
DNA Methylation Analysis in Plasma Cell-Free DNA and Paired CTCs of NSCLC Patients before and after Osimertinib Treatment

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
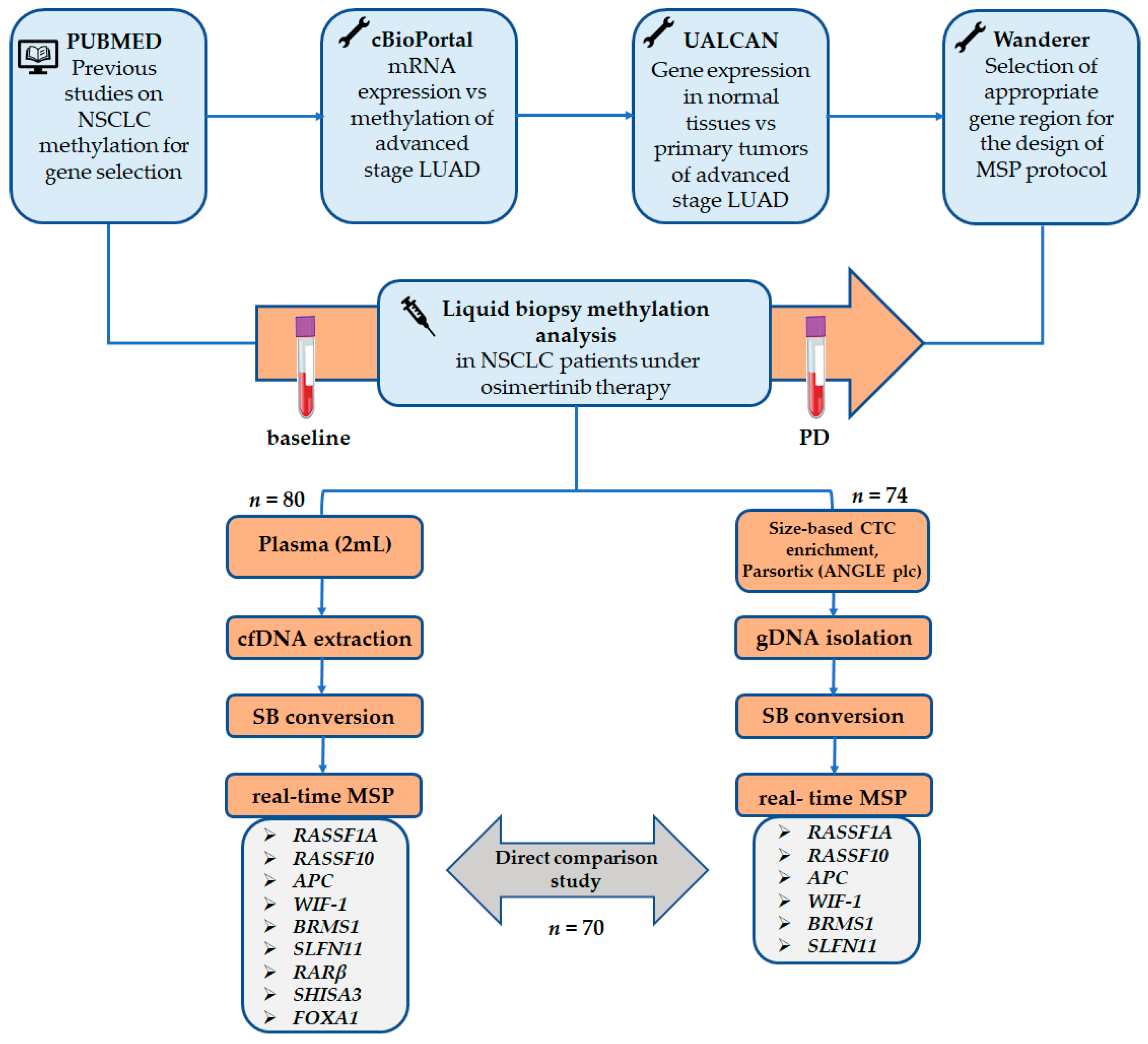
2. Materials and Methods
2.1. TCGA Meta-Analysis
2.2. Patients
2.3. Peripheral Blood Sampling and Processing
2.4. Plasma-cfDNA Extraction
2.5. CTC Enrichment and Genomic DNA Extraction
2.6. Evaluation of DNA Integrity
2.7. Sodium Bisulfite Treatment
2.8. In Silico Primer and Probe Design
2.9. Real-Time Methylation Specific PCR
2.10. Statistical Analysis
3. Results
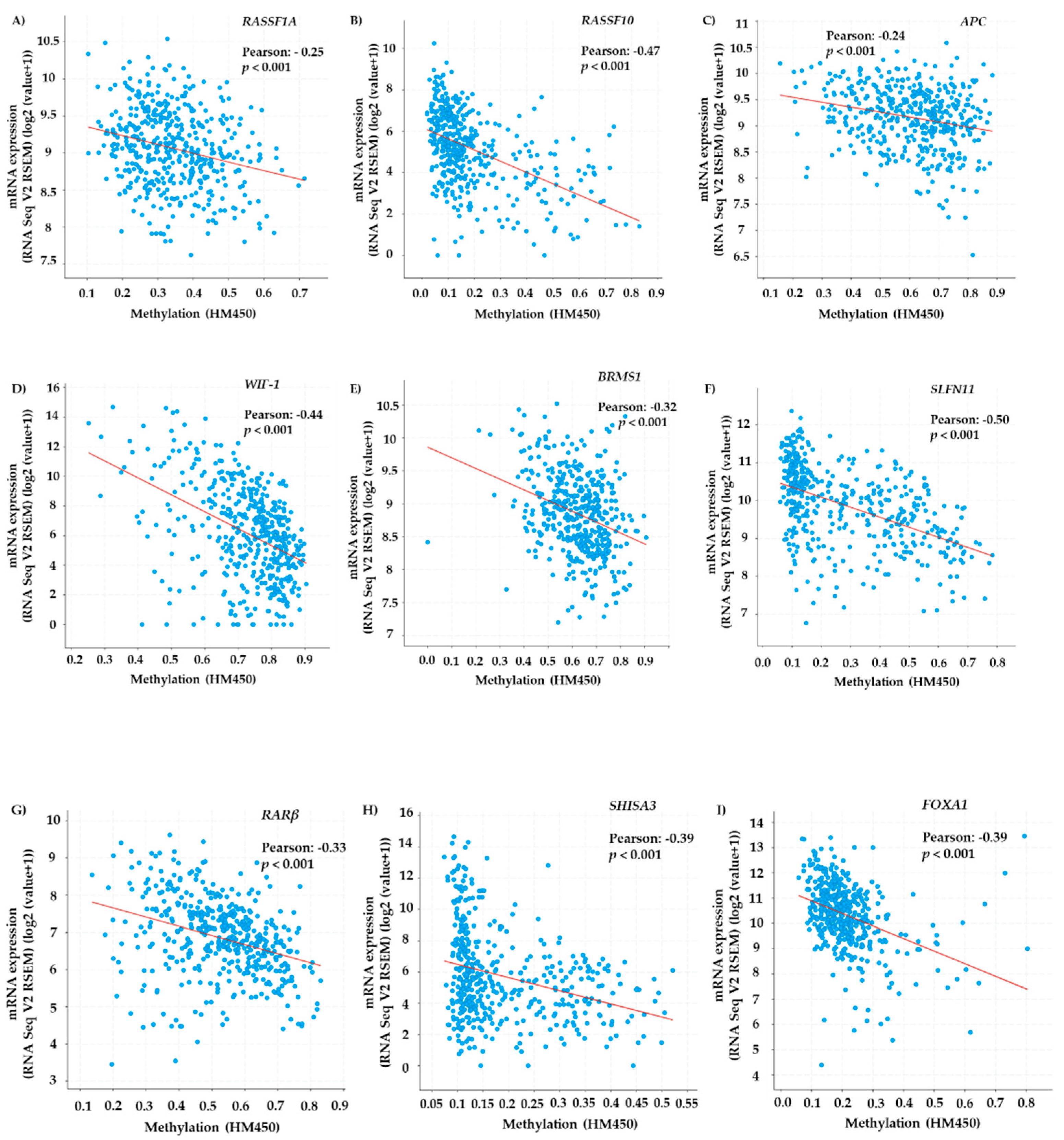
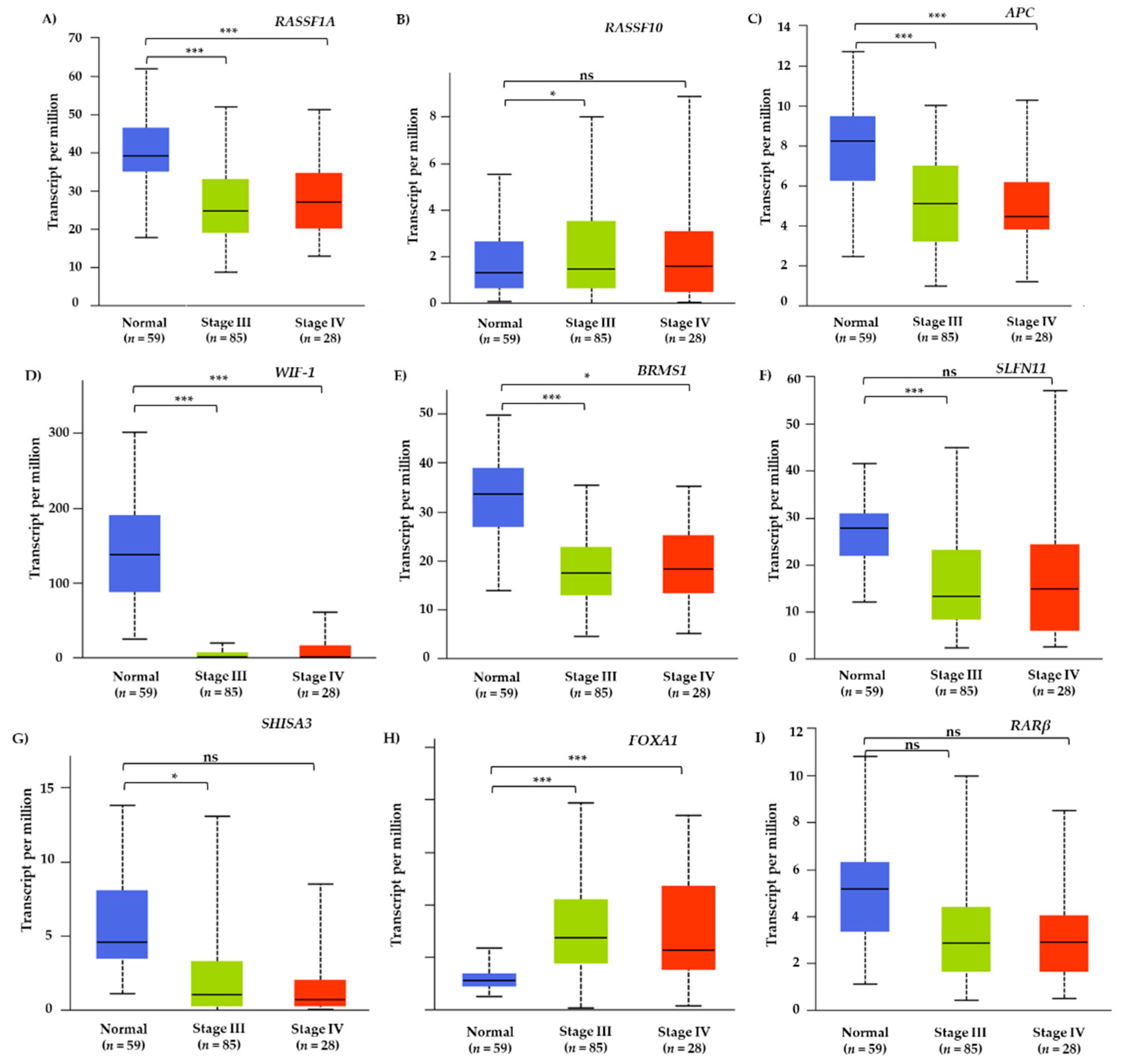
3.1. TCGA Meta-Analysis of Methylated Genes in Lung Adenocarcinoma Tissue Samples
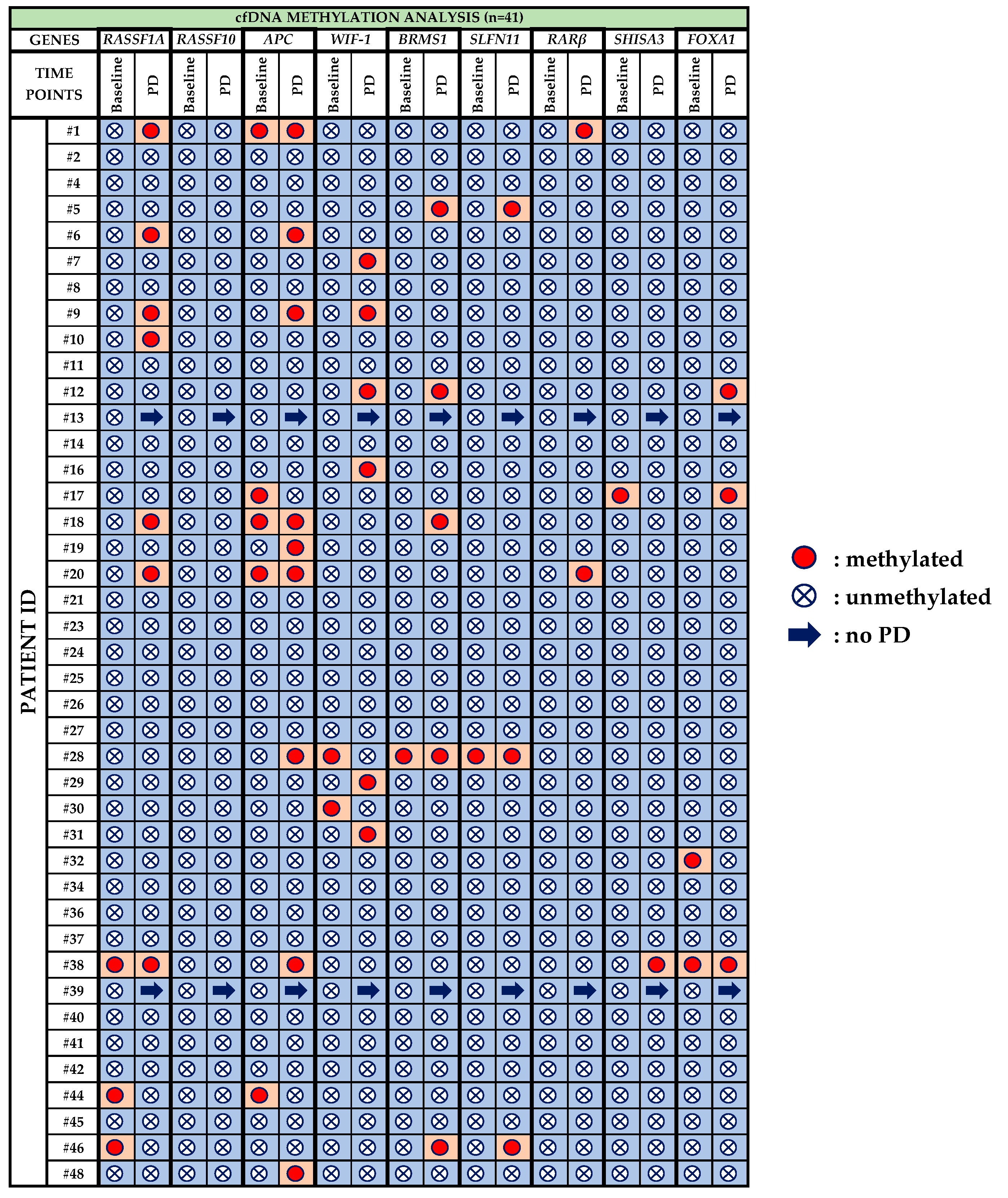
