
Evaluation of NAB2-STAT6 Fusion Variants and Other Molecular Alterations as Prognostic Biomarkers in a Case Series of 83 Solitary Fibrous Tumors

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Sample Series
2.2. Tissue Microarray (TMA) Construction and Immunohistochemistry (IHC)
2.3. Targeted RNA-Seq
2.4. pTERT Mutation Analysis
2.5. Statistical Analysis
3. Results
3.1. NAB2-STAT6 Fusion Variants Correlate with Patient Age, Tumor Size and Location
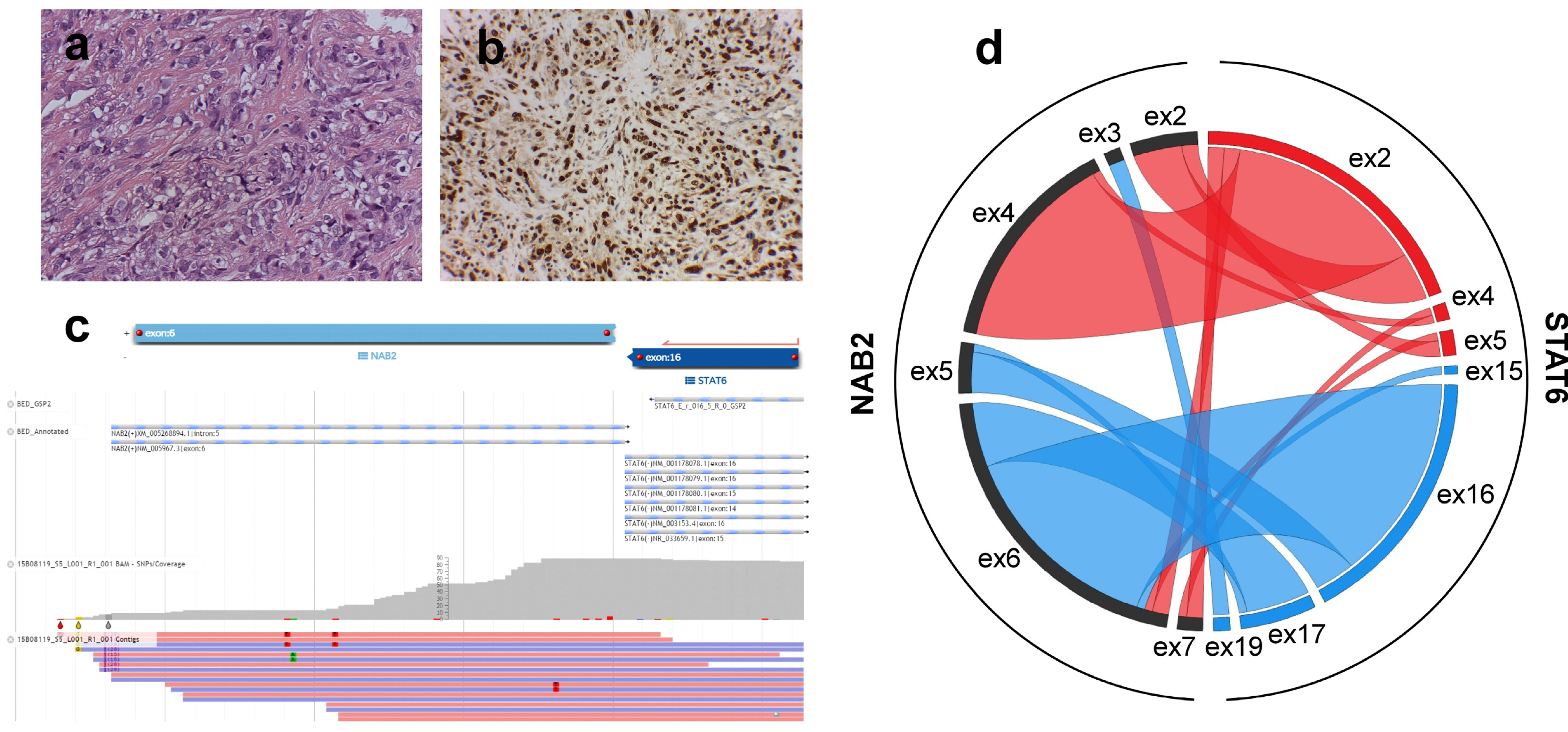
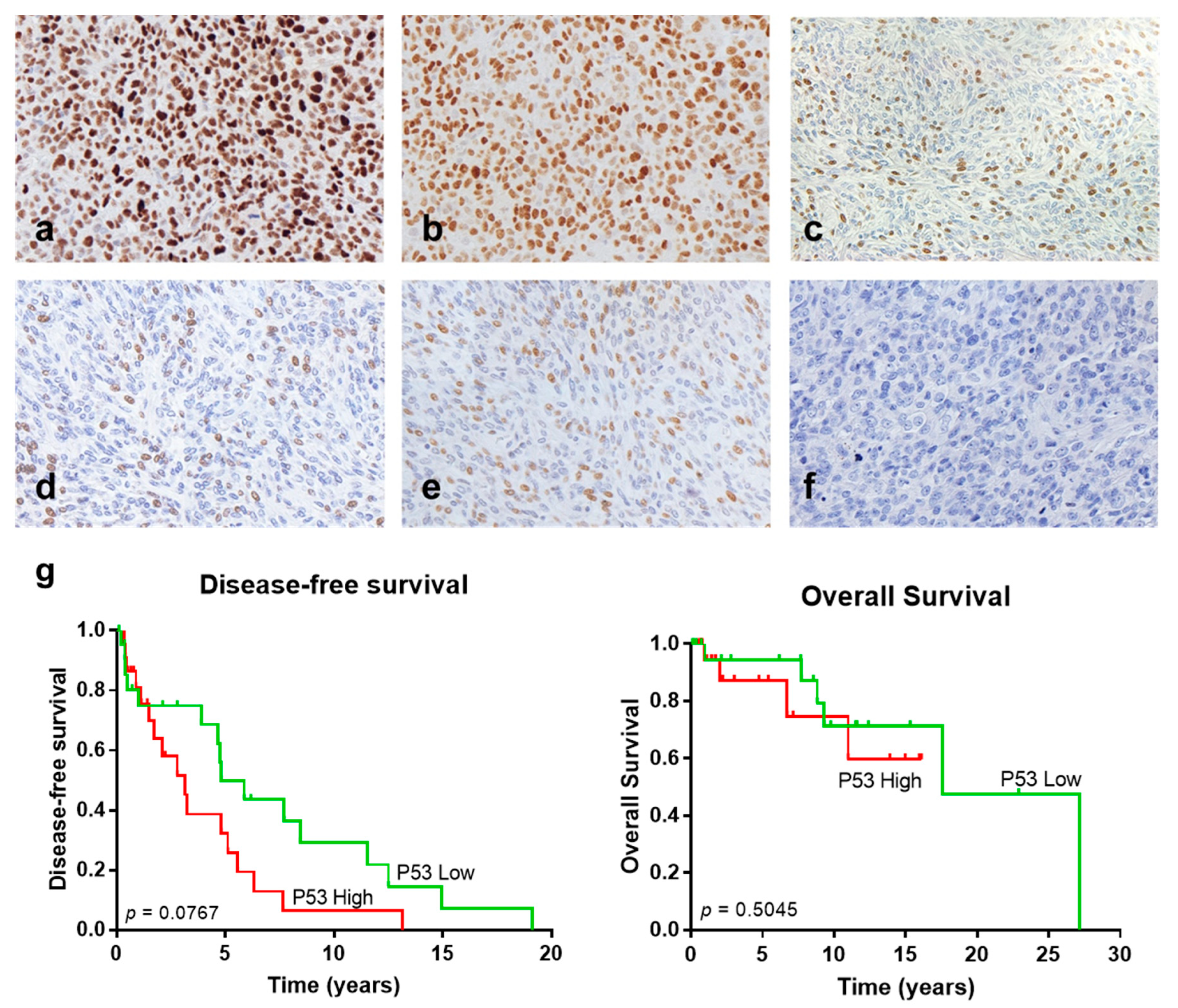
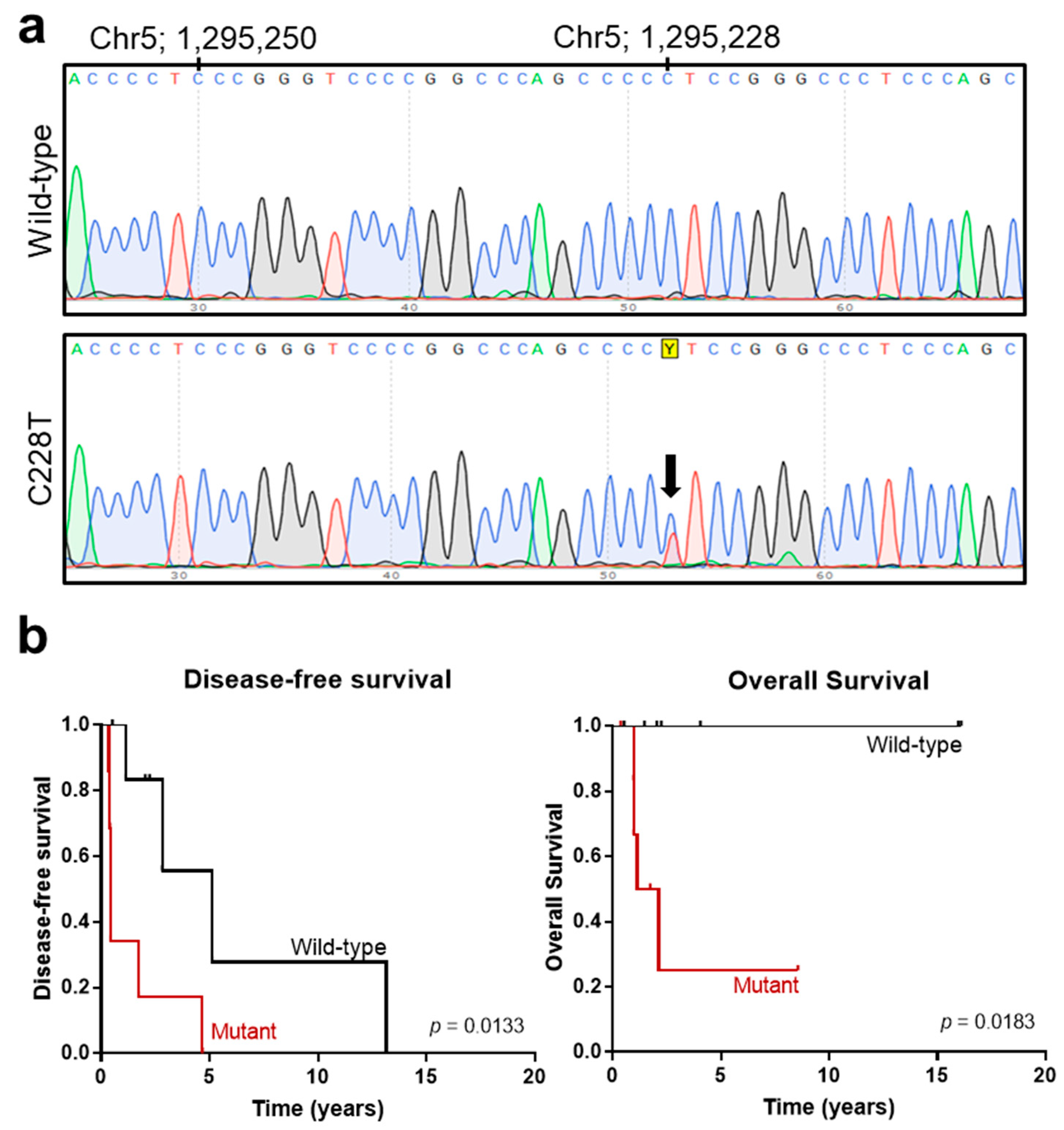
3.2. Analysis of Additional Molecular Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Classification of Tumours Editorial Board. Soft Tissue and Bone Tumours. In WHO Classification of Tumours Series, 5th ed.; IARC Press: Lyon, France, 2020; Volume 3. [Google Scholar]
- Martin-Broto, J.; Mondaza-Hernandez, J.L.; Moura, D.S.; Hindi, N. A Comprehensive Review on Solitary Fibrous Tumor: New Insights for New Horizons. Cancers 2021, 13, 2913. [Google Scholar] [CrossRef] [PubMed]
- Machado, I.; Nieto-Morales, G.; Cruz, J.; Navarro, S.; Giner, F.; Ferrandez, A.; López-Soto, M.V.; Lavernia, J.; Llombart-Bosch, A. Controversial issues in soft tissue solitary fibrous tumors: A pathological and molecular review. Pathol. Int. 2020, 70, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Demicco, E.G.; Wagner, M.J.; Maki, R.G.; Gupta, V.; Iofin, I.; Lazar, A.J.; Wang, W.-L. Risk assessment in solitary fibrous tumors: Validation and refinement of a risk stratification model. Mod. Pathol. 2017, 30, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Georgiesh, T.; Boye, K.; Bjerkehagen, B. A novel risk score to predict early and late recurrence in solitary fibrous tumour. Histopathology 2020, 77, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Demicco, E.G.; Griffin, A.M.; Gladdy, R.A.; Dickson, B.C.; Ferguson, P.C.; Swallow, C.J.; Wunder, J.S.; Wang, W.L. Comparison of published risk models for prediction of outcome in patients with extrameningeal solitary fibrous tumour. Histopathology 2019, 75, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Machado, I.; Nieto Morales, M.G.; Cruz, J.; Lavernia, J.; Giner, F.; Navarro, S.; Ferrandez, A.; Llombart-Bosch, A. Solitary Fibrous Tumor: Integration of Clinical, Morphologic, Immunohistochemical and Molecular Findings in Risk Stratification and Classification May Better Predict Patient outcome. Int. J. Mol. Sci. 2021, 22, 9423. [Google Scholar] [CrossRef] [PubMed]
