
Cell-Extrinsic Differentiation Block Mediated by EphA3 in Pre-Leukaemic Thymus Contributes to Disease Progression

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mouse Strains
2.2. Tissue Processing
2.3. Flow Cytometry: Thymocyte DN Populations, Chimerism, and Cell Cycle
2.4. Thymocyte Transplantation
2.5. Gene Expression Study
2.6. Disease Progression Study
2.7. Statistical Analyses
3. Results
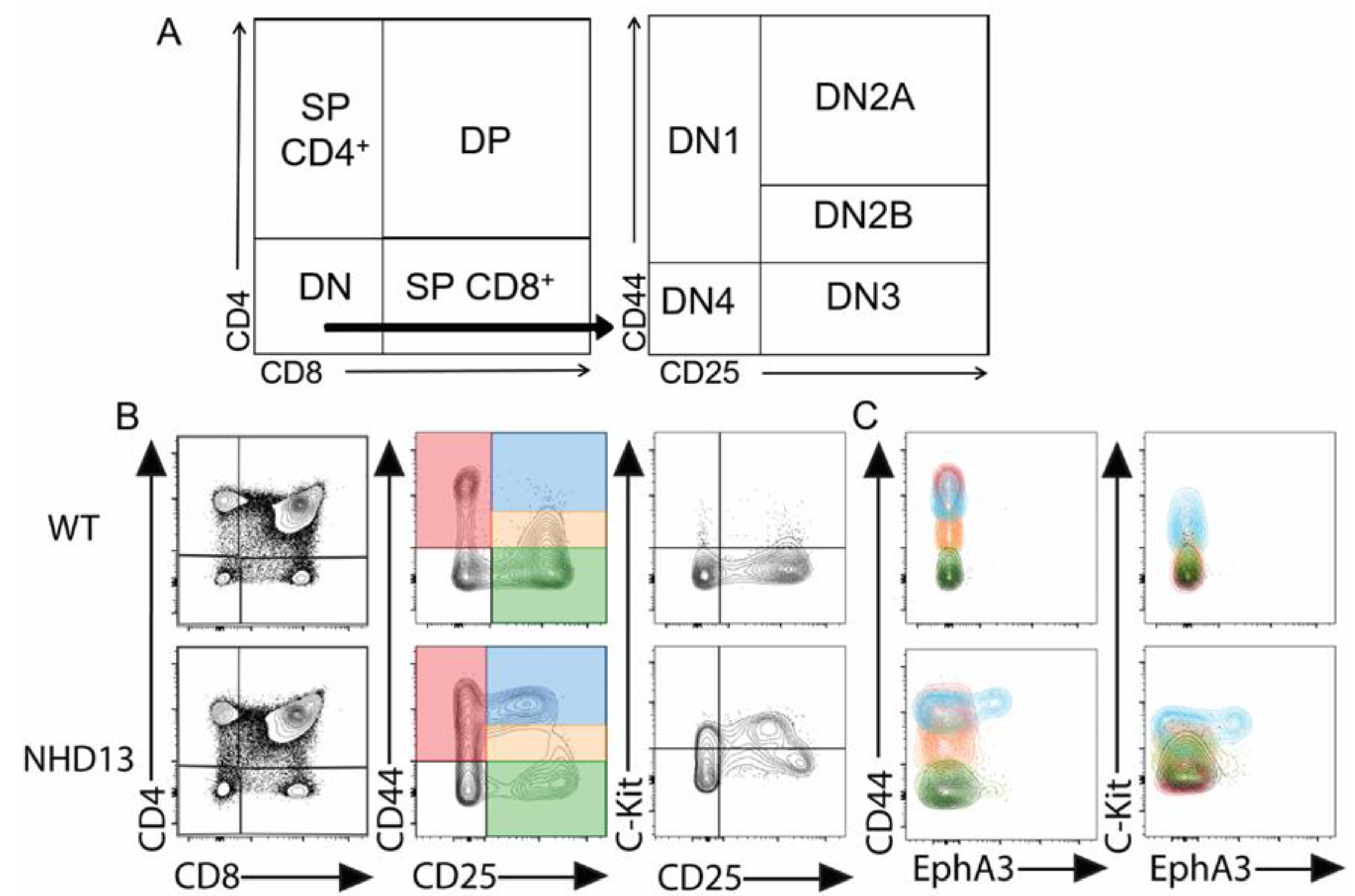
3.1. EphA3 Is Abnormally Expressed in NHD13 Thymocytes
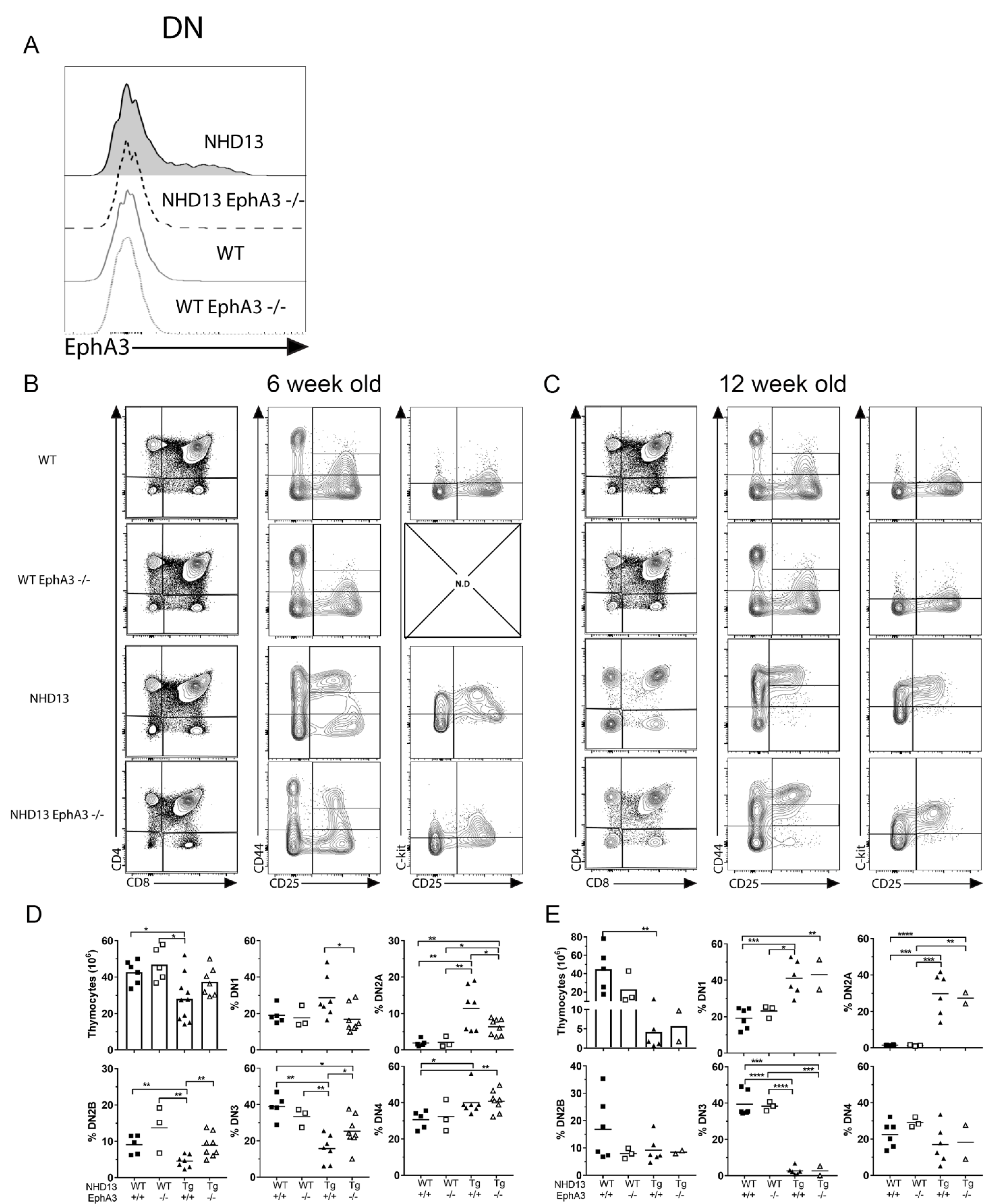
3.2. EphA3 Deletion Delays the NHD13 Thymic Phenotype
3.3. EphA3 Deletion Reduces T-ALL Incidence in NHD13 Mice
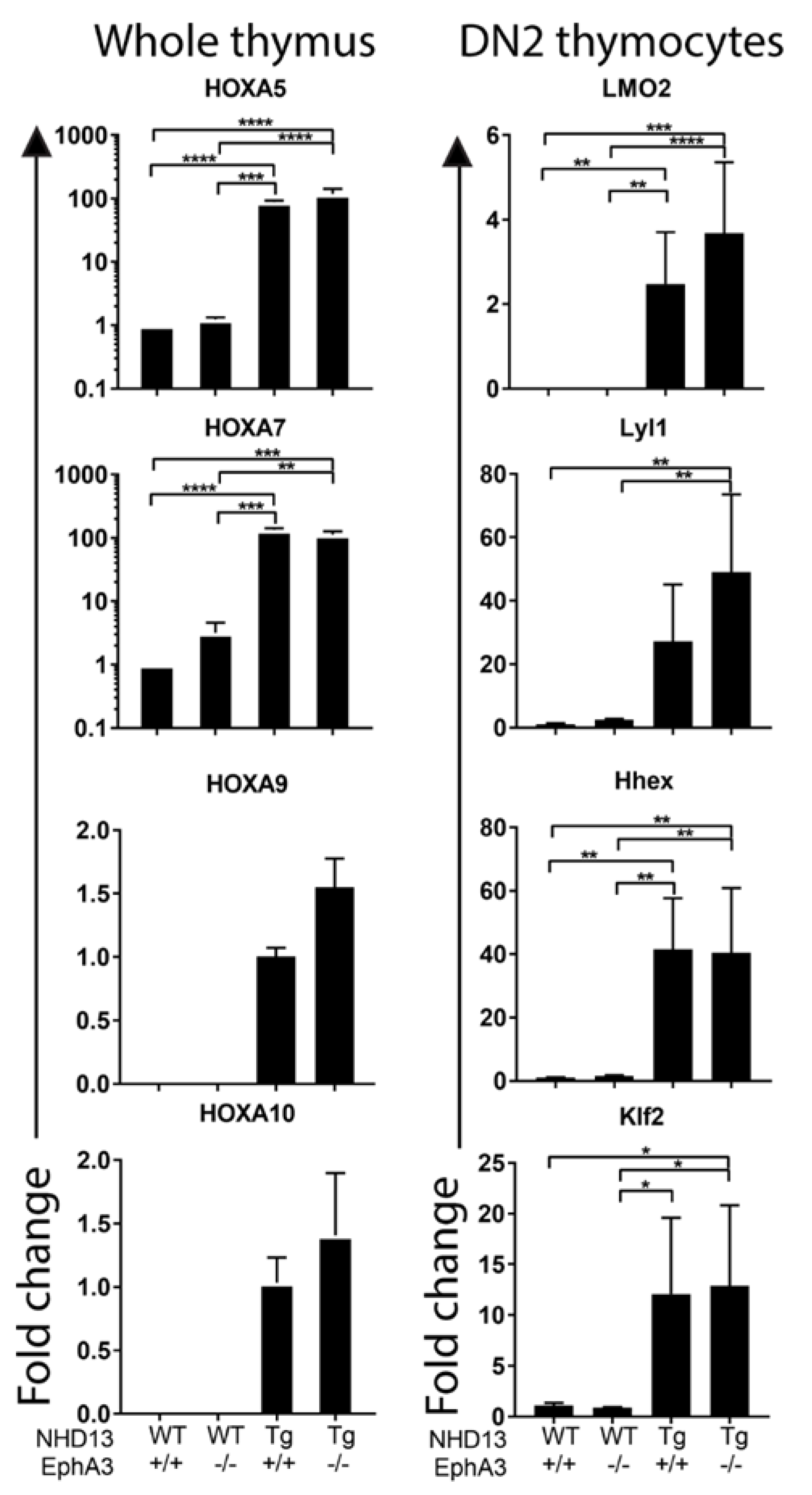
3.4. Sustained Abnormal Expression of Self-Renewal Genes in the Absence of EphA3
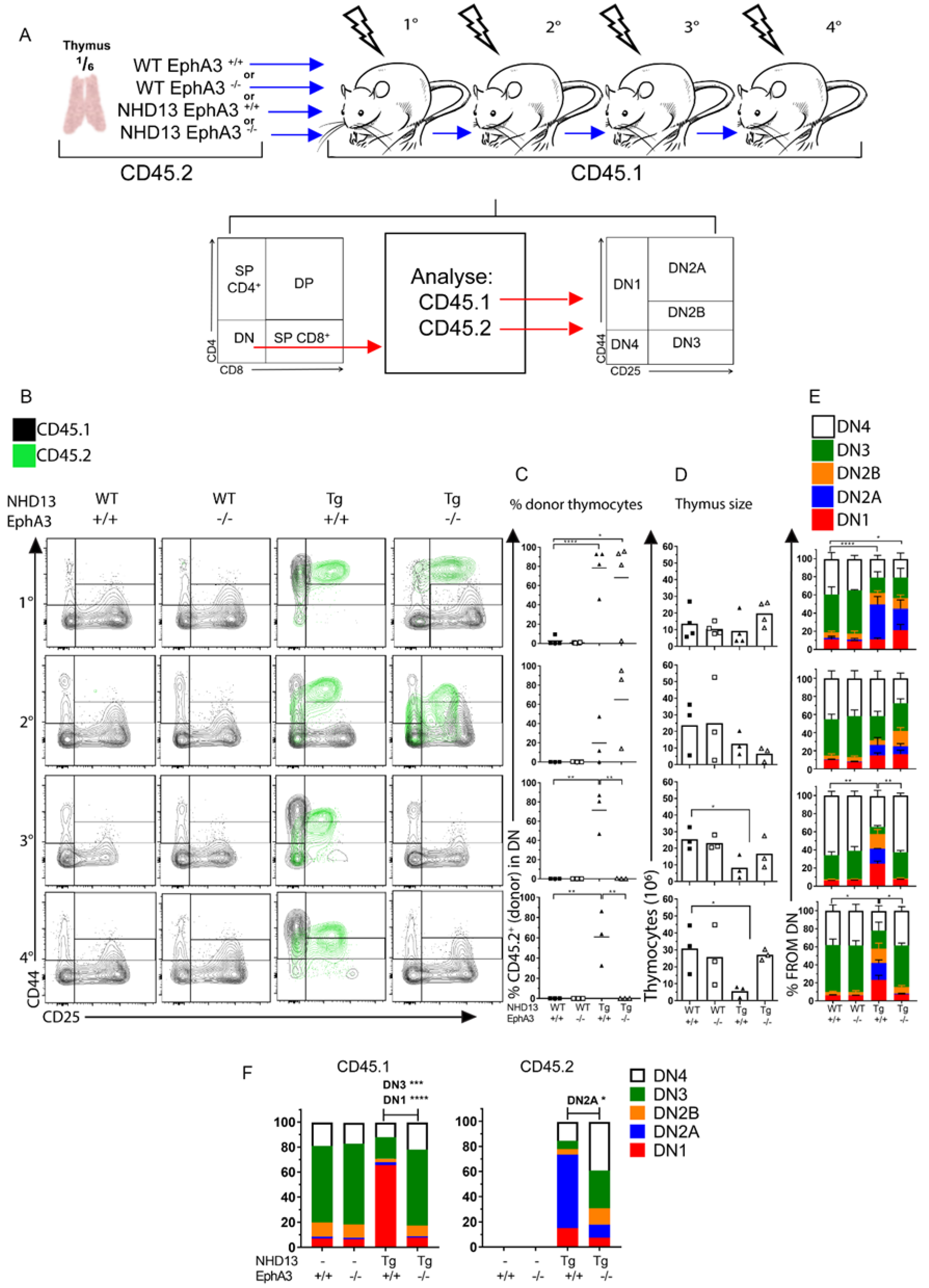
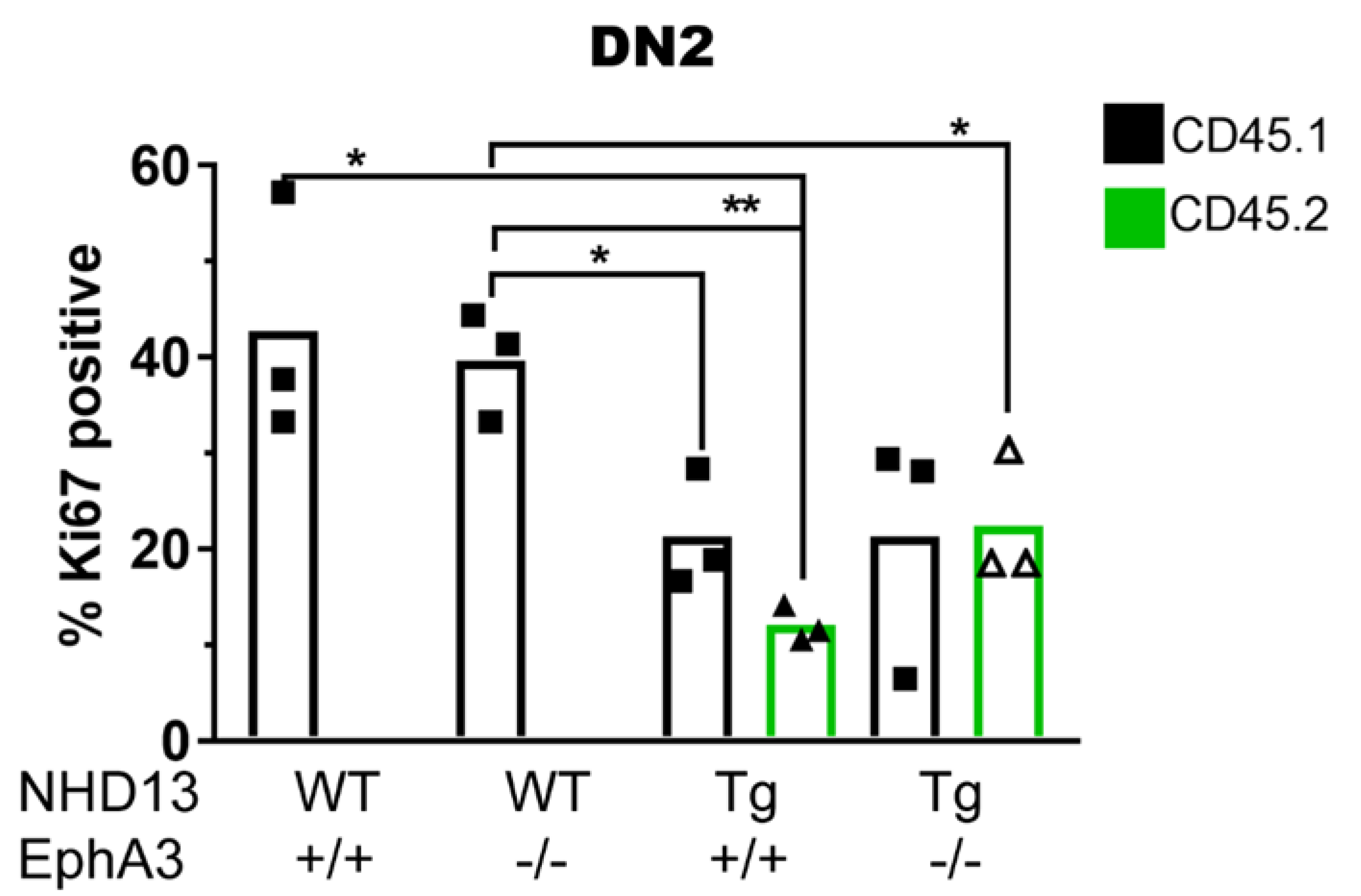
3.5. EphA3 Mediates an Interaction between Donor and Recipient Thymocytes
3.6. NHD13 DN2 Thymocytes Induce Cell Cycle Arrest on WT Cells Independently of EphA3 Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Ding, L.; Holmfeldt, L.; Wu, G.; Heatley, S.L.; Payne-Turner, D.; Easton, J.; Chen, X.; Wang, J.; Rusch, M.; et al. The genetic basis of early T-cell precursor acute lymphoblastic leukaemia. Nature 2012, 481, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Ferrando, A.A.; Neuberg, D.S.; Staunton, J.; Loh, M.L.; Huard, C.; Raimondi, S.C.; Behm, F.G.; Pui, C.H.; Downing, J.R.; Gilliland, D.G.; et al. Gene expression