Notch Signaling Molecules as Prognostic Biomarkers for Acute Myeloid Leukemia

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
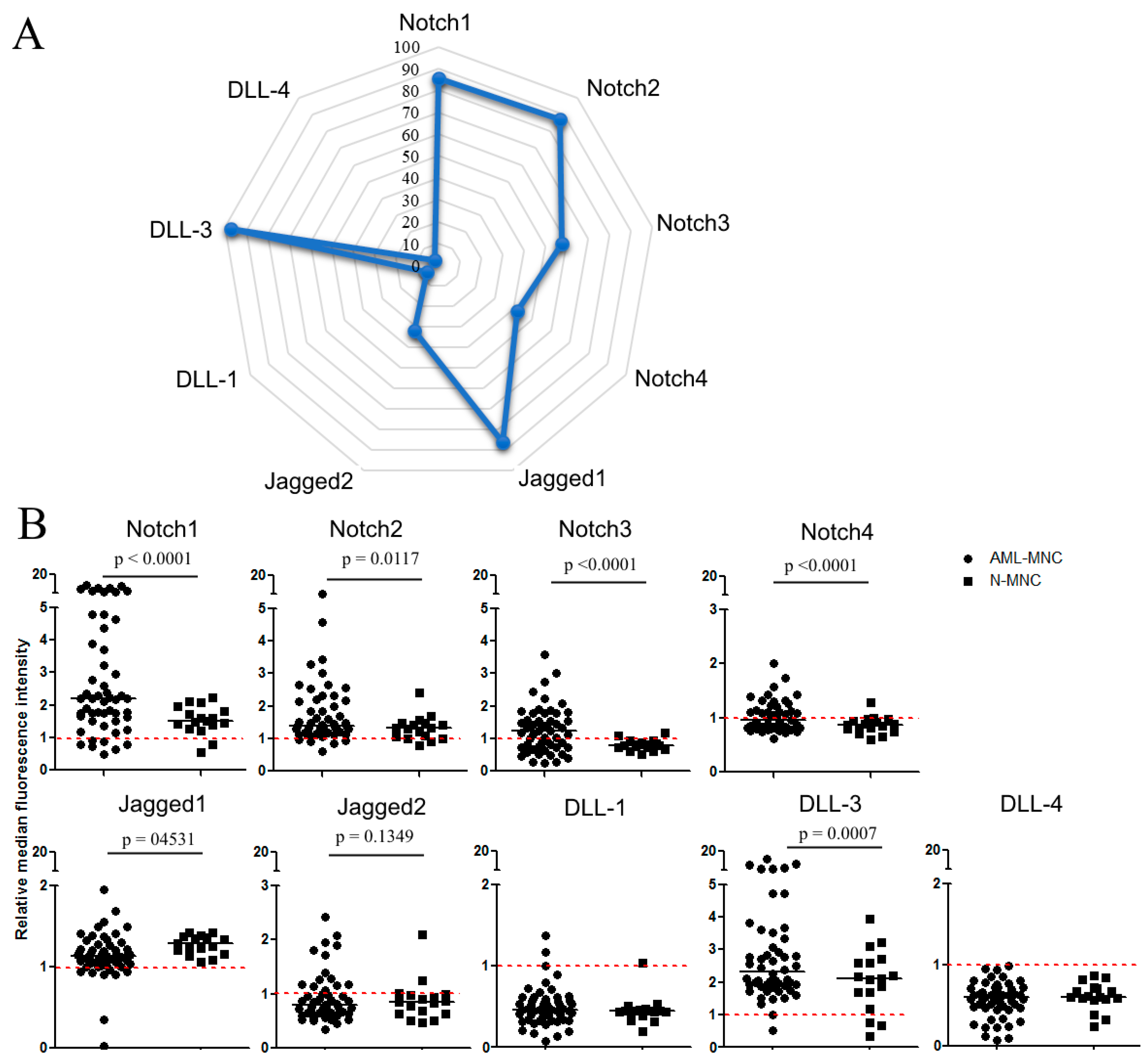
2.1. Flow Cytometric Analysis of Membrane Expression of Notch Molecules
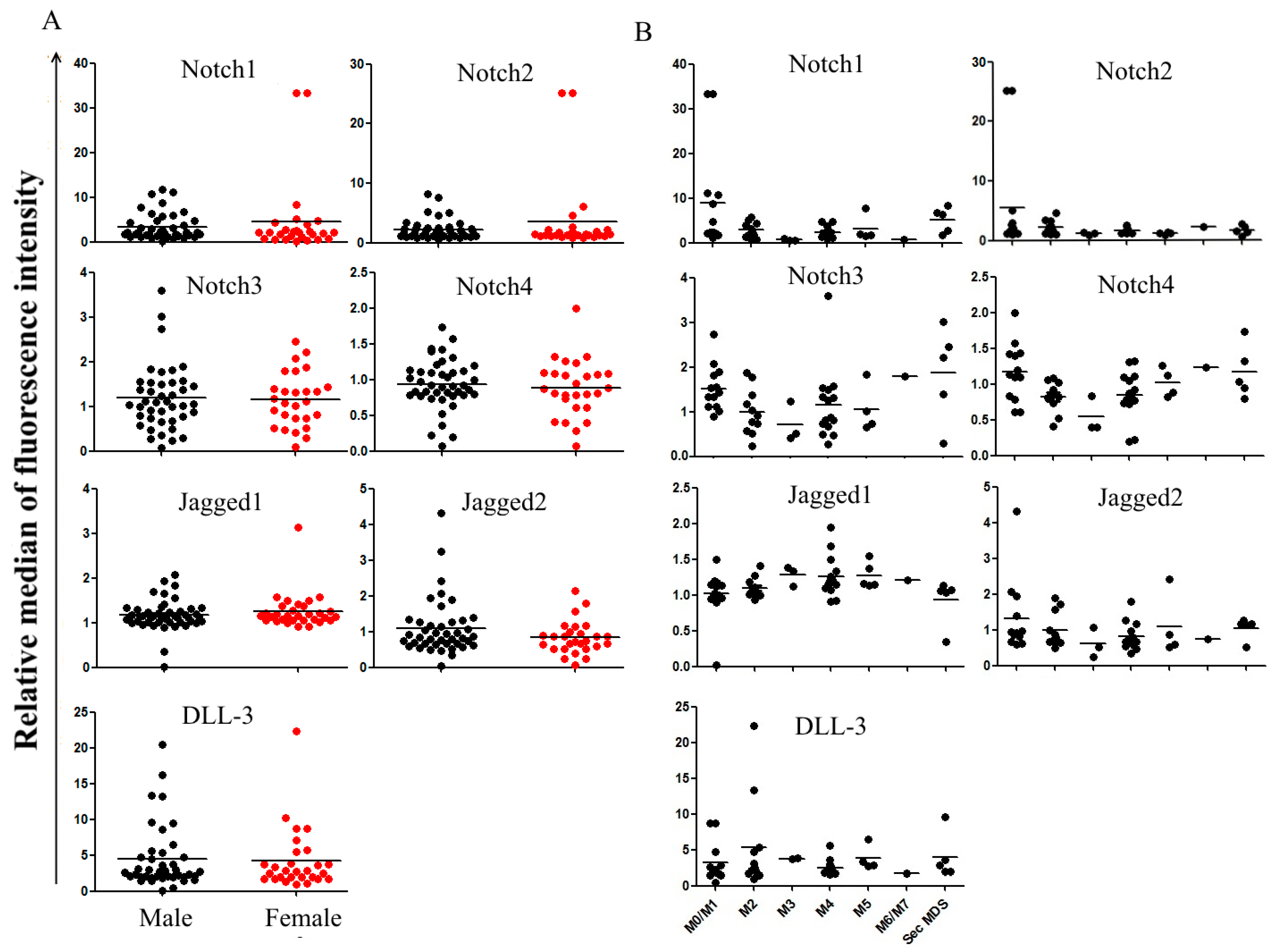
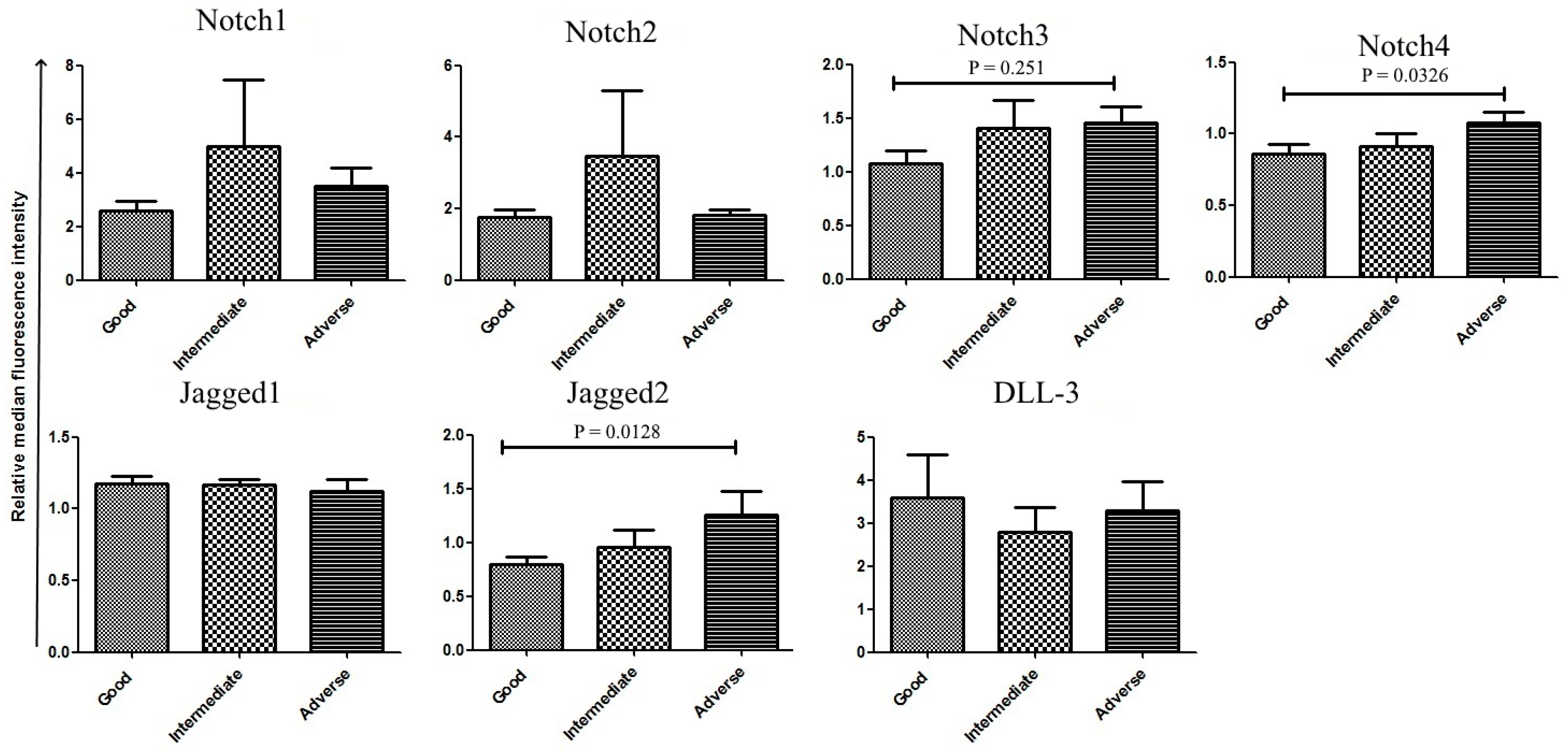
2.2. Correlation between Notch Expression and Known Prognostic Markers
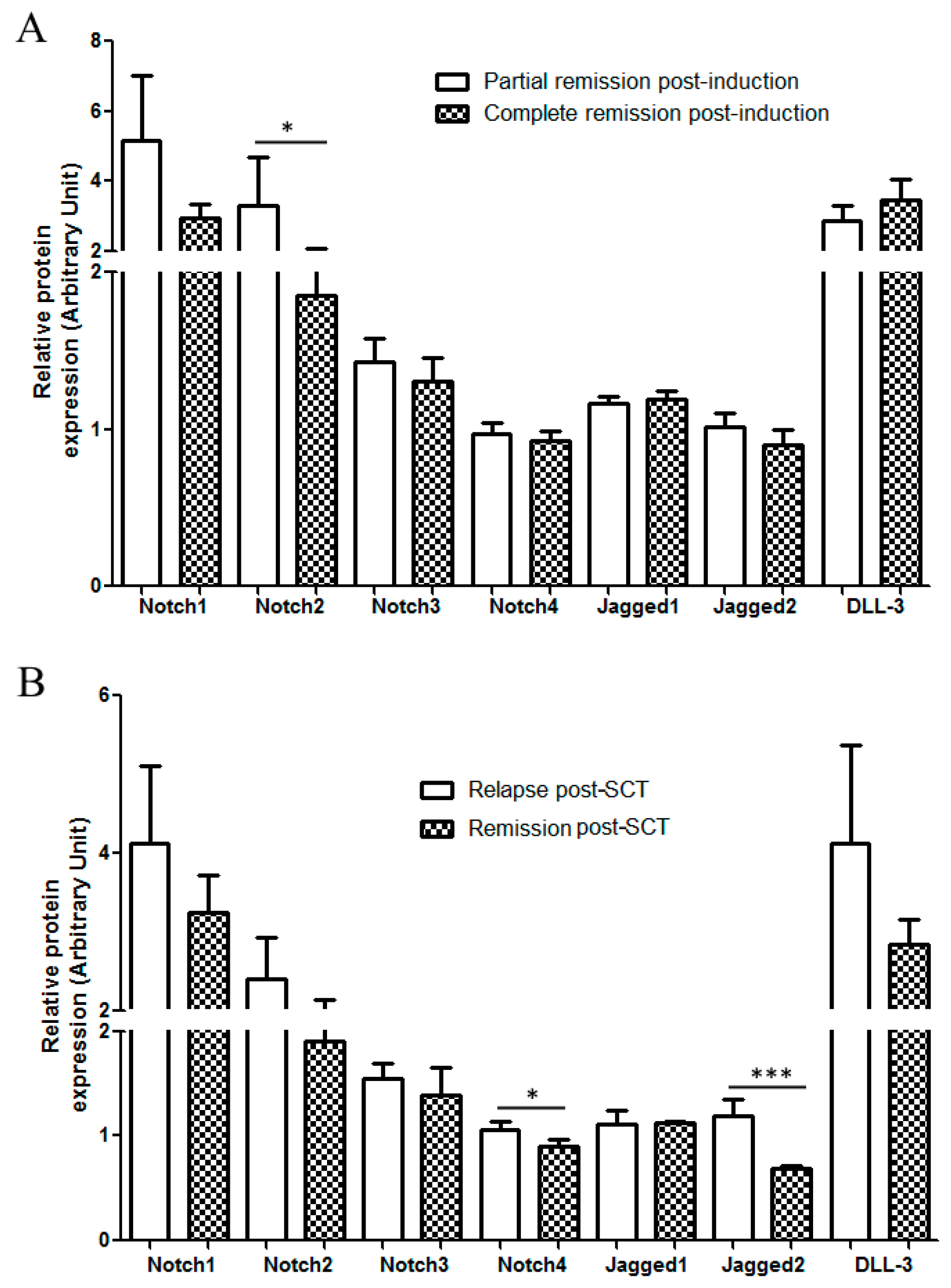
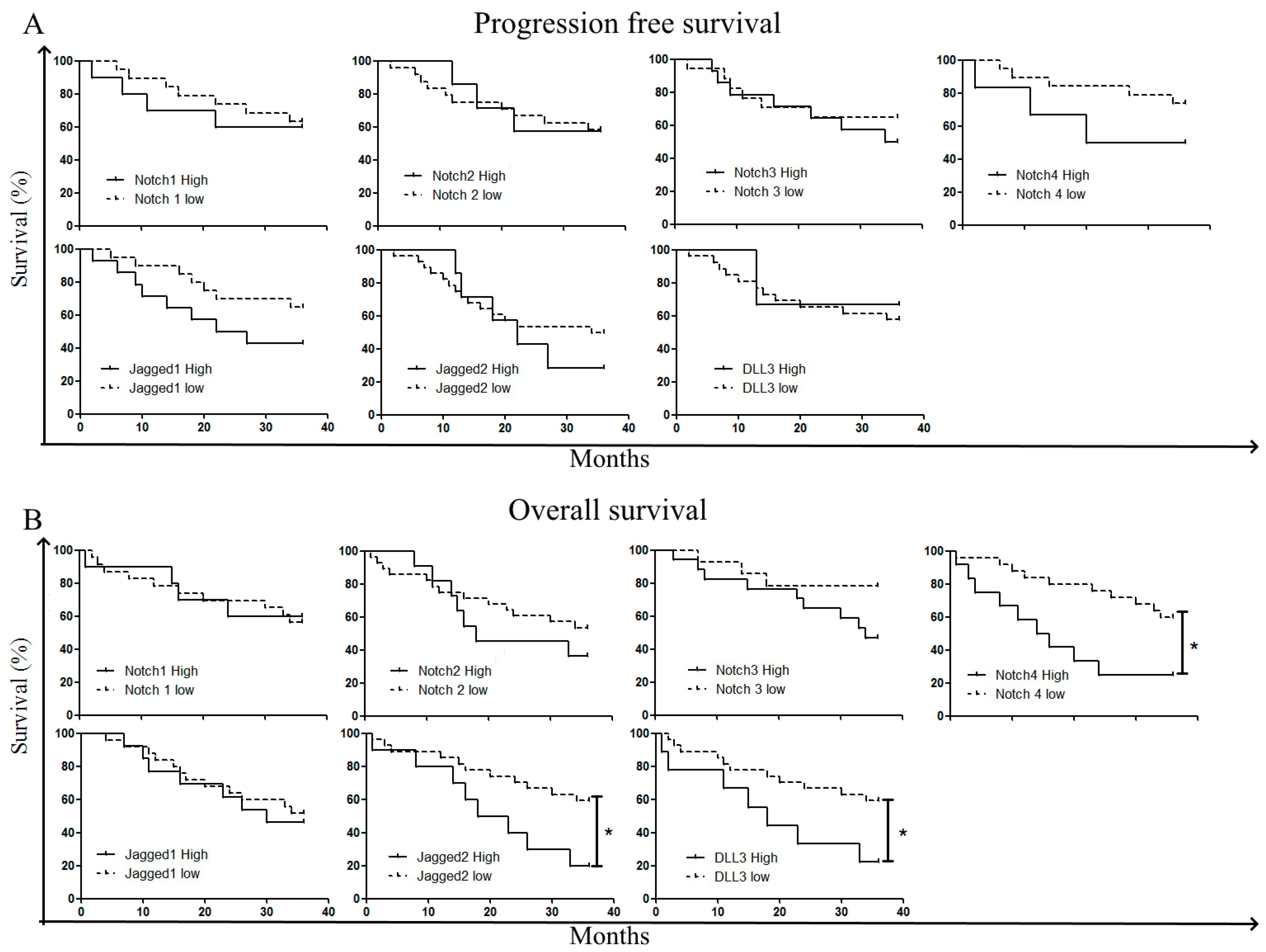
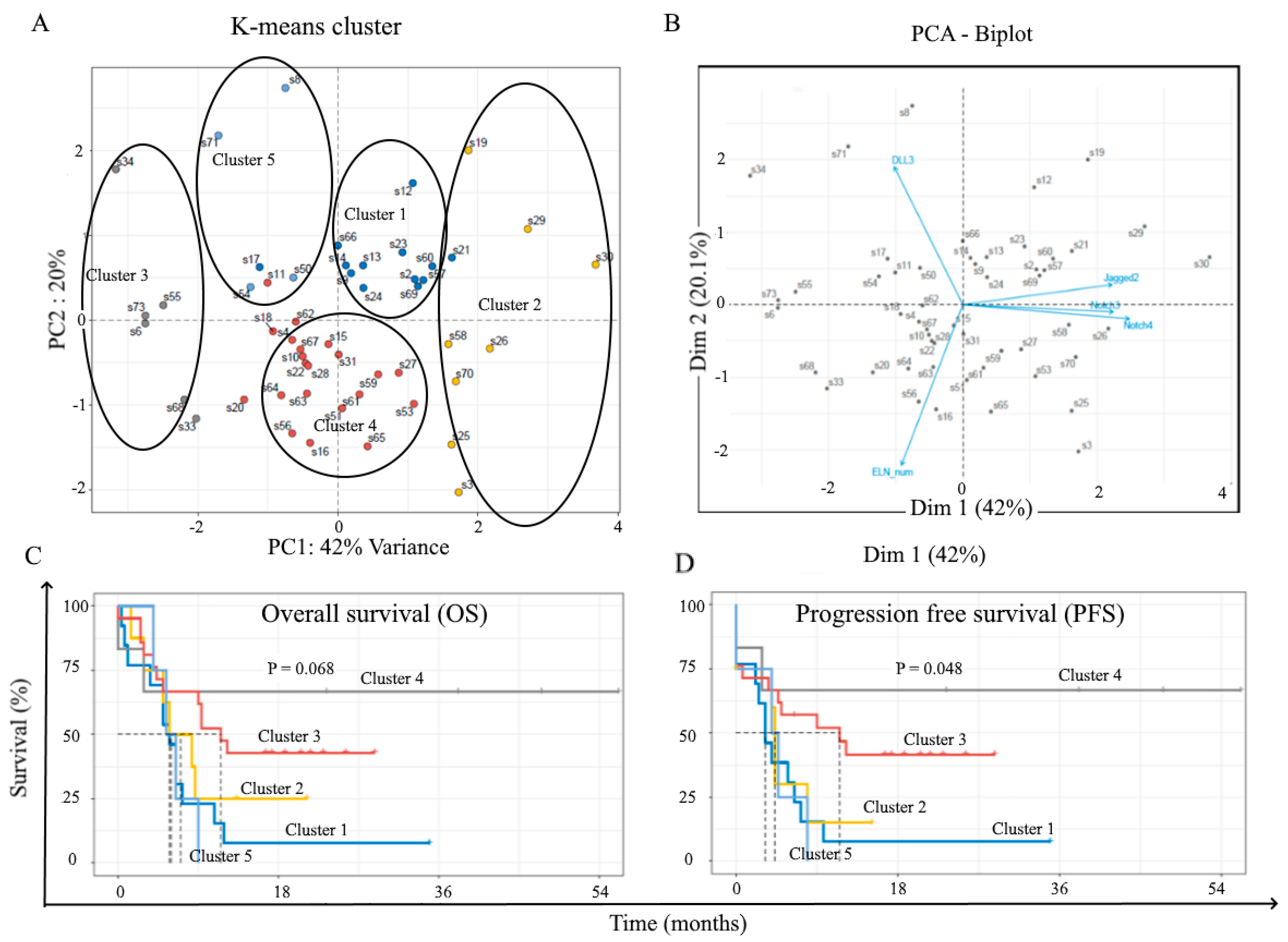
2.3. Correlation with Treatment Outcome and Patient Survival
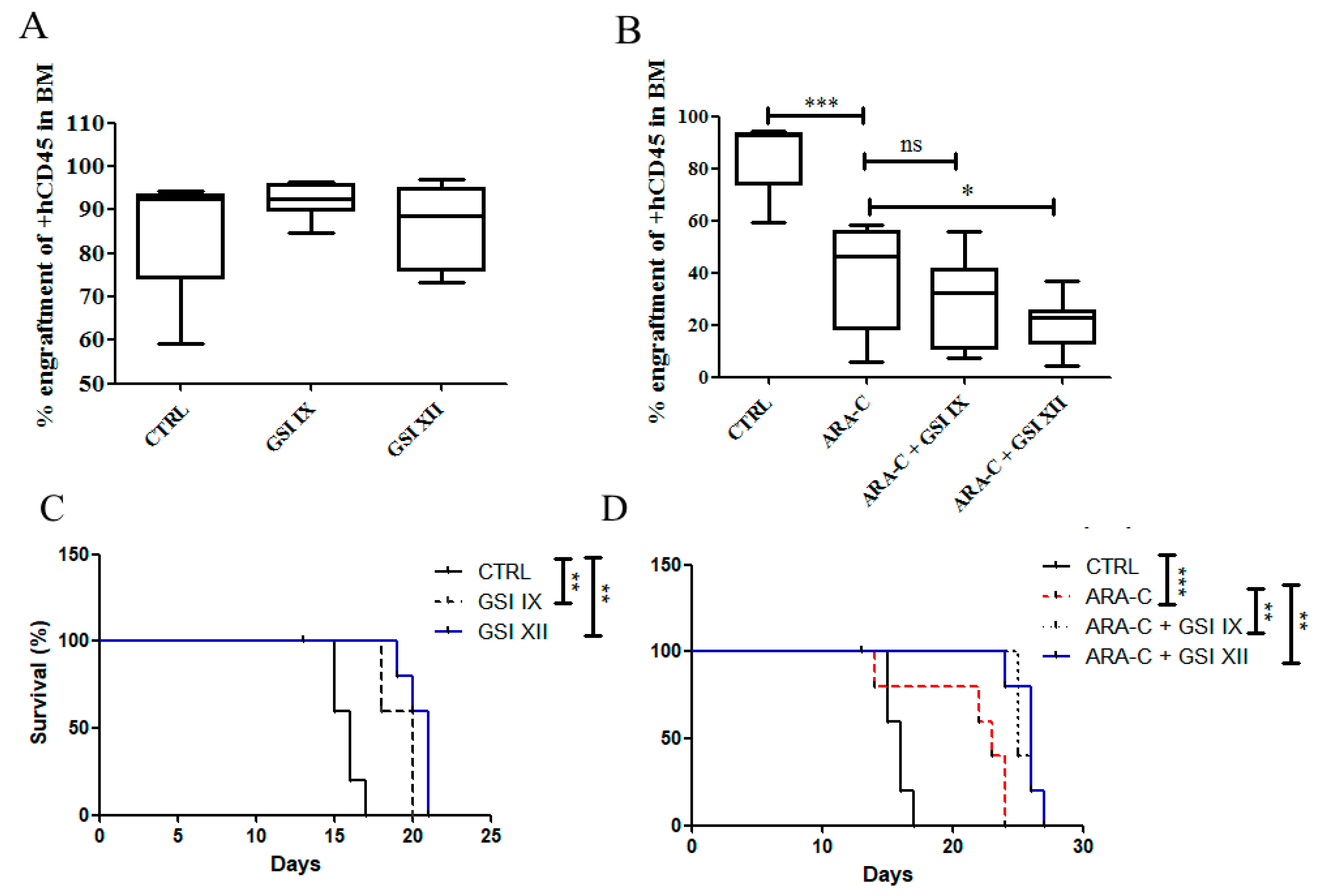
2.4. Notch Inhibition Prolongs Survival of AML Mouse Xenograft Model
3. Discussion
4. Materials and Methods
4.1. Patients, Samples, and Cell Lines
4.2. Chemicals and Antibodies
4.3. Flow Cytometry
4.4. Notch Receptor Immunophenotype
4.5. Xenograft Mouse Model
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kumar, C.C. Genetic abnormalities and challenges in the treatment of acute myeloid leukemia. Genes Cancer 2011, 2, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Longo, D.L.; Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute Myeloid Leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef] [Green Version]
- Single Cell Analysis of Clonal Architecture in Acute Myeloid Leukaemia | Leukemia. Available online: https://0-www-nature-com.brum.beds.ac.uk/articles/s41375-018-0319-2 (accessed on 11 September 2019).
- Prada-Arismendy, J.; Arroyave, J.C.; Röthlisberger, S. Molecular biomarkers in acute myeloid leukemia. Blood Rev. 2017, 31, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef]
