Genomic Instability Signature of Palindromic Non-Coding Somatic Mutations in Bladder Cancer

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Patients and Samples
2.2. RNA and DNA Extractions
2.3. In Silico Identification of New TGAACA Core Motif of Palindromic Sequences in the Human Genome
2.4. DNA Non-Coding Mutations Analysis
2.5. Targeted Next Generation Sequencing (NGS)
2.6. Assessment of the Microsatellite Instability (MSI) Status
2.7. Real-Time Quantitative RT-PCR
2.8. Statistical Analysis
3. Results
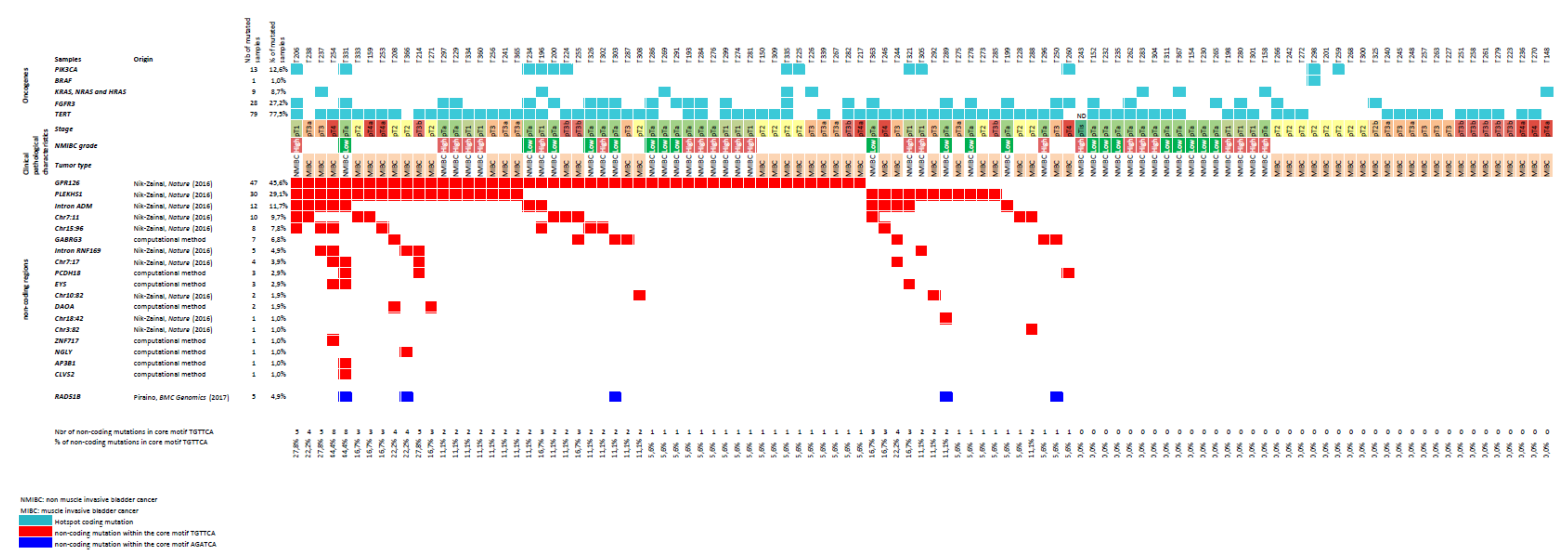
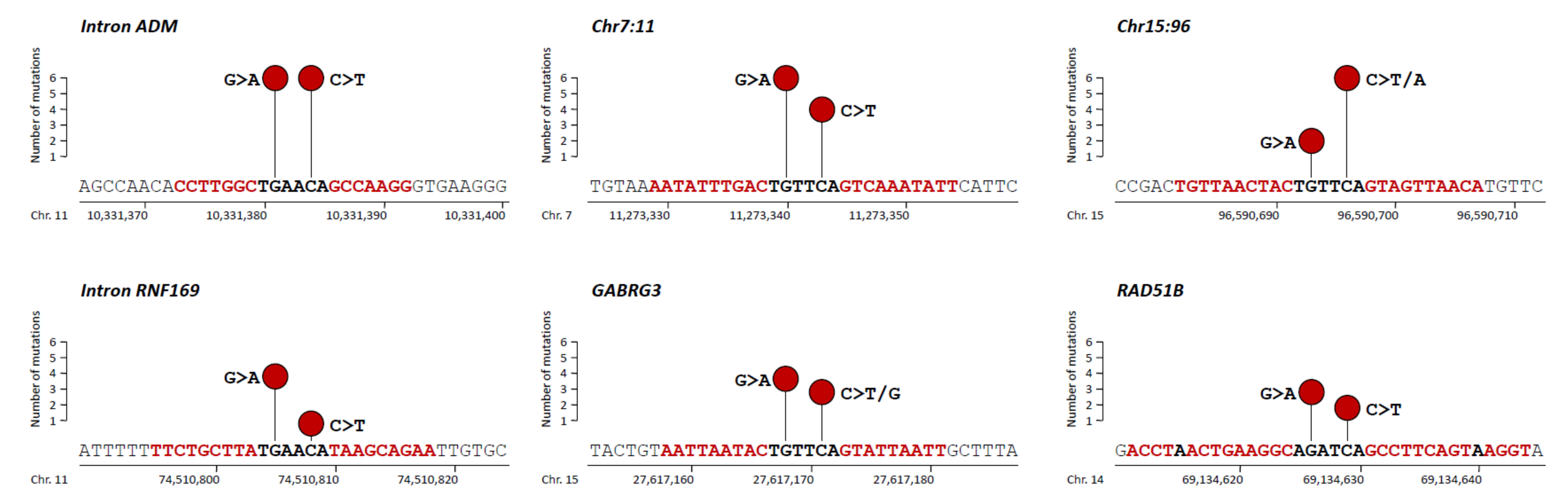
3.1. Non-Coding Mutations within a TGAACA Core Motif of 10 Palindromic Sequences
3.2. Additional Non-Coding Mutations within a TGAACA Core Motif of Palindromic Sequences Identified by in Silico Analysis of the Human Genome
3.3. Non-Coding Mutations within an AGATCA Core Motif of Palindromic Sequences in an Intron of RAD51B
3.4. Relationship between APOBEC3 RNA Level and TGAACA Core Motif Mutations
3.5. Association between TGAACA Core Motif Mutations and Clinico-Biological Parameters
3.6. Protein-Coding Gene Mutations in Bladder Tumors Showing High Levels of Non-Coding TGAACA Mutations
3.7. Association between TGAACA Core Motif Mutational Signature and Expression Levels of Immune-Related Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| HRM | high-resolution melting |
| MIBC | muscle-invasive bladder cancer |
| NMIBC | non-muscle-invasive bladder cancer |
| WHO | World Health Organization |
| OS | overall survival |
| PFS | progression-free survival |
| RFS | recurrence-free survival |
| PCR | polymerase chain reaction |
| TNM | tumor node metastasis |
| TMB | tumor mutational burden |
| HNSCC | Head and Neck Squamous Cell Carcinoma |
| HPV | human papilloma virus |
| TSG | tumor suppressor gene |
| NGS | next-generation sequencing |
References
- Antoni, S.; Ferlay, J.; Soerjomataram, I.; Znaor, A.; Jemal, A.; Bray, F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur. Urol. 2017, 71, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Kluth, L.A.; Black, P.C.; Bochner, B.H.; Catto, J.; Lerner, S.P.; Stenzl, A.; Sylvester, R.; Vickers, A.J.; Xylinas, E.; Shariat, S.F. Prognostic and Prediction Tools in Bladder Cancer: A Comprehensive Review of the Literature. Eur. Urol. 2015, 68, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Bellmunt, J.; Orsola, A.; Leow, J.J.; Wiegel, T.; De Santis, M.; Horwich, A. Bladder cancer: ESMO Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2014, 25 (Suppl. 3), 40–48. [Google Scholar] [CrossRef] [PubMed]