- Machado, I.; Morales, G.N.; Cruz, J.; Lavernia, J.; Giner, F.; Navarro, S.; Ferrandez, A.; Llombart-Bosch, A. Solitary fibrous tumor: A case series identifying pathological adverse factors-implications for risk stratification and classification. Virchows Arch. Int. J. Pathol. 2020, 476, 597–607. [Google Scholar] [CrossRef]
- Chmielecki, J.; Crago, A.M.; Rosenberg, M.; O’Connor, R.; Walker, S.R.; Ambrogio, L.; Auclair, D.; McKenna, A.; Heinrich, M.C.; Frank, D.A.; et al. Whole-exome sequencing identifies a recurrent NAB2-STAT6 fusion in solitary fibrous tumors. Nat. Genet. 2013, 45, 131–132. [Google Scholar] [CrossRef] [Green Version]
- Robinson, D.R.; Wu, Y.M.; Kalyana-Sundaram, S.; Cao, X.; Lonigro, R.J.; Sung, Y.S.; Chen, C.L.; Zhang, L.; Wang, R.; Su, F.; et al. Identification of recurrent NAB2-STAT6 gene fusions in solitary fibrous tumor by integrative sequencing. Nat. Genet. 2013, 45, 180–185. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.C.; Huang, H.Y. Solitary fibrous tumor: An evolving and unifying entity with unsettled issues. Histol. Histopathol. 2019, 34, 313–334. [Google Scholar] [CrossRef]
- Bieg, M.; Moskalev, E.A.; Will, R.; Hebele, S.; Schwarzbach, M.; Schmeck, S.; Hohenberger, P.; Jakob, J.; Kasper, B.; Gaiser, T.; et al. Gene Expression in Solitary Fibrous Tumors (SFTs) Correlates with Anatomic Localization and NAB2-STAT6 Gene Fusion Variants. Am. J. Pathol. 2021, 191, 602–617. [Google Scholar] [CrossRef]
- Georgiesh, T.; Namløs, H.M.; Sharma, N.; Lorenz, S.; Myklebost, O.; Bjerkehagen, B.; Meza-Zepeda, L.A.; Boye, K. Clinical and molecular implications of NAB2-STAT6 fusion variants in solitary fibrous tumour. Pathology 2021, 53, 713–719. [Google Scholar] [CrossRef]
- Barthelmess, S.; Geddert, H.; Boltze, C.; Moskalev, E.A.; Bieg, M.; Sirbu, H.; Brors, B.; Wiemann, S.; Hartmann, A.; Agaimy, A.; et al. Solitary fibrous tumors/hemangiopericytomas with different variants of the NAB2-STAT6 gene fusion are characterized by specific histomorphology and distinct clinicopathological features. Am. J. Pathol. 2014, 184, 1209–1218. [Google Scholar] [CrossRef]
- Park, H.K.; Yu, D.B.; Sung, M.; Oh, E.; Kim, M.; Song, J.Y.; Lee, M.S.; Jung, K.; Noh, K.W.; An, S.; et al. Molecular changes in solitary fibrous tumor progression. J. Mol. Med. 2019, 97, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.C.; Li, C.F.; Kao, Y.C.; Chuang, I.C.; Tai, H.C.; Tsai, J.W.; Yu, S.C.; Huang, H.Y.; Lan, J.; Yen, S.L.; et al. The clinicopathological significance of NAB2-STAT6 gene fusions in 52 cases of intrathoracic solitary fibrous tumors. Cancer Med. 2016, 5, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Akaike, K.; Kurisaki-Arakawa, A.; Hara, K.; Suehara, Y.; Takagi, T.; Mitani, K.; Kaneko, K.; Yao, T.; Saito, T. Distinct clinicopathological features of NAB2-STAT6 fusion gene variants in solitary fibrous tumor with emphasis on the acquisition of highly malignant potential. Hum. Pathol. 