NRF2 Regulation by Noncoding RNAs in Cancers: The Present Knowledge and the Way Forward


Laboratory of Oncology, Fondazione IRCCS Casa Sollievo della Sofferenza, San Giovanni Rotondo, 71013 Foggia, Italy
*
Author to whom correspondence should be addressed.
Cancers 2020, 12(12), 3621; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123621
Submission received: 27 October 2020
/
Revised: 23 November 2020
/
Accepted: 30 November 2020
/
Published: 3 December 2020
(This article belongs to the Special Issue The KEAP1-NRF2 Pathway in Cancer)
Abstract
:Simple Summary
The NRF2 pathway represents one of the most intriguing pathways that promotes chemo- and radioresistance of neoplastic cells. Increasing findings suggest that the NRF2 signaling can be modulated by multiple epigenetic factors such as noncoding RNAs, which influence a large number of oncogenic mechanisms, both at transcriptional and at post-transcriptional levels. As a consequence, the identification and characterization of specific noncoding RNAs as biomarkers related to oxidative stress may help to clarify the relationship between them and NRF2 signaling in the tumor context, in terms of positive and negative modulation, also referring to their intersection with other NRF2 crosstalking pathways. In this review, we summarize the recent updates on NRF2 network regulation by noncoding RNAs in tumors, thus paving the way toward the potential translational role of these small RNAs as key tumor biomarkers of neoplastic processes.
Abstract
Nuclear factor erythroid 2-related factor 2 (NRF2) is the key transcription factor triggered by oxidative stress that moves in cells of the antioxidant response element (ARE)-antioxidant gene network against reactive oxygen species (ROS) cellular damage. In tumors, the NRF2 pathway represents one of the most intriguing pathways that promotes chemo- and radioresistance of neoplastic cells and its activity is regulated by genetic and epigenetic mechanisms; some of these being poorly investigated in cancer. The noncoding RNA (ncRNA) network is governed by microRNAs (miRNAs) and long noncoding RNAs (lncRNAs) and modulates a variety of cellular mechanisms linked to cancer onset and progression, both at transcriptional and post-transcriptional levels. In recent years, the scientific findings about the effects of ncRNA landscape variations on NRF2 machines are rapidly increasing and need to be continuously updated. Here, we review the latest knowledge about the link between NRF2 and ncRNA networks in cancer, thus focusing on their potential translational significance as key tumor biomarkers.
1. Introduction
The NRF2 protein, encoded by the Nuclear factor erythroid 2-related factor 2 (NFE2L2) gene, is a transcription factor endogenously expressed in eukaryotic cells and represents the main protector of the cellular antioxidant and cytoprotective response to harmful insults, as well as to xenobiotic damage and oxidative stress [1].
The activity of NRF2 is strictly dependent on a battery of transcriptional modulators that govern its physiologic activity under basal conditions as well as under internal and external stimuli arising from an oxidative stress condition [2]. In normal state, NRF2 orchestrates the maintenance of basal expression levels of more than 200 target antioxidant response element (ARE) genes through its direct link to this specific consensus sequences located at their promoter regions [3]. During the cellular adaptation to environmental modifications, the activation of NRF2 signaling is triggered by competitive interactions with the three ubiquitin ligase complexes—Cullin 3-RING-box protein-Kelch-like ECH-Associated Protein 1 (CUL3-RBX1-KEAP1), Skp1-cullin-F-box protein-transducin repeat-containing proteins (SCF/β-TrCP) and ERAD-associated E3 ubiquitin-protein ligase (HRD1). This complex interaction controls the ubiquitination and proteasomal degradation of NRF2 and, as a consequence, its abundance in specific subcellular compartments [4]. When NRF2 moves into the nucleus, it specifically binds to the ARE gene regions by heterodimerizing with small MAF (sMAF, avian musculo aponeurotic fibrosarcoma oncogene homolog) proteins and turning on the transcription of a large number of antioxidant and detoxification genes [5,6].
The main well-characterized mechanism of NRF2 regulation is tightly linked to its negative repressor, the KEAP1 protein. The KEAP1 is able to form an ubiquitin ligase complex with CUL3 and RBX1 to bind the NRF2 and enhancing its proteolitic degradation (in normal cell conditions) or detach from NRF2, because of KEAP1 cysteine modification (upon stress exposure) and favoring its nuclear translocation [7,8,9]. A KEAP1-independent modulation of the NRF2 signaling pathway may also occur both at transcriptional, post-transcriptional and at post-translational levels, with a consequent modification of its cellular localization, protein folding/stability and its DNA-binding ability [10,11].
Over the recent decades, the NRF2 transcription factor has been found to be overexpressed in various human disease, and a growing number of studies have identified abnormal NRF2 functions that go over the physiologic stress-regulating processes, including cancerous processes [12]. Fascinating scientific evidence indicates that the potential double role of NRF2 in cancer suppression and promotion [12,13] is under the control of genetic and epigenetic events, the latest ones strictly related to the activity of ncRNAs, such as microRNAs (miRNAs) and long noncoding RNAs (lncRNAs) [14,15]. These small pieces of RNAs are noncoding nucleotides that modulate transcriptional and post-transcriptional variations of oncogene and tumor suppressor genes, also not in physiologic processes such as cell differentiation, proliferation, cell cycle regulation, self-renewal capacity of stem cells and apoptosis, but also in the chemo- and radioresistance ability of tumors [16,17].
In the present review, we summarized the mechanisms by which ncRNAs affect NRF2 activity, their relationship with NRF2 regulators, their effects on ARE gene expression and other protein of different signaling pathways related to NRF2 activity.
2. Brief Overview of the NRF2 Signaling Pathway: Negative and Positive Regulators and ARE Genes’ Targets
The NRF2 transcription factor was firstly described thanks to its ability to bind to the NF-E2 site in the β-globin gene cluster and belongs to the cap’n’collar (CNC) subfamily of the basic region-leucine zipper (bZip) transcription factors (TFs) [18]. NRF2 protein counts seven NRF2-ECH homology domains, namely Neh1-Neh7, that are a highly conserved regions having different functions such as resistance to oxidative stress, autophagy, inflammation and inflammasome signaling modulation, apoptosis, mitochondrial biogenesis and stem cell maintenance [19].
The well-characterized mechanism that contributes in the regulation of NRF2 protein level is tightly linked to its negative repressor, Kelch-like ECH-associated protein 1 (KEAP1), which is able to modulate the cytoplasmatic levels of NRF2 by stimulating its proteolytic demolition or inhibiting this process thanks to the modification of NRF2 cysteine residues depending on cell status [7,8,20]. In normal cell conditions, NRF2 is kept in the cytosol by binding a dimer of KEAP1 protein, the main cysteine-rich protein that acts as a key sensor for oxidative/electrophilic stress molecules [21,22]. The KEAP1 guides NRF2 into the E3 ligase complex by means of bric-a-brac (BTB) and Kelch domains, which physically interact with CUL3 and NRF2. After the binding between NRF2 and E3 complexes, a polyubiquitination occurs at seven NRF2 lysine residues located in the Neh2 domain, followed by an ubiquitin-dependent degradation by the 26S proteasome [22,23]. Upon stimuli induction, the inactivation of KEAP1 occurs. As a result of this, the NRF2 translocates into the nucleus and dimerizes with sMAF proteins to bind the promoter region of several target antioxidants genes through their ARE consensus sequences [24].
In addition to the above-described master KEAP1 modulation, it has also been demonstrated that several complexes such as SCF/β-TrCP (S-phase kinase-associated protein 1 (SKP1), Cullin-1 (CUL1) and F-box protein E3 ubiquitin ligase are fundamental for the NRF2 stabilization process [25]. It has also been proved that p21 (CDKN1A), a well-known p53-downstream gene, can directly bind to ETGE and DLG motifs of NRF2, thus avoiding the interaction with KEAP1 and facilitating the NRF2 ubiquitination process [11]. Interestingly, the cyclin kinase inhibitor p21 and NRF2 are linked through a feedback mechanism, thanks to which it has been shown that the increase in p21 might increase the NRF2 activity [26]. Breast cancer type 1 susceptibility (BRCA1) protein also directly interacts with NRF2 and their binding impairs KEAP1-mediated NRF2 ubiquitination process, which is essential for NRF2’s stability and activation [27]. Lastly, in-depth investigations showed that the p62, also known as sequestosome 1 (SQSTM1), can mediate a selective autophagic KEAP1 degradation. These events trigger NRF2 through a positive feedback loop via p62/NRF2/KEAP1 pathway interactions, thus stimulating the transcription of several NRF2 ARE target genes [28,29].
3. NRF2 Deregulation in Cancer: Focus on the Epigenetic Modifications
Cancer development and progression depends on multiple genetic and epigenetic alterations. Specifically, the epigenetic processes are mechanisms that alter the expression level of the genes without modifying the sequence of DNA nucleotides. They include several changes in DNA methylation patterns, histone modifications and small noncoding microRNAs (miRNAs) and long noncoding RNAs (lncRNAs) expression [30].
A large amount of scientific evidence suggested that NRF2 favors the survival of normal cells as well as of cancer cells, thus corroborating the idea that its activation might promote the neoplastic progression. The identification of a “dark side of NRF2” has been debated over the time, but it still appears quite uncertain, since NRF2 could act both as a tumor suppressor and oncogene [12,13]. As for many genes, the occurrence of genetic and epigenetic modifications affecting the NRF2 pathway promotes cancer-related molecular events such as tumor initiation, growth, invasion and metastasis. The main interesting effect of all reported genetic/epigenetic alterations of the NRF2 pathway is a potential translational impact in terms of patients’ survival and response to chemo-radio and targeted therapies, firstly reported in lung tumors [31,32,33,34,35]. The biallelic inactivation of the KEAP1 gene was reported to have a great impact on the upregulation of NRF2 and was firstly described in non small cell lung cancer (NSCLC) and then widely notified in many other solid tumors [36,37,38,39]. Point missense mutations of NFE2L2 gene were also reported to have a similar effect on the KEAP1/NRF2 binding affinity and frequently recurred not only in lung cancer [40], but also in head and neck carcinoma [41], hepatocellular carcinoma (HCC) [42], papillary renal cell carcinoma (PRCC) [43] as well as esophageal and skin cancers [44]. In lung squamous carcinoma (LUSC) and head and neck cancers, an alternative splice variant of NFE2L2 gene lacking exon 2 was described to play a significant role in the loss of interaction of NRF2 with the Kelch domain of KEAP1. This in turn caused the stabilization of NRF2 and the induction of its transcriptional response [45]. Epigenetic modifications have been widely described to impact on the modulation of the KEAP1/NRF2 system in cancer. Aberrant methylation at the promoter island of the KEAP1 gene has been widely reported as one of the most important mechanisms of KEAP1 silencing in solid tumors, such as glioma, breast, prostate, colorectal, thyroid cancers, clear renal cell carcinoma and lung cancer and was linked to tumor development, chemoresistance and mortality risk [14]. More recently, in NSCLC Kirsten rat sarcoma viral oncogene homolog (KRAS) wild-type subpopulations, a novel and significant correlation was observed among promoter and in intragenic exon 3 cytosine-guanine dinucleotide island (CpG-I) methylation and the transcription levels of KEAP1, NFE2L2 and many ARE genes [46,47].
Given the real complexity and multiplicity of the cellular processes controlled by NRF2, one of the most recent and attractive pieces of evidence suggests that the regulation NRF2 can be also guided by the ncRNAs, such as miRNAs and lncRNAs [48]. The following two main sections will summarize the findings about ncRNAs related to oxidative stress and their relationship with the NRF2 network.
4. LncRNAs Intercepting the NRF2 Axis
LncRNAs are noncoding RNA molecules not more than 200 nucleotides in length involved in the inhibition of gene expression as well as in the regulation of many different physiological mechanisms such as cell differentiation, cycle regulation, apoptosis and proliferation [49,50].
lncRNAs are characterized by higher stage and cell subtype specificities in various tumors. Since lncRNAs participate in several cells signaling pathways and lead to the alteration of tissue-specific pattern of expression, their importance is related to the development of novel strategies for specific cancer subtype diagnosis and targeting. The modulation of a large number of lncRNAs, which could be up- or downregulated in several cancers, is important to determine their tissue specificity and for linking them to tumor stage [51].