- Soria, F.; Krabbe, L.M.; Todenhöfer, T.; Dobruch, J.; Mitra, A.P.; Inman, B.A.; Gust, K.M.; Lotan, Y.; Shariat, S.F. Molecular markers in bladder cancer. World J. Urol. 2019, 37, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Research Network. Comprehensive molecular characterization of urothelial bladder carcinoma. Nature 2014, 507, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Enokida, H.; Yoshino, H.; Matsushita, R.; Nakagawa, M. The role of microRNAs in bladder cancer. Investig. Clin. Urol. 2016, 57 (Suppl. 1), S60–S76. [Google Scholar] [CrossRef]
- Li, Q.; Wang, H.; Peng, H.; Huang, Q.; Huyan, T.; Huang, Q.; Yang, H.; Shi, J. MicroRNAs: Key Players in Bladder Cancer. Mol. Diagn. Ther. 2019, 23, 579–601. [Google Scholar] [CrossRef]
- Weinhold, N.; Jacobsen, A.; Schultz, N.; Sander, C.; Lee, W. Genome-wide analysis of noncoding regulatory mutations in cancer. Nat. Genet. 2014, 46, 1160–1165. [Google Scholar] [CrossRef]
- Rheinbay, E.; Parasuraman, P.; Grimsby, J.; Tiao, G.; Engreitz, J.M.; Kim, J.; Lawrence, M.S.; Taylor-Weiner, A.; Rodriguez-Cuevas, S.; Rosenberg, M.; et al. Recurrent and functional regulatory mutations in breast cancer. Nature 2017, 547, 55–60. [Google Scholar] [CrossRef]
- Fujimoto, A.; Furuta, M.; Totoki, Y.; Tsunoda, T.; Kato, M.; Shiraishi, Y.; Tanaka, H.; Taniguchi, H.; Kawakami, Y.; Ueno, M.; et al. Whole-genome mutational landscape and characterization of noncoding and structural mutations in liver cancer. Nat. Genet. 2016, 48, 500–509. [Google Scholar] [CrossRef]
- Puente, X.S.; Beà, S.; Valdés-Mas, R.; Villamor, N.; Gutiérrez-Abril, J.; Martín-Subero, J.I.; Munar, M.; Rubio-Pérez, C.; Jares, P.; Aymerich, M.; et al. Non-coding recurrent mutations in chronic lymphocytic leukaemia. Nature 2015, 526, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Piraino, S.W.; Furney, S.J. Identification of coding and non-coding mutational hotspots in cancer genomes. BMC Genom. 2017, 18, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nik-Zainal, S.; Davies, H.; Staaf, J.; Ramakrishna, M.; Glodzik, D.; Zou, X.; Martincorena, I.; Alexandrov, L.B.; Martin, S.; Wedge, D.C.; et al. Landscape of somatic mutations in 560 breast cancer whole-genome sequences. Nature 2016, 534, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Buisson, R.; Langenbucher, A.; Bowen, D.; Kwan, E.E.; Benes, C.H.; Zou, L.; Lawrence, M.S. Passenger hotspot mutations in cancer driven by APOBEC3A and mesoscale genomic features. Science 2019, 28, 364. [Google Scholar] [CrossRef]