2015, 46, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Tai, H.C.; Chuang, I.C.; Chen, T.C.; Li, C.F.; Huang, S.C.; Kao, Y.C.; Lin, P.C.; Tsai, J.W.; Lan, J.; Yu, S.C.; et al. NAB2-STAT6 fusion types account for clinicopathological variations in solitary fibrous tumors. Mod. Pathol. 2015, 28, 1324–1335. [Google Scholar] [CrossRef] [PubMed]
- Chuang, I.C.; Liao, K.C.; Huang, H.Y.; Kao, Y.C.; Li, C.F.; Huang, S.C.; Tsai, J.W.; Chen, K.C.; Lan, J.; Lin, P.C. NAB2-STAT6 gene fusion and STAT6 immunoexpression in extrathoracic solitary fibrous tumors: The association between fusion variants and locations. Pathol. Int. 2016, 66, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Fritchie, K.; Jensch, K.; Moskalev, E.A.; Caron, A.; Jenkins, S.; Link, M.; Brown, P.D.; Rodriguez, F.J.; Guajardo, A.; Brat, D.; et al. The impact of histopathology and NAB2-STAT6 fusion subtype in classification and grading of meningeal solitary fibrous tumor/hemangiopericytoma. Acta Neuropathol. 2019, 137, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, Y.; Kohashi, K.; Kinoshita, I.; Yamamoto, H.; Iwasaki, T.; Yoshimoto, M.; Ishihara, S.; Toda, Y.; Itou, Y.; Koga, Y.; et al. Clinicopathological review of solitary fibrous tumors: Dedifferentiation is a major cause of patient death. Virchows Arch. Int. J. Pathol. 2019, 475, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Demicco, E.G.; Wani, K.; Ingram, D.; Wagner, M.; Maki, R.G.; Rizzo, A.; Meeker, A.; Lazar, A.J.; Wang, W.L. TERT promoter mutations in solitary fibrous tumour. Histopathology 2018, 73, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Vogels, R.; Macagno, N.; Griewank, K.; Groenen, P.; Verdijk, M.; Fonville, J.; Kusters, B.; Bouvier, C.; Cornu, P.; Dufour, H.; et al. Prognostic significance of NAB2–STAT6 fusion variants and TERT promotor mutations in solitary fibrous tumors/hemangiopericytomas of the CNS: Not (yet) clear. Acta Neuropathol. 2019, 137, 679–682. [Google Scholar] [CrossRef]