- Bewersdorf, J.P.; Stahl, M.; Zeidan, A.M. Are we witnessing the start of a therapeutic revolution in acute myeloid leukemia? Leuk. Lymphoma 2019, 60, 1354–1369. [Google Scholar] [CrossRef]
- Davis, J.R.; Benjamin, D.J.; Jonas, B.A. New and emerging therapies for acute myeloid leukaemia. J. Investig. Med. Off. Publ. Am. Fed. Clin. Res. 2018, 66, 1088–1095. [Google Scholar] [CrossRef]
- Ntziachristos, P.; Lim, J.S.; Sage, J.; Aifantis, I. From Fly Wings to Targeted Cancer Therapies: A Centennial for Notch Signaling. Cancer Cell 2014, 25, 318–334. [Google Scholar] [CrossRef] [Green Version]
- Ferrando, A. NOTCH mutations as prognostic markers in T-ALL. Leukemia 2010, 24, 2003–2004. [Google Scholar] [CrossRef] [Green Version]
- Rosati, E.; Sabatini, R.; Rampino, G.; Tabilio, A.; Di Ianni, M.; Fettucciari, K.; Bartoli, A.; Coaccioli, S.; Screpanti, I.; Marconi, P. Constitutively activated Notch signaling is involved in survival and apoptosis resistance of B-CLL cells. Blood 2009, 113, 856–865. [Google Scholar] [CrossRef]
- Di Ianni, M.; Baldoni, S.; Del Papa, B.; Aureli, P.; Dorillo, E.; De Falco, F.; Albi, E.; Varasano, E.; Di Tommaso, A.; Giancola, R.; et al. NOTCH1 Is Aberrantly Activated in Chronic Lymphocytic Leukemia Hematopoietic Stem Cells. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Palomero, T.; McKenna, K.; O-Neil, J.; Galinsky, I.; Stone, R.; Suzukawa, K.; Stiakaki, E.; Kalmanti, M.; Fox, E.A.; Caligiuri, M.A.; et al. Activating mutations in NOTCH1 in acute myeloid leukemia and lineage switch leukemias. Leukemia 2006, 20, 1963–1966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Zhao, Y.; Xu, M.; Dai, Q.; Meng, W.; Yang, J.; Qin, R. Activation of Notch signal pathway is associated with a poorer prognosis in acute myeloid leukemia. Med. Oncol. 2010, 28, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ma, D.; Ye, J.; Zang, S.; Lu, F.; Yang, M.; Qu, X.; Sun, X.; Ji, C. Prognostic impact of δ-like ligand 4 and Notch1 in acute myeloid leukemia. Oncol. Rep. 2012, 28, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Czemerska, M.; Pluta, A.; Szmigielska-Kaplon, A.; Wawrzyniak, E.; Cebula-Obrzut, B.; Medra, A.; Smolewski, P.; Robak, T.; Wierzbowska, A. Jagged-1: A new promising factor associated with favorable prognosis in patients with acute myeloid leukemia. Leuk. Lymphoma 2015, 56, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Sliwa, T.; Awsa, S.; Vesely, M.; Rokitte, D.; Grossschmidt, P.; Jilch, R.; Ulrich, W.; Geissler, K. Hyperexpression of NOTCH-1 is found in immature acute myeloid leukemia. Int. J. Clin. Exp. Pathol. 2014, 7, 882–889. [Google Scholar]
- Turbeville, S.; Francis, K.M.; Behm, I.; Chiu, G.R.; Sanchez, H.; Morrison, B.A.; Rowe, J.M. Prevalence and Incidence of Acute Myeloid Leukemia May be Higher Than Currently Accepted Estimates Among the ≥65 Year-Old Population in the United States. Blood 2014, 124, 958. [Google Scholar] [CrossRef]