- Rheinbay, E.; Nielsen, M.M.; Abascal, F.; Wala, J.A.; Shapira, O.; Tiao, G.; Hornshøj, H.; Hess, J.M.; Juul, R.I.; Lin, Z.; et al. Analyses of non-coding somatic drivers in 2658 cancer whole genomes. Nature 2020, 578, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Rachakonda, P.S.; Hosen, I.; De Verdier, P.J.; Fallah, M.; Heidenreich, B.; Ryk, C.; Wiklund, N.P.; Steineck, G.; Schadendorf, D.; Hemminki, K.; et al. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by a common polymorphism. Proc. Natl. Acad. Sci. USA 2013, 110, 17426–17431. [Google Scholar] [CrossRef] [Green Version]
- Garinet, S.; Pignot, G.; Vacher, S.; Le Goux, C.; Schnitzler, A.; Chemlali, W.; Sirab, N.; Barry Delongchamps, N.; Zerbib, M.; Sibony, M.; et al. High Prevalence of a Hotspot of Noncoding Somatic Mutations in Intron 6 of GPR126 in Bladder Cancer. Mol. Cancer Res. 2019, 17, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Pignot, G.; Le Goux, C.; Vacher, S.; Schnitzler, A.; Radvanyi, F.; Allory, Y.; Lallemand, F.; Barry Delongchamps, N.; Zerbib, M.; Terris, B.; et al. PLEKHS1: A new molecular marker predicting risk of progression of non-muscle-invasive bladder cancer. Oncol. Lett. 2019, 18, 3471–3480. [Google Scholar] [CrossRef] [Green Version]
- Sobin, L.; Gospodarowicz, M.; Wittekind, C. TNM classification of malignant tumors. UICC 2010, 262–265. [Google Scholar]
- Bièche, I.; Onody, P.; Tozlu, S.; Driouch, K.; Vidaud, M.; Lidereau, R. Prognostic value of ERBB family mRNA expression in breast carcinomas. Int. J. Cancer 2003, 106, 758–765. [Google Scholar] [CrossRef]
- Ohl, F.; Jung, M.; Radonić, A.; Sachs, M.; Loening, S.A.; Jung, K. Identification and validation of suitable endogenous reference genes for gene expression studies of human bladder cancer. J. Urol. 2006, 175, 1915–1920. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Kim, J.; Haradhvala, N.J.; Huang, M.N.; Tian Ng, A.W.; Wu, Y.; Boot, A.; Covington, K.R.; Gordenin, D.A.; Bergstrom, E.N.; et al. The repertoire of mutational signatures in human cancer. Nature 2020, 578, 94–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samstein, R.M.; Lee, C.H.; Shoushtari, A.N.; Hellmann, M.D.; Shen, R.; Janjigian, Y.Y.; Barron, D.A.; Zehir, A.; Jordan, E.J.; Omuro, A.; et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat. Genet. 