3.2. Plasma-cfDNA Methylation Analysis
- RASSF1A: Before treatment with osimertinib, methylation of RASSF1A was detected in 3/41 (7.3%) patients, whereas at PD it was observed in 7/39 (17.9%) patients. One patient (pt#38) maintained RASSF1A methylation at both time points, whereas pt#44 and pt#46 were negative at PD.
- RASSF10: Methylation for RASSF10 was not detected at any time point.
- APC: 5/41 (12.2%) patients were positive for APC methylation during baseline and three of these five (pt#1, pt#18, pt#20) maintained methylation also at PD. In total, 9/39 (23.1%) patients were detected with APC methylation at PD.
- WIF-1: WIF-1 methylation was detected in 2/41 (4.9%) patients at baseline and in 6/39 (15.4%) patients at PD.
- BRMS1: Before osimertinib treatment BRMS1 methylation was detected in only one patient (pt#28) who was also positive at PD. At PD, 5/39 (12.8%) patients were positive for BRMS1 methylation.
- SLFN11: Only one patient was detected with SLFN11 methylation at baseline while 3/39 (7.7%) patients were positive at PD.
- RARβ: Before treatment with osimertinib, no methylation was detected at all whereas 2/39 (5.1%) patients were positive at PD.
- SHISA3: SHISA3 methylation was detected in only one patient (pt#17) at baseline and in only one (pt#38) at PD.
- FOXA1: 2/41(4.9%) patients were detected positive for FOXA1 methylation at baseline whereas 3/39 (7.7%) patients at PD. Only one patient (pt#38) was positive for FOXA1 methylation at both time points.
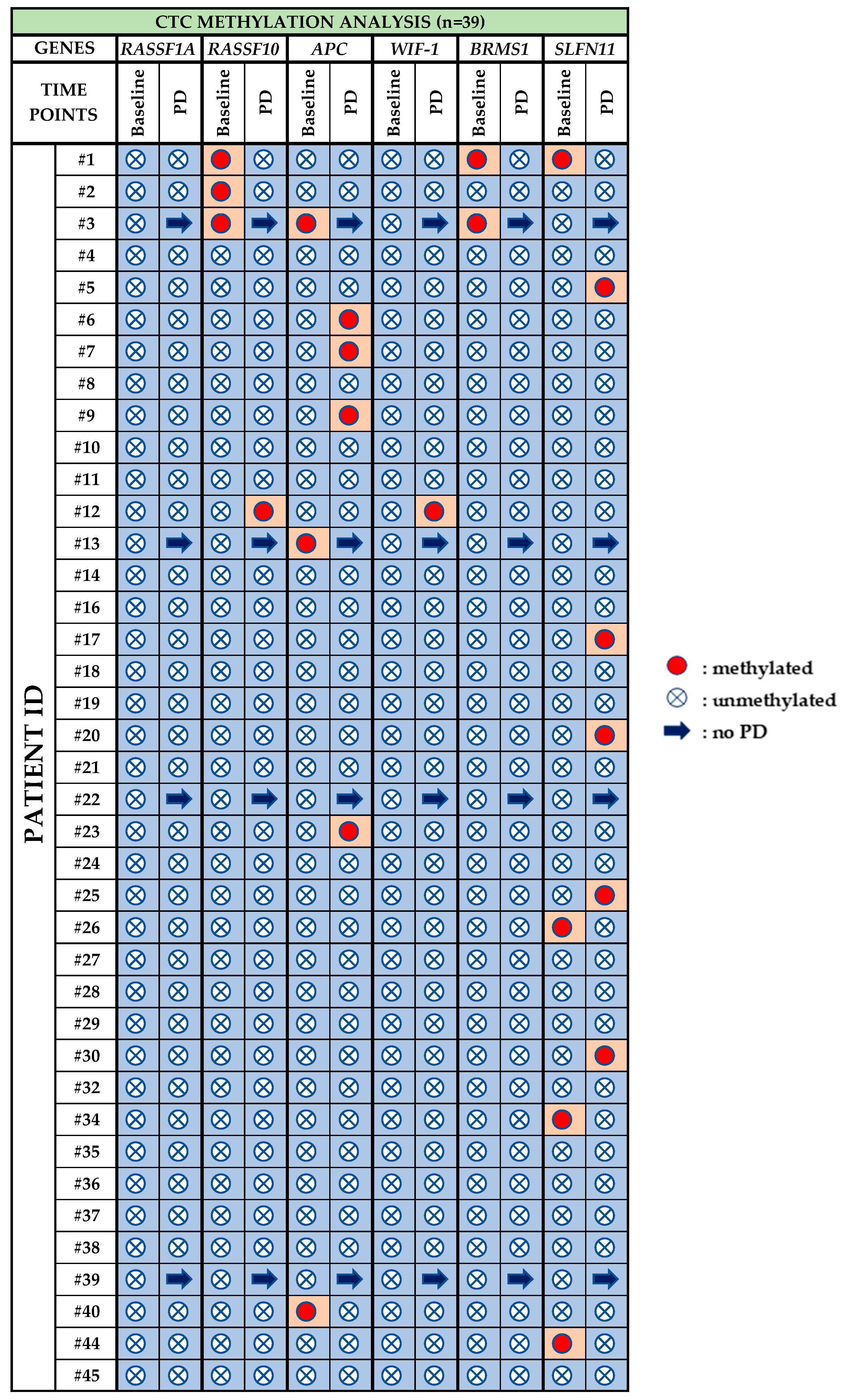
3.3. CTC Methylation Analysis
- RASSF1A: Methylation for RASSF1A was not detected at any time point.