- Lohse, I.; Statz-Geary, K.; Brothers, S.P.; Wahlestedt, C. Precision medicine in the treatment stratification of AML patients: Challenges and progress. Oncotarget 2018, 9, 37790–37797. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Zhang, J.; Ji, C. The emerging roles of Notch signaling in leukemia and stem cells. Biomark. Res. 2013, 1, 23. [Google Scholar] [CrossRef] [Green Version]
- Lobry, C.; Ntziachristos, P.; Ndiaye-Lobry, D.; Oh, P.; Cimmino, L.; Zhu, N.; Araldi, E.; Hu, W.; Freund, J.; Abdel-Wahab, O.; et al. Notch pathway activation targets AML-initiating cell homeostasis and differentiation. J. Exp. Med. 2013, 210, 301–319. [Google Scholar] [CrossRef]
- Kannan, S.; Sutphin, R.M.; Hall, M.G.; Golfman, L.S.; Fang, W.; Nolo, R.M.; Akers, L.J.; Hammitt, R.A.; McMurray, J.S.; Kornblau, S.M.; et al. Notch activation inhibits AML growth and survival: A potential therapeutic approach. J. Exp. Med. 2013, 210, 321–337. [Google Scholar] [CrossRef]
- Tohda, S.; Kogoshi, H.; Murakami, N.; Sakano, S.; Nara, N. Diverse effects of the Notch ligands Jagged1 and Delta1 on the growth and differentiation of primary acute myeloblastic leukemia cells. Exp. Hematol. 2005, 33, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Nwabo Kamdje, A.H.; Takam Kamga, P.; Tagne Simo, R.; Vecchio, L.; Seke Etet, P.F.; Muller, J.M.; Bassi, G.; Lukong, E.; Kumar Goel, R.; Mbo Amvene, J.; et al. Developmental pathways associated with cancer metastasis: Notch, Wnt, and Hedgehog. Cancer Biol. Med. 2017, 14, 109–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampreia, F.P.; Carmelo, J.G.; Anjos-Afonso, F. Notch Signaling in the Regulation of Hematopoietic Stem Cell. Curr. Stem Cell Rep. 2017, 3, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aster, J.C.; Pear, W.S.; Blacklow, S.C. The Varied Roles of Notch in Cancer. Annu. Rev. Pathol. 