
Integrative Transcriptome Profiling Reveals SKA3 as a Novel Prognostic Marker in Non-Muscle Invasive Bladder Cancer


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Samples
2.2. RNA Sequencing Analysis
2.3. Public Expression Profiles
2.4. DEG Analysis
2.5. Single-Cell RNA-Seq Dataset Analysis of Public Data
2.6. Cell Culture
2.7. RNA Interference
2.8. RNA Extraction and Real-Time Quantitative PCR
2.9. Colony Formation Assay
2.10. Flow Cytometry
2.11. Statistical Analysis
3. Results
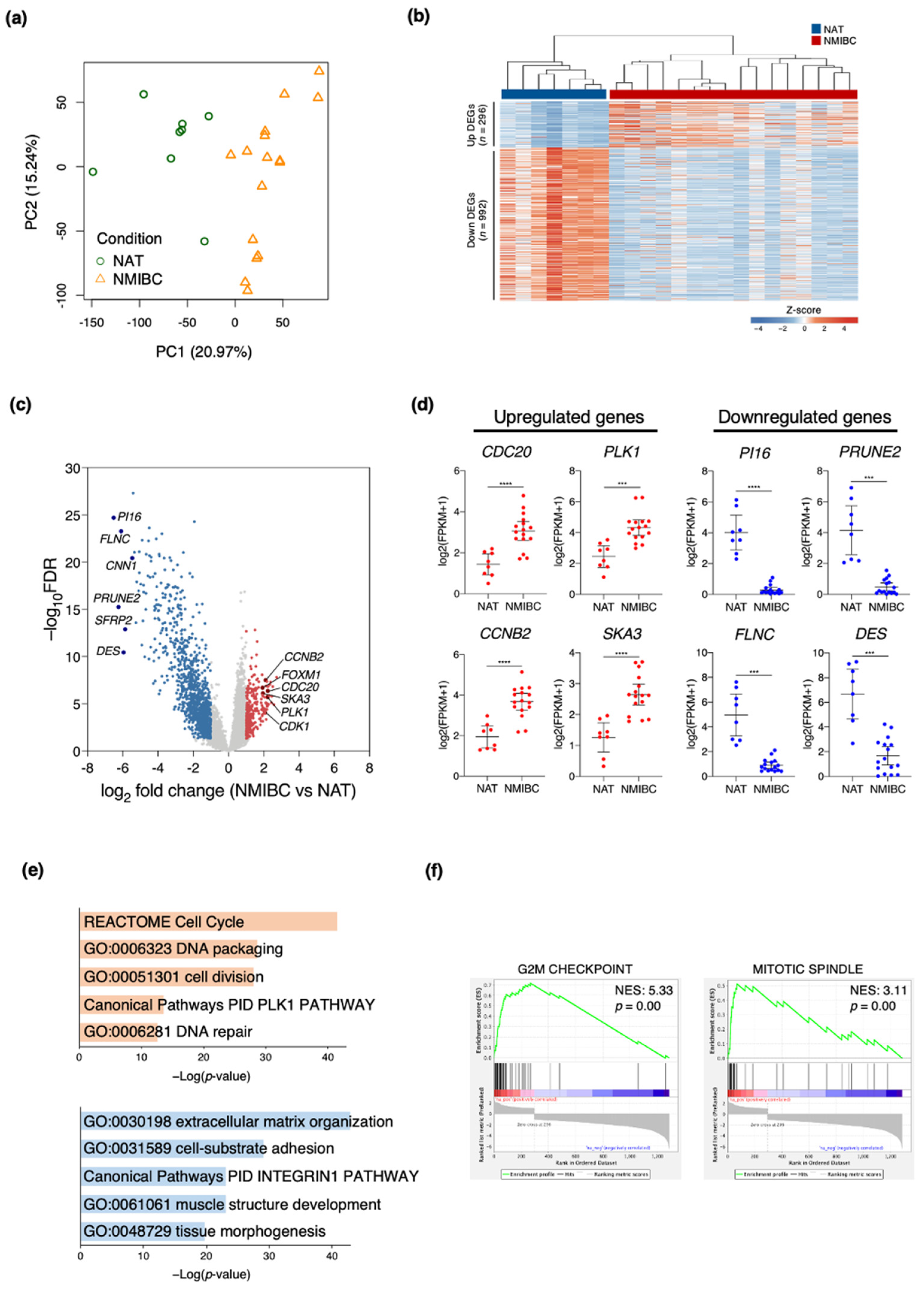
3.1. Transcriptome Profiles Showing Molecular Signatures in Patients with NMIBC
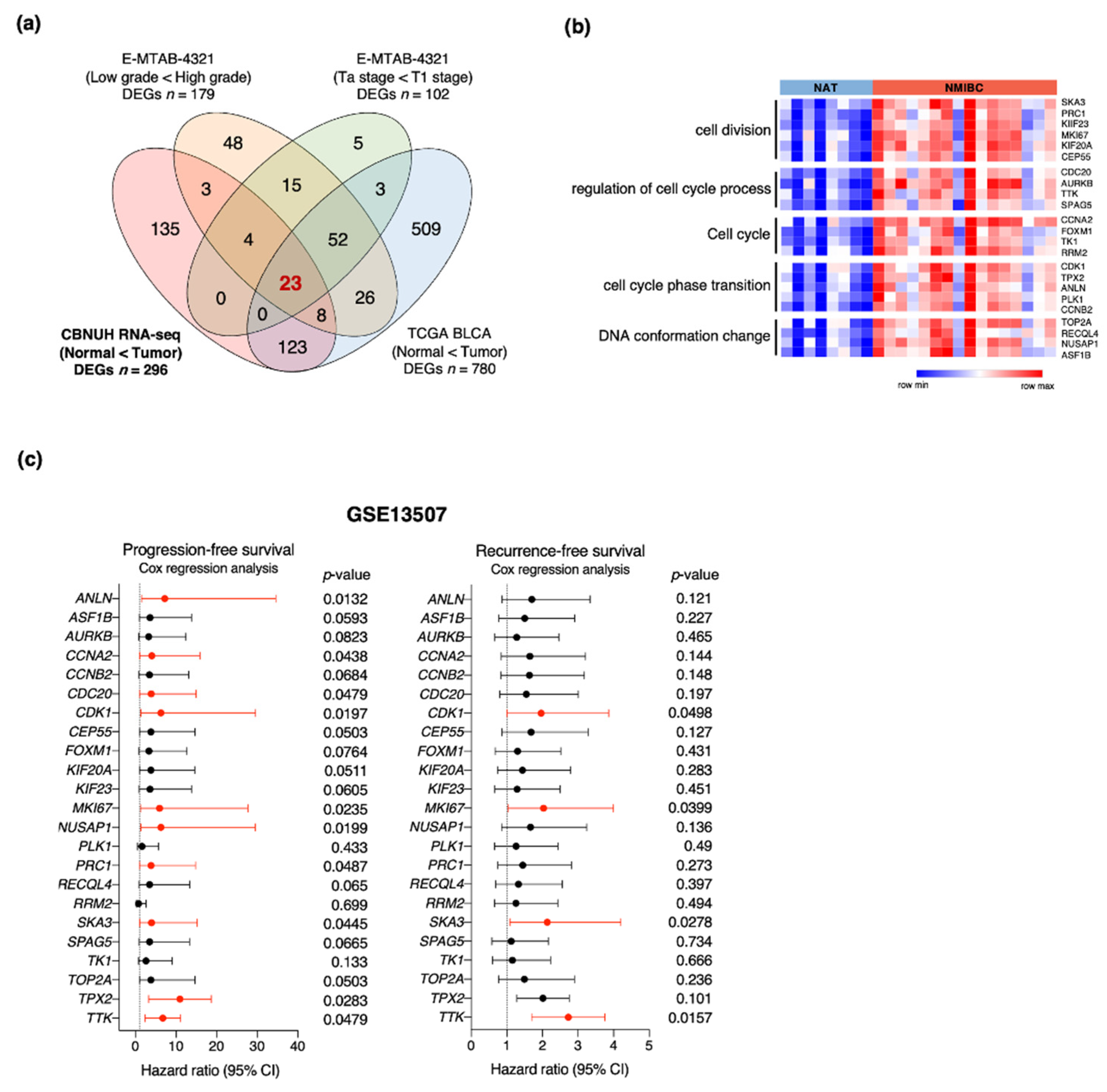
3.2. Identification of Genes Associated with NMIBC Prognosis
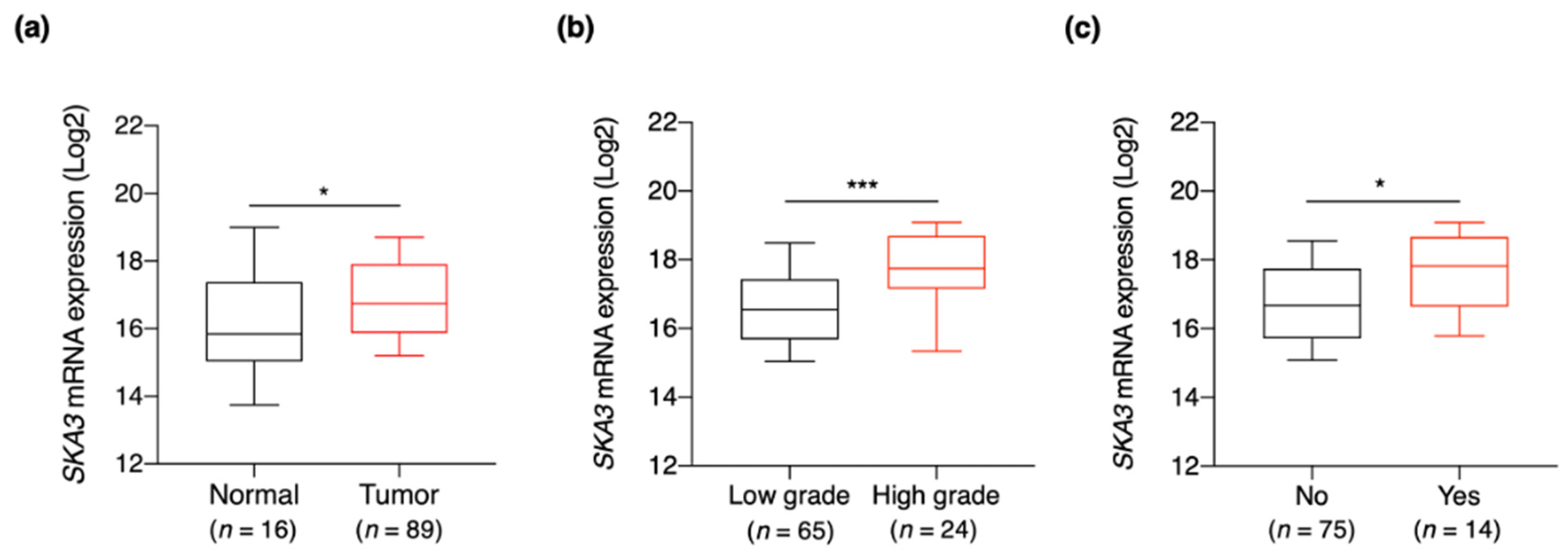
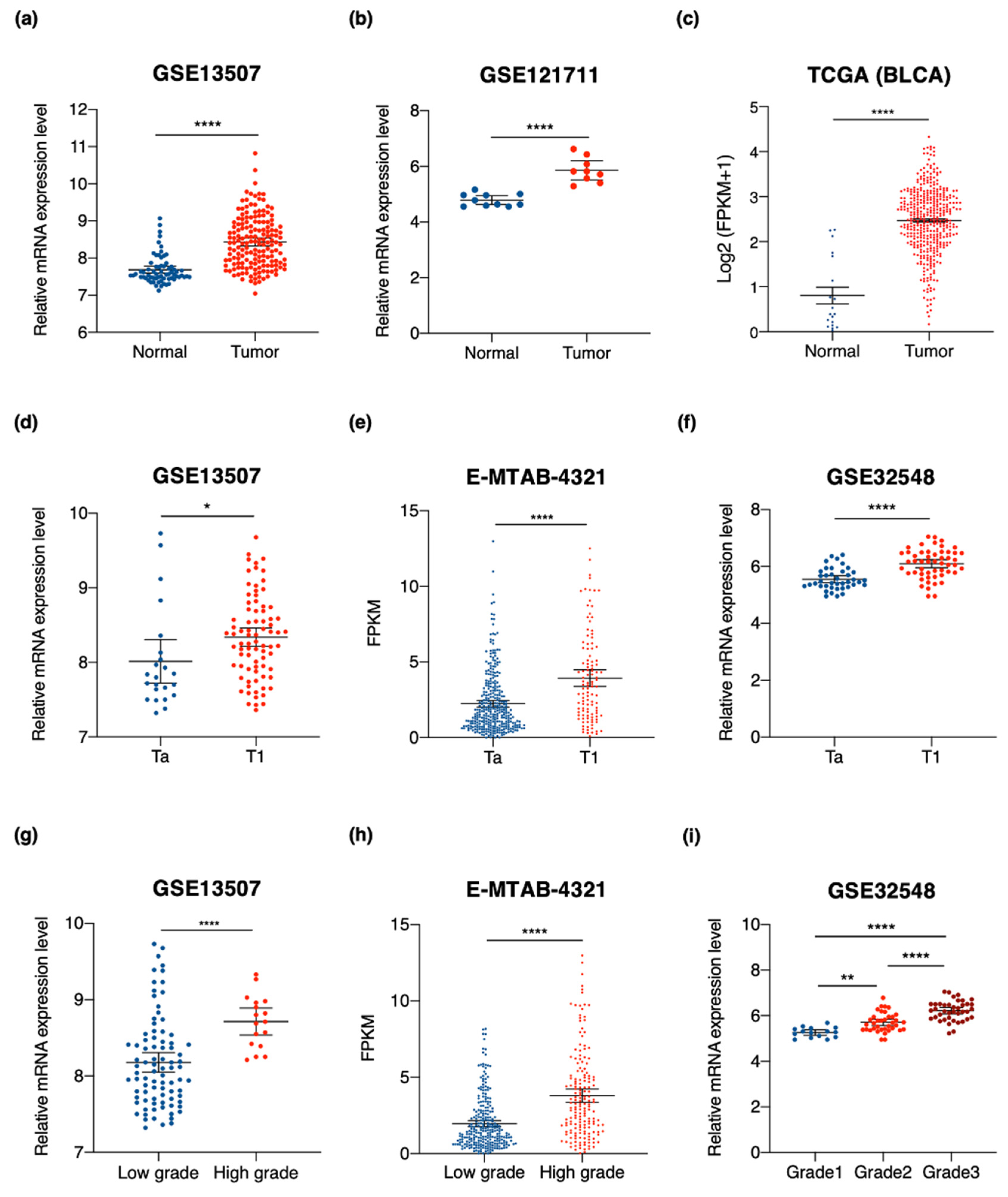
3.3. SKA3 Expression Is Correlated with Clinicopathological Characteristics of Bladder Cancer
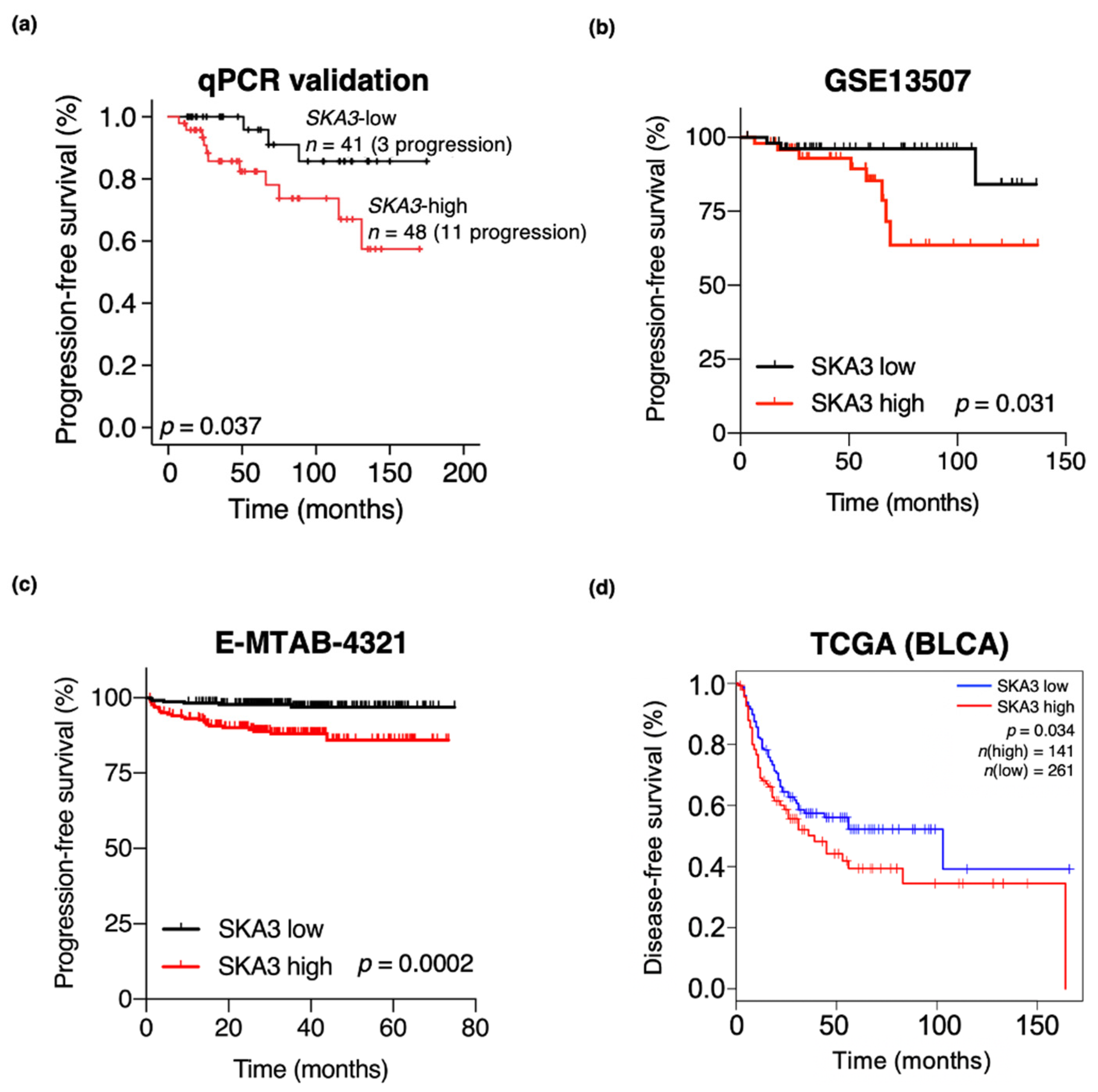
3.4. High SKA3 Expression Is Correlated with NMIBC Patient Prognosis
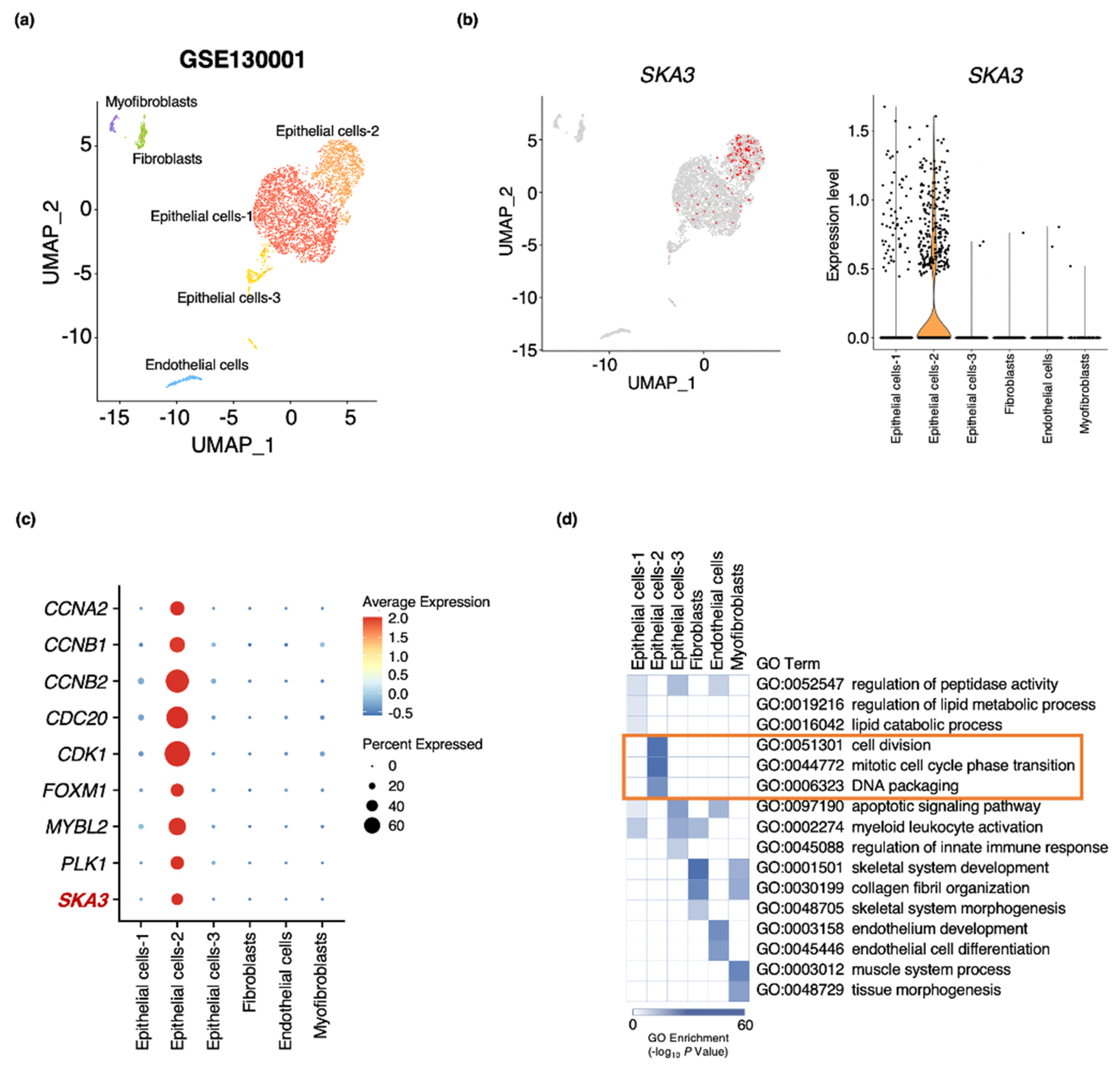
3.5. SKA3 Expression Identified in an Epithelial Cell Subpopulation of Bladder Cancer
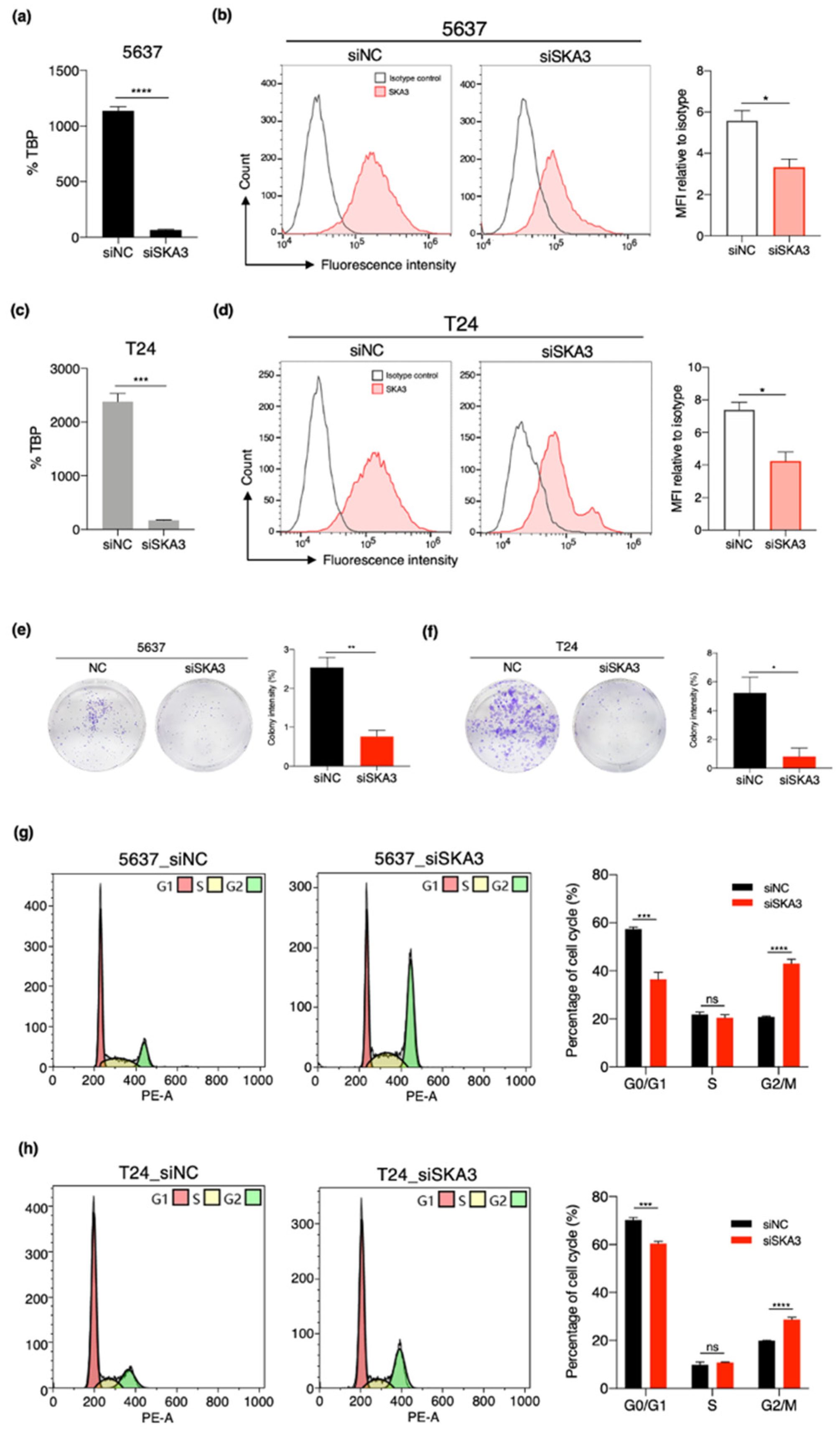
3.6. SKA3 Promotes Cell Proliferation and Cell Cycle Progression in Bladder Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antoni, S.; Ferlay, J.; Soerjomataram, I.; Znaor, A.; Jemal, A.; Bray, F. Bladder cancer incidence and mortality: A global overview and recent trends. Eur. Urol. 2017, 71, 96–108. [Google Scholar] [CrossRef]
- Kobayashi, T.; Owczarek, T.B.; McKiernan, J.M.; Abate-Shen, C. Modelling bladder cancer in mice: Opportunities and challenges. Nat. Rev. Cancer 2015, 15, 42–54. [Google Scholar] [CrossRef] [Green Version]
- Berdik, C. Unlocking bladder cancer. Nature 2017, 551, S34–S35. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.A.; Hurst, C.D. Molecular biology of bladder cancer: New insights into pathogenesis and clinical diversity. Nat. Rev. Cancer 2015, 15, 25–41. [Google Scholar] [CrossRef]
- Sanli, O.; Dobruch, J.; Knowles, M.A.; Burger, M.; Alemozaffar, M.; Nielsen, M.E.; Lotan, Y. Bladder cancer. Nat. Rev. Dis. Primers 2017, 3, 17022. [Google Scholar] [CrossRef]