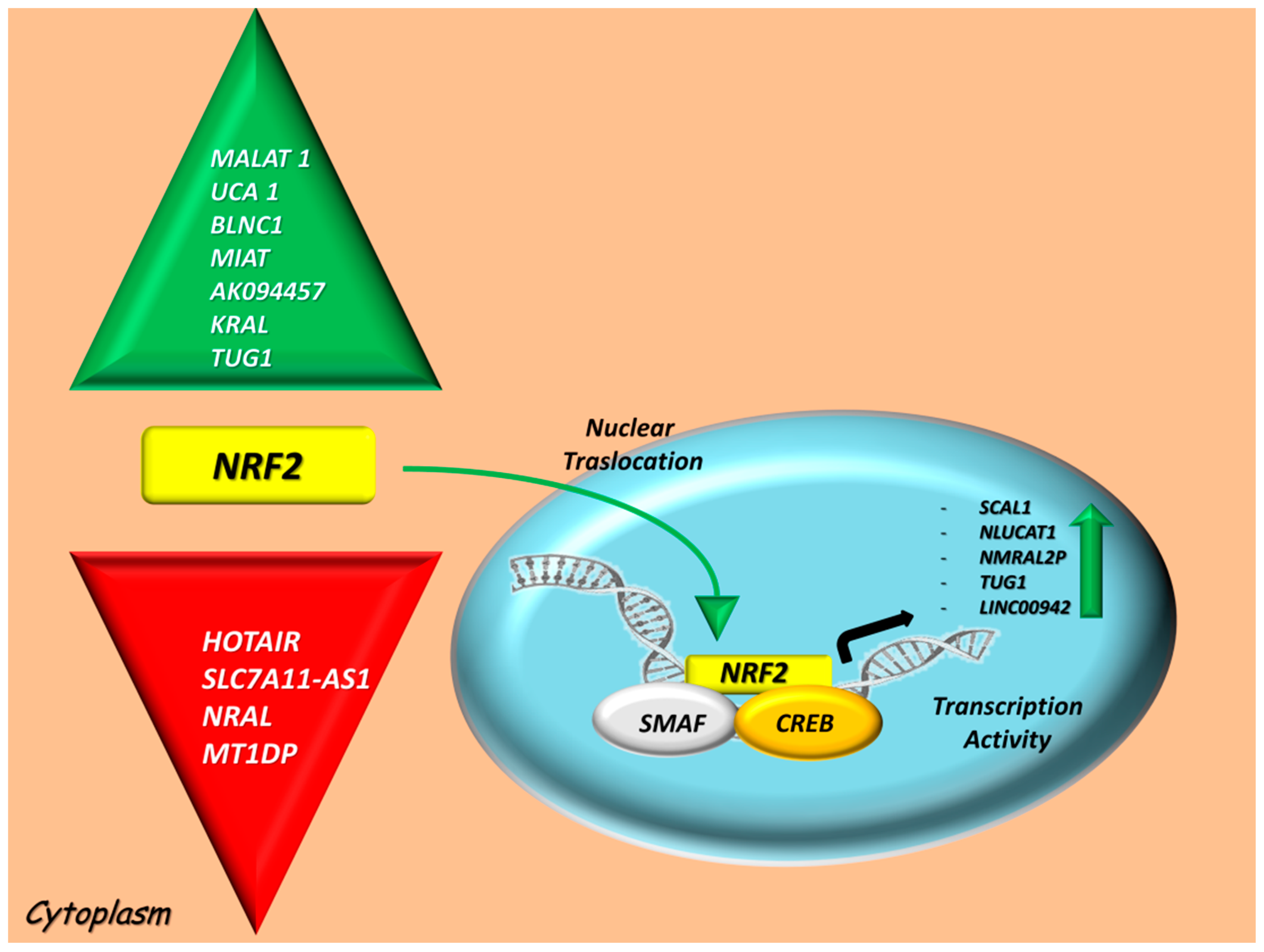
LncRNAs may also play a pivotal role in the onset, progression and stem cell pluripotency of cancer cells [52] and in the oxidation/antioxidant system by acting as negative or positive regulators via NRF2 interactions [53]. According to these, we identified two different groups of lncRNAs linked to the NRF2-oxidative stress system: transcriptional targets and regulators that are summarized in Figure 1.
4.1. LncRNAs Modulated by NRF2 Signaling
Emerging reports on NRF2-associated lcRNAs described a large number of identified transcriptional targets of NRF2, but only a small part of these were functionally validated. The most well-characterized LncRNAs whose transcription were directly modulated by NRF2 include SCAL1 (LUCAT1), nuclear smoke- and cancer-associated lncRNA (NLUCAT1), NmrA-like redox sensor 2 pseudogene (NMRAL2P), Taurine-upregulated gene 1 (TUG1), and long intergenic nonprotein coding RNA 942 (LINC00942) and are listed in Table 1.
4.1.1. SCAL1 (LUCAT1)
The smoke- and cancer-associated lncRNA 1 (SCAL1 or LUCAT1) is a cancer lncRNA related to smoke exposure that is strictly associated with NRF2 activity. The NRF2 binds the SCAL1 promoter region through a putative nuclear factor-erythroid-2 (NFE2) binding site, thus suggesting that this lncRNA could be transcriptionally regulated by NRF2. On the other hand, the silencing of SCAL1 in human bronchial epithelial cells (HBE) induces a significant enhancement of cytotoxicity under the cigarette smoke extract (CSE) exposure. These findings suggested that SCAL1 may be considered as a downstream mediator of NRF2 against reactive oxygen species (ROS)-induced oxidative stress in airway epithelial cells [54]. Additionally, a transcriptomic analysis in a large cohort of adenocarcinoma and squamous cell carcinoma tumors of the lung showed that SCAL1 correlated with KEAP1 and NFE2L2 mutational statuses [55].
4.1.2. NLUCAT1
The NLUCAT1 is a nuclear transcript variant of LUCAT1 that is composed of six exons, which are under the control of NRF2 transcription factors. In the A549 lung adenocarcinoma (LUAD) cell line, it was observed that the targeted deletion of NLUCAT1 led to a decrease in cell proliferation and invasion, increasing the oxidative stress and cisplatin-induced NRF2-mediated apoptosis. More interestingly, a recent transcriptomic analysis revealed that NLUCAT1 may exert a positive loop on the NRF2 network [56].
4.1.3. NMRAL2P
NmrA-like redox sensor 2 pseudogene (NMRAL2P) is the first functional pseudogene that was identified as a direct target of NRF2 and downstream regulator of NRF2-dependent NQO1 activation in sulforaphane (SFN)-treated colon cancer cells. By using functional and pharmacological assays, it was observed that siRNA-mediated knockdown of NFE2L2 gene was able to interfere with the ability of SFN to induce NMRAL2P expression [57].
4.1.4. TUG1
Taurine-upregulated gene 1 (TUG1) has been reported to be involved in different pathogenic cellular mechanisms, as well as carcinogenesis and chemoresistance of cancer cells [58]. In prostate and esophageal squamous cell carcinoma (ESCC), TUG1 directly binds NRF2, thus upregulating its expression at protein level together with its downstream members (HO-1 and NQO1) [76]. The silencing of TUG1 could offer an interesting point of view aimed at overcoming drug resistance in ESCC using NRF2 inhibitors [73]. Moreover, new recent evidence also suggested a synergic and oncogenic role of NRF2 and TUG1 in stimulating proliferation, apoptosis, migration and resistance to doxorubicin treatment in urothelial carcinoma of the bladder (UCB) cells [59].
4.1.5. LINC00942
Little evidence is now available on the role of long intergenic nonprotein coding RNA 942 (LINC00942) in the modulation of the NRF2 antioxidant pathway. Transcriptomic analyses demonstrated that LINC00942 could directly interact with NRF2 through its binding to the NFE2L2 promoter region, thus decreasing the Glutamate-Cysteine Ligase Catalytic Subunit (GCLC) mRNA and protein levels. This observation has been linked to the fact that human tumors having NFE2L2 gain-of-function mutations showed an increased expression of LINC00942 [60].
4.2. Positive and Negative LncRNA Regulators of the NRF2 Activity
This group of LncRNA participates in the recruitment of chromatin modifiers [77] or transcription factors [16] and can modulate post-transcriptional events, as well as the translational repression and/or the enhance mRNA degradation, or serve as natural “miRNA sponges” through sequestration of miRNAs [78,79]. They are also involved in the modulation of the oxidation/antioxidant system by acting as negative or positive regulators via NRF2 interactions, as explained below and summarized in Table 1.
4.2.1. HOTAIR
The first evidence about the regulation of NRF2 expression by Hox transcript antisense intergenic RNA (HOTAIR) was provided by Zhang and coworkers, who demonstrated that HOTAIR regulates NRF2 levels via mediating histone H4 acetylation at the NFE2L2 gene promoter in Gc1-Spg (germ cell-1 spermatogonial) cells. Once the silencing of HOTAIR occurred, a positive correlation between the downregulation of NRF2 levels and H4 acetylation within the NFE2L2 promoter was observed [61]. In addition, it is hypothesized that HOTAIR participates in the epigenetic suppression of the NRF2-multidrug resistance-associated protein 2/4 (MRP2/4) pathway and stimulates the transcription of a lot of ARE genes [62].
4.2.2. MALAT1
A cooperation between the KEAP1/NRF2 axis and the lncRNA metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) was recently documented in human umbilical vein endothelial cells (HUVECs). Under hydrogen peroxide exposure, the expression of MALAT1 stimulates the downregulation of KEAP1 at mRNA and protein levels, thus leading the NRF2 nuclear accumulation and ARE target gene activation [63]. MALAT1 can also directly bind NRF2, thus activating the NRF2 signaling cascade and ARE-related gene upregulation [64]. Moreover, MALAT1 also interacts with Polycomb repressive complex 2 (PRC2) components, including Enhancer of zeste homolog 2 (EZH2). In fact, Li and colleagues showed in lung cancer cells that EZH2 targets NFE2L2 promoter, thus suppressing its expression by regulating trimethylation of histone H3 at lysine 27 (H3K27me3) [80].
4.2.3. UCA1
The Urothelial carcinoma-associated 1 (UCA1) lncRNA was associated with miR-495 activity, which targets and negatively regulates it in renal cell carcinoma, after the epigenetic repression of p21 [65]. Li and colleagues showed that a high expression of UCA1 could develops cisplatin resistance in lung cancer cells via UCA1/miR-495/NRF2 crosstalk [66]. In hippocampal tissues and neurons, UCA1 activates NRF2-dependent cytoprotective pathway by sponging miR-495 and, overall, leads to an inhibition of apoptosis during seizure-induced brain injury [67].
4.2.4. BLNC1
LncRNA BLNC1 was found overexpressed in diabetic nephropathy (DN) and contributes to attenuate chemical and physical insults, inflammation and renal fibrosis by hyperactivating the NRF2/HO-1 pathway in renal HK-2 proximal tubules kidney cells. Since the BLNC1 could also inhibit the activation of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway, it was suggested that the crosstalk between NRF2/HO-1 and NF-κB pathways might serve as a potential therapeutic target against DN [81].
4.2.5. MIAT and AK094457
The lncRNA Myocardial infarction-associated transcript (MIAT) targets NRF2 by mediating the high glucose-induced tubular injury in renal HK-2 epithelial cells and modulates cell viability in proximal tubules in renal via balancing NRF2 levels [68].
The downregulation of AK094457 was found to be inversely correlated to the upregulation of NRF2 and HO-1 in vascular smooth muscle cells (VSMCs) and this may suggest how both are involved in antioxidant mechanisms [69].
4.2.6. SLC7A11-AS1
Recent evidence about the relationship between Solute Carrier Family 7 Member 11-antisense 1 (SLC7A11-AS1) and post-transcriptional regulation of NRF2 via protein–protein interactions was reported in pancreatic cancer. The SLC7A11-AS1 was observed to hamper with the stability of NRF2, thus impairing the E3 Ub-ligases processes as well as targeting proteins for degradation and controlling protein turnover by modification of ubiquitin–proteasome system (UPS)-related proteins. In pancreatic ductal adenocarcinoma cells (PDACs), it was observed that the block of SCFβ-TRCP-mediated degradation of NRF2 produced the reduction in intracellular ROS and stimulated stemness and chemoresistance properties [70].
4.2.7. KRAL and NRAL
The dual and opposite roles of NRF2 regulation-associated lncRNA (NRAL) and KEAP1 regulation-associated lncRNA (KRAL) have been recently outlined in hepatocellular carcinoma (HCC) cell lines. KRAL activation promotes chemo-sensitization to HCC cells by targeting the miR-141/KEAP1 axis as a competing endogenous RNA (ceRNA), with a consequent upregulation of KEAP1 and a silencing of NFE2L2, respectively [71]. On the other hand, NRAL inhibits the miR-340-5p/NRF2 crosstalk by exerting a transcriptional repression through an interaction with the NFE2L2 3′-untranslated (3′UTR) regions and this causes an increase in its expression and drug resistance on HepG2 cells [71,72].