- Bahrami, A.; Lee, S.; Schaefer, I.-M.; Boland, J.M.; Patton, K.T.; Pounds, S.; Fletcher, C.D. TERT promoter mutations and prognosis in solitary fibrous tumor. Mod. Pathol. 2016, 29, 1511–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Seger, N.; Tsagkozis, P.; Hesla, A.C.; Ghaderi, M.; Chen, Y.; Ehnman, M.; Warsito, D.; Wejde, J.; Larsson, O.; et al. Telomerase promoter mutations and copy number alterations in solitary fibrous tumours. J. Clin. Pathol. 2018, 71, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Schirosi, L.; Lantuejoul, S.; Cavazza, A.; Murer, B.; Yves Brichon, P.; Migaldi, M.; Sartori, G.; Sgambato, A.; Rossi, G. Pleuro-pulmonary solitary fibrous tumors: A clinicopathologic, immunohistochemical, and molecular study of 88 cases confirming the prognostic value of de Perrot staging system and p53 expression, and evaluating the role of c-kit, BRAF, PDGFRs (alpha/beta), c-met, and EGFR. Am. J. Surg. Pathol. 2008, 32, 1627–1642. [Google Scholar] [CrossRef]
- Argani, P.; Kao, Y.C.; Zhang, L.; Sung, Y.S.; Alaggio, R.; Swanson, D.; Matoso, A.; Dickson, B.C.; Antonescu, C.R. BCOR Overexpression in Renal Malignant Solitary Fibrous Tumors: A Close Mimic of Clear Cell Sarcoma of Kidney. Am. J. Surg. Pathol. 2019, 43, 773–782. [Google Scholar] [CrossRef]
- Kao, Y.C.; Sung, Y.S.; Argani, P.; Swanson, D.; Alaggio, R.; Tap, W.; Wexler, L.; Dickson, B.C.; Antonescu, C.R. NTRK3 overexpression in undifferentiated sarcomas with YWHAE and BCOR genetic alterations. Mod. Pathol. 2020, 33, 1341–1349. [Google Scholar] [CrossRef]
- Martin-Broto, J.; Cruz, J.; Penel, N.; Le Cesne, A.; Hindi, N.; Luna, P.; Moura, D.S.; Bernabeu, D.; de Alava, E.; Lopez-Guerrero, J.A.; et al. Pazopanib for treatment of typical solitary fibrous tumours: A multicentre, single-arm, phase 2 trial. Lancet Oncol. 2020, 21, 456–466. [Google Scholar] [CrossRef]
- Martin-Broto, J.; Stacchiotti, S.; Lopez-Pousa, A.; Redondo, A.; Bernabeu, D.; de Alava, E.; Casali, P.G.; Italiano, A.; Gutierrez, A.; Moura, D.S.; et al. Pazopanib for treatment of advanced malignant and dedifferentiated solitary fibrous tumour: A multicentre, single-arm, phase 2 trial. Lancet Oncol. 2019, 20, 134–144. [Google Scholar] [CrossRef]
- Rodriguez-Nunez, P.; Romero-Perez, L.; Amaral, A.T.; Puerto-Camacho, P.; Jordan, C.; Marcilla, D.; Grunewald, T.G.; Alonso, J.; de Alava, E.; Diaz-Martin, J. Hippo pathway effectors YAP1/TAZ induce an EWS-FLI1-opposing gene signature and associate with disease progression in Ewing sarcoma. J. Pathol. 2020, 250, 374–386. [Google Scholar] [CrossRef] [Green Version]
- Rudzinski, E.R.; Lockwood, C.M.; Stohr, B.A.; Vargas, S.O.; Sheridan, R.; Black, J.O.; Rajaram, V.; Laetsch, T.W.; Davis, J.L. Pan-Trk Immunohistochemistry Identifies NTRK Rearrangements in Pediatric Mesenchymal Tumors. Am. J. Surg. Pathol. 2018, 42, 927–935. [Google Scholar] [CrossRef]