signatures define novel oncogenic pathways in T cell acute lymphoblastic leukemia. Cancer Cell 2002, 1, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Van Vlierberghe, P.; Pieters, R.; Beverloo, H.B.; Meijerink, J.P. Molecular-genetic insights in paediatric T-cell acute lymphoblastic leukaemia. Br. J. Haematol. 2008, 143, 153–168. [Google Scholar] [CrossRef]
- Girardi, T.; Vicente, C.; Cools, J.; De Keersmaecker, K. The genetics and molecular biology of T-ALL. Blood 2017, 129, 1113–1123. [Google Scholar] [CrossRef]
- Eguchi-Ishimae, M.; Eguchi, M.; Kempski, H.; Greaves, M. NOTCH1 mutation can be an early, prenatal genetic event in T-ALL. Blood 2008, 111, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Shields, B.J.; Slape, C.I.; Vo, N.; Jackson, J.T.; Pliego-Zamora, A.; Ranasinghe, H.; Shi, W.; Curtis, D.J.; McCormack, M.P. The NUP98-HOXD13 fusion oncogene induces thymocyte self-renewal via Lmo2/Lyl1. Leukemia 2019, 33, 1868–1880. [Google Scholar] [CrossRef]
- McCormack, M.P.; Young, L.F.; Vasudevan, S.; de Graaf, C.A.; Codrington, R.; Rabbitts, T.H.; Jane, S.M.; Curtis, D.J. The Lmo2 oncogene initiates leukemia in mice by inducing thymocyte self-renewal. Science 2010, 327, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.W.; Chung, Y.J.; Slape, C.; Aplan, P.D. A NUP98-HOXD13 fusion gene impairs differentiation of B and T lymphocytes and leads to expansion of thymocytes with partial TCRB gene rearrangement. J. Immunol. 