4.2.8. MT1DP
An intriguing hypothesis about a potential correlation between metallothionein 1D pseudogene (MT1DP) lncRNA and NRF2 was provided by Gao and colleagues. The main finding was that MT1DP sustained the activation of miR-365 through a 3′UTR consensus site, which in turn, is able to repress the NFE2L2 gene and impair cadmium-stimulated oxidative stress [74]. More recently, it was reported that MT1DP weakened the expression of NRF2 and enhanced the sensitivity of NRF2-overexpressed NSCLC cells to erastin-induced ferroptosis by stabilizing miR-365a-3p [75].
5. miRNAs Intercepting the NRF2 Axis
miRNAs are proximately 22 nucleotides single-stranded noncoding RNA molecules able to modulate the translation or stability of mRNA molecules through interaction with specific mRNAs with complementary base sequences [82]. In human cancer, the miRNA machinery is deregulated by several mechanisms and impacts on the tumorigenic activities of neoplastic cells mainly promoting cell invasion and metastasis [83].
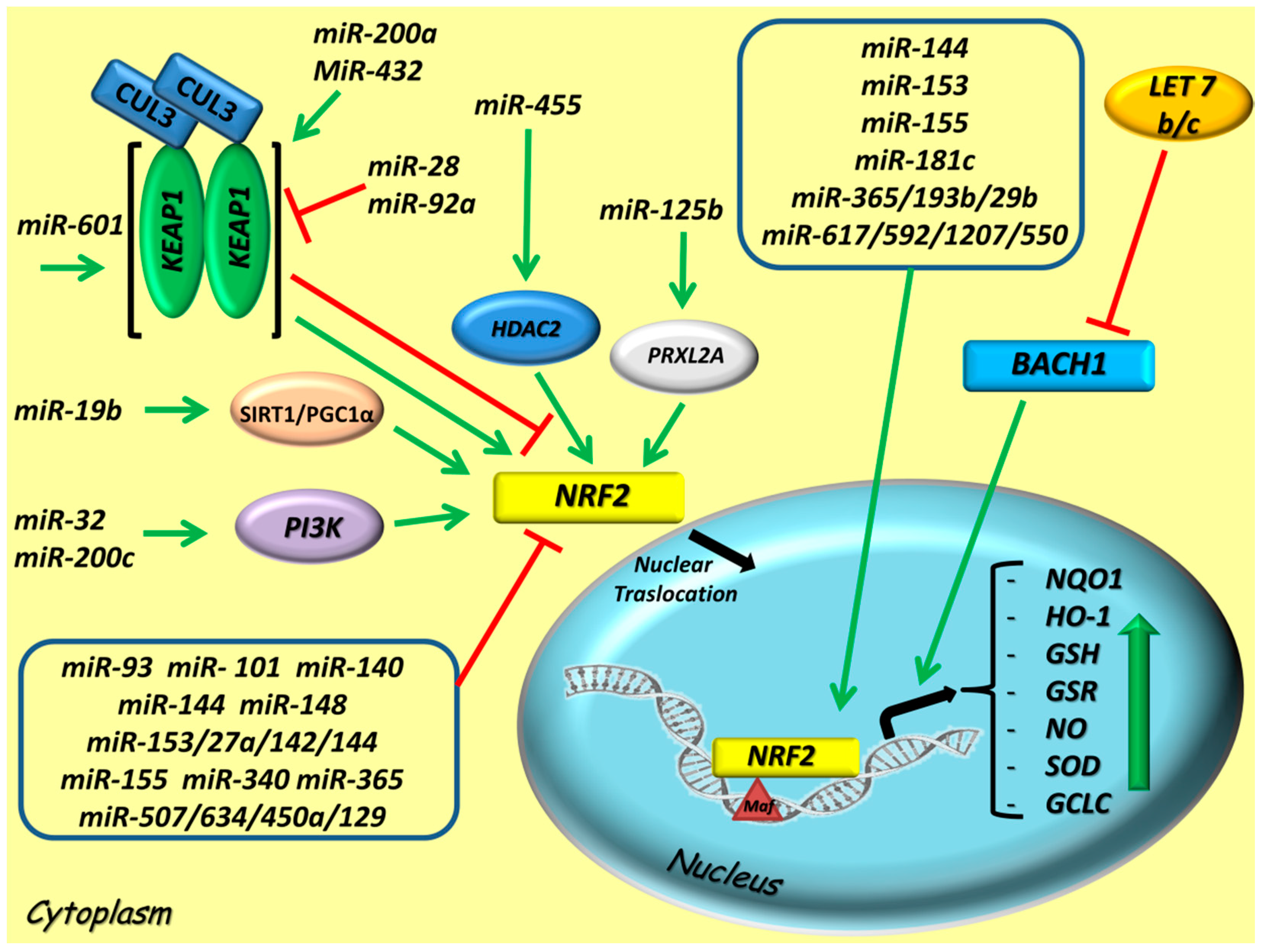
The exact molecular mechanisms by which miRNAs can modulate the antioxidant defense system in cells are only partially understood. However, the most recent scientific evidence suggests that several miRNAs crosstalking with the NRF2 signaling pathway by directly regulating the expression of NRF2, by influencing the nuclear translocation of NRF2 or by indirectly modulating KEAP1 and other upstream mediators of the NRF2 pathway [84,85] (Figure 2).
5.1. Positive miRNA Regulators of NRF2 Activity
The first miRNAs reported to exert a positive stimulation on NRF2 transcription were let 7b and let 7c, which surely regulate the transcription factor BTB and CNC homology 1 (BACH1) expression in human hepatoma Huh-7 and liver cells. As a result, an increase of HO-1 expression mediated by NRF2 and an attenuation of the oxidative stress mechanism were demonstrated [86,87].
In LO2 liver cells, a reduction in miR-19b levels was observed under acetaminophen (APAP) treatment. This, in turn, leads to the stimulation of sirtuin-1 (SIRT1), which activates the NRF2 cascade and its related downstream target genes and enhances drug hepatotoxicity [88].
miR-32 contributes to the activation of Phosphoinositide 3-kinase (PI3K) in prostate cancer which, in turn, upregulates NRF2. In this way, NRF2 creates a positive feedback loop by promoting survival in cultured human retinal pigment epithelium (RPE) cells [89].
In oral squamous cell carcinoma (OSCC), it was observed that the overexpression of the Peroxiredoxin-like 2A (PRXL2A) gene was induced by miR-125b, which successively suppresses the oxidative stress damage driven by positive feedback loops involving the NRF2 signaling pathway [90].
In neuroblastoma SH-SY5Y cells, the transfection experiments with a miR-144 mimic was seen to accelerate apoptosis and reduced the expression of several enzymes regulated by NRF2 that are involved in glutathione (GSH) synthesis and reactive oxygen species scavenging [91].
Evidence of the link between miR-153-3p and NRF2 emerged in breast cancer cell lines, where lower miR-153-3p expression levels correlated to an increase in NRF2 and downstream gene levels with a consequent stimulation of tumor cell migration and invasion [92].
In lung cancer cell lines it has been found that miR-155 confers resistance to arsenic trioxide (ATO) by activating the NRF2 signaling pathway. The consequent upregulation of HO-1 and NQO1 inversely correlated with a relative decrease in apoptosis, thus promoting the enhancement of the B-cell lymphoma 2/BCL2-Associated X protein (BCL-2/BAX) ratio [93].
Bioinformatics predictions coupled with functional analysis revealed the presence of a miR-181c binding site in the 3′UTR of NFE2L2, thus enhancing the activation of multiple pro-survival pathways.
The family of miR-200 may play an important role in the KEAP1-dependent mechanism of NRF2 regulation. It has been demonstrated that miR-200a stimulates the NRF2 signaling pathway by suppressing KEAP1, mainly due to the decrease in ROS concentration, thus ensuring the survival of osteoblasts upon dexamethasone treatment in osteoblast (OB)-6 cells [94].
It is well-known that the Zinc Finger E-Box Binding Homeobox 1 (ZEB1) protein could inhibit E-cadherin, thus promoting the epithelial mesenchymal transition (EMT) and the overexpression of NRF2 [95]. miR-200c was also observed in both breast and ovarian cancers to induce AKT upregulation. Given that the upstream regulation of PI3K/AKT pathway activates NRF2 and its related antioxidant genes, the downregulation of miR-200c by NRF2 could potentially induce a negative feedback mechanism [89].
miR-365, miR-193b and miR-29b are also involved in the scenario of cancer NRF2-related deregulation. Enforced expression of miR-29b occurs in response to NRF2 activation [96,97]. The presence of a feedback loop between NRF2 and miR-29b was suggested by Chan and colleagues, who firstly described the localization of the Sp1 transcription factor binding site at the NFE2L2 promoter region. NRF2 could suppress miR-29b and induce an upregulation of both SP-1 and NRF2, thus protecting cells from apoptosis [98,99].
Evidence of a positive modulation of NRF2 by miR-432-3p was reported in esophageal squamous cell carcinoma (ESCC). By using ARE reporter plasmids, miR-432-3p was supposed to promote the NRF2 protein stabilization by directly binding the KEAP1 coding region, thus increasing the resistance of cancer cells to cisplatin (CDDP) [100].
miR-455-3p activates NRF2 via its upstream mediator, the histone deacetylase 2 (HDAC2), thus ensuring more protection to osteoblasts against oxidative injury by activating NQO1, HO-1 and GCLC expressions [101,102].
miR-601 targets CUL3 and contributes to the activation of the NRF2 signaling pathway in protecting retinal pigment epithelium (PRE) cells against oxidative damage. The study of the effects of miR-601 expression on CUL3 and NRF2 signaling revealed that this miRNA could participate in a dependent CUL3-KEAP1-NRF2 activation mechanism, thus suggesting a possible therapeutic strategy for treatment or prevention of Age-related macular degeneration (AMD) [103].
Finally, the group containing miR-617, miR-592, miR-1207 and miR-550 was recently correlated to the NRF2 activity in different cancer profiles. Specifically, it was found that NRF2 upregulates miR-617, thus protecting the esophageal cancer cells from chemical and physical insults. It has also been observed that NRF2 is involved in a drug resistance mechanism in colorectal and liver carcinoma, thus suggesting a possible link between the reduction in miR-592 levels and NRF2 activation in these tumors [104,105]. In prostate cancer, a concomitant and synergic upregulation of miRNA-1207 and NRF2 was demonstrated [106].
Likewise, little is known about miR-550. The bioinformatics DIANA-microT V3.0 tool was used to identify downstream NRF2 targets, such as tumor suppressor BAF Chromatin Remodeling Complex Subunit BCL11B (BCL11B), which correlates to miR-550 activity, thus corroborating the importance of NRF2 upregulation and promoting cell growth and proliferation [107].
miRNA-positive regulators of NRF2 signaling are presented in Table 2.
5.2. Negative miRNA Regulators of NRF2 Activity
The first investigated NRF2-related miRNA in MCF-7 breast cancer cells was miR-28, which exerts a negative effect on NRF2, independently from its master regulator, the KEAP1 protein [109].
An inverse relationship was proposed for miR-92a expression, whose suppression induces the activation of the NRF2-KEAP1-ARE pathway in cultured human umbilical vein endothelial cells (HUVECs). By luciferase assays, it has been demonstrated that miR-92a was able to promote cell proliferation, thus contributing to a decrease in apoptosis-inducing factors such as caspase 3, tumor necrosis factor (TNF)-α and KEAP1 expression with an upregulation of NRF2 and many of its ARE target genes [110].
Singh and colleagues suggested that miR-93 could act as an oncogenic negative regulator of NRF2 activity by targeting specific sites at the 3′UTR region in MCF-10A breast cancer cells. To prove the tumor-enhancer effect of miR-93 mediated by NRF2 signaling, the authors proved an inverse correlation between an inhibition of miR-93 and an increased expression of NRF2 with a consequent significant reduction in mammosphere propagation, apoptosis and DNA damage upon 17β-estradiol treatment [111].
Interesting evidence also came from the study on miR-101 in breast cancer. The miR-101 binds to the 3′UTR region of NFE2L2, thus influencing its expression, promoting the suppression of cell proliferation and enhancing the sensitivity of cancer cells to ROS [112].
NRF2/miR-140 crosstalk may play an important role in major protection in fibrotic lung tissues against transforming growth factor (TGF)-β1-mediated inflammation and fibroblast differentiation induced by radio treatment [113,114].
The question looks intriguing when we looked at several studies focusing on miR-144. This miRNA was seen to be upregulated not only in the peripheral blood of acute myeloid leukemia (AML) affected patients but also in AML HL-60 cells, thus suggesting that its inhibition can promote apoptosis and suppress NRF2 activation [115]. Of interest, the molecular link between NRF2 and miR-144 correlates to an enhancement of toxicity of 5-fluorouracil (5-FU) in hepatocellular cancer cell lines [116].
