
Sphingolipids and Lymphomas: A Double-Edged Sword

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Sphingoid Bases
3. Sphingosine-1-Phosphate
4. Ceramide
5. Ceramide-1-Phosphate
6. Sphingomyelin
7. Glycosphingolipids
7.1. Cerebrosides
7.2. Globosides
7.3. Gangliosides
8. Targeting Sphingolipid Metabolism in Lymphoma Treatment
9. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Bultman, S.J. Interplay between diet, gut microbiota, epigenetic events, and colorectal cancer. Mol. Nutr. Food Res. 2017, 61, 1500902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bédard, A.-S.V.; Hien, E.D.; Lafontaine, D.A. Riboswitch regulation mechanisms: RNA, metabolites and regulatory proteins. Biochim. et Biophys. Acta 2020, 1863, 194501. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bermudez, J.; Baudrier, L.; Bayraktar, E.; Shen, Y.; La, K.; Guarecuco, R.; Yucel, B.; Fiore, D.; Tavora, B.; Freinkman, E.; et al. Squalene accumulation in cholesterol auxotrophic lymphomas prevents oxidative cell death. Nature 2019, 567, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Yan, X.J.; Rosen, L.; McCormick, S.; Chiorazzi, N.; Mongini, P.K.A. Effects of prostaglandin E2on p53 mRNA transcription and p53 mutagenesis during T-cell-independent human B-cell clonal expansion. FASEB J. 2014, 28, 627–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberlin, L.S.; Gabay, M.; Fan, A.C.; Gouw, A.M.; Tibshirani, R.J.; Felsher, D.W.; Zare, R.N. Alteration of the lipid profile in lymphomas induced by MYC overexpression. Proc. Natl. Acad. Sci. USA 2014, 111, 10450–10455. [Google Scholar] [CrossRef] [Green Version]
- Pernes, G.; Flynn, M.C.; Lancaster, G.I.; Murphy, A.J. Fat for fuel: Lipid metabolism in haematopoiesis. Clin. Transl. Immunol. 2019, 8, e1098. [Google Scholar] [CrossRef]
- Olson, D.; Fröhlich, F.; Farese, R.; Walther, T. Taming the sphinx: Mechanisms of cellular sphingolipid homeostasis. Biochim. et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1861, 784–792. [Google Scholar] [CrossRef] [Green Version]
- Bartke, N.; Hannun, Y.A. Bioactive sphingolipids: Metabolism and function. J. Lipid Res. 2009, 50, S91–S96. [Google Scholar] [CrossRef] [Green Version]
- Mollinedo, F.; Gajate, C. Lipid rafts as major platforms for signaling regulation in cancer. Adv. Biol. Regul. 2015, 57, 130–146. [Google Scholar] [CrossRef]
- Hakomori, S.-I.; Handa, K. GM3 and cancer. Glycoconj. J. 2015, 32, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hammadi, M.; Youinou, P.; Tempescul, A.; Tobón, G.; Berthou, C.; Bordron, A.; Pers, J.-O. Membrane microdomain sphingolipids are required for anti-CD20-induced death of chronic lymphocytic leukemia B cells. Haematologica 2012, 97, 288–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, D.; Saba, J.D. Sphingolipid Signaling and Hematopoietic Malignancies: To the Rheostat and Beyond. Anti-Cancer Agents Med. Chem. 2011, 11, 782–793. [Google Scholar] [CrossRef]
- Gault, C.R.; Obeid, L.M.; Hannun, Y.A. An overview of sphingolipid metabolism: From synthesis to breakdown. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2010. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Blom, T. Trafficking and Functions of Bioactive Sphingolipids: Lessons from Cells and Model Membranes. Lipid Insights 2015, 8, LPI-S31615. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Nagahashi, M.; Kim, E.Y.; Harikumar, K.B.; Yamada, A.; Huang, W.-C.; Hait, N.C.; Allegood, J.C.; Price, M.M.; Avni, D.; et al. Sphingosine-1-Phosphate Links Persistent STAT3 Activation, Chronic Intestinal Inflammation, and Development of Colitis-Associated Cancer. Cancer Cell 2013, 23, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Vockerodt, M.; Vrzalikova, K.; Ibrahim, M.; Nagy, E.; Margielewska, S.; Hollows, R.; Lupino, L.; Tooze, R.; Care, M.; Simmons, W.; et al. Regulation of S1PR2 by the EBV oncogene LMP1 in aggressive ABC-subtype diffuse large B-cell lymphoma. J. Pathol. 2019, 248, 142–154. [Google Scholar] [CrossRef]
- Patwardhan, G.A.; Liu, Y.-Y. Sphingolipids and expression regulation of genes in cancer. Prog. Lipid Res. 2011, 50, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, H.; Chen, T.; Wang, H.; Liang, X.; Zhang, Y.; Duan, J.; Qian, S.; Qiao, K.; Zhang, L.; et al. C24-Ceramide Drives Gallbladder Cancer Progression Through Directly Targeting Phosphatidylinositol 5-Phosphate 4-Kinase Type-2 Gamma to Facilitate Mammalian Target of Rapamycin Signaling Activation. Hepatology 2021, 73, 692–712. [Google Scholar] [CrossRef]
- Hartmann, D.; Lucks, J.; Fuchs, S.; Schiffmann, S.; Schreiber, Y.; Ferreirós, N.; Merkens, J.; Marschalek, R.; Geisslinger, G.; Grösch, S. Long chain ceramides and very long chain ceramides have opposite effects on human breast and colon cancer cell growth. Int. J. Biochem. Cell Biol. 2012, 44, 620–628. [Google Scholar] [CrossRef]
- Snider, A.J.; Gandy, K.A.O.; Obeid, L.M. Sphingosine kinase: Role in regulation of bioactive sphingolipid mediators in inflammation. Biochimie 2010, 92, 707–715. [Google Scholar] [CrossRef] [Green Version]
- Separovic, D.; Semaan, L.; Tarca, A.L.; Maitah, M.Y.A.; Hanada, K.; Bielawski, J.; Villani, M.; Luberto, C. Suppression of sphingomyelin synthase 1 by small interference RNA is associated with enhanced ceramide production and apoptosis after photodamage. Exp. Cell Res. 2008, 314, 1860–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holm, L.J.; Krogvold, L.; Hasselby, J.P.; Kaur, S.; Claessens, L.A.; Russell, M.A.; Mathews, C.E.; Hanssen, K.F.; Morgan, N.; Koeleman, B.P.C.; et al. Abnormal islet sphingolipid metabolism in type 1 diabetes. Diabetologia 2018, 61, 1650–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sui, J.; He, M.; Wang, Y.; Zhao, X.; He, Y.; Shi, B. Sphingolipid metabolism in type 2 diabetes and associated cardiovascular complications. Exp. Ther. Med. 2019, 18, 3603–3614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mielke, M.M.; Haughey, N.J.; Han, D.; An, Y.; Bandaru, V.V.R.; Lyketsos, C.G.; Ferrucci, L.; Resnick, S.M. The Association between Plasma Ceramides and Sphingomyelins and Risk of Alzheimer’s Disease Differs by Sex and APOE in the Baltimore Longitudinal Study of Aging. J. Alzheimer’s Dis. 2017, 60, 819–828. [Google Scholar] [CrossRef] [Green Version]
- WHO. Globocan 2020. Available online: https://gco.iarc.fr/today/home (accessed on 15 November 2021).