2009, 183, 6227–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.W.; Slape, C.; Zhang, Z.; Aplan, P.D. NUP98-HOXD13 transgenic mice develop a highly penetrant, severe myelodysplastic syndrome that progresses to acute leukemia. Blood 2005, 106, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kania, A.; Klein, R. Mechanisms of ephrin-Eph signalling in development, physiology and disease. Nat. Rev. Mol. Cell Biol. 2016, 17, 240–256. [Google Scholar] [CrossRef]
- Holder, N.; Klein, R. Eph receptors and ephrins: Effectors of morphogenesis. Development 1999, 126, 2033–2044. [Google Scholar] [CrossRef] [PubMed]
- Anderton, M.; van der Meulen, E.; Blumenthal, M.J.; Schafer, G. The role of the Eph receptor family in tumorigenesis. Cancers 2021, 13, 206. [Google Scholar] [CrossRef]
- Janes, P.W.; Vail, M.E.; Ernst, M.; Scott, A.M. Eph receptors in the immunosuppressive tumor environment. Cancer Res. 2021, 81, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Brantley-Sieders, D.M. Clinical relevance of Ephs and ephrins in cancer: Lessons from breast, colorectal, and lung cancer profiling. Semin. Cell Dev. Biol. 2012, 23, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, W.; Hogan, C. Normal epithelial cells trigger EphA2-dependent RasV12 cell repulsion at the single cell level. Small GTPases 2019, 10, 305–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, A.W.; Ward, L.D.; Wicks, I.P.; Simpson, R.J.; Salvaris, E.; Wilks, A.; Welch, K.; Loudovaris, M.; Rockman, S.; Busmanis, I. Isolation and characterization of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line. J. Biol. Chem. 1992, 267, 3262–3267. [Google Scholar] [CrossRef]