2017, 12, 245–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notch signalling drives bone marrow stromal cell-mediated chemoresistance in acute myeloid leukemia. Oncotarget 2015. [CrossRef] [Green Version]
- Kamga, P.T.; Collo, G.D.; Midolo, M.; Adamo, A.; Delfino, P.; Mercuri, A.; Cesaro, S.; Mimiola, E.; Bonifacio, M.; Andreini, A.; et al. Inhibition of Notch signaling enhances chemosensitivity in B cell precursor acute lymphoblastic leukemia. Cancer Res. 2019, 79, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Bernasconi-Elias, P.; Hu, T.; Jenkins, D.; Firestone, B.; Gans, S.; Kurth, E.; Capodieci, P.; Deplazes-Lauber, J.; Petropoulos, K.; Thiel, P.; et al. Characterization of activating mutations of NOTCH3 in T cell acute lymphoblastic leukemia and anti-leukemic activity of NOTCH3 inhibitory antibodies. Oncogene 2016, 35, 6077–6086. [Google Scholar] [CrossRef] [Green Version]
- Tottone, L.; Zhdanovskaya, N.; Carmona Pestaña, Á.; Zampieri, M.; Simeoni, F.; Lazzari, S.; Ruocco, V.; Pelullo, M.; Caiafa, P.; Felli, M.P.; et al. Histone Modifications Drive Aberrant Notch3 Expression/Activity and Growth in T-ALL. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Aburjania, Z.; Jang, S.; Whitt, J.; Jaskula-Stzul, R.; Chen, H.; Rose, J.B. The Role of Notch3 in Cancer. Oncologist 2018, 23, 900–911. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-W.; Wei, X.