- Tran, L.; Xiao, J.F.; Agarwal, N.; Duex, J.E.; Theodorescu, D. Advances in bladder cancer biology and therapy. Nat. Rev. Cancer 2021, 21, 104–121. [Google Scholar] [CrossRef]
- Burger, M.; Catto, J.W.; Dalbagni, G.; Grossman, H.B.; Herr, H.; Karakiewicz, P.; Kassouf, W.; Kiemeney, L.A.; La Vecchia, C.; Shariat, S.; et al. Epidemiology and risk factors of urothelial bladder cancer. Eur. Urol. 2013, 63, 234–241. [Google Scholar] [CrossRef]
- Mertens, L.S.; Neuzillet, Y.; Horenblas, S.; van Rhijn, B.W. Landmarks in non-muscle-invasive bladder cancer. Nat. Rev. Urol. 2014, 11, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Matulewicz, R.S.; Steinberg, G.D. Non-muscle-invasive bladder cancer: Overview and contemporary treatment landscape of neoadjuvant chemoablative therapies. Rev. Urol. 2020, 22, 43–51. [Google Scholar] [PubMed]
- Batista, R.; Vinagre, N.; Meireles, S.; Vinagre, J.; Prazeres, H.; Leao, R.; Maximo, V.; Soares, P. Biomarkers for bladder cancer diagnosis and surveillance: A comprehensive review. Diagnostics 2020, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeks, J.J.; Al-Ahmadie, H.; Faltas, B.M.; Taylor, J.A., 3rd; Flaig, T.W.; DeGraff, D.J.; Christensen, E.; Woolbright, B.L.; McConkey, D.J.; Dyrskjot, L. Genomic heterogeneity in bladder cancer: Challenges and possible solutions to improve outcomes. Nat. Rev. Urol. 2020, 17, 259–270. [Google Scholar] [CrossRef]
- Nordentoft, I.; Lamy, P.; Birkenkamp-Demtroder, K.; Shumansky, K.; Vang, S.; Hornshoj, H.; Juul, M.; Villesen, P.; Hedegaard, J.; Roth, A.; et al. Mutational context and diverse clonal development in early and late bladder cancer. Cell Rep. 2014, 7, 1649–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minoli, M.; Kiener, M.; Thalmann, G.N.; Kruithof-de Julio, M.; Seiler, R. Evolution of urothelial bladder cancer in the context of molecular classifications. Int. J. Mol. Sci. 2020, 21, 5670. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.G.; Oh, W.K.; Galsky, M.D. Treatment of muscle-invasive and advanced bladder cancer in 2020. CA Cancer J. Clin. 2020, 70, 404–423. [Google Scholar] [CrossRef] [PubMed]