- Dottori, M.; Down, M.; Huttmann, A.; Fitzpatrick, D.R.; Boyd, A.W. Cloning and characterization of EphA3 (Hek) gene promoter: DNA methylation regulates expression in hematopoietic tumor cells. Blood 1999, 94, 2477–2486. [Google Scholar] [CrossRef]
- Guan, M.; Liu, L.; Zhao, X.; Wu, Q.; Yu, B.; Shao, Y.; Yang, H.; Fu, X.; Wan, J.; Zhang, W. Copy number variations of EphA3 are associated with multiple types of hematologic malignancies. Clin. Lymphoma Myeloma Leuk. 2011, 11, 50–53. [Google Scholar] [CrossRef] [Green Version]
- Tomasevic, N.; Luehrsen, K.; Baer, M.; Palath, V.; Martinez, D.; Williams, J.; Yi, C.; Sujatha-Bhaskar, S.; Lanke, R.; Leung, J.; et al. A high affinity recombinant antibody to the human EphA3 receptor with enhanced ADCC activity. Growth Factors 2014, 32, 223–235. [Google Scholar] [CrossRef]
- Ashton, J.M.; Balys, M.; Neering, S.J.; Hassane, D.C.; Cowley, G.; Root, D.E.; Miller, P.G.; Ebert, B.L.; McMurray, H.R.; Land, H.; et al. Gene sets identified with oncogene cooperativity analysis regulate in vivo growth and survival of leukemia stem cells. Cell Stem Cell 2012, 11, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Day, B.W.; Stringer, B.W.; Al-Ejeh, F.; Ting, M.J.; Wilson, J.; Ensbey, K.S.; Jamieson, P.R.; Bruce, Z.C.; Lim, Y.C.; Offenhauser, C.; et al. EphA3 maintains tumorigenicity and is a therapeutic