-L.; Dou, X.-W.; Huang, W.-H.; Du, C.-W.; Zhang, G.-J. The association between Notch4 expression, and clinicopathological characteristics and clinical outcomes in patients with breast cancer. Oncol. Lett. 2018, 15, 8749–8755. [Google Scholar] [CrossRef] [Green Version]
- Nagamatsu, I.; Onishi, H.; Matsushita, S.; Kubo, M.; Kai, M.; Imaizumi, A.; Nakano, K.; Hattori, M.; Oda, Y.; Tanaka, M.; et al. NOTCH4 Is a Potential Therapeutic Target for Triple-negative Breast Cancer. Anticancer Res. 2014, 34, 69–80. [Google Scholar] [PubMed]
- Fukusumi, T.; Guo, T.W.; Sakai, A.; Ando, M.; Ren, S.; Haft, S.; Liu, C.; Amornphimoltham, P.; Gutkind, J.S.; Califano, J.A. The NOTCH4–HEY1 Pathway Induces Epithelial–Mesenchymal Transition in Head and Neck Squamous Cell Carcinoma. Clin. Cancer Res. 2018, 24, 619–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelliher, M.A.; Roderick, J.E. NOTCH Signaling in T-Cell-Mediated Anti-Tumor Immunity and T-Cell-Based Immunotherapies. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, F.; Osborne, B.A. Non-canonical notch signaling in cancer and immunity. Front. Oncol. 2014, 4, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamal, A.; Handayani, A.; Septiandri, A.A.; Ripmiatin, E.; Effendi, Y. Dimensionality Reduction using PCA and K-Means Clustering for Breast Cancer Prediction. Lontar Komput. J. Ilm. Teknol. Inf. 2018, 192–201. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | N = 79 |
|---|---|
| Gender | Females = 31 |
| Males = 48 | |
| Median age | 52 (16–74) |