- RASSF10: Before treatment with osimertinib, methylation of RASSF10 was detected in 3/39 (7.3%) patients, whereas at PD it was observed in 1/35 (2.8%) patient.
- APC: 3/39 (7.7%) patients were positive for APC methylation at baseline, while at PD, 4/35 (11.4%) were positive for APC methylation.
- WIF-1: WIF-1 methylation was not detected at baseline whereas at PD only one sample (pt#12) was found positive.
- BRMS1: Before osimertinib treatment BRMS1 methylation was detected in 2/39 (5.1%) patients, while at PD, no BRMS1 methylation was detected.
- SLFN11: At baseline, SLFN11 methylation was detected in 4/39 (10.2%) patients that were found negative at PD. At PD 5/35 (14.3%) patients were found positive for SLFN11 methylation.
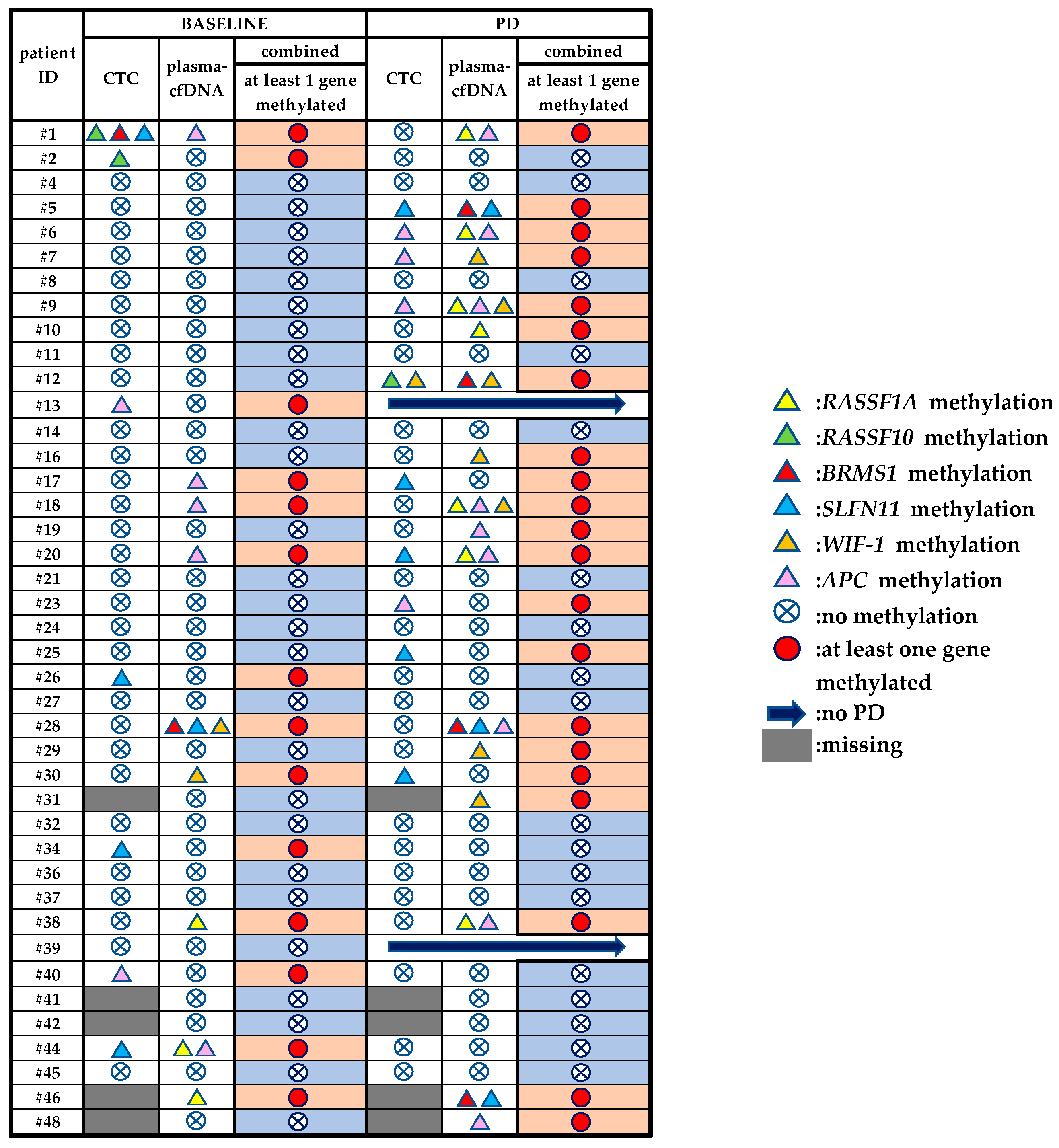
3.4. Direct Comparison of the Methylation Status of Six Genes between Plasma-cfDNA and Paired CTCs
3.5. DNA Methylation Analysis of Plasma-cfDNA and Paired CTCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mok, T.S.; Wu, Y.; Thongprasert, S.; Yang, C.; Saijo, N.; Sunpaweravong, P.; Han, B.; Margono, B.; Ichinose, Y.; Nishiwaki, Y.; et al. Gefitinib or Carboplatin–Paclitaxel in Pulmonary Adenocarcinoma. N. Engl. J. Med. 2009, 361, 947–957. [Google Scholar] [CrossRef]
- Maemondo, M.; Inoue, A.; Kobayashi, K.; Sugawara, S.; Oizumi, S.; Isobe, H.; Gemma, A.; Harada, M.; Yoshizawa, H.; Kinoshita, I.; et al. Gefitinib or chemotherapy for non-small-cell lung cancer with mutated EGFR. N. Engl. J. Med. 2010, 362, 2380–2388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosell, R.; Carcereny, E.; Gervais, R.; Vergnenegre, A.; Massuti, B.; Felip, E.; Palmero, R.; Garcia-Gomez, R.; Pallares, C.; Sanchez, J.M.; et al. Erlotinib versus standard chemotherapy as first-line treatment for European patients with advanced EGFR mutation-positive non-small-cell lung cancer (EURTAC): A multicentre, open-label, randomised phase 3 trial. Lancet Oncol. 2012, 13, 239–246. [Google Scholar] [CrossRef]
- Park, K.; Tan, E.H.; O’Byrne, K.; Zhang, L.; Boyer, M.; Mok, T.; Hirsh, V.; Yang, J.C.H.; Lee, K.H.; Lu, S.; et al. Afatinib versus gefitinib as first-line treatment of patients with EGFR mutation-positive non-small-cell lung cancer (LUX-Lung 7): A phase 2B, open-label, randomised controlled trial. Lancet Oncol. 2016, 17, 577–589. [Google Scholar] [CrossRef]
- Jänne, P.A.; Chih-Hsin Yang, J.; Kim, D.W.; Planchard, D.; Ohe, Y.; Ramalingam, S.S.; Ahn, M.J.; Kim, S.W.; Su, W.C.; Horn, L.; et al. AZD9291 in EGFR inhibitor-resistant non-small-cell lung cancer. N. Engl. J. Med. 2015, 372, 1689–1699. [Google Scholar] [CrossRef]
- Soria, J.C.; Ohe, Y.; Vansteenkiste, J.; Reungwetwattana, T.; Chewaskulyong, B.; Lee, K.H.; Dechaphunkul, A.; Imamura, F.; Nogami, N.; Kurata, T.; et al. Osimertinib in untreated EGFR-Mutated advanced non-small-cell lung cancer. N. Engl. J. Med. 2018, 378, 113–125. [Google Scholar] [CrossRef]
- Ramalingam, S.S.; Vansteenkiste, J.; Planchard, D.; Cho, B.C.; Gray, J.E.; Ohe, Y.; Zhou, C.; Reungwetwattana, T.; Cheng, Y.; Chewaskulyong, B.; et al. Overall Survival with Osimertinib in Untreated, EGFR-Mutated Advanced NSCLC. N. Engl. J. Med. 2020, 382, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Oxnard, G.R.; Hu, Y.; Mileham, K.F.; Husain, H.; Costa, D.B.; Tracy, P.; Feeney, N.; Sholl, L.M.; Dahlberg, S.E.; Redig, A.J.; et al. Assessment of Resistance Mechanisms and Clinical Implications in Patients with EGFR T790M-Positive Lung Cancer and Acquired Resistance to Osimertinib. JAMA Oncol. 2018, 4, 1527–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, X.; Puri, S.; Negrao, M.V.; Nilsson, M.B.; Boyle, T.; Hicks, J.K.; Lovinger, K.L.; Roarty, E.; Tang, M.; Sun, H.; et al. Landscape of EGFR -dependent and -independent resistance mechanisms to osimertinib and continuation therapy post- progression in EGFR-mutant NSCLC. Clin. Cancer Res. 2019, 24, 6195–6203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoni-Rugiu, E.; Melchior, L.C.; Urbanska, E.M.; Jakobsen, J.N.; De Stricker, K.; Grauslund, M.; Sørensen, J.B. Intrinsic resistance to EGFR-tyrosine kinase inhibitors in EGFR-mutant non-small cell lung cancer: Differences and similarities with acquired resistance. Cancers 2019, 11, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintanal-villalonga, Á.; Molina-pinelo, S. Epigenetics of lung cancer: A translational perspective. Cell Oncol. 2019, 42, 739–756. [Google Scholar] [CrossRef] [PubMed]
