. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11121891
Chicago/Turabian StyleNishiuchi, Aya, Shigeo Hisamori, Masazumi Sakaguchi, Keita Fukuyama, Nobuaki Hoshino, Yoshiro Itatani, Shusaku Honma, Hisatsugu Maekawa, Tatsuto Nishigori, Shigeru Tsunoda, and et al. 2019. "MicroRNA-9-5p-CDX2 Axis: A Useful Prognostic Biomarker for Patients with Stage II/III Colorectal Cancer" Cancers 11, no. 12: 1891. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11121891