Qu and colleagues recently proposed a novel mechanism of NRF2 regulation in response to oxidative stress in endometrial cancer cells via miR-148b. They observed that HIF-1 and NRF2 expressions were significantly reduced upon miR-148b overexpression; endoplasmic reticulum metalloprotease 1 (ERMP1) related and suggested a tumor-suppressive role of this miRNA in RL95-2 cells of human endometrial cancer [117].
Another piece of evidence of a negative regulation of NRF2 by miRNAs was provided by Wang and colleagues, when they observed that miR-153 acts as an oncogenic effector of the apoptosis, thus reinforcing the proliferation ability of breast cancer cells via suppression of NRF2 and its downstream ARE genes’ expression [92]. Finally, alternative approaches, such as in silico and in vitro models of cancer cell lines, added novel molecular insights and elucidated the link between NRF2 and miRNAs. A group of novel miRNAs (miR-153/miR-27a/miR-142-5p/miR-144) was identified as exerting a modulator effect on NRF2 expression and on oxidative stress balance in neuronal cells. The upregulation of this group of miRNAs resulted in suppressing NRF2 signaling, thus turning off both glutamate-cysteine ligase catalytic (GCLC) and glutathione-disulfide reductase (GSR) expressions [118].
In bronchial epithelial cells, it was demonstrated that, under arsenic treatment, the activity of miR-155 directly accelerates apoptosis by blocking NRF2 activity and those of related cytoprotective genes as well as glutathione (GSH), nitric oxide (NO) and superoxide dismutase (SOD) [119]. Moreover, a notable correlation between the overexpression of miR-155 and the radiation-induced pulmonary fibrosis by NRF2 was reported in a mouse model [120].
In human neuroblastoma SH-SY5Y cells, the miR-181s are able to suppress the Sirtuin 1/Peroxisome proliferator-activated receptor gamma coactivator 1-alpha/NRF2 (SIRT1/PGC-1α/NRF2) signaling pathway [121], whereas in hepatocellular carcinoma patterns it was shown that miR-340 targets the NRF2 pathway and alters the chemoresistant phenotype of cells after cisplatin treatment [122].
The molecular bridge between metallothionein 1D pseudogene (MT1DP) and NRF2 was linked to the miR-365 activity in human hepatocellular carcinoma cell line HepG2. Upon cadmium-induced oxidative stress, the upregulation of miR-365 acts as a sensor for MT1DP activation, thus thereby repressing the NRF2 activity via direct binding to its 3′UTR [74].
Lastly, miR-507, miR-634, miR-450a and miR-129-5p were shown to negatively modulate to NRF2 oncogenic activity by directly targeting NFE2L2 in the neuroblastoma SH-SY5Y cell line [123], whereas it was demonstrated that they could exert a synergic effect of increasing sensitivity to cisplatin treatment in NSCLC A549 cells [124].
As follows, Table 3 shows miRNA-positive regulators of NRF2 signaling.
6. Conclusions and Perspectives
It is becoming increasingly clear that the NRF2 signaling pathway can be epigenetically modulated in different ways and that the identification of new sensitive and reliable biomarkers (miRNAs and lncRNAs) that guide this mechanism in solid tumors could be useful to better understand the link between oxidative stress and cancer. This review aimed to provide a concise summary of the most recent updates in this field, but further massive investigations are required to allow the translational utility of ncRNAs as potential biomarkers related to NRF2 activity.
The oxidative stress processes related to the NRF2 activity have become a main issue in cancer biology investigation over recent years, so the identification of ncRNA alterations related to these dynamic processes will surely provide new opportunities to understand cancer biology and treatment. Moreover, further characterization of the fundamental mechanisms by which altered processing of these specific ncRNAs contributes to tumor onset and progression will be crucial to ensure great effects of many promising cancer therapies that specifically target the altered processing of RNA, with minimal effects on normal cells. From a practical and more futuristic point of view, the advent of next-generation sequencing (NGS) and liquid biopsy techniques able to detect point mutations and ncRNA level variations in blood will also allow a better clarification of the existing relationship between ncRNAs and the NRF2 pathway in a dynamic and longitudinal monitoring context of cancer patients during the course of the disease.
Author Contributions
Conceptualization, F.P.F. and L.A.M.; data curation, F.P.F. and A.S.; writing—original draft preparation, F.P.F.; writing—review and editing, L.A.M. and A.S.; visualization, A.S.; supervision, L.A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Italian Ministry of Health, Ricerca Corrente 2020, by the “5 × 1000” voluntary contributions to Fondazione IRCCS Casa Sollievo della Sofferenza.
Acknowledgments
The authors thank Domenico Trombetta for the critical revision of the manuscript and Stefano Castellana for the bioinformatic support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Jaramillo, M.C.; Zhang, D.D. The emerging role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev. 2013, 27, 2179–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levings, D.C.; Wang, X.; Kohlhase, D.; Bell, D.A.; Slattery, M. A distinct class of antioxidant response elements is consistently activated in tumors with NRF2 mutations. Redox Biol. 2018, 19, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.W.; Jaiswal, A.K. An autoregulatory loop between Nrf2 and Cul3-Rbx1 controls their cellular abundance. J. Biol. Chem. 2010, 285, 21349–21358. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yu, S.; Liu, T.; Kim, J.H.; Blank, V.; Li, H.; Kong, A.N. Heterodimerization with small Maf proteins enhances nuclear retention of Nrf2 via masking the NESzip motif. Biochim. Biophys. Acta 2008, 1783, 1847–1856. [Google Scholar] [CrossRef] [Green Version]
- Otsuki, A.; Yamamoto, M. Cis-element architecture of Nrf2-sMaf heterodimer binding sites and its relation to diseases. Arch. Pharm. Res. 2020, 43, 275–285. [Google Scholar] [CrossRef]
- Yamamoto, T.; Suzuki, T.; Kobayashi, A.; Wakabayashi, J.; Maher, J.; Motohashi, H.; Yamamoto, M. Physiological significance of reactive cysteine residues of Keap1 in determining Nrf2 activity. Mol. Cell. Biol. 2008, 28, 2758–2770. [Google Scholar] [CrossRef] [Green Version]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Villeneuve, N.F.; Lau, A.; Zhang, D.D. Regulation of the Nrf2-Keap1 antioxidant response by the ubiquitin proteasome system: An insight into cullin-ring ubiquitin ligases. Antioxid. Redox Signal. 2010, 13, 1699–1712. [Google Scholar] [CrossRef] [Green Version]
- McMahon, M.; Itoh, K.; Yamamoto, M.; Hayes, J.D. Keap1-dependent proteasomal degradation of transcription factor Nrf2 contributes to the negative regulation of antioxidant response element-driven gene expression. J. Biol. Chem. 2003, 278, 21592–21600. [Google Scholar] [CrossRef] [Green Version]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Rojo Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell. 2018, 34, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Menegon, S.; Columbano, A.; Giordano, S. The dual roles of NRF2 in cancer. Trends Mol. Med. 2016, 22, 578–593. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, F.P.; Sparaneo, A.; Trombetta, D.; Muscarella, L.A. Epigenetic versus genetic deregulation of the KEAP1/NRF2 axis in solid tumors: Focus on methylation and noncoding RNAs. Oxid. Med. Cell. Longev. 2018, 2018, 2492063. [Google Scholar] [CrossRef] [PubMed]
- Cloer, E.W.; Goldfarb, D.; Schrank, T.P.; Weissman, B.E.; Major, M.B. NRF2 activation in cancer: From DNA to protein. Cancer Res. 2019, 79, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long non-coding RNAs in the regulation of gene expression: Physiology and disease. Noncoding RNA 2019, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Lu, H.; Bai, Y. Nrf2 in cancers: A double-edged sword. Cancer Med. 2019, 8, 2252–2267. [Google Scholar] [CrossRef]
- Moi, P.; Chan, K.; Asunis, I.; Cao, A.; Kan, Y.W. Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic leucine zipper transcriptional activator that binds to the tandem NF-E2/AP1 repeat of the beta-globin locus control region. Proc. Natl. Acad. Sci. USA 1994, 91, 9926–9930. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, A.; Kang, M.I.; Okawa, H.; Ohtsuji, M.; Zenke, Y.; Chiba, T.; Igarashi, K.; Yamamoto, M. Oxidative stress sensor Keap1 functions as an adaptor for Cul3-based E3 ligase to regulate proteasomal degradation of Nrf2. Mol. Cell. Biol. 2004, 24, 7130–7139. [Google Scholar] [CrossRef] [Green Version]
- Baird, L.; Yamamoto, M. The molecular mechanisms regulating the KEAP1-NRF2 pathway. Mol. Cell. Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D. Regulation of the CNC-bZIP transcription factor Nrf2 by Keap1 and the axis between GSK-3 and β-TrCP. Curr. Opin. Toxicol. 2016, 1, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.D.; Lo, S.C.; Cross, J.V.; Templeton, D.J.; Hannink, M. Keap1 is a redox-regulated substrate adaptor protein for a Cul3-dependent ubiquitin ligase complex. Mol. Cell. Biol. 2004, 24, 10941–10953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tebay, L.E.; Robertson, H.; Durant, S.T.; Vitale, S.R.; Penning, T.M.; Dinkova-Kostova, A.T.; Hayes, J.D. Mechanisms of activation of the transcription factor Nrf2 by redox stressors, nutrient cues, and energy status and the pathways through which it attenuates degenerative disease. Free Radic. Biol. Med. 2015, 88, 108–146. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Chowdhry, S.; Dinkova-Kostova, A.T.; Sutherland, C. Dual regulation of transcription factor Nrf2 by Keap1 and by the combined actions of beta-TrCP and GSK-3. Biochem. Soc. Trans. 2015, 43, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Sun, Z.; Wang, X.J.; Jiang, T.; Huang, Z.; Fang, D.; Zhang, D.D. Direct interaction between Nrf2 and p21(Cip1/WAF1) upregulates the Nrf2-mediated antioxidant response. Mol. Cell 2009, 34, 663–673. [Google Scholar] [CrossRef] [Green Version]
- Gorrini, C.; Baniasadi, P.S.; Harris, I.S.; Silvester, J.; Inoue, S.; Snow, B.; Joshi, P.A.; Wakeham, A.; Molyneux, S.D.; Martin, B.; et al. BRCA1 interacts with Nrf2 to regulate antioxidant signaling and cell survival. J. Exp. Med. 2013, 210, 1529–1544. [Google Scholar] [CrossRef] [Green Version]
- Ichimura, Y.; Komatsu, M. Activation of p62/SQSTM1-Keap1-nuclear factor erythroid 2-related factor 2 pathway in cancer. Front. Oncol. 2018, 8, 210. [Google Scholar] [CrossRef]
- Jain, A.; Lamark, T.; Sjøttem, E.; Larsen, K.B.; Awuh, J.A.; Øvervatn, A.; McMahon, M.; Hayes, J.D.; Johansen, T. p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J. Biol. Chem. 2010, 285, 22576–22591. [Google Scholar] [CrossRef] [Green Version]
- Perdigoto, C.N. Epigenetic cancer evolution, one cell at a time. Nat. Rev. Genet. 2019, 20, 434–435. [Google Scholar] [CrossRef]
- Arbour, K.C.; Jordan, E.; Kim, H.R.; Dienstag, J.; Yu, H.A.; Sanchez-Vega, F.; Lito, P.; Berger, M.; Solit, D.B.; Hellmann, M.; et al. Effects of co-occurring genomic alterations on outcomes in patients with KRAS-mutant non-small cell lung cancer. Clin. Cancer Res. 2018, 24, 334–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.H.; Kim, J.H.; Ko, E.; Kim, J.Y.; Park, M.J.; Kim, M.J.; Seo, H.; Li, S.; Lee, J.Y. Resistance to gefitinib and cross-resistance to irreversible EGFR-TKIs mediated by disruption of the Keap1-Nrf2 pathway in human lung cancer cells. FASEB J. 2018, 32, 5862–5873. [Google Scholar] [CrossRef] [PubMed]