- Artz, A.S.; Ershler, W.B. Management of the older patient. In Hematopoietic Stem Cell Transplantation in Clinical Practice; Elsevier Health Sciences: Amsterdam, The Netherlands, 2009; ISBN 9780443101472. [Google Scholar]
- Singh, R.; Shaik, S.; Negi, B.; Rajguru, J.; Patil, P.; Parihar, A.; Sharma, U. Non-Hodgkin’s lymphoma: A review. J. Fam. Med. Prim. Care 2020, 9, 1834–1840. [Google Scholar] [CrossRef]
- Solimando, A.G.; Ribatti, D.; Vacca, A.; Einsele, H. Targeting B-cell non Hodgkin lymphoma: New and old tricks. Leuk. Res. 2016, 42, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Sukswai, N.; Lyapichev, K.; Khoury, J.D.; Medeiros, L.J. Diffuse large B-cell lymphoma variants: An update. Pathology 2020, 52, 53–67. [Google Scholar] [CrossRef] [Green Version]
- de Leval, L.; Jaffe, E.S. Lymphoma Classification. Cancer J. 2020, 26, 176–185. [Google Scholar] [CrossRef]
- Ysebaert, L.; Quillet-Mary, A.; Tosolini, M.; Pont, F.; Laurent, C.; Fournié, J.-J. Lymphoma Heterogeneity Unraveled by Single-Cell Transcriptomics. Front. Immunol. 2021, 12, 202. [Google Scholar] [CrossRef]
- Wright, G.W.; Huang, D.W.; Phelan, J.D.; Coulibaly, Z.A.; Roulland, S.; Young, R.M.; Wang, J.Q.; Schmitz, R.; Morin, R.; Tang, J.; et al. A Probabilistic Classification Tool for Genetic Subtypes of Diffuse Large B Cell Lymphoma with Therapeutic Implications. Cancer Cell 2020, 37, 551–568.e14. [Google Scholar] [CrossRef] [PubMed]
- Esmeray, E.; Küçük, C. Genetic alterations in B cell lymphoma subtypes as potential biomarkers for non-invasive diagnosis, prognosis, therapy, and disease monitoring. Turk. J. Biol. 2020, 44, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Grubor, V.; Love, C.L.; Banerjee, A.; Richards, K.L.; Mieczkowski, P.A.; Dunphy, C.; Choi, W.; Au, W.Y.; Srivastava, G.; et al. Genetic heterogeneity of diffuse large B-cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1398–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maan, M.; Peters, J.; Dutta, M.; Patterson, A.D. Lipid metabolism and lipophagy in cancer. Biochem. Biophys. Res. Commun. 2018, 504, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Corn, K.C.; Windham, M.A.; Rafat, M. Lipids in the tumor microenvironment: From cancer progression to treatment. Prog. Lipid Res. 2020, 80, 101055. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Liang, J.-H.; Wang, L.; Zhu, H.-Y.; Wu, W.; Cao, L.; Fan, L.; Li, J.-Y.; Yang, T.; Xu, W. Low serum cholesterol levels predict inferior prognosis and improve NCCN-IPI scoring in diffuse large B cell lymphoma. Int. J. Cancer 2018, 143, 1884–1895. [Google Scholar] [CrossRef]
- Kleinstern, G.; Camp, N.J.; Berndt, S.I.; Birmann, B.M.; Nieters, A.; Bracci, P.M.; McKay, J.D.; Ghesquières, H.; Lan, Q.; Hjalgrim, H.; et al. Lipid Trait Variants and the Risk of Non-Hodgkin Lymphoma Subtypes: A Mendelian Randomization Study. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1074–1078. [Google Scholar] [CrossRef] [Green Version]
- Cinque, B.; Di Marzio, L.; Centi, C.; Di Rocco, C.; Riccardi, C.; Cifone, M.G. Sphingolipids and the immune system. Pharmacol. Res. 2003, 47, 421–437. [Google Scholar] [CrossRef]
- Bode, C.; Gräler, M.H. Immune Regulation by Sphingosine 1-Phosphate and Its Receptors. Arch. Immunol. et Ther. Exp. 2012, 60, 3–12. [Google Scholar] [CrossRef]
- Baeyens, A.A.; Schwab, S.R. Finding a Way Out: S1P Signaling and Immune Cell Migration. Annu. Rev. Immunol. 2020, 38, 759–784. [Google Scholar] [CrossRef]
- Raza, Y.; Salman, H.; Luberto, C. Sphingolipids in Hematopoiesis: Exploring Their Role in Lineage Commitment. Cells 2021, 10, 2507. [Google Scholar] [CrossRef]
- Zhang, T.; de Waard, A.; Wuhrer, M.; Spaapen, R.M. The Role of Glycosphingolipids in Immune Cell Functions. Front. Immunol. 2019, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Saito, M. Bioactive gangliosides: Differentiation inducers for hematopoietic cells and their mechanism(s) of actions. Adv. Lipid Res. 1993, 25, 303–327. [Google Scholar] [PubMed]
- Kaucic, K.; Grovas, A.; Li, R.; Quinones, R.; Ladisch, S. Modulation of human myelopoiesis by human gangliosides. Exp. Hematol. 1994, 22, 52–59. [Google Scholar] [PubMed]
- Katayama, Y.; Frenette, P.S. Galactocerebrosides Are Required Postnatally for Stromal-Dependent Bone Marrow Lymphopoiesis. Immunity 2003, 18, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Bharti, A.C.; Singh, S.M. Induction of apoptosis in bone marrow cells by gangliosides produced by a T cell lymphoma. Immunol. Lett. 2000, 72, 39–48. [Google Scholar] [CrossRef]
- Tang, Y.-C.; Yuwen, H.; Wang, K.; Bruno, P.M.; Bullock, K.; Deik, A.; Santaguida, S.; Trakala, M.; Pfau, S.J.; Zhong, N.; et al. Aneuploid Cell Survival Relies upon Sphingolipid Homeostasis. Cancer Res. 2017, 77, 5272–5286. [Google Scholar] [CrossRef] [Green Version]
- Kilbey, A.; Terry, A.; Jenkins, A.; Borland, G.; Zhang, Q.; Wakelam, M.; Cameron, E.R.; Neil, J.C. Runx Regulation of Sphingolipid Metabolism and Survival Signaling. Cancer Res. 2010, 70, 5860–5869. [Google Scholar] [CrossRef] [Green Version]
- Lone, W.; Bouska, A.; Sharma, S.; Amador, C.; Saumyaranjan, M.; Herek, T.A.; Heavican, T.B.; Yu, J.; Lim, S.T.; Ong, C.K.; et al. Genome-Wide miRNA Expression Profiling of Molecular Subgroups of Peripheral T-cell Lymphoma. Clin. Cancer Res. 2021, 27, 6039–6053. [Google Scholar] [CrossRef]
- Piszcz, J.; Lemancewicz, D.; Dudzik, D.; Ciborowski, M. Differences and similarities between LC-MS derived serum fingerprints of patients with B-cell malignancies. Electrophoresis 2013, 34, 2857-2664. [Google Scholar] [CrossRef]
- Barré, F.P.; Claes, B.S.; Dewez, F.; Peutz-Kootstra, C.J.; Munch-Petersen, H.F.; Grønbæk, K.; Lund, A.H.; Heeren, R.M.; Côme, C.R.M.; Cillero-Pastor, B. Specific Lipid and Metabolic Profiles of R-CHOP-Resistant Diffuse Large B-Cell Lymphoma Elucidated by Matrix-Assisted Laser Desorption Ionization Mass Spectrometry Imaging and in Vivo Imaging. Anal. Chem. 2018, 90, 14198–14206. [Google Scholar] [CrossRef] [Green Version]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nat. Cancer 2017, 18, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Farhat, M.; Poissonnier, A.; Hamze, A.; Ouk-Martin, C.; Brion, J.-D.; Alami, M.; Feuillard, J.; Jayatvignoles, C. Reversion of apoptotic resistance of TP53-mutated Burkitt lymphoma B-cells to spindle poisons by exogenous activation of JNK and p38 MAP kinases. Cell Death Dis. 2014, 5, e1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.M.; Makena, M.R.; Hindle, A.; Koneru, B.; Nguyen, T.H.; Verlekar, D.U.; Cho, H.; Maurer, B.J.; Kang, M.H.; Reynolds, C.P. Cytotoxicity and molecular activity of fenretinide and metabolites in T-cell lymphoid malignancy, neuroblastoma, and ovarian cancer cell lines in physiological hypoxia. Anti-Cancer Drugs 2019, 30, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.H.; Chang, C.-C.; Schroeder, J.J. Evaluation of Sphinganine and Sphingosine as Human Breast Cancer Chemotherapeutic and Chemopreventive Agents. Exp. Biol. Med. 2006, 231, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.H.; Yang, H.; Hsieh, C.-Y.; Sun, W.; Chang, C.-C.; Schroeder, J.J. Evaluation of chemotherapeutic and cancer-protective properties of sphingosine and C2-ceramide in a human breast stem cell derived carcinogenesis model. Int. J. Oncol. 2019, 54, 655–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, C.; Berlin, M.; Röstel, F.; Teichmann, B.; Gräler, M.H. Evaluating Sphingosine and its Analogues as Potential Alternatives for Aggressive Lymphoma Treatment. Cell. Physiol. Biochem. 2014, 34, 1686–1700. [Google Scholar] [CrossRef] [Green Version]