target in glioblastoma multiforme. Cancer Cell 2013, 23, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Song, W.; Amato, K. Eph receptor tyrosine kinases in cancer stem cells. Cytokine Growth Factor Rev. 2015, 26, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Janes, P.W.; Vail, M.E.; Gan, H.K.; Scott, A.M. Antibody Targeting of Eph Receptors in Cancer. Pharmaceuticals 2020, 13, 88. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Cher, L.; Inglis, P.; Lwin, Z.; Lau, E.; Ackermann, U.; Coombs, N.; Remen, K.; Guo, N.; Lee, S.T.; et al. Preliminary Findings of a Phase I Safety and Bioimaging Trial of Kb004 (Ifabotuzumab) in Patients with Glioblastoma. Neuro Oncol. 2019, 21, 6. [Google Scholar] [CrossRef]
- Offenhauser, C.; Al-Ejeh, F.; Puttick, S.; Ensbey, K.S.; Bruce, Z.C.; Jamieson, P.R.; Smith, F.M.; Stringer, B.W.; Carrington, B.; Fuchs, A.V.; et al. EphA3 Pay-Loaded Antibody Therapeutics for the Treatment of Glioblastoma. Cancers 2018, 10, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caivano, A.; La Rocca, F.; Laurenzana, I.; Annese, T.; Tamma, R.; Famigliari, U.; Simeon, V.; Trino, S.; De Luca, L.; Villani, O.; et al. Epha3 acts as proangiogenic factor in multiple myeloma. Oncotarget 2017, 8, 34298–34309. [Google Scholar] [CrossRef]
- Charmsaz, S.; Al-Ejeh, F.; Yeadon, T.M.; Miller, K.J.; Smith, F.M.; Stringer, B.W.; Moore, A.S.; Lee, F.T.; Cooper, L.T.; Stylianou, C.; et al. EphA3 as a target for antibody immunotherapy in acute lymphoblastic leukemia. Leukemia 2017, 31, 1779–1787. [Google Scholar] [CrossRef] [Green Version]