| FAB Subtypes (n = 57) | M1/M0 = 14 |
| M2 = 13 | |
| M3 = 3 | |
| M4 = 16 | |
| M5 = 5 | |
| M6/M7 = 1 | |
| Secondary to myelodysplasia = 5 | |
| Other = 1 | |
| Hemoglobin | Hb (g/dL) = 8.7 (3.16–12.9) |
| Platelets | PLT count (109/L) = 67.5 (6–319) |
| White blood cells | WBC count (109/L) = 20 (0.5–21.8) |
| ELN stratification (n = 51) | Good = 22 |
| Intermediate = 17 | |
| Adverse =18 | |
| Induction therapy (58) | 7+3 = 26 |
| MICE = 26 | |
| FLAI = 2 | |
| Other = 4 | |
| HSCT (n = 29) | Three year remission = 10 |
| Relapse within 3 years = 19 |
| A | WBC | Notch1 | Notch2 | Notch3 | Notch4 | Jagged1 | Jagged2 | DLL-3 |
| 54 | 54 | 54 | 54 | 54 | 54 | 53 | ||
| r | −0.3051 | −0.2424 | 0.07906 | −0.08409 | 0.2046 | −0.1569 | 0.03657 | |
| p-value | 0.0124 | 0.0387 | 0.2849 | 0.2727 | 0.0689 | 0.1286 | 0.3974 | |
| Statistics | * | * | ns | ns | ns | ns | ns | |
| B | Hb | Notch1 | Notch2 | Notch3 | Notch4 | Jagged1 | Jagged2 | DLL-3 |
| 54 | 54 | 54 | 54 | 54 | 54 | 53 | ||
| r | 0.01171 | −0.1183 | 0.03234 | 0.0008389 | 0.2720 | −0.2440 | −0.1473 | |
| p-value | 0.4665 | 0.1971 | 0.4082 | 0.4976 | 0.0233 | 0.0377 | 0.1462 | |
| Statistics | ns | ns | ns | ns | * | * | ns | |
| C | PLT | Notch1 | Notch2 | Notch3 | Notch4 | Jagged1 | Jagged2 | DLL-3 |
| 54 | 54 | 54 | 54 | 54 | 54 | 53 | ||
| r | 0.2905 | 0.02284 | 0.05585 | 0.05406 | 0.1449 | 0.03023 | −0.02843 | |
| p-value | 0.0166 | 0.4349 | 0.3442 | 0.3489 | 0.1480 | 0.4141 | 0.4199 | |
| Statistics | * | ns | ns | ns | ns | ns | ns |
| Notch1 | Notch2 | Notch3 | Notch4 | Jagged1 | Jagged2 | DLL-3 | |