- Park, M.-T.; Kang, J.A.; Choi, J.-A.; Kang, C.-M.; Kim, T.-H.; Bae, S.; Kang, S.; Kim, S.; Choi, W.-I.; Cho, C.-K.; et al. Phytosphingosine induces apoptotic cell death via caspase 8 activation and Bax translocation in human cancer cells. Clin. Cancer Res. 2003, 9, 878–885. [Google Scholar]
- Yuza, K.; Nakajima, M.; Nagahashi, M.; Tsuchida, J.; Hirose, Y.; Miura, K.; Tajima, Y.; Abe, M.; Sakimura, K.; Takabe, K.; et al. Different Roles of Sphingosine Kinase 1 and 2 in Pancreatic Cancer Progression. J. Surg. Res. 2018, 232, 186–194. [Google Scholar] [CrossRef]
- Tsuchida, J.; Nagahashi, M.; Nakajima, M.; Moro, K.; Tatsuda, K.; Ramanathan, R.; Takabe, K.; Wakai, T. Breast cancer sphingosine-1-phosphate is associated with phospho-sphingosine kinase 1 and lymphatic metastasis. J. Surg. Res. 2016, 205, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Hirose, Y.; Nagahashi, M.; Katsuta, E.; Yuza, K.; Miura, K.; Sakata, J.; Kobayashi, T.; Ichikawa, H.; Shimada, Y.; Kameyama, H.; et al. Generation of sphingosine-1-phosphate is enhanced in biliary tract cancer patients and is associated with lymphatic metastasis. Sci. Rep. 2018, 8, 10814. [Google Scholar] [CrossRef] [Green Version]
- Nunes, J.; Naymark, M.; Sauer, L.; Muhammad, A.; Keun, H.; Sturge, J.; Stebbing, J.; Waxman, J.; Pchejetski, D. Circulating sphingosine-1-phosphate and erythrocyte sphingosine kinase-1 activity as novel biomarkers for early prostate cancer detection. Br. J. Cancer 2012, 106, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Yokota, T.; Nojima, H.; Kuboki, S.; Yoshitomi, H.; Furukawa, K.; Takayashiki, T.; Takano, S.; Ohtsuka, M. Sphingosine-1-phosphate Receptor-1 Promotes Vascular Invasion and EMT in Hepatocellular Carcinoma. J. Surg. Res. 2021, 259, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Cheng, Y.; Chang, H.-M.; Deguchi, M.; Hsueh, A.J.; Leung, P.C.K. Sphingosine-1-phosphate promotes ovarian cancer cell proliferation by disrupting Hippo signaling. Oncotarget 2017, 8, 27166–27176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, M.; Nagahashi, M.; Rashid, O.; Takabe, K.; Wakai, T. The role of sphingosine-1-phosphate in the tumor microenvironment and its clinical implications. Tumor Biol. 2017, 39, 1010428317699133. [Google Scholar] [CrossRef] [Green Version]
- Riboni, L.; Hadi, L.A.; Navone, S.E.; Guarnaccia, L.; Campanella, R.; Marfia, G. Sphingosine-1-Phosphate in the Tumor Microenvironment: A Signaling Hub Regulating Cancer Hallmarks. Cells 2020, 9, 337. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, Y.I.; Campos, L.E.; Castro, M.G.; Aladhami, A.; Oskeritzian, C.A.; Alvarez, S.E. Sphingosine-1 Phosphate: A New Modulator of Immune Plasticity in the Tumor Microenvironment. Front. Oncol. 2016, 6, 218. [Google Scholar] [CrossRef] [Green Version]
- Weichand, B.; Popp, R.; Dziumbla, S.; Mora, J.; Strack, E.; Elwakeel, E.; Frank, A.-C.; Scholich, K.; Pierre, S.; Syed, S.N.; et al. S1PR1 on tumor-associated macrophages promotes lymphangiogenesis and metastasis via NLRP3/IL-1β. J. Exp. Med. 2017, 214, 2695–2713. [Google Scholar] [CrossRef]
- El Buri, A.; Adams, D.R.; Smith, D.; Tate, R.; Mullin, M.; Pyne, S.; Pyne, N.J. The sphingosine 1-phosphate receptor 2 is shed in exosomes from breast cancer cells and is N-terminally processed to a short constitutively active form that promotes extracellular signal regulated kinase activation and DNA synthesis in fibroblasts. Oncotarget 2018, 9, 29453–29467. [Google Scholar] [CrossRef] [Green Version]
- Albinet, V.; Bats, M.-L.; Huwiler, A.; Rochaix, P.; Chevreau, C.; Ségui, B.; Levade, T.; Andrieu-Abadie, N. Dual role of sphingosine kinase-1 in promoting the differentiation of dermal fibroblasts and the dissemination of melanoma cells. Oncogene 2014, 33, 3364–3373. [Google Scholar] [CrossRef]
- Yester, J.W.; Bryan, L.; Waters, M.R.; Mierzenski, B.; Biswas, D.D.; Gupta, A.S.; Bhardwaj, R.; Surace, M.J.; Eltit, J.M.; Milstien, S.; et al. Sphingosine-1-phosphate inhibits IL-1-induced expression of C-C motif ligand 5 via c-Fos-dependent suppression of IFN-β amplification loop. FASEB J. 2015, 29, 4853–4865. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Huang, P.; Dai, H.; Li, Q.; Hu, L.; Peng, J.; Jiang, S.; Xu, Y.; Wu, Z.; Nie, H.; et al. TIMELESS regulates sphingolipid metabolism and tumor cell growth through Sp1/ACER2/S1P axis in ER-positive breast cancer. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, Y.; Li, J.; Ji, B.; Kang, N.; Yang, L.; Simonetto, D.A.; Kwon, J.H.; Kamath, M.; Cao, S.; Shah, V. Sphingosine-1-Phosphate Mediates a Reciprocal Signaling Pathway between Stellate Cells and Cancer Cells that Promotes Pancreatic Cancer Growth. Am. J. Pathol. 2014, 184, 2791–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Zhao, S.; Wang, S.; Pan, X.; Zhang, Y.; Xu, J.; Jiang, Y.; Li, H.; Zhang, Q.; Gao, J.; et al. S1P/S1PR3 axis promotes aerobic glycolysis by YAP/c-MYC/PGAM1 axis in osteosarcoma. EBioMedicine 2019, 40, 210–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, P.; Kim, D.; You, E.; Jung, J.; Oh, S.; Kim, J.; Lee, K.-H.; Rhee, S. Extracellular Matrix Rigidity-dependent Sphingosine-1-phosphate Secretion Regulates Metastatic Cancer Cell Invasion and Adhesion. Sci. Rep. 2016, 6, 21564. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Liu, X.; Yan, Z.; Xie, L. Sphingosine 1-phosphate regulates proliferation, cell cycle and apoptosis of hepatocellular carcinoma cells via syndecan-1. Prog. Biophys. Mol. Biol. 2019, 148, 32–38. [Google Scholar] [CrossRef]