- Comperat, E.; Varinot, J.; Moroch, J.; Eymerit-Morin, C.; Brimo, F. A practical guide to bladder cancer pathology. Nat. Rev. Urol. 2018, 15, 143–154. [Google Scholar] [CrossRef]
- Mitra, A.P.; Hansel, D.E.; Cote, R.J. Prognostic value of cell-cycle regulation biomarkers in bladder cancer. Semin. Oncol. 2012, 39, 524–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blick, C.G.; Nazir, S.A.; Mallett, S.; Turney, B.W.; Onwu, N.N.; Roberts, I.S.; Crew, J.P.; Cowan, N.C. Evaluation of diagnostic strategies for bladder cancer using computed tomography (CT) urography, flexible cystoscopy and voided urine cytology: Results for 778 patients from a hospital haematuria clinic. BJU Int. 2012, 110, 84–94. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, F.; Jia, C.; Liu, M.; Nan, Y.; Qu, L.; Kong, Q.; Hou, F.; Luo, W.; Na, W.; et al. Comprehensive gene expression analysis in NMIBC using RNA-seq reveals new therapy strategies. Front. Oncol. 2019, 9, 523. [Google Scholar] [CrossRef] [Green Version]
- Zuo, S.; Dai, G.; Ren, X. Identification of a 6-gene signature predicting prognosis for colorectal cancer. Cancer Cell Int. 2019, 19, 6. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Bao, X.; Rogowski, P.; Schafer, C.; Schmidt-Hegemann, N.S.; Unger, K.; Lu, S.; Sun, J.; Buchner, A.; Stief, C.; et al. Establishment and validation of an individualized cell cycle process-related gene signature to predict cancer-specific survival in patients with bladder cancer. Cancers 2020, 12, 1146. [Google Scholar] [CrossRef]
- Kim, S.K.; Roh, Y.G.; Park, K.; Kang, T.H.; Kim, W.J.; Lee, J.S.; Leem, S.H.; Chu, I.S. Expression signature defined by FOXM1-CCNB1 activation predicts disease recurrence in non-muscle-invasive bladder cancer. Clin. Cancer Res. 2014, 20, 3233–3243. [Google Scholar] [CrossRef] [Green Version]
- Rinaldetti, S.; Wirtz, R.; Worst, T.S.; Hartmann, A.; Breyer, J.; Dyrskjot, L.; Erben, P. FOXM1 predicts disease progression in non-muscle invasive bladder cancer. J. Cancer Res. Clin. Oncol. 2018, 144, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Roh, Y.G.; Mun, J.Y.; Kim, S.K.; Park, W.; Jeong, M.S.; Kim, T.N.; Kim, W.T.; Choi, Y.H.; Chu, I.S.; Leem, S.H. Fanconi anemia pathway activation by FOXM1 is critical to bladder cancer recurrence and anticancer drug resistance. Cancers 2020, 12, 1417. [Google Scholar] [CrossRef]