2019, 51, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Borcoman, E.; De La Rochere, P.; Richer, W.; Vacher, S.; Chemlali, W.; Krucker, C.; Sirab, N.; Radvanyi, F.; Allory, Y.; Pignot, G.; et al. Inhibition of PI3K pathway increases immune infiltrate in muscle-invasive bladder cancer. Oncoimmunology 2019, 8, e1581556. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.; Chakravarthy, A.; Su, X.; Boshoff, C.; Fenton, T.R. APOBEC-mediated cytosine deamination links PIK3CA helical domain mutations to human papillomavirus-driven tumor development. Cell Rep. 2014, 7, 1833–1841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudley, J.C.; Schroers-Martin, J.; Lazzareschi, D.V.; Shi, W.Y.; Chen, S.B.; Esfahani, M.S.; Trivedi, D.; Chabon, J.J.; Chaudhuri, A.A.; Stehr, H.; et al. Detection and Surveillance of Bladder Cancer Using Urine Tumor DNA. Cancer Discov. 2019, 9, 500–509. [Google Scholar] [CrossRef]
- Christensen, E.; Birkenkamp-Demtröder, K.; Sethi, H.; Shchegrova, S.; Salari, R.; Nordentoft, I.; Wu, H.T.; Knudsen, M.; Lamy, P.; Lindskrog, S.V.; et al. Early Detection of Metastatic Relapse and Monitoring of Therapeutic Efficacy by Ultra-Deep Sequencing of Plasma Cell-Free DNA in Patients With Urothelial Bladder Carcinoma. J. Clin. Oncol. 2019, 37, 1547–1557. [Google Scholar] [CrossRef]
- Marabelle, A.; Le, D.T.; Ascierto, P.A.; Di Giacomo, A.M.; De Jesus-Acosta, A.; Delord, J.P.; Geva, R.; Gottfried, M.; Penel, N.; Hansen, A.R.; et al. Efficacy of Pembrolizumab in Patients with Noncolorectal High Microsatellite Instability/Mismatch Repair-Deficient Cancer: Results From the Phase II KEYNOTE-158 Study. J. Clin. Oncol. 2020, 38, 1–10. [Google Scholar] [CrossRef]
- Roper, N.; Gao, S.; Maity, T.K.; Banday, A.R.; Zhang, X.; Venugopalan, A.; Cultraro, C.M.; Patidar, R.; Sindiri, S.; Brown, A.L.; et al. APOBEC Mutagenesis and Copy-Number Alterations Are Drivers of Proteogenomic Tumor Evolution and Heterogeneity in Metastatic Thoracic Tumors. Cell Rep. 2019, 26, 2651–2666. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Parameters | Whole Population | No Recurrence | Recurrence | Muscle-Invasive Progression | ||
|---|---|---|---|---|---|---|
| Number of Patients (%) | Number (%) | Number (%) | p-Value * | Number (%) | p-Value ** | |
| Total population | 44 (100.0) | 14 (31.8) | 22 (50.0) | 8 (18.2) | ||
| Age (years) | ||||||
| ≥60 | 34 (77.3) | 10 (29.4) | 16 (47.1) | 0.77 | 8 (23.5) | 0.17 |
| <60 | 10 (22.7) | 4 (40.0) | 6 (60.0) | 0 (0.0) | ||
| Sex | ||||||
| Male | 41 (93.2) | 13 (31.7) | 20 (48.8) | >0.99 | 8 (19.5) | >0.99 |