- Sassoli, C.; Pierucci, F.; Tani, A.; Frati, A.; Chellini, F.; Matteini, F.; Vestri, A.; Anderloni, G.; Nosi, D.; Zecchi-Orlandini, S.; et al. Sphingosine 1-Phosphate Receptor 1 Is Required for MMP-2 Function in Bone Marrow Mesenchymal Stromal Cells: Implications for Cytoskeleton Assembly and Proliferation. Stem Cells Int. 2018, 2018, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Young, N.; Pearl, D.K.; Van Brocklyn, J.R. Sphingosine-1-Phosphate Regulates Glioblastoma Cell Invasiveness through the Urokinase Plasminogen Activator System and CCN1/Cyr61. Mol. Cancer Res. 2009, 7, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-Y.; Choi, B.; Kim, J.-E.; Park, S.-O.; Kim, S.-M.; Chang, E.-J. Interleukin-22 Mediates the Chemotactic Migration of Breast Cancer Cells and Macrophage Infiltration of the Bone Microenvironment by Potentiating S1P/SIPR Signaling. Cells 2020, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hind, T.; Lam, B.W.S.; Herr, D.R. Sphingosine 1–phosphate signaling induces SNAI2 expression to promote cell invasion in breast cancer cells. FASEB J. 2019, 33, 7180–7191. [Google Scholar] [CrossRef]
- Cattaneo, M.G.; Vanetti, C.; Samarani, M.; Aureli, M.; Bassi, R.; Sonnino, S.; Giussani, P. Cross-talk between sphingosine-1-phosphate and EGFR signaling pathways enhances human glioblastoma cell invasiveness. FEBS Lett. 2018, 592, 949–961. [Google Scholar] [CrossRef] [Green Version]
- Adada, M.M.; Canals, D.; Jeong, N.; Kelkar, A.D.; Hernandez-Corbacho, M.; Pulkoski-Gross, M.J.; Donaldson, J.C.; Hannun, Y.A.; Obeid, L.M. Intracellular sphingosine kinase 2-derived sphingosine-1-phosphate mediates epidermal growth factor-induced ezrin-radixin-moesin phosphorylation and cancer cell invasion. FASEB J. 2015, 29, 4654–4669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muehlich, S. Induction of connective tissue growth factor (CTGF) in human endothelial cells by lysophosphatidic acid, sphingosine-1-phosphate, and platelets. Atherosclerosis 2004, 175, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Ségui, B.; Andrieu-Abadie, N.; Jaffrézou, J.-P.; Benoist, H.; Levade, T. Sphingolipids as modulators of cancer cell death: Potential therapeutic targets. Biochim. et Biophys. Acta (BBA)-Biomembr. 2006, 1758, 2104–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, S.; Keino-Masu, K.; Ohto, T.; Masu, M. The N-terminal hydrophobic sequence of autotaxin (ENPP2) functions as a signal peptide. Genes Cells 2006, 11, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Härmä, V.; Knuuttila, M.; Virtanen, J.; Mirtti, T.; Kohonen, P.; Kovanen, P.; Happonen, A.; Kaewphan, S.; Ahonen, I.; Kallioniemi, O.; et al. Lysophosphatidic acid and sphingosine-1-phosphate promote morphogenesis and block invasion of prostate cancer cells in three-dimensional organotypic models. Oncogene 2012, 31, 2075–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, A.; Nagahashi, M.; Aoyagi, T.; Huang, W.-C.; Lima, S.; Hait, N.C.; Maiti, A.; Kida, K.; Terracina, K.P.; Miyazaki, H.; et al. ABCC1-Exported Sphingosine-1-phosphate, Produced by Sphingosine Kinase 1, Shortens Survival of Mice and Patients with Breast Cancer. Mol. Cancer Res. 2018, 16, 1059–1070. [Google Scholar] [CrossRef] [Green Version]
- Saddoughi, S.A.; Song, P.; Ogretmen, B. Roles of Bioactive Sphingolipids in Cancer Biology and Therapeutics. In Lipids in Health and Disease; Springer: Dordrecht, The Netherlands, 2008; pp. 413–440. [Google Scholar] [CrossRef] [Green Version]
- Ihlefeld, K.; Vienken, H.; Claas, R.F.; Blankenbach, K.; Rudowski, A.; ter Braak, M.; Koch, A.; Van Veldhoven, P.P.; Pfeilschifter, J.; zu Heringdorf, D.M. Upregulation of ABC transporters contributes to chemoresistance of sphingosine 1-phosphate lyase-deficient fibroblasts. J. Lipid Res. 2015, 56, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Matula, K.; Collie-Duguid, E.; Murray, G.; Parikh, K.; Grabsch, H.; Tan, P.; Lalwani, S.; Garau, R.; Ong, Y.; Bain, G.; et al. Regulation of cellular sphingosine-1-phosphate by sphingosine kinase 1 and sphingosine-1-phopshate lyase determines chemotherapy resistance in gastroesophageal cancer. BMC Cancer 2015, 15, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Pyne, N.J.; Pyne, S. Recent advances in the role of sphingosine 1-phosphate in cancer. FEBS Lett. 2020, 594, 3583–3601. [Google Scholar] [CrossRef]
- Le, Y.; Shen, X.; Kang, H.; Wang, Q.; Li, K.; Zheng, J.; Yu, Y. Accelerated, untargeted metabolomics analysis of cutaneous T-cell lymphoma reveals metabolic shifts in plasma and tumor adjacent skins of xenograft mice. Biol. Mass Spectrom. 2018, 53, 172–182. [Google Scholar] [CrossRef]
- Bayerl, M.G.; Bruggeman, R.D.; Conroy, E.J.; Hengst, J.A.; King, T.S.; Jimenez, M.; Claxton, D.F.; Yun, J.K. Sphingosine kinase 1 protein and mRNA are overexpressed in non-Hodgkin lymphomas and are attractive targets for novel pharmacological interventions. Leuk. Lymphoma 2008, 49, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Sun, W.; Webb, T.J. Sphingosine Kinase Blockade Leads to Increased Natural Killer T Cell Responses to Mantle Cell Lymphoma. Cells 2020, 9, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagchi, S.; Li, S.; Wang, C.-R. CD1b-autoreactive T cells recognize phospholipid antigens and contribute to antitumor immunity against a CD1b+ T cell lymphoma. OncoImmunology 2016, 5, e1213932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupino, L.; Perry, T.; Margielewska, S.; Hollows, R.; Ibrahim, M.; Care, M.; Allegood, J.; Tooze, R.; Sabbadini, R.; Reynolds, G.; et al. Sphingosine-1-phosphate signalling drives an angiogenic transcriptional programme in diffuse large B cell lymphoma. Leukemia 2019, 33, 2884–2897. [Google Scholar] [CrossRef] [Green Version]
- Seymour, J.F.; Pfreundschuh, M.; Trnĕný, M.; Sehn, L.H.; Catalano, J.; Csinady, E.; Moore, N.; Coiffier, B.; on behalf of the MAIN Study Investigators. R-CHOP with or without bevacizumab in patients with previously untreated diffuse large B-cell lymphoma: Final MAIN study outcomes. Haematologica 2014, 99, 1343–1349. [Google Scholar] [CrossRef] [Green Version]
- Solimando, A.; Annese, T.; Tamma, R.; Ingravallo, G.; Maiorano, E.; Vacca, A.; Specchia, G.; Ribatti, D. New Insights into Diffuse Large B-Cell Lymphoma Pathobiology. Cancers 2020, 12, 1869. [Google Scholar] [CrossRef]
- Kluk, M.J.; Ryan, K.P.; Wang, B.; Zhang, G.; Rodig, S.J.; Sanchez, T. Sphingosine-1-phosphate receptor 1 in classical Hodgkin lymphoma: Assessment of expression and role in cell migration. Lab. Investig. 2013, 93, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, Z.; Wan, W.; Liu, Y.; Jing, H.; Dong, F. FAM19A5/S1PR1 signaling pathway regulates the viability and proliferation of mantle cell lymphoma. J. Recept. Signal Transduct. 2021, 1–5. [Google Scholar] [CrossRef]