- Yang, J.L.; Wang, C.C.N.; Cai, J.H.; Chou, C.Y.; Lin, Y.C.; Hung, C.C. Identification of GSN and LAMC2 as key prognostic genes of bladder cancer by integrated bioinformatics analysis. Cancers 2020, 12, 1809. [Google Scholar] [CrossRef]
- Bolenz, C.; Lotan, Y. Molecular biomarkers for urothelial carcinoma of the bladder: Challenges in clinical use. Nat. Clin. Pract. Urol. 2008, 5, 676–685. [Google Scholar] [CrossRef]
- Gaitanos, T.N.; Santamaria, A.; Jeyaprakash, A.A.; Wang, B.; Conti, E.; Nigg, E.A. Stable kinetochore-microtubule interactions depend on the Ska complex and its new component Ska3/C13Orf3. EMBO J. 2009, 28, 1442–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daum, J.R.; Wren, J.D.; Daniel, J.J.; Sivakumar, S.; McAvoy, J.N.; Potapova, T.A.; Gorbsky, G.J. Ska3 is required for spindle checkpoint silencing and the maintenance of chromosome cohesion in mitosis. Curr. Biol. 2009, 19, 1467–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Wang, Z.; Huang, S.; Sun, C.; Zhao, J.; Shi, J.; Li, Z.; Wang, Z.; He, X.; Tam, N.L.; et al. SKA3 Promotes tumor growth by regulating CDK2/P53 phosphorylation in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, R.; Wang, M.Q.; Niu, W.B.; Wang, Y.J.; Liu, Y.Y.; Liu, L.Y.; Wang, M.; Zhong, J.; You, H.Y.; Wu, X.H.; et al. SKA3 promotes cell proliferation and migration in cervical cancer by activating the PI3K/Akt signaling pathway. Cancer Cell Int. 2018, 18, 183. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Ksheersagar, P.; Sharma, P. Diagnosis and treatment of bladder cancer. Am. Fam. Physician 2009, 80, 717–723. [Google Scholar]
- Hall, M.C.; Chang, S.S.; Dalbagni, G.; Pruthi, R.S.; Seigne, J.D.; Skinner, E.C.; Wolf, J.S., Jr.; Schellhammer, P.F. Guideline for the management of nonmuscle invasive bladder cancer (stages Ta, T1, and Tis): 2007 update. J. Urol. 2007, 178, 2314–2330. [Google Scholar] [CrossRef]
- Babjuk, M.; Bohle, A.; Burger, M.; Capoun, O.; Cohen, D.; Comperat, E.M.; Hernandez, V.; Kaasinen, E.; Palou, J.; Roupret, M.; et al. EAU guidelines on non-muscle-invasive urothelial carcinoma of the bladder: Update 2016. Eur. Urol. 2017, 71, 447–461. [Google Scholar] [CrossRef]