| Female | 3 (6.8) | 1 (33.3) | 2 (66.7) | 0 (0.0) | ||
| Smoking status | ||||||
| Non-smoker | 19 (43.2) | 4 (21.1) | 12 (63.2) | 0.13 | 3 (15.8) | 0.97 |
| Smoker | 25 (56.8) | 10 (40.0) | 10 (40.0) | 5 (20.0) | ||
| History of NMIBC | ||||||
| No | 24 (54.5) | 11 (45.8) | 10 (41.7) | 0.049 | 3 (12.5) | 0.50 |
| Yes | 20 (45.5) | 3 (15.0) | 12 (60.0) | 5 (25.0) | ||
| Associated pTis | ||||||
| No | 42 (95.4) | 14 (33.3) | 22 (52.4) | NA | 6 (14.3) | 0.030 |
| Yes | 2 (4.6) | 0 (0.0) | 0 (0.0) | 2 (100) | ||
| Grade | ||||||
| Low grade | 20 (45.5) | 9 (45.0) | 10 (50.0) | 0.27 | 1 (5.0) | 0.094 |
| High grade | 24 (54.5) | 5 (20.8) | 12 (50.0) | 7 (29.2) | ||
| Tumor stage | ||||||
| Ta | 30 (68.2) | 10 (33.3) | 18 (60.0) | 0.75 | 2 (6.7) | 0.008 |
| T1 | 14 (31.8) | 4 (28.6) | 4 (28.6) | 6 (42.9) | ||
| FGFR3 status | ||||||
| Mutated | 22 (50.0) | 5 (22.7) | 16 (72.7) | 0.028 | 1 (4.5) | 0.051 |
| Not mutated | 22 (50.0) | 9 (40.9) | 6 (27.3) | 7 (31.8) | ||
| PIK3CA status | ||||||
| Mutated | 7 (15.9) | 4 (57.1) | 2 (28.6) | 0.18 | 1 (14.3) | >0.99 |
| Not mutated | 37 (84.1) | 10 (27.0) | 20 (54.1) | 7 (18.9) | ||
| TERT status a | ||||||
| Mutated | 35 (81.4) | 10 (28.6) | 18 (51.4) | 0.75 | 7 (20.0) | 0.32 |
| Not mutated | 8 (18.6) | 4 (50.0) | 4 (50.0) | 0 (0.0) | ||
| Parameters | Whole Population | Recurrence-Free Survival | Overall Survival | ||
|---|---|---|---|---|---|
| Number of Patients (%) | Number of Events (%) a | p-Value * | Number of Events (%) b | p-value * | |
| Total population | 59 (100.0) | 39 (66.1) | 34 (57.6) | ||
| Age (years) | |||||
| ≥60 | 43 (72.8) | 33 (76.7) | 0.0046 | 30 (69.8) | 0.002 |
| <60 | 16 (27.2) | 6 (37.5) | 4 (25.0) | ||
| Sex | |||||
| Male | 45 (76.3) | 27 (60.0) | 0.15 | 26 (57.8) | 0.97 |
| Female | 14 (23.7) | 12 (85.7) | 8 (57.1) | ||
| Smoking status c | |||||
| Non-smoker | 9 (17.0) | 7 (77.8) | 0.50 | 7 (77.8) | 0.25 |
| Smoker | 44 (83.0) | 26 (59.1) | 22 (50.0) | ||
| History of NMIBC | |||||
| No | 35 (59.3) | 21 (60.0) | 0.23 | 20 (57.1) | 0.93 |
| Yes | 24 (40.7) | 18 (75.0) | 14 (58.3) | ||
| Associated pTis | |||||
| No | 52 (88.1) | 36 (69.2) | 0.21 | 32 (61.5) | 0.12 |
| Yes | 7 (11.9) | 3 (42.9) | 2 (28.6) | ||
| Tumor stage | |||||
| T2 | 21 (35.6) | 13 (61.9) | 0.61 | 8 (38.1) | 0.024 |
| ≥T3 | 38 (64.4) | 26 (68.4) | 26 (68.4) | ||
| Lymph node status d | |||||
| N− | 37 (64.9) | 20 (54.1) | 0.019 | 17 (45.9) | 0.004 |
| N+ | 20 (35.1) | 17 (85.0) | 17 (85.0) | ||