- Rao, S.; Cai, K.Q.; Stadanlick, J.E.; Greenberg-Kushnir, N.; Solanki-Patel, N.; Lee, S.-Y.; Fahl, S.; Testa, J.R.; Wiest, D.L. Ribosomal Protein Rpl22 Controls the Dissemination of T-cell Lymphoma. Cancer Res. 2016, 76, 3387–3396. [Google Scholar] [CrossRef] [Green Version]
- Koresawa, R.; Yamazaki, K.; Oka, D.; Fujiwara, H.; Nishimura, H.; Akiyama, T.; Hamasaki, S.; Wada, H.; Sugihara, T.; Sadahira, Y. Sphingosine-1-phosphate receptor 1 as a prognostic biomarker and therapeutic target for patients with primary testicular diffuse large B-cell lymphoma. Br. J. Haematol. 2016, 174, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Paik, J.H.; Nam, S.J.; Kim, T.M.; Heo, D.S.; Kim, C.-W.; Jeon, Y.K. Overexpression of sphingosine-1-phosphate receptor 1 and phospho-signal transducer and activator of transcription 3 is associated with poor prognosis in rituximab-treated diffuse large B-cell lymphomas. BMC Cancer 2014, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Middle, S.; Coupland, S.E.; Taktak, A.; Kidgell, V.; Slupsky, J.R.; Pettitt, A.R.; Till, K.J. Immunohistochemical analysis indicates that the anatomical location of B-cell non-Hodgkin’s lymphoma is determined by differentially expressed chemokine receptors, sphingosine-1-phosphate receptors and integrins. Exp. Hematol. Oncol. 2015, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Kawaaz, M.; Sanchez, T.; Kluk, M.J. Evaluation of S1PR1, pSTAT3, S1PR2, and FOXP1 expression in aggressive, mature B cell lymphomas. J. Hematop. 2019, 12, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Nedelkovska, H.; Rosenberg, A.F.; Hilchey, S.P.; Hyrien, O.; Burack, W.R.; Quataert, S.A.; Baker, C.M.; Azadniv, M.; Welle, S.L.; Ansell, S.M.; et al. Follicular Lymphoma Tregs Have a Distinct Transcription Profile Impacting Their Migration and Retention in the Malignant Lymph Node. PLoS ONE 2016, 11, e0155347. [Google Scholar] [CrossRef] [Green Version]
- Wasik, A.M.; Wu, C.; Mansouri, L.; Rosenquist, R.; Pan-Hammarstrom, Q.; Sander, B. Clinical and functional impact of recurrent S1PR1 mutations in mantle cell lymphoma. Blood Adv. 2018, 2, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Hill, H.A.; Qi, X.; Jain, P.; Nomie, K.; Wang, Y.; Zhou, S.; Wang, M.L. Genetic mutations and features of mantle cell lymphoma: A systematic review and meta-analysis. Blood Adv. 2020, 4, 2927–2938. [Google Scholar] [CrossRef]
- Sadeghi, L.; Arvidsson, G.; Merrien, M.; Wasik, A.M.; Görgens, A.; Smith, C.E.; Sander, B.; Wright, A.P. Differential B-Cell Receptor Signaling Requirement for Adhesion of Mantle Cell Lymphoma Cells to Stromal Cells. Cancers 2020, 12, 1143. [Google Scholar] [CrossRef]
- Nishimura, H.; Akiyama, T.; Monobe, Y.; Matsubara, K.; Igarashi, Y.; Abe, M.; Sugihara, T.; Sadahira, Y. Expression of sphingosine-1-phosphate receptor 1 in mantle cell lymphoma. Mod. Pathol. 2010, 23, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Bouska, A.; Zhang, W.; Gong, Q.; Iqbal, J.; Scuto, A.; Vose, J.; Ludvigsen, M.; Fu, K.; Weisenburger, D.D.; Greiner, T.C.; et al. Combined copy number and mutation analysis identifies oncogenic pathways associated with transformation of follicular lymphoma. Leukemia 2017, 31, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Stelling, A.; Hashwah, H.; Bertram, K.; Manz, M.; Tzankov, A.; Müller, A. The tumor suppressive TGF-β/SMAD1/S1PR2 signaling axis is recurrently inactivated in diffuse large B-cell lymphoma. Blood 2018, 131, 2235–2246. [Google Scholar] [CrossRef] [Green Version]
- Flori, M.; Schmid, C.A.; Sumrall, E.T.; Tzankov, A.; Law, C.W.; Robinson, M.; Müller, A. The hematopoietic oncoprotein FOXP1 promotes tumor cell survival in diffuse large B-cell lymphoma by repressing S1PR2 signaling. Blood 2016, 127, 1438–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orgueira, A.M.; Ferro, R.F.; Arias, J.D.; Santos, C.A.; Rodríguez, B.A.; Pérez, L.B.; Vence, N.A.; López, B.; Blanco, A.A.; Valentín, P.M.; et al. Detection of new drivers of frequent B-cell lymphoid neoplasms using an integrated analysis of whole genomes. PLoS ONE 2021, 16, e0248886. [Google Scholar] [CrossRef]
- Green, J.A.; Suzuki, K.; Cho, B.; Willison, L.D.; Palmer, D.; Allen, C.D.C.; Schmidt, T.H.; Xu, Y.; Proia, R.L.; Coughlin, S.R.; et al. The sphingosine 1-phosphate receptor S1P2 maintains the homeostasis of germinal center B cells and promotes niche confinement. Nat. Immunol. 2011, 12, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Cattoretti, G.; Mandelbaum, J.; Lee, N.; Chaves, A.H.; Mahler, A.M.; Chadburn, A.; Dalla-Favera, R.; Pasqualucci, L.; MacLennan, A.J. Targeted Disruption of the S1P2 Sphingosine 1-Phosphate Receptor Gene Leads to Diffuse Large B-Cell Lymphoma Formation. Cancer Res. 2009, 69, 8686–8692. [Google Scholar] [CrossRef] [Green Version]
- Muppidi, J.; Schmitz, R.; Green, J.A.; Xiao, W.; Larsen, A.B.; Braun, S.E.; An, J.; Xu, Y.; Rosenwald, A.; Ott, G.; et al. Loss of signalling via Gα13 in germinal centre B-cell-derived lymphoma. Nature 2014, 516, 254–258. [Google Scholar] [CrossRef] [Green Version]
- Castro, B.M.; Prieto, M.; Silva, L.C. Ceramide: A simple sphingolipid with unique biophysical properties. Prog. Lipid Res. 2014, 54, 53–67. [Google Scholar] [CrossRef]
- Nganga, R.; Oleinik, N.; Ogretmen, B. Mechanisms of Ceramide-Dependent Cancer Cell Death. Adv Cancer Res. 2018, 140, 1–25. [Google Scholar] [CrossRef]
- Hait, N.C.; Maiti, A. The Role of Sphingosine-1-Phosphate and Ceramide-1-Phosphate in Inflammation and Cancer. Mediat. Inflamm. 2017, 2017, 1–17. [Google Scholar] [CrossRef]
- Abou-Ghali, M.; Stiban, J. Regulation of ceramide channel formation and disassembly: Insights on the initiation of apoptosis. Saudi J. Biol. Sci. 2015, 22, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C.A.; Hunn, M.; Rojas, P.; Torres, V.; Leyton, L.; Quest, A.F.G. Caspase-dependent initiation of apoptosis and necrosis by the Fas receptor in lymphoid cells: Onset of necrosis is associated with delayed ceramide increase. J. Cell Sci. 2002, 115, 4671–4683. [Google Scholar] [CrossRef] [Green Version]
- Govindarajah, N.; Clifford, R.; Bowden, D.; Sutton, P.; Parsons, J.; Vimalachandran, D. Sphingolipids and acid ceramidase as therapeutic targets in cancer therapy. Crit. Rev. Oncol. 2019, 138, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Cremesti, A.; Paris, F.; Grassmé, H.; Holler, N.; Tschopp, J.; Fuks, Z.; Gulbins, E.; Kolesnick, R. Ceramide Enables Fas to Cap and Kill. J. Biol. Chem. 2001, 276, 23954–23961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalfant, C.E.; Rathman, K.; Pinkerman, R.L.; Wood, R.E.; Obeid, L.M.; Ogretmen, B.; Hannun, Y.A. De Novo Ceramide Regulates the Alternative Splicing of Caspase 9 and Bcl-x in A549 Lung Adenocarcinoma Cells. Dependence on protein phosphatase-1. J. Biol. Chem. 2002, 277, 12587–12595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.-T.; Wu, C.-Y.; Lin, Y.-C.; Wu, M.-T.; Su, K.-L.; Yuan, S.-S.; Wang, H.-M.D.; Fong, Y.; Lin, Y.-H.; Chiu, C.-C. C2-Ceramide-Induced Rb-Dominant Senescence-Like Phenotype Leads to Human Breast Cancer MCF-7 Escape from p53-Dependent Cell Death. Int. J. Mol. Sci. 2019, 20, 4292. [Google Scholar] [CrossRef] [Green Version]