- Frank, R.; Scheffler, M.; Merkelbach-Bruse, S.; Ihle, M.A.; Kron, A.; Rauer, M.; Ueckeroth, F.; König, K.; Michels, S.; Fischer, R.; et al. Clinical and pathological characteristics of KEAP1- and NFE2L2-mutated Non-Small Cell Lung Carcinoma (NSCLC). Clin. Cancer Res. 2018, 24, 3087–3096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellyer, J.A.; Stehr, H.; Das, M.; Padda, S.K.; Ramchandran, K.; Neal, J.W.; Diehn, M.; Wakelee, H.A. Impact of KEAP1/NFE2L2/CUL3 mutations on duration of response to EGFR tyrosine kinase inhibitors in EGFR mutated non-small cell lung cancer. Lung Cancer 2019, 134, 42–45. [Google Scholar] [CrossRef]
- Riely GJ, J.E.; Kim, H.R.; Yu, H.A.; Berger, M.F.; Solit, D.B. Association of outcomes and co-occurring genomic alterations in patients with KRAS-mutant non-small cell lung cancer. J. Clin. Oncol. 2016, 34, 90192016. [Google Scholar] [CrossRef]
- Papagiannakopoulos, T.; Warren, L. The pleiotropic role of the KEAP1/NRF2 pathway in cancer. Annu. Rev. Cancer Biol. 2020, 4, 413–435. [Google Scholar]
- Singh, A.; Misra, V.; Thimmulappa, R.K.; Lee, H.; Ames, S.; Hoque, M.O.; Herman, J.G.; Baylin, S.B.; Sidransky, D.; Gabrielson, E.; et al. Dysfunctional KEAP1-NRF2 interaction in non-small-cell lung cancer. PLoS Med. 2006, 3, e420. [Google Scholar] [CrossRef] [Green Version]
- Ciriello, G.; Miller, M.L.; Aksoy, B.A.; Senbabaoglu, Y.; Schultz, N.; Sander, C. Emerging landscape of oncogenic signatures across human cancers. Nat. Genet. 2013, 45, 1127–1133. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The cancer genome atlas pan-cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Shibata, T.; Ohta, T.; Tong, K.I.; Kokubu, A.; Odogawa, R.; Tsuta, K.; Asamura, H.; Yamamoto, M.; Hirohashi, S. Cancer related mutations in NRF2 impair its recognition by Keap1-Cul3 E3 ligase and promote malignancy. Proc. Natl. Acad. Sci. USA 2008, 105, 13568–13573. [Google Scholar] [CrossRef] [Green Version]
- Shibata, T.; Kokubu, A.; Saito, S.; Narisawa-Saito, M.; Sasaki, H.; Aoyagi, K.; Yoshimatsu, Y.; Tachimori, Y.; Kushima, R.; Kiyono, T.; et al. NRF2 mutation confers malignant potential and resistance to chemoradiation therapy in advanced esophageal squamous cancer. Neoplasia 2011, 13, 864–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guichard, C.; Amaddeo, G.; Imbeaud, S.; Ladeiro, Y.; Pelletier, L.; Maad, I.B.; Calderaro, J.; Bioulac-Sage, P.; Letexier, M.; Degos, F. Integrated analysis of somatic mutations and focal copy-number changes identifies key genes and pathways in hepatocellular carcinoma. Nat. Genet. 2012, 44, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Ooi, A.; Dykema, K.; Ansari, A.; Petillo, D.; Snider, J.; Kahnoski, R.; Anema, J.; Craig, D.; Carpten, J.; Teh, B.T.; et al. CUL3 and NRF2 mutations confer an NRF2 activation phenotype in a sporadic form of papillary renal cell carcinoma. Cancer Res. 2013, 73, 2044–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.R.; Oh, J.E.; Kim, M.S.; Kang, M.R.; Park, S.W.; Han, J.Y.; Eom, H.S.; Yoo, N.J.; Lee, S.H. Oncogenic NRF2 mutations in squamous cell carcinomas of oesophagus and skin. J. Pathol. 2010, 220, 446–451. [Google Scholar] [CrossRef]
- Goldstein, L.D.; Lee, J.; Gnad, F.; Klijn, C.; Schaub, A.; Reeder, J.; Daemen, A.; Bakalarski, C.E.; Holcomb, T.; Shames, D.S.; et al. Recurrent loss of NFE2L2 exon 2 is a mechanism for Nrf2 pathway activation in human cancers. Cell Rep. 2016, 16, 2605–2617. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, F.P.; Mazza, T.; Castellana, S.; Sparaneo, A.; Muscarella, L.A. Epigenetic scanning of KEAP1 CpG sites uncovers new molecular-driven patterns in lung adeno and squamous cell carcinomas. Antioxidants 2020, 9, 904. [Google Scholar] [CrossRef]
- Fabrizio, F.P.; Sparaneo, A.; Centra, F.; Trombetta, D.; Storlazzi, C.T.; Graziano, P.; Maiello, E.; Fazio, V.M.; Muscarella, L.A. Methylation density pattern of KEAP1 gene in lung cancer cell lines detected by quantitative methylation specific PCR and pyrosequencing. Int. J. Mol. Sci. 2019, 20, 2697. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Yu, S.; Zhang, C.; Kong, A.N. Epigenetic regulation of Keap1-Nrf2 signaling. Free Radic. Biol. Med. 2015, 88, 337–349. [Google Scholar] [CrossRef]
- Rinn, J.L.; Chang, H.Y. Genome regulation by long noncoding RNAs. Annu. Rev. Biochem. 2012, 81, 145–166. [Google Scholar] [CrossRef] [Green Version]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2012, 505, 344–352. [Google Scholar] [CrossRef] [Green Version]
- Bhan, A.; Soleimani, M.; Mandal, S.S. Long noncoding RNA and cancer: A new paradigm. Cancer Res. 2017, 77, 3965–3981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, V.A.; Perera, R.J.; Khalil, A.M. Emerging functional and mechanistic paradigms of mammalian long non-coding RNAs. Nucleic Acids Res. 2012, 40, 6391–6400. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, C.; Zhu, J.; Shen, G.; Li, Z.; Dong, J. Long noncoding RNAs in the regulation of oxidative stress. Oxid. Med. Cell. Longev. 2019, 2019, 1318795. [Google Scholar] [CrossRef] [PubMed]
- Thai, P.; Statt, S.; Chen, C.H.; Liang, E.; Campbell, C.; Wu, R. Characterization of a novel long noncoding RNA, SCAL1, induced by cigarette smoke and elevated in lung cancer cell lines. Am. J. Respir. Cell. Mol. Biol. 2013, 49, 204–211. [Google Scholar] [CrossRef] [Green Version]
- White, N.M.; Cabanski, C.R.; Silva-Fisher, J.M.; Dang, H.X.; Govindan, R.; Maher, C.A. Transcriptome sequencing reveals altered long intergenic non-coding RNAs in lung cancer. Genome Biol. 2014, 15, 429. [Google Scholar] [CrossRef]
- Moreno Leon, L.; Gautier, M.; Allan, R.; Ilié, M.; Nottet, N.; Pons, N.; Paquet, A.; Lebrigand, K.; Truchi, M.; Fassy, J.; et al. The nuclear hypoxia-regulated NLUCAT1 long non-coding RNA contributes to an aggressive phenotype in lung adenocarcinoma through regulation of oxidative stress. Oncogene 2019, 38, 7146–7165. [Google Scholar] [CrossRef]
- Johnson, G.S.; Li, J.; Beaver, L.M.; Dashwood, W.M.; Sun, D.; Rajendran, P.; Williams, D.E.; Ho, E.; Dashwood, R.H. A functional pseudogene, NMRAL2P, is regulated by Nrf2 and serves as a coactivator of NQO1 in sulforaphane-treated colon cancer cells. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Zhou, H.; Sun, L.; Wan, F. Molecular mechanisms of TUG1 in the proliferation, apoptosis, migration and invasion of cancer cells. Oncol. Lett. 2019, 18, 4393–4402. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Huang, G.; Cheng, H. Transcription factor Nrf2 induces the up-regulation of lncRNA TUG1 to promote progression and Adriamycin resistance in urothelial carcinoma of the bladder. Cancer Manag. Res. 2019, 11, 6079–6090. [Google Scholar] [CrossRef] [Green Version]
- Ashouri, A.; Sayin, V.I.; Van den Eynden, J.; Singh, S.X.; Papagiannakopoulos, T.; Larsson, E. Pan-cancer transcriptomic analysis associates long non-coding RNAs with key mutational driver events. Nat. Commun. 2016, 7, 13197. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, Z.; Li, X.; Zhang, P.; Wang, J.; Zhu, D.; Chen, X.; Ye, L. Low long non-coding RNA HOTAIR expression is associated with down-regulation of Nrf2 in the spermatozoa of patients with asthenozoospermia or oligoasthenozoospermia. Int. J. Clin. Exp. Pathol. 2015, 8, 14198–14205. [Google Scholar] [PubMed]
- Zhang, X.; Wang, T.; Yang, Y.; Li, R.; Chen, Y.; Li, R.; Jiang, X.; Wang, L. Tanshinone IIA attenuates acetaminophen-induced hepatotoxicity through HOTAIR-Nrf2-MRP2/4 signaling pathway. Biomed. Pharmacother. 2020, 130, 110547. [Google Scholar] [CrossRef] [PubMed]
- Zeng, R.; Zhang, R.; Song, X.; Ni, L.; Lai, Z.; Liu, C.; Ye, W. The long non-coding RNA MALAT1 activates Nrf2 signaling to protect human umbilical vein endothelial cells from hydrogen peroxide. Biochem. Biophys. Res. Commun. 2018, 495, 2532–2538. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ke, S.; Zhong, L.; Wu, J.; Tseng, A.; Morpurgo, B.; Golovko, A.; Wang, G.; Cai, J.J.; Ma, X.; et al. Long noncoding RNA MALAT1 regulates generation of reactive oxygen species and the insulin responses in male mice. Biochem. Pharmacol. 2018, 152, 94–103. [Google Scholar] [CrossRef]
- Lu, Y.; Liu, W.G.; Lu, J.H.; Liu, Z.J.; Li, H.B.; Liu, G.J.; She, H.Y.; Li, G.Y.; Shi, X.H. LncRNA UCA1 promotes renal cell carcinoma proliferation through epigenetically repressing p21 expression and negatively regulating miR-495. Tumour. Biol. 2017, 39, 1010428317701632. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Fan, K.; Qu, Y.; Zhai, W.; Huang, A.; Sun, X.; Xing, S. Deregulation of UCA1 expression may be involved in the development of chemoresistance to cisplatin in the treatment of non-small-cell lung cancer via regulating the signaling pathway of microRNA-495/NRF2. J. Cell Physiol. 2020, 235, 3721–3730. [Google Scholar] [CrossRef]
- Geng, J.F.; Liu, X.; Zhao, H.B.; Fan, W.F.; Geng, J.J.; Liu, X.Z. LncRNA UCA1 inhibits epilepsy and seizure-induced brain injury by regulating miR-495/Nrf2-ARE signal pathway. Int. J. Biochem. Cell Biol. 2018, 99, 133–139. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, D.Y.; Sha, W.G.; Shen, L.; Lu, G.Y.; Yin, X. Long non-coding MIAT mediates high glucose-induced renal tubular epithelial injury. Biochem. Biophys. Res. Commun. 2015, 468, 726–732. [Google Scholar] [CrossRef]
- Liu, M.; Song, Y.; Han, Z. Study on the effect of LncRNA AK094457 on OX-LDL induced vascular smooth muscle cells. Am. J. Transl. Res. 2019, 11, 5623–5633. [Google Scholar]
- Yang, Q.; Li, K.; Huang, X.; Zhao, C.; Mei, Y.; Li, X.; Jiao, L.; Yang, H. lncRNA SLC7A11-AS1 promotes chemoresistance by blocking SCF (beta-TRCP)-mediated degradation of NRF2 in pancreatic cancer. Mol. Ther. Nucleic Acids 2020, 19, 974–985. [Google Scholar] [CrossRef]
- Wu, L.; Pan, C.; Wei, X.; Shi, Y.; Zheng, J.; Lin, X.; Shi, L. lncRNA KRAL reverses 5-fluorouracil resistance in hepatocellular carcinoma cells by acting as a ceRNA against miR-141. Cell Commun. Signal. 2018, 16, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.L.; Cai, W.P.; Lei, X.; Shi, K.Q.; Lin, X.Y.; Shi, L. NRAL mediates cisplatin resistance in hepatocellular carcinoma via miR-340-5p/Nrf2 axis. J. Cell. Commun. Signal. 2019, 13, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xiong, R.; Li, C.; Xu, M.; Guo, M. LncRNA TUG1 promotes cisplatin resistance in esophageal squamous cell carcinoma cells by regulating Nrf2. Acta Biochim. Biophys. Sin. 2019, 51, 826–833. [Google Scholar] [CrossRef]
- Gao, M.; Li, C.; Xu, M.; Liu, Y.; Cong, M.; Liu, S. LncRNA MT1DP aggravates cadmium-induced oxidative stress by repressing the function of Nrf2 and is dependent on interaction with miR-365. Adv. Sci. (Weinh) 2018, 5, 1800087. [Google Scholar] [CrossRef] [PubMed]
- Gai, C.; Liu, C.; Wu, X.; Yu, M.; Zheng, J.; Zhang, W.; Lv, S.; Li, W. MT1DP loaded by folate-modified liposomes sensitizes erastin-induced ferroptosis via regulating miR-365a-3p/NRF2 axis in non-small cell lung cancer cells. Cell Death Dis. 2020, 11, 751. [Google Scholar] [CrossRef]
- Yang, G.; Yin, H.; Lin, F.; Gao, S.; Zhan, K.; Tong, H.; Tang, X.; Pan, Q.; Gou, X. Long noncoding RNA TUG1 regulates prostate cancer cell proliferation, invasion and migration via the Nrf2 signaling axis. Pathol. Res. Pract. 2020, 216, 152851. [Google Scholar] [CrossRef]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Li, J.; Zhou, K.; Liang, T. Competing endogenous RNA: A novel posttranscriptional regulatory dimension associated with the progression of cancer. Oncol. Lett. 2015, 10, 2683–2690. [Google Scholar] [CrossRef] [Green Version]