- Dagrada, G.P.; Spagnuolo, R.D.; Mauro, V.; Tamborini, E.; Cesana, L.; Gronchi, A. Solitary fibrous tumors: Loss of chimeric protein expression and genomic instability mark dedifferentiation. Mod. Pathol. 2015, 28, 1074–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoi, T.; Tsuzuki, T.; Yatabe, Y.; Suzuki, M.; Kurumaya, H.; Koshikawa, T.; Kuhara, H.; Kuroda, M.; Nakamura, N.; Nakatani, Y.; et al. Solitary fibrous tumour: Significance of p53 and CD34 immunoreactivity in its malignant transformation. Histopathology 1998, 32, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Koelsche, C.; Sahm, F.; Capper, D.; Reuss, D.; Sturm, D.; Jones, D.T.; Kool, M.; Northcott, P.A.; Wiestler, B.; Böhmer, K.; et al. Distribution of TERT promoter mutations in pediatric and adult tumors of the nervous system. Acta Neuropathol. 2013, 126, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, G.; Sambri, A.; Pedrini, E.; Pazzaglia, L.; Sangiorgi, L.; Ruengwanichayakun, P.; Donati, D.; Benassi, M.S.; Righi, A. Histological and molecular features of solitary fibrous tumor of the extremities: Clinical correlation. Virchows Arch. 2020, 476, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, F.; Bouvier-Labit, C.; Finetti, P.; Metellus, P.; Adelaide, J.; Mokhtari, K.; Figarella-Branger, D.; Decouvelaere, A.V.; Miquel, C.; Coindre, J.M.; et al. Gene expression profiling of solitary fibrous tumors. PLoS ONE 2013, 8, e64497. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Kim, H.S.; Kim, J.H.; Choi, S.H.; Kim, D.S.; Ryoo, Z.Y.; Kim, J.Y.; Lee, S. NAB2-STAT6 fusion protein mediates cell proliferation and oncogenic progression via EGR-1 regulation. Biochem. Biophys. Res. Commun. 2020, 526, 287–292. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | GEIS Cohort | Local Cohort | Whole Series |
|---|---|---|---|
| Sex | n = 50 | n = 31 | n = 81 |
| Male | 22 (44) | 13 (42) | 35 (43) |
| Female | 28 (56) | 18 (58) | 46 (57) |
| Age (years) | n = 50 | n = 30 a | n = 80 a |
| Mean ± SD | 54.5 ± 16.5 | 55.8 ± 13.5 | 55.1 ± 15.4 |
| Median (Range) | 54 (20–87) | 56 (27–79) | 55 (20–87) |
| Size, mm | n = 34 a | n = 21 a | n = 55 a |
| Mean ± SD | 95.2 ± 59.7 | 90.9 ± 51.0 | 93.5 ± 56.1 |
| Median (Range) | 85 (15–270) | 95 (15–210) | 85 (15–270) |
| Location | n = 50 | n = 31 * | n = 81 |
| Abdominal/Visceral | 11 (22) | 13 (42) | 24 (30) |
| Pleuro-pulmonary | 13 (26) | 8 (26) | 21 (26) |
| Meningeal | 9 (18) | 2 (6) | 11 (14) |
| Extremity | 6 (12) | 4 (13) | 10 (12) |
| Head and Neck | 7 (14) | 2 (6) | 9 (11) |
| Trunk | 4 (8) | 2 (6) | 6 (7) |