- Swords, R.T.; Greenberg, P.L.; Wei, A.H.; Durrant, S.; Advani, A.S.; Hertzberg, M.S.; Jonas, B.A.; Lewis, I.D.; Rivera, G.; Gratzinger, D.; et al. KB004, a first in class monoclonal antibody targeting the receptor kinase EphA3, in patients with advanced hematologic malignancies: Results from a phase 1 study. Leuk. Res. 2016, 50, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, A.; Pniak, A.; Lemke, G.; Brown, A. EphA3 null mutants do not demonstrate motor axon guidance defects. Mol. Cell Biol. 2003, 23, 8092–8098. [Google Scholar] [CrossRef] [Green Version]
- Barbier, V.; Nowlan, B.; Levesque, J.P.; Winkler, I.G. Flow cytometry analysis of cell cycling and proliferation in mouse hematopoietic stem and progenitor cells. Methods Mol. Biol. 2012, 844, 31–43. [Google Scholar] [CrossRef]
- Janes, P.W.; Slape, C.I.; Farnsworth, R.H.; Atapattu, L.; Scott, A.M.; Vail, M.E. EphA3 biology and cancer. Growth Factors 2014, 32, 176–189. [Google Scholar] [CrossRef]
- Buono, M.; Facchini, R.; Matsuoka, S.; Thongjuea, S.; Waithe, D.; Luis, T.C.; Giustacchini, A.; Besmer, P.; Mead, A.J.; Jacobsen, S.E.; et al. A dynamic niche provides Kit ligand in a stage-specific manner to the earliest thymocyte progenitors. Nat. Cell Biol. 2016, 18, 157–167. [Google Scholar] [CrossRef]