- Kornienko, A.E.; Guenzl, P.M.; Barlow, D.P.; Pauler, F.M. Gene regulation by the act of long non-coding RNA transcription. BMC Biol. 2013, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Xu, L.; Tang, N.; Xu, Y.; Ye, X.; Shen, S.; Niu, X.; Lu, S.; Chen, Z. The polycomb group protein EZH2 inhibits lung cancer cell growth by repressing the transcription factor Nrf2. FEBS Lett. 2014, 588, 3000–3007. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Zhao, J.; Ding, J.; Shen, X.; Zhou, J.; Xu, Z. LncRNA Blnc1 expression and its effect on renal fibrosis in diabetic nephropathy. Am. J. Transl. Res. 2019, 11, 5664–5672. [Google Scholar] [PubMed]
- Bartel, D.P. Metazoan microRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, N.M.; Rushworth, S.A.; Murray, M.Y.; Bowles, K.M.; MacEwan, D.J. Understanding the role of NRF2-regulated miRNAs in human malignancies. Oncotarget 2013, 4, 1130–1142. [Google Scholar] [CrossRef] [Green Version]
- Ashrafizadeh, M.; Ahmadi, Z.; Samarghandian, S.; Mohammadinejad, R.; Yaribeygi, H.; Sathyapalan, T.; Sahebkar, A. MicroRNA-mediated regulation of Nrf2 signaling pathway: Implications in disease therapy and protection against oxidative stress. Life Sci. 2020, 244, 117329. [Google Scholar] [CrossRef]
- Reichard, J.F.; Motz, G.T.; Puga, A. Heme oxygenase-1 induction by NRF2 requires inactivation of the transcriptional repressor BACH1. Nucleic Acids Res. 2007, 35, 7074–7086. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.; Tian, Q.; Steuerwald, N.M.; Schrum, L.W.; Bonkovsky, H.L. The let-7 microRNA enhances heme oxygenase-1 by suppressing Bach1 and attenuates oxidant injury in human hepatocytes. Biochim. Biophys. Acta 2012, 1819, 1113–1122. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhao, H.; Luo, C.; Du, D.; Huang, J.; Ming, Q.; Jin, F.; Wang, D.; Huang, W. Acetaminophen responsive miR-19b modulates SIRT1/Nrf2 signaling pathway in drug-induced hepatotoxicity. Toxicol. Sci. 2019, 170, 476–488. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Sternberg, P.; Cai, J. Essential roles of the PI3 kinase/Akt pathway in regulating Nrf2-dependent antioxidant functions in the RPE. Invest. Ophthalmol. Vis. Sci. 2008, 49, 1671–1678. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.F.; Wei, Y.Y.; Yang, C.C.; Liu, C.J.; Yeh, L.Y.; Chou, C.H.; Chang, K.W.; Lin, S.C. miR-125b suppresses oral oncogenicity by targeting the anti-oxidative gene PRXL2A. Redox Biol. 2019, 22, 101140. [Google Scholar] [CrossRef]
- Zhou, C.; Zhao, L.; Zheng, J.; Wang, K.; Deng, H.; Liu, P.; Chen, L.; Mu, H. MicroRNA-144 modulates oxidative stress tolerance in SH-SY5Y cells by regulating nuclear factor erythroid 2-related factor 2-glutathione axis. Neurosci. Lett. 2016, 655, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Teng, Y.; Liu, Q. MicroRNA-153 regulates NRF2 expression and is associated with breast carcinogenesis. Clin. Lab. 2016, 62, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Lai, Y.; Chen, H.; Liu, Y.; Zhang, Z. miR-155 mediates arsenic trioxide resistance by activating Nrf2 and suppressing apoptosis in lung cancer cells. Sci. Rep. 2017, 7, 12155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Mao, L.; Wang, S.G.; Chen, F.L.; Ji, F.; Fei, H.D. MicroRNA-200a activates Nrf2 signaling to protect osteoblasts from dexamethasone. Oncotarget 2017, 8, 104867–104876. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.D.; Kim, Y.W.; Cho, I.J.; Lee, C.H.; Kim, S.G. E-cadherin inhibits nuclear accumulation of Nrf2: Implications for chemoresistance of cancer cells. J. Cell. Sci. 2012, 125, 1284–1295. [Google Scholar] [CrossRef] [Green Version]
- Chorley, B.N.; Campbell, M.R.; Wang, X.; Karaca, M.; Sambandan, D.; Bangura, F.; Xue, P.; Pi, J.; Kleeberger, S.R.; Bell, D.A. Identification of novel NRF2-regulated genes by ChIP-Seq: Influence on retinoid X receptor alpha. Nucleic Acids Res. 2012, 40, 7416–7429. [Google Scholar] [CrossRef] [Green Version]
- Luna, C.; Li, G.; Qiu, J.; Epstein, D.L.; Gonzalez, P. Role of miR-29b on the regulation of the extracellular matrix in human trabecular meshwork cells under chronic oxidative stress. Mol. Vis. 2009, 15, 2488–2497. [Google Scholar]
- Chan, K.; Lu, R.; Chang, J.C.; Kan, Y.W. NRF2, a member of the NFE2 family of transcription factors, is not essential for murine erythropoiesis, growth, and development. Proc. Natl. Acad. Sci. USA 1996, 93, 13943–13948. [Google Scholar] [CrossRef] [Green Version]
- Amodio, N.; Di Martino, M.T.; Foresta, U.; Leone, E.; Lionetti, M.; Leotta, M.; Gullà, A.M.; Pitari, M.R.; Conforti, F.; Rossi, M.; et al. miR-29b sensitizes multiple myeloma cells to bortezomib-induced apoptosis through the activation of a feedback loop with the transcription factor Sp1. Cell Death Dis. 2012, 3, e436. [Google Scholar] [CrossRef] [Green Version]
- Akdemir, B.; Nakajima, Y.; Inazawa, J.; Inoue, J. miR-432 Induces NRF2 stabilization by directly targeting KEAP1. Mol. Cancer Res. 2017, 15, 1570–1578. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wu, W.; Jiao, G.; Li, C.; Liu, H. MiR-455-3p activates Nrf2/ARE signaling via HDAC2 and protects osteoblasts from oxidative stress. Int. J. Biol. Macromol. 2018, 107, 2094–2101. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhu, H.; Wang, C.; Zhu, X.; Liu, G.; Chen, C.; Cui, Z. microRNA-455 targets cullin 3 to activate Nrf2 signaling and protect human osteoblasts from hydrogen peroxide. Oncotarget 2017, 8, 59225–59234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.J.; Rong, L.; Huang, D.; Jiang, Q. Targeting cullin 3 by miR-601 activates Nrf2 signaling to protect retinal pigment epithelium cells from hydrogen peroxide. Biochem. Biophys. Res. Commun. 2019, 515, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Akhdar, H.; Loyer, P.; Rauch, C.; Corlu, A.; Guillouzo, A.; Morel, F. Involvement of Nrf2 activation in resistance to 5-fluorouracil in human colon cancer HT-29 cells. Eur. J. Cancer 2009, 45, 2219–2227. [Google Scholar] [CrossRef]
- Inami, Y.; Waguri, S.; Sakamoto, A.; Kouno, T.; Nakada, K.; Hino, O.; Watanabe, S.; Ando, J.; Iwadate, M.; Yamamoto, M.; et al. Persistent activation of Nrf2 through p62 in hepatocellular carcinoma cells. J. Cell Biol. 2011, 193, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Moltzahn, F.; Olshen, A.B.; Baehner, L.; Peek, A.; Fong, L.; Stöppler, H.; Simko, J.; Hilton, J.F.; Carroll, P.; Blelloch, R. Microfluidic-based multiplex qRT-PCR identifies diagnostic and prognostic microRNA signatures in the sera of prostate cancer patients. Cancer Res. 2011, 71, 550–560. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Su, Z.; Sanai, N.; Xue, X.; Lu, L.; Chen, Y.; Wu, J.; Zheng, W.; Zhuge, Q.; Wu, Z.B. microRNA expression profile and differentially-expressed genes in prolactinomas following bromocriptine treatment. Oncol. Rep. 2012, 27, 1312–1320. [Google Scholar]
- Jung, K.A.; Lee, S.; Kwak, M.K. NFE2L2/NRF2 activity is linked to mitochondria and AMP-activated protein kinase signaling in cancers through miR-181c/mitochondria-encoded cytochrome c oxidase regulation. Antioxid. Redox Signal. 2017, 27, 945–961. [Google Scholar] [CrossRef]
- Yang, M.; Yao, Y.; Eades, G.; Zhang, Y.; Zhou, Q. MiR-28 regulates Nrf2 expression through a Keap1-independent mechanism. Breast Cancer Res. Treat. 2011, 129, 983–991. [Google Scholar] [CrossRef]
- Liu, H.; Wu, H.Y.; Wang, W.Y.; Zhao, Z.L.; Liu, X.Y.; Wang, L.Y. Regulation of miR-92a on vascular endothelial aging via mediating Nrf2-KEAP1-ARE signal pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2734–2742. [Google Scholar]
- Singh, B.; Ronghe, A.M.; Chatterjee, A.; Bhat, N.K.; Bhat, H.K. MicroRNA-93 regulates NRF2 expression and is associated with breast carcinogenesis. Carcinogenesis 2013, 34, 1165–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, J.; Huang, W.Z.; Wen, Y.Q.; Yi, Y.C. Effect of miR-101 on proliferation and oxidative stress-induced apoptosis of breast cancer cells via Nrf2 signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8931–8939. [Google Scholar] [PubMed]
- Duru, N.; Gernapudi, R.; Zhang, Y.; Yao, Y.; Lo, P.K.; Wolfson, B.; Zhou, Q. NRF2/miR-140 signaling confers radioprotection to human lung fibroblasts. Cancer Lett. 2015, 369, 184–191. [Google Scholar] [CrossRef] [Green Version]
- Duru, N.; Zhang, Y.; Gernapudi, R.; Wolfson, B.; Lo, P.K.; Yao, Y.; Zhou, Q. Loss of miR-140 is a key risk factor for radiation-induced lung fibrosis through reprogramming fibroblasts and macrophages. Sci. Rep. 2017, 6, 39572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Liu, D.; Xue, Y.; Hu, X. Enforced miR-144-3p expression as a non-invasive biomarker for the acute myeloid leukemia patients mainly by targeting NRF2. Clin. Lab. 2017, 63, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Ye, W.; Zhang, Y.; Yu, D.; Shao, Q.; Liang, J.; Zhang, M. miR-144 reverses chemoresistance of hepatocellular carcinoma cell lines by targeting Nrf2-dependent antioxidant pathway. Am. J. Transl. Res. 2010, 8, 2992–3002. [Google Scholar]
- Qu, J.; Zhang, L.; Li, L.; Su, Y. miR-148b functions as a tumor suppressor by targeting endoplasmic reticulum Metallo protease 1 in human endometrial cancer cells. Oncol. Res. 2018, 27, 81–88. [Google Scholar] [CrossRef]
- Narasimhan, M.; Patel, D.; Vedpathak, D.; Rathinam, M.; Henderson, G.; Mahimainathan, L. Identification of novel microRNAs in post-transcriptional control of Nrf2 expression and redox homeostasis in neuronal, SH-SY5Y cells. PLoS ONE 2012, 7, e51111. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Jiang, X.; Gu, S.; Zhang, Z. MicroRNA-155 regulates arsenite-induced malignant transformation by targeting Nrf2-mediated oxidative damage in human bronchial epithelial cells. Toxicol. Lett. 2017, 278, 38–47. [Google Scholar] [CrossRef]
- Pottier, N.; Maurin, T.; Chevalier, B.; Puisségur, M.P.; Lebrigand, K.; Robbe-Sermesant, K.; Bertero, T.; Lino Cardenas, C.L.; Courcot, E.; Rios, G.; et al. Identification of keratinocyte growth factor as a target of microRNA-155 in lung fibroblasts: Implication in epithelial-mesenchymal interactions. PLoS ONE 2009, 4, e6718. [Google Scholar] [CrossRef]
- Zhao, M.W.; Yang, P.; Zhao, L.L. Chlorpyrifos activates cell pyroptosis and increases susceptibility on oxidative stress-induced toxicity by miR-181/SIRT1/PGC-1alpha/Nrf2 signaling pathway in human neuroblastoma SH-SY5Y cells: Implication for association between chlorpyrifos and Parkinson's disease. Environ. Toxicol. 2019, 34, 699–707. [Google Scholar] [PubMed]