|---|---|---|---|---|---|---|---|
| Chi-score | 0.1822 | 2.737 | 3.424 | 0.01225 | 0.3810 | 2.915 | 0.1822 |
| p-value | 0.3348 | 0.0490 | 0.0321 | 0.4559 | 0.2685 | 0.0878 | 0.3348 |
| Statistics | ns | * | * | ns | ns | ns | ns |
| Notch1 | Notch2 | Notch3 | Notch4 | Jagged1 | Jagged2 | DLL-3 | |
|---|---|---|---|---|---|---|---|
| Chi-score | 0.03 | 0.69 | 1.15 | 2.65 | 0.69 | 7.64 | 1.01 |
| p-Value | 0.43 | 0.20 | 0.14 | 0.05 | 0.20 | 0.003 | 0.15 |
| Statistics | ns | ns | ns | * | ns | * | ns |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takam Kamga, P.; Collo, G.D.; Resci, F.; Bazzoni, R.; Mercuri, A.; Quaglia, F.M.; Tanasi, I.; Delfino, P.; Visco, C.; Bonifacio, M.; et al. Notch Signaling Molecules as Prognostic Biomarkers for Acute Myeloid Leukemia. Cancers 2019, 11, 1958. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11121958
Takam Kamga P, Collo GD, Resci F, Bazzoni R, Mercuri A, Quaglia FM, Tanasi I, Delfino P, Visco C, Bonifacio M, et al. Notch Signaling Molecules as Prognostic Biomarkers for Acute Myeloid Leukemia. Cancers. 2019; 11(12):1958. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11121958
Chicago/Turabian StyleTakam Kamga, Paul, Giada Dal Collo, Federica Resci, Riccardo Bazzoni, Angela Mercuri, Francesca Maria Quaglia, Ilaria Tanasi, Pietro Delfino, Carlo Visco, Massimiliano Bonifacio, and et al. 2019. "Notch Signaling Molecules as Prognostic Biomarkers for Acute Myeloid Leukemia" Cancers 11, no. 12: 1958. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers11121958