| Mitotic count (/10 HPFs) | n = 50 | n = 29 a | n = 79 a |
| Mean ± SD | 5.5 ± 8.3 | 3.8 ± 5.0 | 4.9 ± 7.3 |
| Median (Range) | 3 (0–48) | 2 (0–17) | 3 (0–48) |
| <4 | 27 (54) | 20 (69) | 47 (59) |
| ≥4 | 23 (46) | 9 (31) | 32 (41) |
| Necrosis | n = 49 a | n = 25 a | n = 74 a |
| Absent | 41 (84) | 21 (84) | 62 (84) |
| Present | 8 (16) | 4 (16) | 12 (16) |
| Follow-up for OS | |||
| Median, months (Range) | 86 (9–326) | 17 (1–132) | 48 (1–326) |
| Follow-up for DFS | |||
| Median, months (Range) | 47 (4–230) | 17 (1–74) | 28 (1–229) |
| Recurrence/progression | n = 49 a | n = 27 a | n = 76 a |
| No | 0 (0) | 21 (78) | 21 (28) |
| Yes | 49 (100) | 6 (22) | 55 (72) |
| Outcome | n = 49 a | n = 27 a | n = 76 a |
| Alive | 33 (67) | 26 (96) | 59 (78) |
| Dead of disease | 16 (33) | 1 (4) | 17 (22) |
| GEIS Cohort | Local Cohort | Whole Series | |||||||
|---|---|---|---|---|---|---|---|---|---|
| NAB2-STAT6 Variant | TAD | FULL | p Value | TAD | FULL | p Value | TAD | FULL | p Value |
| Relative frequency | |||||||||
| N | 25 (50) | 25 (50) | 18 (55) | 15 (45) | 43 (52) | 40 (48) | |||
| Sex | 0.612 | 0.722 | 0.8241 | ||||||
| Male | 11 (50) | 11 (50) | 6 (46) | 7 (54) | 17 (49) | 18 (51) | |||
| Female | 14 (50) | 14 (50) | 10 (56) | 8 (44) | 24 (52) | 22 (48) | |||
| Age (years) | 0.001 | 0.448 | 0.001 | ||||||
| Mean (± SD) | 46.7 ± 15.0 | 62.3 ± 14.8 | 53.9 ± 13.4 | 57.6 ± 13.9 | 49.6 ± 14.7 | 61.0 ± 14.1 | |||
| Median (Range) | 51 (20–79) | 66 (31–87) | 54 (27–79) | 58 (32–77) | 52 (20–79) | 65 (31–87) | |||
| Size (mm) | 0.041 | 0.468 | 0.225 | ||||||
| Means ± SD | 76.0 ± 51.9 | 114.4 ± 63.6 | 98.3 ± 55.9 | 84.1 ± 47.8 | 84.6 ± 53.5 | 101.5 ± 58.1 | |||
| Median (Range) | 65 (15–200) | 100 (30–270) | 98 (15–210) | 75 (35–200) | 73 (15–210) | 95 (30–270) | |||
| Location | 0.048 | 0.019 | 0.0029 | ||||||
| Abdominal/Visceral | 4 (36) | 7 (64) | 11 (85) | 2 (15) | 15 (67) | 9 (37) | |||
| Pleuro-pulmonary | 3 (23) | 10 (77) | 1 (12) | 7 (88) | 4 (19) | 17 (81) | |||
| Meningeal | 8 (89) | 1 (11) | 2 (100) | 0 (0) | 10 (91) | 1 (9) | |||
| Extremity | 3 (50) | 3 (50) | 1 (25) | 3 (75) | 4 (40) | 6 (60) | |||
| Head and Neck | 4 (57) | 3 (43) | 1 (50) | 1 (50) | 5 (56) | 4 (44) | |||
| Trunk | 3 (75) | 1 (25) | 1 (50) | 1 (50) | 4 (67) | 2 (33) | |||
| Necrosis | 0.327 | 0.645 | 0.2159 | ||||||
| Absent | 22 (54) | 19 (46) | 10 (55) | 8 (45) | 34 (55) | 28 (45) | |||
| Present | 3 (51) | 5 (49) | 7 (40) | 4 (60) | 4 (33) | 8 (67) | |||
| Mitotic count/10 HPFs | 0.571 | 0.717 | 0.6543 | ||||||
| <4 | 15 (56) | 12 (44) | 10 (56) | 8 (44) | 26 (55) | 21 (45) | |||