- Rodewald, H.R.; Kretzschmar, K.; Swat, W.; Takeda, S. Intrathymically expressed c-kit ligand (stem cell factor) is a major factor driving expansion of very immature thymocytes in vivo. Immunity 1995, 3, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.M.; Walsh, P.T.; Rudiger, T.; Cotter, T.G.; Mc Carthy, T.V.; Marx, A.; O’Connor, R. EphA3 is induced by CD28 and IGF-1 and regulates cell adhesion. Exp. Cell Res. 2004, 292, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Shields, B.J.; Alserihi, R.; Nasa, C.; Bogue, C.; Alexander, W.S.; McCormack, M.P. Hhex regulates Kit to promote radioresistance of self-renewing thymocytes in Lmo2-transgenic mice. Leukemia 2015, 29, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros-Arias, L.; Silva, J.G.; Paiva, R.A.; Carbonetto, B.; Faisca, P.; Martins, V.C. T Cell Acute Lymphoblastic Leukemia as a Consequence of Thymus Autonomy. J. Immunol. 2019, 202, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Martins, V.C.; Busch, K.; Juraeva, D.; Blum, C.; Ludwig, C.; Rasche, V.; Lasitschka, F.; Mastitsky, S.E.; Brors, B.; Hielscher, T.; et al. Cell competition is a tumour suppressor mechanism in the thymus. Nature 2014, 509, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.C.; Ruggiero, E.; Schlenner, S.M.; Madan, V.; Schmidt, M.; Fink, P.J.; von Kalle, C.; Rodewald, H.R. Thymus-autonomous T cell development in the absence of progenitor import. J. Exp. Med. 2012, 209, 1409–1417. [Google Scholar] [CrossRef] [Green Version]