- Witjes, J.A.; Comperat, E.; Cowan, N.C.; De Santis, M.; Gakis, G.; Lebret, T.; Ribal, M.J.; Van der Heijden, A.G.; Sherif, A. EAU guidelines on muscle-invasive and metastatic bladder cancer: Summary of the 2013 guidelines. Eur. Urol. 2014, 65, 778–792. [Google Scholar] [CrossRef]
- Kim, W.T.; Kim, J.; Yan, C.; Jeong, P.; Choi, S.Y.; Lee, O.J.; Chae, Y.B.; Yun, S.J.; Lee, S.C.; Kim, W.J. S100A9 and EGFR gene signatures predict disease progression in muscle invasive bladder cancer patients after chemotherapy. Ann. Oncol. 2014, 25, 974–979. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, S.; Pohl, M.O.; Zhou, Y.; Rodriguez-Frandsen, A.; Wang, G.; Stein, D.A.; Moulton, H.M.; DeJesus, P.; Che, J.; Mulder, L.C.; et al. Meta- and orthogonal integration of influenza “OMICs” data defines a role for UBR4 in virus budding. Cell Host Microbe 2015, 18, 723–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Sebra, R.P.; Sfakianos, J.P.; Allette, K.; Wang, W.; Yoo, S.; Bhardwaj, N.; Schadt, E.E.; Yao, X.; Galsky, M.D.; et al. A reference profile-free deconvolution method to infer cancer cell-intrinsic subtypes and tumor-type-specific stromal profiles. Genome Med. 2020, 12, 24. [Google Scholar] [CrossRef]
- Guzman, C.; Bagga, M.; Kaur, A.; Westermarck, J.; Abankwa, D. ColonyArea: An ImageJ plugin to automatically quantify colony formation in clonogenic assays. PLoS ONE 2014, 9, e92444. [Google Scholar] [CrossRef] [PubMed]
- Hedegaard, J.; Lamy, P.; Nordentoft, I.; Algaba, F.; Hoyer, S.; Ulhoi, B.P.; Vang, S.; Reinert, T.; Hermann, G.G.; Mogensen, K.; et al. Comprehensive transcriptional analysis of early-stage urothelial carcinoma. Cancer Cell 2016, 30, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Sylvester, R.J.; van der Meijden, A.P.; Oosterlinck, W.; Witjes, J.A.; Bouffioux, C.; Denis, L.; Newling, D.W.; Kurth, K. Predicting recurrence and progression in individual patients with stage Ta T1 bladder cancer using EORTC risk tables: A combined analysis of 2596 patients from seven EORTC trials. Eur. Urol. 2006, 49, 466–477. [Google Scholar] [CrossRef] [PubMed]