- Sundararaj, K.P.; Wood, R.E.; Ponnusamy, S.; Salas, A.M.; Szulc, Z.; Bielawska, A.; Obeid, L.M.; Hannun, Y.A.; Ogretmen, B. Rapid Shortening of Telomere Length in Response to Ceramide Involves the Inhibition of Telomere Binding Activity of Nuclear Glyceraldehyde-3-phosphate Dehydrogenase. J. Biol. Chem. 2004, 279, 6152–6162. [Google Scholar] [CrossRef] [Green Version]
- Chalfant, C.E.; Kishikawa, K.; Mumby, M.C.; Kamibayashi, C.; Bielawska, A.; Hannun, Y.A. Long Chain Ceramides Activate Protein Phosphatase-1 and Protein Phosphatase-2A. J. Biol. Chem. 1999, 274, 20313–20317. [Google Scholar] [CrossRef] [Green Version]
- Ruvolo, P. Ceramide regulates cellular homeostasis via diverse stress signaling pathways. Leukemia 2001, 15, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Nepali, P.R.; Haimovitz-Friedman, A. Chemotherapeutic Agents-Induced Ceramide-Rich Platforms (CRPs) in Endothelial Cells and Their Modulation. In Methods in Molecular Biology; Springer: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Furuya, H.; Shimizu, Y.; Kawamori, T. Sphingolipids in cancer. Cancer Metastasis Rev. 2011, 30, 567–576. [Google Scholar] [CrossRef]
- Schiffmann, S.; Sandner, J.; Birod, K.; Wobst, I.; Angioni, C.; Ruckhäberle, E.; Kaufmann, M.; Ackermann, H.; Lötsch, J.; Schmidt, H.; et al. Ceramide synthases and ceramide levels are increased in breast cancer tissue. Carcinogenesis 2009, 30, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Roh, J.-L.; Park, J.Y.; Kim, E.H.; Jang, H.J. Targeting acid ceramidase sensitises head and neck cancer to cisplatin. Eur. J. Cancer 2016, 52, 163–172. [Google Scholar] [CrossRef]
- White-Gilbertson, S.; Lu, P.; Norris, J.S.; Voelkel-Johnson, C. Genetic and pharmacological inhibition of acid ceramidase prevents asymmetric cell division by neosis. J. Lipid Res. 2019, 60, 1225–1235. [Google Scholar] [CrossRef]
- Cheng, J.C.; Bai, A.; Beckham, T.H.; Tucker Marrison, S.; Yount, C.L.; Young, K.; Lu, P.; Bartlett, A.M.; Wu, B.X.; Keane, B.J.; et al. Radiation-induced acid ceramidase confers prostate cancer resistance and tumor relapse. J. Clin. Investig. 2013, 123, 4344–4358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Realini, N.; Palese, F.; Pizzirani, D.; Pontis, S.; Basit, A.; Bach, A.; Ganesan, A.; Piomelli, D. Acid Ceramidase in Melanoma. J. Biol. Chem. 2016, 291, 2422–2434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klobučar, M.; Grbčić, P.; Pavelić, S.K.; Jonjić, N.; Visentin, S.; Sedić, M. Acid ceramidase inhibition sensitizes human colon cancer cells to oxaliplatin through downregulation of transglutaminase 2 and β1 integrin/FAK−mediated signalling. Biochem. Biophys. Res. Commun. 2018, 503, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Doan, N.B.; Nguyen, H.S.; Al-Gizawiy, M.M.; Mueller, W.M.; Sabbadini, R.A.; Rand, S.D.; Connelly, J.M.; Chitambar, C.R.; Schmainda, K.; Mirza, S.P. Acid ceramidase confers radioresistance to glioblastoma cells. Oncol. Rep. 2017, 38, 1932–1940. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.-F.; Pearson, J.M.; Feith, D.J.; Loughran, T.P. The emergence of acid ceramidase as a therapeutic target for acute myeloid leukemia. Expert Opin. Ther. Targets 2017, 21, 583–590. [Google Scholar] [CrossRef]
- Swanton, C.; Marani, M.; Pardo, O.; Warne, P.H.; Kelly, G.; Sahai, E.; Elustondo, F.; Chang, J.; Temple, J.; Ahmed, A.A.; et al. Regulators of Mitotic Arrest and Ceramide Metabolism Are Determinants of Sensitivity to Paclitaxel and Other Chemotherapeutic Drugs. Cancer Cell 2007, 11, 498–512. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, M.; Ogiso, H.; Takeuchi, T.; Kitatani, K.; Umehara, H.; Okazaki, T. Lysosomal ceramide generated by acid sphingomyelinase triggers cytosolic cathepsin B-mediated degradation of X-linked inhibitor of apoptosis protein in natural killer/T lymphoma cell apoptosis. Cell Death Dis. 2015, 6, e1717. [Google Scholar] [CrossRef] [Green Version]
- Rimokh, R.; Gadoux, M.; Bertheas, M.; Berger, F.; Garoscio, M.; Deleage, G.; Germain, D.; Magaud, J. FVT-1, a novel human transcription unit affected by variant translocation t(2;18)(p11;q21) of follicular lymphoma. Blood 1993, 81, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Czuchlewski, D.R.; Csernus, B.; Bubman, D.; Hyjek, E.; Martin, P.; Chadburn, A.; Knowles, D.M.; Cesarman, E. Expression of the Follicular Lymphoma Variant Translocation 1 Gene in Diffuse Large B-Cell Lymphoma Correlates With Subtype and Clinical Outcome. Am. J. Clin. Pathol. 2008, 130, 957–962. [Google Scholar] [CrossRef]
- Dai, L.; Trillo-Tinoco, J.; Bai, A.; Chen, Y.; Bielawski, J.; Del Valle, L.; Smith, C.D.; Ochoa, A.C.; Qin, Z.; Parsons, C. Ceramides promote apoptosis for virus-infected lymphoma cells through induction of ceramide synthases and viral lytic gene expression. Oncotarget 2015, 6, 24246–24260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Z.; Dai, L.; Trillo-Tinoco, J.; Senkal, C.; Wang, W.; Reske, T.; Bonstaff, K.; Del Valle, L.; Rodriguez, P.; Flemington, E.; et al. Targeting Sphingosine Kinase Induces Apoptosis and Tumor Regression for KSHV-Associated Primary Effusion Lymphoma. Mol. Cancer Ther. 2013, 13, 154–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Wu, S.; Zhang, Y.; Zi, Y.; Yang, M.; Guo, Y.; Zhang, L.; Wang, L. Ceramide participates in cell programmed death induced by Type II anti-CD20 mAb. J. Cent. South Univ. Med. Sci. 2015, 40, 1292–1297. [Google Scholar] [CrossRef]
- Gustafsson, K.; Sander, B.; Bielawski, J.; Hannun, Y.A.; Flygare, J. Potentiation of Cannabinoid-Induced Cytotoxicity in Mantle Cell Lymphoma through Modulation of Ceramide Metabolism. Mol. Cancer Res. 2009, 7, 1086–1098. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Henriquez, M.; Torres, V.; Moraga, F.; Díaz-Elizondo, J.; Arredondo, C.; Chiong, M.; Olea-Azar, C.; Stutzin, A.; Lavandero, S.; et al. Ceramide-induced formation of ROS and ATP depletion trigger necrosis in lymphoid cells. Free Radic. Biol. Med. 2008, 44, 1146–1160. [Google Scholar] [CrossRef]
- Chen, J.; Goyal, N.; Dai, L.; Lin, Z.; Del Valle, L.; Zabaleta, J.; Liu, J.; Post, S.R.; Foroozesh, M.; Qin, Z. Developing new ceramide analogs and identifying novel sphingolipid-controlled genes against a virus-associated lymphoma. Blood 2020, 136, 2175–2187. [Google Scholar] [CrossRef]