- Peaudecerf, L.; Lemos, S.; Galgano, A.; Krenn, G.; Vasseur, F.; Di Santo, J.P.; Ezine, S.; Rocha, B. Thymocytes may persist and differentiate without any input from bone marrow progenitors. J. Exp. Med. 2012, 209, 1401–1408. [Google Scholar] [CrossRef] [Green Version]
- Brannan, J.M.; Dong, W.; Prudkin, L.; Behrens, C.; Lotan, R.; Bekele, B.N.; Wistuba, I.; Johnson, F.M. Expression of the receptor tyrosine kinase EphA2 is increased in smokers and predicts poor survival in non-small cell lung cancer. Clin. Cancer Res. 2009, 15, 4423–4430. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pliego Zamora, A.C.; Ranasinghe, H.; Lisle, J.E.; Ng, C.K.; Huang, S.; Wadlow, R.; Scott, A.M.; Boyd, A.W.; Slape, C.I. Cell-Extrinsic Differentiation Block Mediated by EphA3 in Pre-Leukaemic Thymus Contributes to Disease Progression. Cancers 2021, 13, 3858. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153858
Pliego Zamora AC, Ranasinghe H, Lisle JE, Ng CK, Huang S, Wadlow R, Scott AM, Boyd AW, Slape CI. Cell-Extrinsic Differentiation Block Mediated by EphA3 in Pre-Leukaemic Thymus Contributes to Disease Progression. Cancers. 2021; 13(15):3858. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153858
Chicago/Turabian StylePliego Zamora, Adriana C., Hansini Ranasinghe, Jessica E. Lisle, Chun Ki Ng, Stephen Huang, Racheal Wadlow, Andrew M. Scott, Andrew W. Boyd, and Christopher I. Slape. 2021. "Cell-Extrinsic Differentiation Block Mediated by EphA3 in Pre-Leukaemic Thymus Contributes to Disease Progression" Cancers 13, no. 15: 3858. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13153858