- van den Bosch, S.; Alfred Witjes, J. Long-term cancer-specific survival in patients with high-risk, non-muscle-invasive bladder cancer and tumour progression: A systematic review. Eur. Urol. 2011, 60, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Theis, M.; Slabicki, M.; Junqueira, M.; Paszkowski-Rogacz, M.; Sontheimer, J.; Kittler, R.; Heninger, A.K.; Glatter, T.; Kruusmaa, K.; Poser, I.; et al. Comparative profiling identifies C13orf3 as a component of the Ska complex required for mammalian cell division. EMBO J. 2009, 28, 1453–1465. [Google Scholar] [CrossRef]
- Chuang, T.P.; Wang, J.Y.; Jao, S.W.; Wu, C.C.; Chen, J.H.; Hsiao, K.H.; Lin, C.Y.; Chen, S.H.; Su, S.Y.; Chen, Y.J.; et al. Over-expression of AURKA, SKA3 and DSN1 contributes to colorectal adenoma to carcinoma progression. Oncotarget 2016, 7, 45803–45818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Li, J.; Guo, D.; Chen, B.; Liu, P.; Xiao, Y.; Yang, K.; Liu, Z.; Liu, Q. Identification of 13 Key genes correlated with progression and prognosis in hepatocellular carcinoma by weighted gene co-expression network analysis. Front. Genet. 2020, 11, 153. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, Z.R.; Huang, X.; Che, Y.C.; Liu, Q. Identification of spindle and kinetochore-associated family genes as therapeutic targets and prognostic biomarkers in pancreas ductal adenocarcinoma microenvironment. Front. Oncol. 2020, 10, 553536. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.H.; Stoeber, K. The cell cycle and cancer. J. Pathol. 2012, 226, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Lindskrog, S.V.; Prip, F.; Lamy, P.; Taber, A.; Groeneveld, C.S.; Birkenkamp-Demtroder, K.; Jensen, J.B.; Strandgaard, T.; Nordentoft, I.; Christensen, E.; et al. An integrated multi-omics analysis identifies prognostic molecular subtypes of non-muscle-invasive bladder cancer. Nat. Commun. 2021, 12, 2301. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.G.; Groeneveld, C.S.; Jordan, B.; Lin, X.; McLaughlin, K.A.; Das, A.; Fall, L.A.; Fantini, D.; Taxter, T.J.; Mogil, L.S.; et al. Identification of differential tumor subtypes of T1 bladder cancer. Eur. Urol. 2020, 78, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Pettenati, C.; Ingersoll, M.A. Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat. Rev. Urol. 2018, 15, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Gomez, J.; Madero, R.; Solsona, E.; Unda, M.; Martinez-Pineiro, L.; Gonzalez, M.; Portillo, J.; Ojea, A.; Pertusa, C.; Rodriguez-Molina, J.; et al. Predicting nonmuscle invasive bladder cancer recurrence and progression in patients treated with bacillus Calmette-Guerin: The CUETO scoring model. J. Urol. 2009, 182, 2195–2203. [Google Scholar] [CrossRef] [PubMed]
- Palou, J.; Rodriguez-Rubio, F.; Millan, F.; Algaba, F.; Rodriguez-Faba, O.; Huguet, J.; Villavicencio, H. Recurrence at three months and high-grade recurrence as prognostic factor of progression in multivariate analysis of T1G2 bladder tumors. Urology 2009, 73, 1313–1317. [Google Scholar] [CrossRef] [Green Version]
- Solsona, E.; Iborra, I.; Dumont, R.; Rubio-Briones, J.; Casanova, J.; Almenar, S. The 3-month clinical response to intravesical therapy as a predictive factor for progression in patients with high risk superficial bladder cancer. J. Urol. 2000, 164, 685–689. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | NMIBC | NAT | p Value |
|---|---|---|---|
| No. | 16 | 8 | |
| Mean age ± SD | 60.5 ± 10.3 | 60.0 ± 11.5 | 0.809 |
| Gender (%) | 0.722 | ||
| Male | 13 (81.25%) | 6 (75%) | |
| Female | 3 (18.75%) | 2 (25%) | |
| Grade, 2004 WHO grading system (%) | |||
| Low | 11 (68.75%) | ||
| High | 5 (31.25%) | ||
| Stage (%) | |||
| TaN0M0 | 11 (68.75%) | ||
| T1N0M0 | 5 (31.25%) |
| Variable | NMIBC | Control | p Value |
|---|---|---|---|
| No. | 89 | 16 | |
| Mean age ± SD | 68.00 ± 13.68 | 68.00 ± 15.63 | 0.879 |
| Gender (%) | 0.104 | ||
| Male | 76 (85.4%) | 16(100%) | |
| Female | 13 (14.6%) | ||
| Tumor size (%) | |||
| ≤1 cm | 51 (57.3%) | ||
| 2–3 cm | 38 (42.7%) | ||
| >3 cm | 0 | ||
| Multiplicity (%) | |||
| Single | 50 (56.2%) | ||
| 2–7 | 26 (29.2%) | ||
| >7 | 13 (14.6) | ||
| Grade, 2004 WHO grading system (%) | |||
| Low | 65 (73.0%) | ||
| High | 24 (27.0%) | ||
| Stage (%) | |||
| TaN0M0 | 26 (29.2%) | ||