| ≥4 | 10 (43) | 13 (57) | 7 (64) | 4 (36) | 16 (50) | 16 (50) | |||
| Recurrence/progression | - | 0.165 | 0.445 | ||||||
| No | 0 (0) | 0 (0) | 9 (43) | 12 (57) | 9 (43) | 12 (57) | |||
| Yes | 25 (51) | 24 (49) | 5 (83) | 1 (17) | 30 (55) | 25 (44) | |||
| Outcome | 1 | 1 | 1 | ||||||
| Alive | 17 (52) | 16 (48) | 13 (50) | 13 (50) | 30 (51) | 29 (49) | |||
| Dead of disease | 8 (50) | 8 (50) | 1 (100) | 0 (0) | 9 (53) | 8 (47) | |||
| P53 IHC (n = 44) | BCOR IHC (n = 45) | pan–TRK IHC (n = 44) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Low | High | p Value | Low | High | p Value | Negative | Positive | p Value | |
| n = 22 | n = 22 | n = 27 | n = 18 | n = 13 | n = 31 | ||||
| Gender | 0.7546 | 0.5372 | 0.4964 | ||||||
| Male | 7 (44) | 9 (56) | 9 (53) | 8 (47) | 6 (37) | 10 (66) | |||
| Female | 15 (54) | 13 (46) | 18 (64) | 10 (36) | 7 (25) | 21 (75) | |||
| Age, years | 0.3126 | 0.3248 | 0.0679 | ||||||
| Mean ± SD | 48.7 ± 14.9 | 53.9 ± 17.2 | 53.2 ± 16.9 | 48.7 ± 14.5 | 44.7 ± 15.3 | 54.1 ± 15.9 | |||
| Median (Range) | 50 (28–87) | 50.5 (20–79) | 52 (20–87) | 45 (27–79) | 40 (20–78) | 52 (27–87) | |||
| Size (mm) | 0.2988 | 0.0013 | 0.7031 | ||||||
| Mean ± SD | 108.9 ± 57.9 | 91.9 ± 67.5 | 125.7 ± 62.8 | 62.0 ± 33.1 | 89.6 ± 49.0 | 106.9 ± 68.4 | |||
| Median (Range) | 100 (27–210) | 85 (15–270) | 110 (30–270) | 60 (15–130) | 98 (15–200) | 90 (27–270) | |||
| Locations | 0.1915 | 0.0221 | 0.1894 | ||||||
| Abdominal/Visceral | 6 (55) | 5 (45) | 8 (73) | 3 (27) | 2 (18) | 9 (82) | |||
| Pleuro–pulmonary | 8 (62) | 5 (38) | 12 (92) | 1 (8) | 6 (46) | 7 (54) | |||
| Meningeal | 3 (37) | 5 (63) | 3 (37) | 5 (63) | 1 (12) | 7 (88) | |||
| Extremity | 0 (0) | 4 (100) | 2 (40) | 3 (60) | 0 (0) | 4 (100) | |||
| Head and Neck | 4 (80) | 1 (20) | 1 (20) | 4 (80) | 3 (60) | 2 (40) | |||
| Trunk | 1 (33) | 2 (67) | 1 (33) | 2 (67) | 1 (33) | 2 (67) | |||
| Mitotic counts/10 HPFs | 0.0278 | 0.1678 | 0.4832 | ||||||
| <4 | 13 (72) | 5 (28) | 17 (57) | 13 (43) | 6 (33) | 12 (67) | |||
| ≥4 | 8 (33) | 16 (67) | 9 (82) | 2 (18) | 5 (21) | 19 (79) | |||
| Necrosis | 0.4882 | 0.1678 | 0.2268 | ||||||
| Absent | 14 (48) | 15 (52) | 17 (57) | 13 (43) | 5 (17) | 24 (83) | |||
| Present | 7 (64) | 4 (36) | 9 (82) | 2 (18) | 4 (36) | 7 (64) | |||
| Fusion Variant | 0.364 | 0.0027 | 0.522 | ||||||
| STAT6–TAD | 14 (58) | 10 (42) | 10 (40) | 15 (60) | 6 (25) | 18 (75) | |||
| STAT6–FULL | 8 (40) | 12 (60) | 17 (85) | 3 (15) | 7 (35) | 13 (65) | |||
| Recurrence/progression | >0.9999 | 0.7323 | 0.0078 | ||||||