- Marsit, C.J.; Houseman, E.A.; Nelson, H.H.; Kelsey, K.T. Genetic and Epigenetic Tumor Suppressor Gene Silencing Are Distinct Molecular Phenotypes Driven by Growth Promoting Mutations in Nonsmall Cell Lung Cancer. J. Cancer Epidemiol. 2009, 2008, 215809. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.H.; Knudson, A.G.; Pandolfi, P.P. A continuum model for tumour suppression. Nature 2011, 476, 163–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, J.; Shackelford, R.E.; El-Osta, H. Epigenetics in non-small cell lung cancer: From basics to therapeutics. Transl. Lung Cancer Res. 2016, 5, 155–171. [Google Scholar] [CrossRef] [Green Version]
- Balgkouranidou, I.; Liloglou, T.; Lianidou, E.S. Lung cancer epigenetics: Emerging biomarkers. Biomark. Med. 2013, 7, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.V.; Lee, D.Y.; Li, B.; Quinlan, M.P.; Maheswaran, S.; Mcdermott, U.; Azizian, N.; Zou, L.; Fischbach, M.A.; Wong, K.; et al. A chromatin-mediated reversible drug-tolerant state in cancer cell subpopulations. Cell 2010, 141, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Lianidou, E.; Pantel, K. Liquid biopsies. Genes Chromosom. Cancer 2019, 58, 219–232. [Google Scholar] [CrossRef]
- Alix-Panabières, C.; Pantel, K. Liquid Biopsy: From Discovery to Clinical Application. Cancer Discov. 2021, 11, 858–873. [Google Scholar] [CrossRef]
- Palanca-ballester, C.; Rodriguez-casanova, A.; Torres, S.; Calabuig-fariñas, S.; Jantus-lewintre, E.; Diaz-lagares, A.; Montuenga, L.; Sandoval, J. Cancer Epigenetic Biomarkers in Liquid Biopsy for High Incidence Malignancies. Cancers 2021, 13, 3016. [Google Scholar] [CrossRef]
- Lianidou, E. Detection and relevance of epigenetic markers on ctDNA: Recent advances and future outlook. Mol. Oncol. 2021, 15, 1683–1700. [Google Scholar] [CrossRef]
- Zeng, C.; Stroup, E.K.; Zhang, Z.; Chiu, B.C.H.; Zhang, W. Towards precision medicine: Advances in 5-hydroxymethylcytosine cancer biomarker discovery in liquid biopsy. Cancer Commun. 2019, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lo, Y.M.D.; Han, D.S.C.; Jiang, P.C.R. Epigenetics, fragmentomics, and topology of cell-free DNA in liquid biopsies. Science 2021, 372, eaaw3616. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.C.; Oxnard, G.R.; Klein, E.A.; Swanton, C.S.M.C.C. Sensitive and specific multi-cancer detection and localization using methylation signatures in cell-free DNA. Ann. Oncol. 2020, 31, 745–759. [Google Scholar] [CrossRef]
- Nunes, S.P.; Diniz, F.; Moreira-Barbosa, C.; Constâncio, V.; Silva, A.V.; Oliveira, J.; Soares, M.; Paulino, S.; Cunha, A.L.; Rodrigues, J.; et al. Subtyping Lung Cancer Using DNA Methylation in Liquid Biopsies. J. Clin. Med. 2019, 8, 1500. [Google Scholar] [CrossRef] [Green Version]
- Chimonidou, M.; Strati, A.; Tzitzira, A.; Sotiropoulou, G.; Malamos, N.; Georgoulias, V.; Lianidou, E.S. DNA methylation of tumor suppressor and metastasis suppressor genes in circulating tumor cells. Clin. Chem. 2011, 57, 1169–1177. [Google Scholar] [CrossRef] [Green Version]
- Mastoraki, S.; Strati, A.; Tzanikou, E.; Chimonidou, M.; Politaki, E.; Voutsina, A.; Psyrri, A.; Georgoulias, V.; Lianidou, E. ESR1 methylation: A Liquid biopsy-based epigenetic assay for the follow up of patients with metastatic breast cancer receiving endocrine treatment: Short running title: Liquid biopsy: ESR1 methylation in CTCs and paired ctDNA. Clin. Cancer Res. 2018, 24, 1500–1511. [Google Scholar] [CrossRef] [Green Version]
- Vasantharajan, S.S.; Eccles, M.R.; Rodger, E.J.; Pattison, S.; McCall, J.L.; Gray, E.S.; Calapre, L.; Chatterjee, A. The Epigenetic landscape of Circulating tumour cells. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188514. [Google Scholar] [CrossRef]