| T1N0M0 | 63 (70.8%) | ||
| BCG therapy (%) | |||
| No | 57 (64.0%) | ||
| Yes | 32 (36.0%) | ||
| Recurrence—no. of patients (%) | |||
| No | 54 (60.7%) | ||
| Yes | 35 (39.3%) | ||
| Progression—no. of patients (%) | |||
| No | 75 (84.3%) | ||
| Yes | 14 (15.7%) | ||
| Survival—no. of patients (%) | |||
| Alive | 60 (67.4%) | ||
| Death | 29 (32.6%) | ||
| Mean follow-up (range)—months | 70.12 (10.70–174.90) |
| Variables | Univariate Cox Analysis | Multivariate Cox Analysis | ||
|---|---|---|---|---|
| HR (95% CI) | p Value | HR (95% CI) | p Value | |
| Age | 1.086 (1.020–1.156) | 0.009 * | 1.083 (1.014–1.157) | 0.017 * |
| Gender | 0.104 | |||
| Male (Ref.) vs. Female | 1.311 (0.290–5.932) | 0.725 | ||
| Tumor size | ||||
| ≤1 cm (Ref.) vs. 2–3 cm | 1.310 (0.378–4.536) | 0.670 | ||
| Multiplicity | ||||
| Single | Ref. | 0.106 | ||
| 2–7 | 2.176 (0.531–8.910) | 0.280 | ||
| >7 | 7.381 (1.153–47.252) | 0.035 * | ||
| 2004 WHO Grade | ||||
| Low (Ref.) vs. High | 5.132 (1.757–14.987) | 0.003 * | 1.926 (0.580–6.390) | 0.284 |
| Stage | ||||
| Ta (Ref.) vs. T1 | 0.698 (0.233–2.087) | 0.520 | ||
| BCG | ||||
| No (Ref.) vs. Yes | 1.663 (0.575–4.808) | 0.348 | ||
| SKA3 expression | ||||
| High expression (Ref.) vs. Low expression | 3.413 (1.065–10.930) | 0.039 * | 4.155 (1.043–16.544) | 0.043 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, C.; Piao, X.-M.; Kang, K.; Kim, Y.-J.; Kang, K. Integrative Transcriptome Profiling Reveals SKA3 as a Novel Prognostic Marker in Non-Muscle Invasive Bladder Cancer. Cancers 2021, 13, 4673. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13184673
You C, Piao X-M, Kang K, Kim Y-J, Kang K. Integrative Transcriptome Profiling Reveals SKA3 as a Novel Prognostic Marker in Non-Muscle Invasive Bladder Cancer. Cancers. 2021; 13(18):4673. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13184673
Chicago/Turabian StyleYou, Chaelin, Xuan-Mei Piao, Keunsoo Kang, Yong-June Kim, and Kyuho Kang. 2021. "Integrative Transcriptome Profiling Reveals SKA3 as a Novel Prognostic Marker in Non-Muscle Invasive Bladder Cancer" Cancers 13, no. 18: 4673. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13184673