- Shi, L.; Chen, Z.G.; Wu, L.L.; Zheng, J.J.; Yang, J.R.; Chen, X.F.; Chen, Z.Q.; Liu, C.L.; Chi, S.Y.; Zheng, J.Y.; et al. miR-340 reverses cisplatin resistance of hepatocellular carcinoma cell lines by targeting Nrf2-dependent antioxidant pathway. Asian Pac. J. Cancer Prev. 2014, 15, 10439–10444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaisiya, M.; Coda Zabetta, C.D.; Bellarosa, C.; Tiribelli, C. Bilirubin mediated oxidative stress involves antioxidant response activation via Nrf2 pathway. Cell Signal. 2014, 26, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Inoue, J.; Kawano, T.; Kozaki, K.; Omura, K.; Inazawa, J. The impact of miRNA-based molecular diagnostics and treatment of NRF2-stabilized tumors. Mol. Cancer Res. 2014, 12, 58–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Experimentally validated long noncoding RNAs (lncRNAs) which contribute to a down and upregulation of Nuclear factor erythroid 2-related factor (NRF2) at mRNA/protein levels in cancers, based on their regulatory (left side) or transcriptional activities (right side). Looking at the regulatory lncRNA list, some of them induce an overexpression of NRF2 (Metastasis-associated lung adenocarcinoma transcript (MALAT1), Urothelial carcinoma-associated 1 (UCA1), BLNC1, Myocardial infarction-associated transcript 1 (MIAT), AK094457, KEAP1 regulation-associated lncRNA (KRAL), Taurine-upregulated gene 1 (TUG1); green arrow), whereas another group causes a reduction in NRF2 levels (Hox transcript antisense intergenic RNA (HOTAIR), Solute Carrier Family 7 Member 11-antisense 1 (SLC7A11-AS1), NRF2 regulation-associated lncRNA (NRAL), metallothionein 1D pseudogene (MT1DP); red arrow). On the other hand, NRF2 stimulates the transcription of the following lncRNAs: SCAL1, nuclear smoke- and cancer-associated lncRNA (NLUCAT1), NmrA-like redox sensor 2 pseudogene (NMRAL2P), TUG1 and long intergenic nonprotein coding RNA 942 (LINC00942). Abbreviations: sMaf, avian musculo aponeurotic fibrosarcoma oncogene homolog; CREB, cAMP response element-binding protein; NRF2, Nuclear factor erythroid 2-related factor 2; HOTAIR, Hox transcript antisense intergenic RNA; MALAT1, Metastasis-associated lung adenocarcinoma transcript 1; UCA1, Urothelial carcinoma-associated 1; MIAT, Myocardial infarction-associated transcript 1; SLC7A11-AS1, Solute Carrier Family 7 Member 11-antisense 1; KRAL, KEAP1 regulation-associated lncRNA; NRAL, NRF2 regulation-associated lncRNA; TUG1, Taurine-upregulated gene 1; MT1DP, metallothionein 1D pseudogene; SCAL1 or LUCAT1, smoke- and cancer-associated lncRNA 1; NLUCAT1, nuclear smoke- and cancer-associated lncRNA 1; NMRAL2P, NmrA-like redox sensor 2 pseudogene; LINC00942, Long Intergenic Nonprotein Coding RNA 942.
Figure 1.
Experimentally validated long noncoding RNAs (lncRNAs) which contribute to a down and upregulation of Nuclear factor erythroid 2-related factor (NRF2) at mRNA/protein levels in cancers, based on their regulatory (left side) or transcriptional activities (right side). Looking at the regulatory lncRNA list, some of them induce an overexpression of NRF2 (Metastasis-associated lung adenocarcinoma transcript (MALAT1), Urothelial carcinoma-associated 1 (UCA1), BLNC1, Myocardial infarction-associated transcript 1 (MIAT), AK094457, KEAP1 regulation-associated lncRNA (KRAL), Taurine-upregulated gene 1 (TUG1); green arrow), whereas another group causes a reduction in NRF2 levels (Hox transcript antisense intergenic RNA (HOTAIR), Solute Carrier Family 7 Member 11-antisense 1 (SLC7A11-AS1), NRF2 regulation-associated lncRNA (NRAL), metallothionein 1D pseudogene (MT1DP); red arrow). On the other hand, NRF2 stimulates the transcription of the following lncRNAs: SCAL1, nuclear smoke- and cancer-associated lncRNA (NLUCAT1), NmrA-like redox sensor 2 pseudogene (NMRAL2P), TUG1 and long intergenic nonprotein coding RNA 942 (LINC00942). Abbreviations: sMaf, avian musculo aponeurotic fibrosarcoma oncogene homolog; CREB, cAMP response element-binding protein; NRF2, Nuclear factor erythroid 2-related factor 2; HOTAIR, Hox transcript antisense intergenic RNA; MALAT1, Metastasis-associated lung adenocarcinoma transcript 1; UCA1, Urothelial carcinoma-associated 1; MIAT, Myocardial infarction-associated transcript 1; SLC7A11-AS1, Solute Carrier Family 7 Member 11-antisense 1; KRAL, KEAP1 regulation-associated lncRNA; NRAL, NRF2 regulation-associated lncRNA; TUG1, Taurine-upregulated gene 1; MT1DP, metallothionein 1D pseudogene; SCAL1 or LUCAT1, smoke- and cancer-associated lncRNA 1; NLUCAT1, nuclear smoke- and cancer-associated lncRNA 1; NMRAL2P, NmrA-like redox sensor 2 pseudogene; LINC00942, Long Intergenic Nonprotein Coding RNA 942.

Figure 2.
Modulation of the NRF2 signaling pathway by microRNAs (miRNAs) in cancers. Representative scheme depicts miRNAs that directly or indirectly target NRF2 pathway, thus impacting on the expression of antioxidant response element (ARE) genes. The red lines indicate an inhibitory effect on NRF2, otherwise the green lines a stimulatory effect on NRF2. A group of miRNAs with NRF2 indirect interaction is indicated in the bottom box, whereas those which stimulate NRF2 direct interaction in the upper box. Abbreviations: CUL3, Cullin 3; KEAP1, Kelch-Like ECH-Associated Protein 1; NRF2, Nuclear factor erythroid 2-related factor 2; sMAF, avian musculo aponeurotic fibrosarcoma oncogene homolog; SIRT1/PGC-1α, SIRT1, Sirtuin 1/PGC-1α, Peroxisome proliferator-activated receptor gamma coactivator 1-alpha; PI3K, Phosphoinositide 3-kinase; HDAC2, Histone deacetylase 2; PRXL2A, Peroxiredoxin-like 2A; BACH1, bric-a-brac (BTB) and cap’n’collar (CNC) homology, basic leucine zipper transcription factor; NQO1, NAD(P)H Quinone Dehydrogenase 1; HO-1, Heme oxygenase-1; GSH, glutathione; GSR, Glutathionereductase; NO, nitric oxide; SOD, superoxide dismutase; GCLC, Glutamate-Cysteine Ligase Catalytic Subunit.
Figure 2.
Modulation of the NRF2 signaling pathway by microRNAs (miRNAs) in cancers. Representative scheme depicts miRNAs that directly or indirectly target NRF2 pathway, thus impacting on the expression of antioxidant response element (ARE) genes. The red lines indicate an inhibitory effect on NRF2, otherwise the green lines a stimulatory effect on NRF2. A group of miRNAs with NRF2 indirect interaction is indicated in the bottom box, whereas those which stimulate NRF2 direct interaction in the upper box. Abbreviations: CUL3, Cullin 3; KEAP1, Kelch-Like ECH-Associated Protein 1; NRF2, Nuclear factor erythroid 2-related factor 2; sMAF, avian musculo aponeurotic fibrosarcoma oncogene homolog; SIRT1/PGC-1α, SIRT1, Sirtuin 1/PGC-1α, Peroxisome proliferator-activated receptor gamma coactivator 1-alpha; PI3K, Phosphoinositide 3-kinase; HDAC2, Histone deacetylase 2; PRXL2A, Peroxiredoxin-like 2A; BACH1, bric-a-brac (BTB) and cap’n’collar (CNC) homology, basic leucine zipper transcription factor; NQO1, NAD(P)H Quinone Dehydrogenase 1; HO-1, Heme oxygenase-1; GSH, glutathione; GSR, Glutathionereductase; NO, nitric oxide; SOD, superoxide dismutase; GCLC, Glutamate-Cysteine Ligase Catalytic Subunit.


{kind=link}
{kind=link}
Table 1.
(A). LncRNAs modulated by NRF2 signaling. (B). Positive and negative lncRNA regulators of the NRF2 activity.
Table 1.
(A). LncRNAs modulated by NRF2 signaling. (B). Positive and negative lncRNA regulators of the NRF2 activity.
| (A) | ||||||
| LncRNA ID | Chr Location * | Role | NRF2 Levels | NRF2-Related Effects | Cell Cultures/Validation Models | Refs. |
| SCAL1 (LUCAT1) | chr5: 91,285,160–91,314,516 | Transcriptional activity | Upregulated | Downstream mediator of NRF2 against ROS-induced oxidative stress, having a pro-tumorigenic role | HBE cells; Lung ADC and SqCC | [54,55] |
| NLUCAT1 | 5q14.3 | Transcriptional activity | Upregulated | It induces a positive loop on the NRF2 network, correlates with hypoxic stress and contributes to tumor invasion and proliferation | A549 ADC cells | [56] |
| NMRAL2P | chr3: 185,959,943–185,980,872 | Transcriptional activity | Upregulated | Downstream regulator of NRF2-dependent NQO1 activation. It exerts an oncogenic action by promoting EMT | Colon cancer cells | [57] |
| TUG1 | chr22: 30,970,182–30,978,847 | Transcriptional activity | Upregulated | Oncogenic roles in apoptosis, proliferation and chemoresistance by modulating the NRF2 signaling | UCB cells; prostate cancer cells | [58,59] |
| LINC00942 | chr12: 1,500,525–1,504,424 | Transcriptional activity | Upregulated | It concurs on NRF2 antioxidant pathway activation | 19 cancer types from TCGA | [60] |
| (B) | ||||||
| LncRNA ID | Chr Locations * | Role | NRF2-Related Effects | Cell Cultures/Validation Models | Refs. | |
| HOTAIR | chr12: 53,962,308–53,974,956 | Negative regulator | Mediates histone H4 acetylation at the NFE2L2 gene promoter and is also involved in the epigenetic suppression via NRF2-MRP2/4 pathway | GC-1 spg; human hepatocytes L02 cells | [61,62] | |
| MALAT1 | chr11: 65,497,688–65,506,431 | Positive regulator | Participates in MALAT1-mediated HUVEC protection from hydrogen peroxide and it is negatively regulated by KEAP1 | HUVEC cells; mouse primary hepatocytes | [63,64] | |
| UCA1 | chr19: 15,828,206–15,836,136 | Positive regulator | It has an oncogenic role promotes proliferation through p21 repression, it enhances chemoresistance to cisplatin via UCA1/NRF2 crosstalk signaling by sponging miR-495 and promoting the inhibition of apoptosis | Human RCC cells; A549 lung ADC cells; HEK-293 cells | [65,66,67] | |
| BLNC1 | chr9: 95,559,657–95,568,023 | Positive regulator | Participates in the hyperactivation of HO-1/NRF2 and crosstalks with NF-κB pathway by attenuating renal fibrosis, inflammation and oxidative stress | Renal HK-2 cells | [62] | |
| MIAT | chr22: 26,657,520–26,676,475 | Positive regulator | Targets NRF2 by mediating high glucose- induced tubular injury | renal HK-2 epithelial cells | [68] | |
| AK094457 | chr10: 124,745,200–124,748,551 | Positive regulator | Inversely correlates with the upregulation of NRF2 and HO-1 | VSMC cells | [69] | |
| SLC7A11-AS1 | chr4: 138,089,014–138,178,177 | Negative regulator | Promotes resistance to gemcitabine by repressing SCFβ-TRCP-mediated degradation of NRF2 | PDAC cells | [70] | |
| KRAL | 19q10.14 | Positive regulator | Increases sensitivity to fluorouracil by targeting miR-141/KEAP1 axis as ceRNA | HCC SMMC-7721 and HepG2 cells | [71] | |
| NRAL | 19q10.14 | Negative regulator | Enhances resistance to cisplatin for NRF2 by binding to miR-340–5p | HCC SMMC-7721 and HepG2 cells | [72] | |
| TUG1 | chr22: 30,970,182–30,978,847 | Positive regulator | Promotes cisplatin resistance by regulating and stabilizing the NRF2 protein | human EC TE-1 cells | [73] | |
| MT1DP | chr16: 56,643,705–56,644,786 | Negative regulator | Enhances the sensitivity of NRF2 overexpression to erastin-induced ferroptosis by stabilizing miR-365a-3p | HCC HepG2 cells; NSCLC A549 and H1299 cells | [74,75] | |
(A) * UCSC Genome Browser on Human December 2013 (GRCh38/hg38). Abbreviations: Chr, chromosomal; SCAL1 or LUCAT1, smoke- and cancer-associated lncRNA 1; NLUCAT1, nuclear smoke- and cancer-associated lncRNA 1; NMRAL2P, NmrA-like redox sensor 2 pseudogene; TUG1, Taurine-upregulated gene 1; LINC00942, Long Intergenic Nonprotein Coding RNA 942; NRF2, Nuclear factor erythroid 2-related factor 2; ROS, reactive oxygen species; HBE, human bronchial epithelial; ADC, adenocarcinoma; SqCC, squamous cell carcinoma; NQO1, NAD(P)H Quinone Dehydrogenase 1; EMT, epithelial–mesenchymal transition; UCB, urothelial carcinoma of the bladder; TCGA, The Cancer Genome Atlas. (B) * UCSC Genome Browser on Human December 2013 (GRCh38/hg38). Abbreviations: HOTAIR, Hox transcript antisense intergenic RNA; MALAT1, Metastasis-associated lung adenocarcinoma transcript 1; UCA1, Urothelial carcinoma-associated 1; MIAT, Myocardial infarction-associated transcript 1; SLC7A11-AS1, Solute Carrier Family 7 Member 11-antisense 1; NRAL, NRF2 regulation-associated lncRNA; KRAL, KEAP1 regulation-associated lncRNA; MT1DP, metallothionein 1D, pseudogene; MRP2/4, Multidrug resistance-associated protein 2/4; GC-1 spg, germ cell-1 spermatogonial; HUVEC, human umbilical vein endothelial cells; KEAP1, Kelch-Like ECH-Associated Protein 1; RCC, Renal cell carcinoma; ADC, adenocarcinoma; HEK-293, Human embryonic kidney-293; HO-1, Heme oxygenase-1; NF-κB, Nuclear factor-κB; HK-2, Human Kidney-2; VSMC, vascular smooth muscle cells; SCFβ-TRCP, Skp1-cullin-F-box protein-transducin repeat- containing proteins; PDAC, Pancreatic ductal adenocarcinoma cells; ceRNA, competing endogenous RNA; HCC, Hepatocellular carcinoma; EC, esophageal carcinoma; NSCLC, Non small cell lung cancer.