| FGFR3 status | |||||
| Mutated | 6 (10.2) | 4 (66.7) | >0.99 | 4 (66.7) | >0.99 |
| Not mutated | 53 (89.9) | 35 (66.0) | 30 (56.6) | ||
| PIK3CA status | |||||
| Mutated | 6 (10.2) | 2 (33.3) | 0.17 | 2 (33.3) | 0.39 |
| Not mutated | 53 (89.9) | 37 (69.8) | 32 (60.4) | ||
| TERT status | |||||
| Mutated | 44 (74.6) | 31 (70.5) | 0.23 | 26 (59.1) | 0.70 |
| Not mutated | 15 (25.4) | 8 (53.3) | 8 (53.3) | ||
| Non-Coding Sites | Status | GPR126 | PLEKHS1 | Intron ADM | Chr7:11 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WT | M | Total | p-Value * | WT | M | Total | p-Value * | WT | M | Total | p-Value * | WT | M | Total | p-Value * | ||
| PLEKHS1 | WT | 45 | 28 | 73 | 0.021 | ||||||||||||
| M | 11 | 19 | 30 | ||||||||||||||
| Total | 56 | 47 | 103 | ||||||||||||||
| Intron ADM | WT | 51 | 40 | 91 | 0.35 (NS) | 70 | 21 | 91 | 0.0007 | ||||||||
| M | 5 | 7 | 12 | 3 | 9 | 12 | |||||||||||
| Total | 56 | 47 | 103 | 73 | 30 | 103 | |||||||||||
| Chr7:11 | WT | 53 | 40 | 93 | 0.20 (NS) | 68 | 25 | 93 | 0.15 (NS) | 84 | 9 | 93 | 0.091 (NS) | ||||
| M | 3 | 7 | 10 | 5 | 5 | 10 | 7 | 3 | 10 | ||||||||
| Total | 56 | 47 | 103 | 73 | 30 | 103 | 91 | 12 | 103 | ||||||||
| Chr15:96 | WT | 55 | 40 | 95 | 0.035 | 70 | 25 | 95 | 0.045 | 88 | 7 | 95 | 0.0004 | 86 | 9 | 95 | 0.57 (NS) |
| M | 1 | 7 | 8 | 3 | 5 | 8 | 3 | 5 | 8 | 7 | 1 | 8 | |||||
| Total | 56 | 47 | 103 | 73 | 30 | 103 | 91 | 12 | 103 | 93 | 10 | 103 | |||||
| Gene | Normal Bladder Tissues (n = 19) | Group 1 - Bladder Tumors No non-Coding Mutations (n = 39) | Group 2 - Bladder Tumors One or Two Non-Coding Mutations (n = 46) | Group 3 - Bladder Tumors Three or More Non-Coding Mutations (n = 18) | p-Value * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Median | Min | Max | Median | Min | Max | Median | Min | Max | Median | Min | Max | ||
| APOBEC3A | 1.0 | 0.07 | 13.5 | 0.63 | 0.00 | 17.3 | 0.73 | 0.00 | 7.44 | 0.91 | 0.08 | 18.6 | NS |
| APOBEC3B | 1.0 | 0.00 | 3.55 | 9.26 | 1.68 | 309 | 12.6 | 1.29 | 114 | 23.0 | 1.00 | 78.6 | 0.044 |
| APOBEC3H | 1.0 | 0.00 | 8.49 | 5.67 | 0.78 | 44.8 | 5.21 | 0.00 | 23.3 | 5.88 | 0.32 | 19.8 | NS |
| Parameters | Total Population (%) | Group 1 No Non-Coding Mutation | Group 2 One or Two Non-Coding Mutations | Group 3 Three or More Non-Coding Mutations | p-Value * |
|---|---|---|---|---|---|
| Total population | 103 (100.0) | 39 (37.9) | 46 (44.7) | 18 (17.5) | |
| Age (years) | |||||
| ≥60 | 77 (74.8) | 29 (74.4) | 34 (73.9) | 14 (77.8) | 0.95 (NS) |