| No | 6 (55) | 5 (45) | 6 (55) | 5 (45) | 7 (64) | 4 (36) | |||
| Yes | 16 (48) | 17 (52) | 21 (62) | 13 (38) | 6 (18) | 27 (82) | |||
| Outcome | 0.7205 | 0.7323 | 0.0212 | ||||||
| Alive | 16 (47) | 18 (53) | 21 (62) | 13 (38) | 13 (38) | 21 (62) | |||
| Died of disease | 6 (60) | 4 (40) | 6 (55) | 5 (45) | 0 (0) | 10 (100) | |||
| pTERT Mutational Status | |||
|---|---|---|---|
| Wild-Type | Mutant | p Value | |
| n = 9 | n = 9 | ||
| Gender | 0.6372 | ||
| Male (%) | 6 (60) | 4 (40) | |
| Female (%) | 3 (37) | 5 (63) | |
| Age, years | 0.0876 | ||
| Mean ± SD | 48.7 ± 17.1 | 63.1 ± 16.5 | |
| Median (Range) | 49 (20–70) | 64.5 (31–87) | |
| Size (mm) | 1 | ||
| Mean ± SD | 122.0 ± 94.4 | 104.4 ± 46.8 | |
| Median (Range) | 120 (15–270) | 100 (37–165) | |
| Locations | 0.0537 | ||
| Abdominal/Visceral (%) | 6 (86) | 1 (14) | |
| Pleuro-pulmonary (%) | 0 (0) | 4 (100) | |
| Meningeal (%) | 2 (67) | 1 (33) | |
| Extremity (%) | 0 (0) | 1 (100) | |
| Head and Neck (%) | 0 (0) | 1 (100) | |
| Trunk (%) | 1 (100) | 0 (0) | |
| Mitotic counts/10 HPFs | 0.6199 | ||
| <4 (%) | 2 (33) | 4 (67) | |
| ≥4 (%) | 7 (58) | 5 (42) | |
| Necrosis | 0.5846 | ||
| Absent (%) | 6 (50) | 6 (50) | |
| Present (%) | 1 (25) | 3 (75) | |
| Fusion variant | 0.0152 | ||
| STAT6-TAD | 7 (88) | 1 (12) | |
| STAT6-FULL | 2 (20) | 8 (80) | |
| Recurrence/progression | 0.5594 | ||
| No (%) | 3 (75) | 1 (25) | |
| Yes (%) | 4 (40) | 6 (60) | |
| Outcome | 0.0699 | ||
| Alive (%) | 7 (70) | 3 (30) | |
| Died of disease (%) | 0 (0) | 4 (100) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salguero-Aranda, C.; Martínez-Reguera, P.; Marcilla, D.; de Álava, E.; Díaz-Martín, J. Evaluation of NAB2-STAT6 Fusion Variants and Other Molecular Alterations as Prognostic Biomarkers in a Case Series of 83 Solitary Fibrous Tumors. Cancers 2021, 13, 5237. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205237
Salguero-Aranda C, Martínez-Reguera P, Marcilla D, de Álava E, Díaz-Martín J. Evaluation of NAB2-STAT6 Fusion Variants and Other Molecular Alterations as Prognostic Biomarkers in a Case Series of 83 Solitary Fibrous Tumors. Cancers. 2021; 13(20):5237. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205237
Chicago/Turabian StyleSalguero-Aranda, Carmen, Paula Martínez-Reguera, David Marcilla, Enrique de Álava, and Juan Díaz-Martín. 2021. "Evaluation of NAB2-STAT6 Fusion Variants and Other Molecular Alterations as Prognostic Biomarkers in a Case Series of 83 Solitary Fibrous Tumors" Cancers 13, no. 20: 5237. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13205237