- Lianidou, E.S. Gene expression profiling and DNA methylation analyses of CTCs. Mol. Oncol. 2016, 10, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Pixberg, C.F.; Raba, K.; Müller, F.; Behrens, B.; Honisch, E.; Niederacher, D.; Neubauer, H.; Fehm, T.; Goering, W.; Schulz, W.A.; et al. Analysis of DNA methylation in single circulating tumor cells. Oncogene 2017, 36, 3223–3231. [Google Scholar] [CrossRef] [PubMed]
- Gkountela, S.; Castro-giner, F.; Szczerba, B.M.; Rochlitz, C.; Weber, W.P.; Gkountela, S.; Castro-giner, F.; Szczerba, B.M.; Vetter, M.; Landin, J.; et al. Circulating Tumor Cell Clustering Shapes DNA Methylation to Enable Metastasis Seeding Article Circulating Tumor Cell Clustering Shapes DNA Methylation. Cell 2019, 176, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wu, X.; Zheng, J.; Dong, D. DNA methylome profiling of circulating tumor cells in lung cancer at single base-pair resolution. Oncogene 2021, 40, 1884–1895. [Google Scholar] [CrossRef]
- Malpeli, G.; Innamorati, G.; Decimo, I.; Bencivenga, M.; Kamdje, A.H.N.; Perris, R.; Bassi, C. Methylation dynamics of RASSF1A and its impact on cancer. Cancers 2019, 11, 959. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, T.; Bi, J.; Song, B.; Zhou, Y.; Zhang, C.; Gao, M. ScienceDirect RASSF10 is epigenetically inactivated and induces apoptosis in lung cancer cell lines. Biomed. Pharmacother. 2014, 68, 321–326. [Google Scholar] [CrossRef]
- Richter, A.M.; Walesch, S.K.; Wu, P. The tumor suppressor RASSF10 is upregulated upon contact inhibition and frequently epigenetically silenced in cancer. Oncogenesis 2012, 1, e18. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.J. Wnt Signaling Pathway in Non-Small Cell Lung Cancer. J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef]
- Virmani, A.K.; Rathi, A.; Sathyanarayana, U.G.; Padar, A.; Huang, C.X.; Cunnigham, H.T.; Farinas, A.J.; Sara, M.; Euhus, D.M.; Gilcrease, M.; et al. Aberrant methylation of the Adenomatous Polyposis Coli (APC) gene promoter 1A in breast and lung carcinomas. Clin. Cancer Res. 2001, 7, 1998–2004. [Google Scholar] [PubMed]
- Guo, S.; Tan, L.; Pu, W.; Wu, J.; Xu, K.; Wu, J.; Li, Q.; Ma, Y.; Xu, J.; Jin, L.; et al. Quantitative assessment of the diagnostic role of APC promoter methylation in non-small cell lung cancer. Clin. Epigenetics 2014, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chimonidou, M.; Kallergi, G.; Georgoulias, V.; Welch, D.R.; Lianidou, E.S. Breast Cancer Metastasis Suppressor-1 promoter methylation in primary breast tumors and corresponding Circulating Tumor Cells. Mol. Cancer Res. 2013, 11, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Nagji, A.S.; Liu, Y.; Stelow, E.B.; Stukenborg, G.J.; Jones, D.R. BRMS1 transcriptional repression correlates with CpG island methylation and advanced pathological stage in non-small cell lung cancer. J. Pathol. 2010, 221, 229–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balgkouranidou, I.; Chimonidou, M.; Milaki, G.; Tsarouxa, E.G.; Kakolyris, S.; Welch, D.R.; Georgoulias, V. Breast cancer metastasis suppressor-1 promoter methylation in cell-free DNA provides prognostic information in non-small cell lung cancer. Br. J. Cancer 2014, 110, 2054–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnaldi, E.; Ferraioli, D.; Ferrando, L.; Brohée, S.; Ferrando, F.; Fregatti, P. Schlafen-11 expression is associated with immune signatures and basal - like phenotype in breast cancer. Breast Cancer Res. Treat. 2019, 177, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Thomas, A.; Murai, J.; Trepel, J.B.; Bates, S.E.; Rajapakse, V.N.; Pommier, Y. Overcoming Resistance to DNA-Targeted Agents by Epigenetic Activation of Schlafen 11 ( SLFN11 ) Expression with Class I Histone Deacetylase Inhibitors. Clin. Cancer Res. 2018, 24, 1944–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogales, V.; Reinhold, W.C.; Varma, S.; Martinez-cardus, A.; Moutinho, C.; Moran, S.; Heyn, H.; Sebio, A.; Barnadas, A.; Pommier, Y.; et al. Epigenetic inactivation of the putative DNA / RNA helicase SLFN11 in human cancer confers resistance to platinum drugs. Oncotarget 2016, 7, 3084–3097. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Su, K.; Hong, Q.; Yan, B.; Chen, C.; Pan, S.; Chang, Y.; Wang, C.; Hung, P.; et al. Shisa3 Is Associated with Prolonged Survival through Promoting b -Catenin Degradation in Lung Cancer. Am. J. Respir. Crit. Care Med. 2014, 190, 433–444. [Google Scholar] [CrossRef]