| <60 | 26 (25.2) | 10 (25.6) | 12 (26.1) | 4 (22.2) | |
| Sex | |||||
| Male | 86 (83.5) | 36 (92.3) | 40 (87.0) | 10 (55.6) | 0.0017 |
| Female | 17 (16.5) | 3 (7.7) | 6 (13.0) | 8 (44.4) | |
| Smoking status a | |||||
| Non-smoker | 28 (28.9) | 6 (17.1) | 16 (36.4) | 6 (33.3) | 0.16 (NS) |
| Smoker | 69 (71.1) | 29 (82.9) | 28 (63.6) | 12 (66.7) | |
| History of NMIBC | |||||
| No | 59 (57.3) | 27 (69.2) | 20 (43.5) | 12 (66.7) | 0.039 |
| Yes | 44 (42.7) | 12 (30.8) | 26 (56.5) | 6 (33.3) | |
| Associated pTis | |||||
| No | 94 (91.3) | 35 (89.7) | 43 (93.5) | 16 (88.9) | 0.77 (NS) |
| Yes | 9 (8.7) | 4 (10.3) | 3 (6.5) | 2 (11.1) | |
| Tumor stage | |||||
| Cis | 1 (1.0) | 1 (2.6) | 0 (0) | 0 (0) | 0.57 (NS) |
| Ta | 30 (29.1) | 11 (28.2) | 17 (37.0) | 2 (11.1) | |
| T1 | 15 (14.6) | 5 (12.8) | 6 (13.0) | 4 (22.2) | |
| T2 | 19 (18.4) | 8 (20.5) | 8 (17.4) | 3 (16.7) | |
| ≥T3 | 38 (36.9) | 14 (35.9) | 15 (32.6) | 9 (50.0) | |
| FGFR3 status | |||||
| Mutated | 28 (27.2) | 6 (15.4) | 19 (41.3) | 3 (16.7) | 0.015 |
| Not mutated | 75 (72.8) | 33 (84.6) | 27 (58.7) | 15 (83.3) | |
| PIK3CA status | |||||
| Mutated | 13 (12.6) | 2 (5.1) | 7 (15.2) | 4 (22.2) | 0.15 (NS) |
| Not mutated | 90 (87.4) | 37 (94.9) | 39 (84.8) | 14 (77.8) | |
| TERT status b | |||||
| Mutated | 79 (76.7) | 25 (65.8) | 38 (82.6) | 16 (88.9) | 0.082 (NS) |
| Not mutated | 23 (22.3) | 13 (34.2) | 8 (17.4) | 2 (33.3) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vacher, S.; Suybeng, V.; Girard, E.; Masliah Planchon, J.; Thomson, G.; Le Goux, C.; Garinet, S.; Schnitzler, A.; Chemlali, W.; Firlej, V.; et al. Genomic Instability Signature of Palindromic Non-Coding Somatic Mutations in Bladder Cancer. Cancers 2020, 12, 2882. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12102882
Vacher S, Suybeng V, Girard E, Masliah Planchon J, Thomson G, Le Goux C, Garinet S, Schnitzler A, Chemlali W, Firlej V, et al. Genomic Instability Signature of Palindromic Non-Coding Somatic Mutations in Bladder Cancer. Cancers. 2020; 12(10):2882. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12102882
Chicago/Turabian StyleVacher, Sophie, Voreak Suybeng, Elodie Girard, Julien Masliah Planchon, Grégory Thomson, Constance Le Goux, Simon Garinet, Anne Schnitzler, Walid Chemlali, Virginie Firlej, and et al. 2020. "Genomic Instability Signature of Palindromic Non-Coding Somatic Mutations in Bladder Cancer" Cancers 12, no. 10: 2882. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12102882