- Cao, Y.; Qiao, J.; Lin, Z.; Zabaleta, J.; Dai, L.; Qin, Z. Up-regulation of tumor suppressor genes by exogenous dhC16-Cer contributes to its anti-cancer activity in primary effusion lymphoma. Oncotarget 2017, 8, 15220–15229. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, R.; Eckes, T.; Imre, G.; Kippenberger, S.; Meissner, M.; Thomas, D.; Trautmann, S.; Merlio, J.-P.; Chevret, E.; Kaufmann, R.; et al. C6 Ceramide (d18:1/6:0) as a Novel Treatment of Cutaneous T Cell Lymphoma. Cancers 2021, 13, 270. [Google Scholar] [CrossRef]
- Voorzanger-Rousselot, N.; Alberti, L.; Blay, J.-Y. CD40L induces multidrug resistance to apoptosis in breast carcinoma and lymphoma cells through caspase independent and dependent pathways. BMC Cancer 2006, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Dai, L.; Bai, A.; Smith, C.D.; Rodriguez, P.C.; Yu, F.; Qin, Z. ABC294640, A Novel Sphingosine Kinase 2 Inhibitor, Induces Oncogenic Virus–Infected Cell Autophagic Death and Represses Tumor Growth. Mol. Cancer Ther. 2017, 16, 2724–2734. [Google Scholar] [CrossRef] [Green Version]
- Bezombes, C.; Grazide, S.; Garret, C.; Fabre, C.; Quillet-Mary, A.; Müller, S.; Jaffrézou, J.-P.; Laurent, G. Rituximab antiproliferative effect in B-lymphoma cells is associated with acid-sphingomyelinase activation in raft microdomains. Blood 2004, 104, 1166–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, H.; Zhang, C.; Su, L.; Bi, X.; Wang, C.; Wang, L.; Wu, B. Type II anti-CD20 mAb-induced lysosome mediated cell death is mediated through a ceramide-dependent pathway. Biochem. Biophys. Res. Commun. 2015, 457, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shu, L.; Wu, J. Ceramide participates in lysosome-mediated cell death induced by type II anti-CD20 monoclonal antibodies. Leuk. Lymphoma 2015, 56, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Alduaij, W.; Ivanov, A.; Honeychurch, J.; Cheadle, E.J.; Potluri, S.; Lim, S.H.; Shimada, K.; Chan, C.H.T.; Tutt, A.; Beers, S.A.; et al. Novel type II anti-CD20 monoclonal antibody (GA101) evokes homotypic adhesion and actin-dependent, lysosome-mediated cell death in B-cell malignancies. Blood 2011, 117, 4519–4529. [Google Scholar] [CrossRef]
- Codini, M.; Garcia-Gil, M.; Albi, E. Cholesterol and Sphingolipid Enriched Lipid Rafts as Therapeutic Targets in Cancer. Int. J. Mol. Sci. 2021, 22, 726. [Google Scholar] [CrossRef]
- Gomez-Larrauri, A.; Ouro, A.; Trueba, M.; Gomez-Muñoz, A. Regulation of cell growth, survival and migration by ceramide 1-phosphate-implications in lung cancer progression and inflammation. Cell. Signal. 2021, 83, 109980. [Google Scholar] [CrossRef]
- Rivera, I.; Ordoñez, M.; Presa, N.; Gangoiti, P.; Gomez-Larrauri, A.; Trueba, M.; Fox, T.; Kester, M.; Muñoz, A.G. Ceramide 1-phosphate regulates cell migration and invasion of human pancreatic cancer cells. Biochem. Pharmacol. 2015, 102, 107–119. [Google Scholar] [CrossRef]
- Schwalm, S.; Erhardt, M.; Römer, I.; Pfeilschifter, J.; Zangemeister-Wittke, U.; Huwiler, A. Ceramide Kinase Is Upregulated in Metastatic Breast Cancer Cells and Contributes to Migration and Invasion by Activation of PI 3-Kinase and Akt. Int. J. Mol. Sci. 2020, 21, 1396. [Google Scholar] [CrossRef] [Green Version]
- Dressler, K.; Kolesnick, R. Ceramide 1-phosphate, a novel phospholipid in human leukemia (HL-60) cells. Synthesis via ceramide from sphingomyelin. J. Biol. Chem. 1990, 265, 14917–14921. [Google Scholar] [CrossRef]
- van der Hoeven, D.; Cho, K.-J.; Zhou, Y.; Ma, X.; Chen, W.; Naji, A.; Montufar-Solis, D.; Zuo, Y.; Kovar, S.E.; Levental, K.R.; et al. Sphingomyelin Metabolism Is a Regulator of K-Ras Function. Mol. Cell. Biol. 2017, 38, e00373-17. [Google Scholar] [CrossRef] [Green Version]
- Yerly, S.; Ding, H.; Tauzin, S.; Van Echten-Deckert, G.; Borisch, B.; Hoessli, D.C. The sphingolipid-rich rafts of ALK+ lymphomas downregulate the Lyn-Cbp/PAG signalosome. Eur. J. Haematol. 2010, 85, 93–98. [Google Scholar] [CrossRef]
- Vykoukal, J.; Fahrmann, J.F.; Gregg, J.R.; Tang, Z.; Basourakos, S.; Irajizad, E.; Park, S.; Yang, G.; Creighton, C.J.; Fleury, A.; et al. Caveolin-1-mediated sphingolipid oncometabolism underlies a metabolic vulnerability of prostate cancer. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Separovic, D.; Shields, A.F.; Philip, P.A.; Bielawski, J.; Bielawska, A.; Pierce, J.S.; Tarca, A.L. Altered Levels of Serum Ceramide, Sphingosine and Sphingomyelin Are Associated with Colorectal Cancer: A Retrospective Pilot Study. Anticancer Res. 2017, 37, 1213–1218. [Google Scholar] [CrossRef] [Green Version]
- Modrak, D.E.; Cardillo, T.M.; Newsome, G.A.; Goldenberg, D.M.; Gold, D.V. Synergistic Interaction between Sphingomyelin and Gemcitabine Potentiates Ceramide-Mediated Apoptosis in Pancreatic Cancer. Cancer Res. 2004, 64, 8405–8410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozhkova, A.V.; Zinovyeva, M.V.; Sass, A.V.; Zborovskaya, I.B.; Limborska, S.A.; Dergunova, L.V. Expression of sphingomyelin synthase 1 (SGMS1) gene varies in human lung and esophagus cancer. Mol. Biol. 2014, 48, 340–346. [Google Scholar] [CrossRef]
- Zheng, K.; Chen, Z.; Feng, H.; Chen, Y.; Zhang, C.; Yu, J.; Luo, Y.; Zhao, L.; Jiang, X.; Shi, F. Sphingomyelin synthase 2 promotes an aggressive breast cancer phenotype by disrupting the homoeostasis of ceramide and sphingomyelin. Cell Death Dis. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Veldman, R.J.; Zerp, S.; Van Blitterswijk, W.J.; Verheij, M. N-hexanoyl-sphingomyelin potentiates in vitro doxorubicin cytotoxicity by enhancing its cellular influx. Br. J. Cancer 2004, 90, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Mazzei, J.C.; Zhou, H.; Brayfield, B.P.; Hontecillas, R.; Bassaganya-Riera, J.; Schmelz, E.M. Suppression of intestinal inflammation and inflammation-driven colon cancer in mice by dietary sphingomyelin: Importance of peroxisome proliferator-activated receptor γ expression. J. Nutr. Biochem. 2011, 22, 1160–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modrak, D.E.; Leon, E.; Goldenberg, D.M.; Gold, D.V. Ceramide Regulates Gemcitabine-Induced Senescence and Apoptosis in Human Pancreatic Cancer Cell Lines. Mol. Cancer Res. 2009, 7, 890–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakor, A.B.A.; Taniguchi, M.; Kitatani, K.; Hashimoto, M.; Asano, S.; Hayashi, A.; Nomura, K.; Bielawski, J.; Bielawska, A.; Watanabe, K.; et al. Sphingomyelin Synthase 1-generated Sphingomyelin Plays an Important Role in Transferrin Trafficking and Cell Proliferation. J. Biol. Chem. 2011, 286, 36053–36062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taniguchi, M.; Ueda, Y.; Matsushita, M.; Nagaya, S.; Hashizume, C.; Arai, K.; Kabayama, K.; Fukase, K.; Watanabe, K.; Wardhani, L.O.; et al. Deficiency of sphingomyelin synthase 2 prolongs survival by the inhibition of lymphoma infiltration through ICAM-1 reduction. FASEB J. 2020, 34, 3838–3854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothwell, J.A.; Murphy, N.; Bešević, J.; Kliemann, N.; Jenab, M.; Ferrari, P.; Achaintre, D.; Gicquiau, A.; Vozar, B.; Scalbert, A.; et al. Metabolic Signatures of Healthy Lifestyle Patterns and Colorectal Cancer Risk in a European Cohort. Clin. Gastroenterol. Hepatol. 2021, 20, e1061–e1082. [Google Scholar] [CrossRef] [PubMed]