- Si, J.; Ma, Y.; Bi, J.W.; Xiong, Y.; Lv, C.; Li, S.; Wu, N.; Yang, Y. Shisa3 brakes resistance to EGFR-TKIs in lung adenocarcinoma by suppressing cancer stem cell properties. J. Exp. Clin. Cancer Res. 2019, 38, 1–15. [Google Scholar] [CrossRef]
- Li, Y.; Lu, D.; Ma, Y.; Liu, H. Association between Retinoic acid receptor-β hypermethylation and NSCLC risk: A meta-analysis and literature review. Oncotarget 2017, 8, 5814–5822. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Zhang, Z.; Qing, X.; Wang, X.; Liang, C.; Liu, D. Promoter methylation of APC and RAR-β genes as prognostic markers in non-small cell lung cancer (NSCLC). Exp. Mol. Pathol. 2016, 100, 109–113. [Google Scholar] [CrossRef]
- Constâncio, V.; Nunes, S.P.; Moreira-Barbosa, C.; Freitas, R.; Oliveira, J.; Pousa, I.; Oliveira, J.; Soares, M.; Dias, C.G.; Dias, T.; et al. Early detection of the major male cancer types in blood-based liquid biopsies using a DNA methylation panel. Clin. Epigenetics 2019, 11, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ntzifa, A.; Strati, A.; Kallergi, G.; Kotsakis, A.; Georgoulias, V.; Lianidou, E. Gene expression in circulating tumor cells reveals a dynamic role of EMT and PD-L1 during osimertinib treatment in NSCLC patients. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef]
- Vorkas, P.A.; Poumpouridou, N.; Agelaki, S.; Kroupis, C.; Georgoulias, V.; Lianidou, E.S. PIK3CA Hotspot Mutation Scanning by a Novel and Highly Sensitive High-Resolution Small Amplicon Melting Analysis Method. J. Mol. Diagn. 2010, 12, 697–704. [Google Scholar] [CrossRef]
- Zavridou, M.; Mastoraki, S.; Strati, A.; Tzanikou, E.; Chimonidou, M.; Lianidou, E. Evaluation of preanalytical conditions and implementation of quality control steps for reliable gene expression and DNA methylation analyses in liquid biopsies. Clin. Chem. 2018, 64, 1522–1533. [Google Scholar] [CrossRef]
- Giannopoulou, L.; Chebouti, I.; Pavlakis, K.; Kasimir-bauer, S. RASSF1A promoter methylation in high-grade serous ovarian cancer: A direct comparison study in primary tumors, adjacent morphologically tumor cell-free tissues and paired circulating tumor DNA. Oncotarget 2017, 8, 21429–21443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavridou, M.; Strati, A.; Bournakis, E.; Smilkou, S.; Tserpeli, V.L.E. Prognostic Significance of Gene Expression and DNA Methylation Markers in Circulating Tumor Cells and Paired Plasma Derived Exosomes in Metastatic Castration Resistant Prostate Cancer. Cancers 2021, 13, 780. [Google Scholar] [CrossRef]
- Ntzifa, A.; Kotsakis, A.; Georgoulias, V.; Lianidou, E. Detection of EGFR mutations in plasma cfDNA and paired CTCs of NSCLC patients before and after osimertinib therapy using crystal digital PCR. Cancers 2021, 13, 2736. [Google Scholar] [CrossRef]
- Oxnard, G.R. The cellular origins of drug resistance in cancer. Nat. Publ. Gr. 2016, 22, 232–234. [Google Scholar] [CrossRef]
- Vaclova, T.; Grazini, U.; Ward, L.; O’Neill, D.; Markovets, A.; Huang, X.; Chmielecki, J.; Hartmaier, R.; Thress, K.S.; Smith, P.D.; et al. Clinical impact of subclonal EGFR T790M mutations in advanced-stage EGFR-mutant non-small-cell lung cancers. Nat. Commun. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Rotow, J.; Bivona, T.G. Understanding and targeting resistance mechanisms in NSCLC. Nat. Rev. Cancer 2017, 17, 637–658. [Google Scholar] [CrossRef] [PubMed]
- Romero-Garcia, S.; Prado-Garcia, H.; Carlos-Reyes, A. Role of DNA Methylation in the Resistance to Therapy in Solid Tumors. Front. Oncol. 2020, 10, 1–20. [Google Scholar] [CrossRef]
- Nagasawa, S.; Kashima, Y.; Suzuki, A.; Suzuki, Y. Single-cell and spatial analyses of cancer cells: Toward elucidating the molecular mechanisms of clonal evolution and drug resistance acquisition. Inflamm. Regen. 2021, 41, 22. [Google Scholar] [CrossRef] [PubMed]