Table 2.
List of miRNA-positive regulators of NRF2 signaling.
| miRNA ID | Direct or Indirect Interaction/ Targets | NRF2-Related Effects | Cell Cultures/Validation Models | Refs. |
|---|---|---|---|---|
| Let 7b/c | Indirect; BACH1, HO-1 | Participates in negative regulation by suppressing BACH1 and thus stimulating the overexpression of HO-1 via NRF2 | HaCaT human keratinocytes cells; HCC (Huh-7 and HepG2) cells | [86,87] |
| MiR-19b | Indirect; SIRT-1 | Stimulates the activation of SIRT1 by activating NRF2 cascade and its downstream target genes; enhancing drug hepatotoxicity | Human liver LO2 cells | [88] |
| MiR-32 | Indirect; PI3K | Exerts an oncogenic role by inducing the upregulation of NRF2and promoting cells survival | Human RPE cells | [89] |
| MiR-125b | Indirect; PRXL2A | Promotes PRXL2A activation by positive feedback loops involving NRF2 signaling pathway | OSCC (SAS, OECM1, HSC3, FaDu, OC3) cells, HEK-293 and primary NOK cells | [90] |
| MiR-144 | Direct | Regulates the ROS scavenging via NRF2 and stimulates the GSH synthesis | Neuroblastoma SH-SY5Y cells | [91] |
| MiR-153 | Direct | Acts as an oncogene by promoting cell migration and invasion via NRF2 signaling | CRC cells; OSCC (CAL 27) cells and tissues | [92] |
| MiR-155 | Direct | Upregulates NRF2 and its downstream targets (HO-1 and NQO1) by inhibiting apoptosis and mediating ATO resistance | Human lung ADC A549 cells | [93] |
| MiR-181c | Direct | Enhances the activation of NRF2 and other multiple pro-survival pathways | Human CRC (HT29) cells | [108] |
| MiR-200a | Indirect; KEAP1 | Stimulates NRF2 signaling pathway by suppressing KEAP1 and decreasing ROS concentration | OB-6 human osteoblastic cells | [94] |
| MiR-200c | Indirect; PI3K/AKT | Its downregulation provides a feedback mechanism by which NRF2 indirectly regulates E-cadherin and metastasis via PI3K/AKT activation | RPE (ARPE-19) cells | [89] |
| MiR-365/193b/29b | Direct | Enforces NRF2 and its downstream targets activation | Lymphoblast cells | [96] |
| MiR-432 | Indirect; KEAP1 | Targets KEAP1 and decreases the sensitivity to cisplatin (CDDP) drug via activation and stabilization of NRF2 | ESCC (KYSE170, KYSE770, and KYSE2270) cells | [100] |
| MiR-455 | Indirect; HDAC2 | Promotes NRF2 expression via its upstream mediator, HDAC2, by ensuring more protection against oxidative injury and activating ARE-related genes (NQO1, HO-1 and GCLC) | MC3T3-E1 and hFOB1.19 osteoblastic cells | [101,102] |
| MiR-601 | Indirect; CUL3/KEAP1 | Targets CUL3 and concurs into a dependent mechanism of CUL3-KEAP1-NRF2 activation against oxidative damage | RPE (ARPE-19) cells | [103] |
| MiR-617/592/1207/550 | Direct | Exert a pro-oncogenic actions on the NRF2 upregulation; promote cell growth and proliferation by activating antioxidant defense system | ESCC cells; colorectal and liver carcinoma cells; prostate cancer cells | [41,104,105,106,107] |
Abbreviations: BACH1, BTB Domain And CNC Homolog 1; HO-1, Heme oxygenase 1; NRF2, Nuclear factor erythroid 2-related factor 2; HCC, Hepatocellular carcinoma; SIRT-1, Sirtuin 1; PI3K, Phosphoinositide 3-kinase; RPE, retinal pigment epithelium; PRXL2A, Peroxiredoxin-Like 2A; OSCC, Oral squamous cell carcinoma; HEK-293, human embryonic kidney-293; NOK, Normal oral keratinocyte; GSH, Glutathione; CRC, Colorectal cancer; NQO1, NAD(P)H Quinone Dehydrogenase 1; ATO, arsenic trioxide; ADC, Adenocarcinoma; KEAP1, Kelch-Like ECH-Associated Protein 1; AKT, Protein kinase B or PKB; ESCC, Esophageal squamous cell carcinoma; HDAC2, Histone deacetylase 2; ARE, Antioxidant responsive element; GCLC, Glutamate-Cysteine Ligase Catalytic Subunit; CUL3, Cullin 3.
Table 3.
List of miRNA-negative regulators of NRF2 signaling.
| miRNA ID | Direct or Indirect Interaction/Targets | NRF2-Related Effects | Cell Cultures/Validation Models | Refs. |
|---|---|---|---|---|
| MiR-28 | Indirect; KEAP1 | Exerts a negative effect by repressing NRF2 and inhibiting tumor cell growth independently from KEAP1, without any changes at protein level | Human breast MCF-7 and HEK-293 cells | [109] |
| MiR-92a | Indirect; KEAP1/ARE | Its inhibition leads to cell proliferation by decreasing apoptosis-inducing factors and KEAP1 expression with a consequent upregulation of NRF2 and its target genes | HUVECs | [110] |
| MiR-93 | Direct | Acts as a oncogenic regulator by downregulating NRF2 gene and by targeting specific 3′UTR sites with a strong impact on decreasing apoptosis, increasing colony formation and cell migration | Human breast epithelial MCF-10A and neoplastic T47D cells | [111] |
| MiR-101 | Direct | Affects the binding to 3′UTR of NRF2, thus promoting NRF2 overexpression and inhibiting cell proliferation | Human breast MCF-7 cells | [112] |
| MiR-140 | Direct | Targets NRF2 gene by ensuring more radioprotection against TGF-β1-mediated inflammation and fibroblasts differentiation | Human normal lung fibroblast (CCD-19Lu) and mammary epithelial MCF10A cells | [113,114] |
| MiR-144 | Direct | Exerts an oncogenic role by inhibiting NRF2 signaling pathway as well as promotes cell viability, suppresses apoptosis and finally reverses chemoresistance NRF2-related | HL-60 AML and Bel-7402 HCC cells | [115,116] |
| MiR-148b | Direct | Suppresses cell proliferation and regulates the oxidative stress response by downregulating HIF-1 and NRF2 and inhibiting ERMP1 | RL95-2 human endometrial cancer cells | [117] |
| MiR-153/27a/142/144 | Direct | Act as oncogenic miRNAs which decrease apoptosis and successively reinforce cells proliferation via repressing NRF2 and its downstream target genes (GCLC, GSR) | Breast cancer and CRC cell lines; SH-SY5Y neuroblastoma cells | [92,118] |
| MiR-155 | Direct | Accelerates cell malignant transformation by targeting NRF2-mediated oxidative damage and repressing NRF2 and its related gene expressions (GSH, NO and SOD) | 16-HBE cells | [119] |
| MiR-181 | Indirect; SIRT1/PGC-1α | Suppresses SIRT1/PGC-1α/NRF2 signaling pathway by activating cell apoptosis and oxidative stress | SH-SY5Y neuroblastoma cells | [121] |
| MiR-340 | Direct | Inhibits NRF2-dependent antioxidant pathway and enhancing cells sensitivity to cisplatin | HCC SMMC-7721 and HepG2 cells | [122] |
| MiR-365 | Indirect; MT1DP | Represses NRF2 activity via direct binding to its 3′UTR and induce an aggravation of oxidative stress by activating MT1DP | HCC HepG2 cells | [74] |
| MiR-507/634/450a/129 | Direct | Negatively affect NRF2 oncogenic regulation by directly targeting NRF2, thus increasing cisplatin sensitivity and suppressing cell growth | A549 ADC cells | [123,124] |
Abbreviations: KEAP1, Kelch-Like ECH-Associated Protein 1; NRF2, Nuclear factor erythroid 2-related factor 2; HEK-293, human embryonic kidney-293; ARE, Antioxidant responsive element; HUVECs, human umbilical vein endothelial cells; 3′-UTR, 3′-untranslated regions; TGF-β1, Transforming growth factor beta 1; HL-60, human leukemic-60; HCC, hepatocellular cancer; HIF-1, Hypoxia-inducible factor 1; ERMP1, Endoplasmic Reticulum Metallopeptidase 1; GCLC, Glutamate-Cysteine Ligase Catalytic Subunit; GSR, Glutathione reductase; GSH, Glutathione; NO, Nitric oxide; SOD, Superoxide dismutase; HBE, human bronchial epithelial; SIRT1, Sirtuin 1; PGC-1α, Peroxisome proliferator-activated receptor gamma coactivator 1-alpha; MT1DP, Metallothionein 1D, pseudogene; ADC, Adenocarcinoma.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fabrizio, F.P.; Sparaneo, A.; Muscarella, L.A. NRF2 Regulation by Noncoding RNAs in Cancers: The Present Knowledge and the Way Forward. Cancers 2020, 12, 3621. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123621
AMA Style
Fabrizio FP, Sparaneo A, Muscarella LA. NRF2 Regulation by Noncoding RNAs in Cancers: The Present Knowledge and the Way Forward. Cancers. 2020; 12(12):3621. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123621
Chicago/Turabian StyleFabrizio, Federico Pio, Angelo Sparaneo, and Lucia Anna Muscarella. 2020. "NRF2 Regulation by Noncoding RNAs in Cancers: The Present Knowledge and the Way Forward" Cancers 12, no. 12: 3621. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12123621
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.