- Kashima, Y.; Shibahara, D.; Suzuki, A.; Muto, K.; Kobayashi, I.S.; Plotnick, D.; Udagawa, H.; Izumi, H.; Shibata, Y.; Tanaka, K.; et al. Single-Cell Analyses Reveal Diverse Mechanisms of Resistance to EGFR Tyrosine Kinase Inhibitors in Lung Cancer. Cancer Res. 2021, 81, 4835–4848. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Murakami, Y.; Watari, K.; Kuwano, M.; Izumi, H.; Ono, M. CpG hypermethylation contributes to decreased expression of PTEN during acquired resistance to gefitinib in human lung cancer cell lines. Lung Cancer 2015, 87, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Wang, Y.; Duan, J.; Bai, H.; Wang, Z.; Wei, L.; Zhao, J.; Zhuo, M.; Wang, S.; Yang, L.; et al. DNA Methylation status of Wnt antagonist SFRP5 can predict the response to the EGFR-tyrosine kinase inhibitor therapy in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2012, 31, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Greve, G.; Schiffmann, I.; Pfeifer, D.; Pantic, M.; Schüler, J.; Lübbert, M. The pan-HDAC inhibitor panobinostat acts as a sensitizer for erlotinib activity in EGFR-mutated and -wildtype non-small cell lung cancer cells. BMC Cancer 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hou, T.; Ma, J.; Hu, C.; Zou, F.; Jiang, S.; Wang, Y.; Han, C.; Zhang, Y. Decitabine reverses gefitinib resistance in PC9 lung adenocarcinoma cells by demethylation of RASSF1A and GADD45β promoter. Int. J. Clin. Exp. Pathol. 2019, 12, 4002–4010. [Google Scholar] [PubMed]
- El Kadi, N.; Wang, L.; Davis, A.; Korkaya, H.; Cooke, A.; Vadnala, V.; Brown, N.A.; Betz, B.L.; Cascalho, M.; Kalemkerian, G.P.H.K. The EGFR T790M mutation is acquired through AICDA-mediated deamination of 5-methylcytosine following TKI treatment in lung cancer. Cancer Res. 2018, 78, 6728–6735. [Google Scholar] [CrossRef] [Green Version]
- Su, S.-F.; Liu, C.-H.; Cheng, C.-L.; Ho, C.-C.; Yang, T.-Y.; Chen, K.-C.; Hsu, K.-H.; Tseng, J.-S.; Chen, H.-W.; Chang, G.-C.; et al. Genome-Wide Epigenetic Landscape of Lung Adenocarcinoma Links HOXB9 DNA Methylation to Intrinsic EGFR-TKI Resistance and Heterogeneous Responses. JCO Precis. Oncol. 2021, 418–431. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Ye, J.; Chen, Y.; Lizaso, A.; Huang, L.; Shi, L.; Su, J.; Han-Zhang, H.; Chuai, S.; Li, L.; et al. Parallel serial assessment of somatic mutation and methylation profile from circulating tumor DNA predicts treatment response and impending disease progression in osimertinib-treated lung adenocarcinoma patients. Transl. Lung Cancer Res. 2019, 8, 1016–1028. [Google Scholar] [CrossRef]
- Nguyen, H.N.; Cao, N.P.T.; Van Nguyen, T.C.; Le, K.N.D.; Nguyen, D.T.; Nguyen, Q.T.T.; Nguyen, T.H.T.; Van Nguyen, C.; Le, H.T.; Nguyen, M.L.T.; et al. Liquid biopsy uncovers distinct patterns of DNA methylation and copy number changes in NSCLC patients with different EGFR-TKI resistant mutations. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
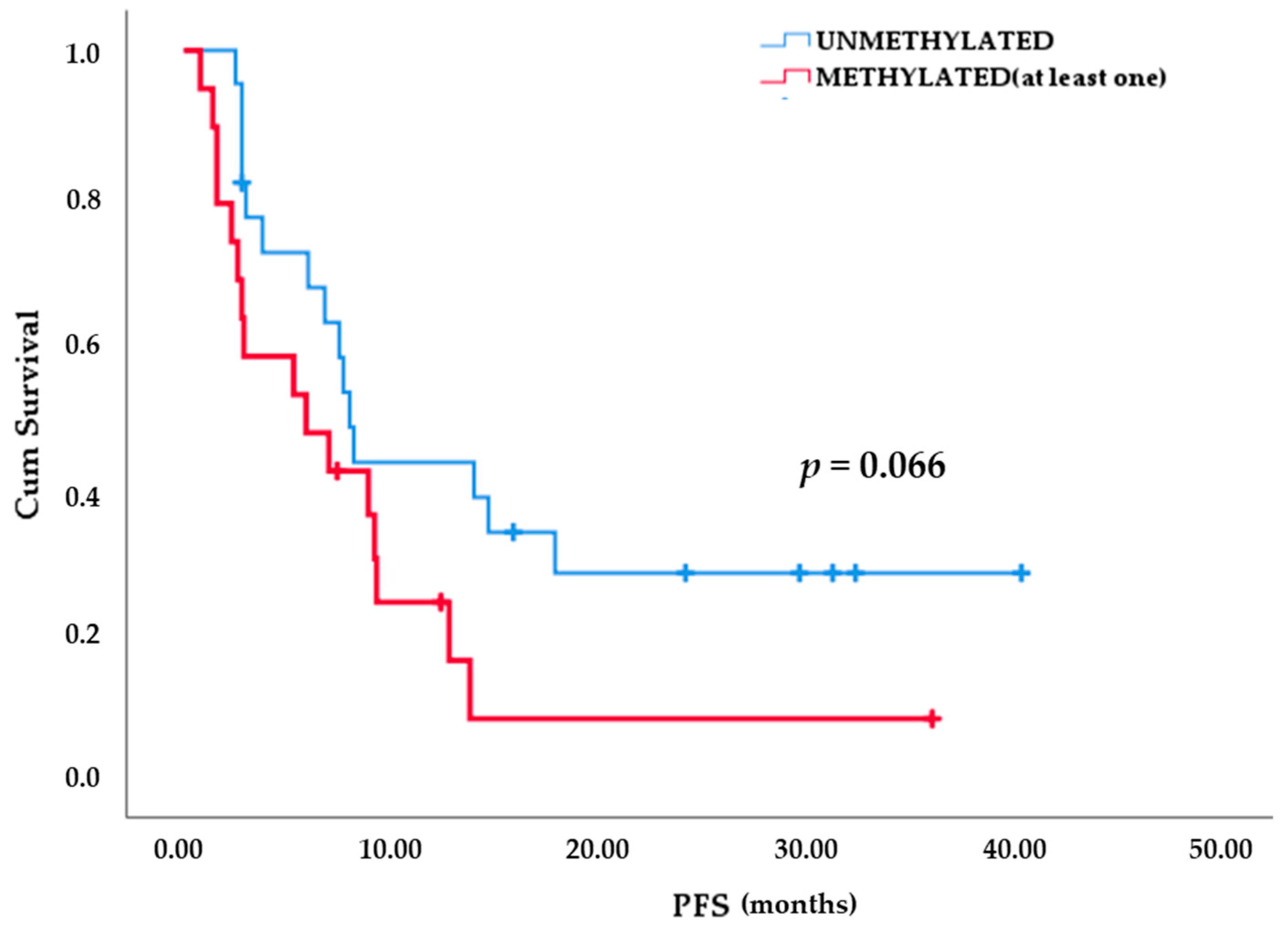
| Methylation Status | Early PD (Up to 13 Months) | Late PD (More Than 13 Months) | Total |
|---|---|---|---|
| Unmethylated | 14 | 8 | 22 |
| Methylated | 18 | 1 | 19 |
| (at least one gene) | |||
| Total | 32 | 9 | 41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ntzifa, A.; Londra, D.; Rampias, T.; Kotsakis, A.; Georgoulias, V.; Lianidou, E. DNA Methylation Analysis in Plasma Cell-Free DNA and Paired CTCs of NSCLC Patients before and after Osimertinib Treatment. Cancers 2021, 13, 5974. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13235974
Ntzifa A, Londra D, Rampias T, Kotsakis A, Georgoulias V, Lianidou E. DNA Methylation Analysis in Plasma Cell-Free DNA and Paired CTCs of NSCLC Patients before and after Osimertinib Treatment. Cancers. 2021; 13(23):5974. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13235974
Chicago/Turabian StyleNtzifa, Aliki, Dora Londra, Theodoros Rampias, Athanasios Kotsakis, Vassilis Georgoulias, and Evi Lianidou. 2021. "DNA Methylation Analysis in Plasma Cell-Free DNA and Paired CTCs of NSCLC Patients before and after Osimertinib Treatment" Cancers 13, no. 23: 5974. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers13235974