- To, N.B.; Nguyen, Y.T.-K.; Moon, J.Y.; Ediriweera, M.K.; Cho, S.K. Pentadecanoic Acid, an Odd-Chain Fatty Acid, Suppresses the Stemness of MCF-7/SC Human Breast Cancer Stem-Like Cells through JAK2/STAT3 Signaling. Nutrients 2020, 12, 1663. [Google Scholar] [CrossRef] [PubMed]
- Ediriweera, M.K.; To, N.B.; Lim, Y.; Cho, S.K. Odd-chain fatty acids as novel histone deacetylase 6 (HDAC6) inhibitors. Biochimie 2021, 186, 147–156. [Google Scholar] [CrossRef]
- Matejcic, M.; Lesueur, F.; Biessy, C.; Renault, A.-L.; Mebirouk, N.; Yammine, S.; Keski-Rahkonen, P.; Li, K.; Hémon, B.; Weiderpass, E.; et al. Circulating plasma phospholipid fatty acids and risk of pancreatic cancer in a large European cohort. Int. J. Cancer 2018, 143, 2437–2448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, A.; Ishida, F.; Nakazawa, H.; Yamaura, M.; Morita, S.; Uehara, T.; Honda, T.; Hidaka, H. Serum sphingomyelin species profile is altered in hematologic malignancies. Clin. Chim. Acta 2021, 514, 29–33. [Google Scholar] [CrossRef]
- Kölzer, M.; Arenz, C.; Ferlinz, K.; Werth, N.; Schulze, H.; Klingenstein, R.; Sandhoff, K. Phosphatidylinositol-3,5-Bisphosphate Is a Potent and Selective Inhibitor of Acid Sphingomyelinase. Biol. Chem. 2003, 384, 1293–1298. [Google Scholar] [CrossRef]
- Cumin, C.; Huang, Y.-L.; Everest-Dass, A.; Jacob, F. Deciphering the Importance of Glycosphingolipids on Cellular and Molecular Mechanisms Associated with Epithelial-to-Mesenchymal Transition in Cancer. Biomolecules 2021, 11, 62. [Google Scholar] [CrossRef]
- Boscher, C.; Zheng, Y.Z.; Lakshminarayan, R.; Johannes, L.; Dennis, J.W.; Foster, L.J.; Nabi, I.R. Galectin-3 Protein Regulates Mobility of N-cadherin and GM1 Ganglioside at Cell-Cell Junctions of Mammary Carcinoma Cells. J. Biol. Chem. 2012, 287, 32940–32952. [Google Scholar] [CrossRef] [Green Version]
- Van Slambrouck, S.; Groux-Degroote, S.; Krzewinski-Recchi, M.-A.; Cazet, A.; Delannoy, P.; Steelant, W.F.A. Carbohydrate-to-carbohydrate interactions between α2,3-linked sialic acids on α2 integrin subunits and asialo-GM1 underlie the bone metastatic behaviour of LNCAP-derivative C4-2B prostate cancer cells. Biosci. Rep. 2014, 34, 546–557. [Google Scholar] [CrossRef]
- Talabnin, K.; Talabnin, C.; Kumagai, T.; Sutatum, N.; Khiaowichit, J.; Dechsukhum, C.; Ishihara, M.; Azadi, P.; Sripa, B. Ganglioside GM2: A potential biomarker for cholangiocarcinoma. J. Int. Med Res. 2020, 48, 0300060520903216. [Google Scholar] [CrossRef] [PubMed]
- Ruckhäberle, E.; Karn, T.; Rody, A.; Hanker, L.; Gätje, R.; Metzler, D.; Holtrich, U.; Kaufmann, M. Gene expression of ceramide kinase, galactosyl ceramide synthase and ganglioside GD3 synthase is associated with prognosis in breast cancer. J. Cancer Res. Clin. Oncol. 2009, 135, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Dobrenkov, K.; Ostrovnaya, I.; Gu, J.; Cheung, I.Y.; Cheung, N.-K.V. Oncotargets GD2 and GD3 are highly expressed in sarcomas of children, adolescents, and young adults. Pediatr. Blood Cancer 2016, 63, 1780–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.; Mikami, M.; Aoki, D.; Kiguchi, K.; Ishiwata, I.; Iwamori, M. Expression of sulfatide and sulfated lactosylceramide among histological types of human ovarian carcinomas. Hum. Cell 2015, 28, 37–43. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Y.; Momin, A.; Shaner, R.; Wang, E.; Bowen, N.J.; Matyunina, L.V.; Walker, L.D.; McDonald, J.F.; Sullards, M.C.; et al. Elevation of sulfatides in ovarian cancer: An integrated transcriptomic and lipidomic analysis including tissue-imaging mass spectrometry. Mol. Cancer 2010, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Moghimi, B.; Muthugounder, S.; Jambon, S.; Tibbetts, R.; Hung, L.; Bassiri, H.; Hogarty, M.D.; Barrett, D.M.; Shimada, H.; Asgharzadeh, S. Preclinical assessment of the efficacy and specificity of GD2-B7H3 SynNotch CAR-T in metastatic neuroblastoma. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Charan, M.; Dravid, P.; Cam, M.; Audino, A.; Gross, A.C.; Arnold, M.A.; Roberts, R.D.; Cripe, T.P.; Pertsemlidis, A.; Houghton, P.J.; et al. GD2-directed CAR-T cells in combination with HGF-targeted neutralizing antibody (AMG102) prevent primary tumor growth and metastasis in Ewing sarcoma. Int. J. Cancer 2019, 146, 3184–3195. [Google Scholar] [CrossRef]
- Andersch, L.; Radke, J.; Klaus, A.; Schwiebert, S.; Winkler, A.; Schumann, E.; Grunewald, L.; Zirngibl, F.; Flemmig, C.; Jensen, M.C.; et al. CD171- and GD2-specific CAR-T cells potently target retinoblastoma cells in preclinical in vitro testing. BMC Cancer 2019, 19, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.S.; Yu, A.L.; Tseng, L.M.; Chow, L.W.C.; Hou, M.F.; Hurvitz, S.A.; Schwab, R.B.; Murray, J.L.; Chang, H.K.; Chang, H.T.; et al. Globo H-KLH vaccine adagloxad simolenin (OBI-822)/OBI-821 in patients with metastatic breast cancer: Phase II randomized, placebo-controlled study. J. Immunother. Cancer 2020, 8, e000342. [Google Scholar] [CrossRef]
- Giussani, P.; Prinetti, A.; Tringali, C. The Role of Sphingolipids in Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 6492. [Google Scholar] [CrossRef]
- Yu, J.; Hung, J.T.; Wang, S.H.; Cheng, J.Y.; Yu, A.L. Targeting glycosphingolipids for cancer immunotherapy. FEBS Lett. 2020, 594, 3602–3618. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-T.; Kim, M.-J.; Kang, Y.-H.; Choi, S.-Y.; Lee, J.-H.; Choi, J.-A.; Kang, C.-M.; Cho, C.-K.; Kang, S.; Bae, S.; et al. Phytosphingosine in combination with ionizing radiation enhances apoptotic cell death in radiation-resistant cancer cells through ROS-dependent and -independent AIF release. Blood 2005, 105, 1724–1733. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Park, M.-T.; Yoon, C.-H.; Byun, J.-Y.; Lee, S.-J. Activation of Lck is critically required for sphingosine-induced conformational activation of Bak and mitochondrial cell death. Biochem. Biophys. Res. Commun. 2008, 370, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Escribà-Garcia, L.; Alvarez-Fernández, C.; Tellez-Gabriel, M.; Sierra, J.; Briones, J. Dendritic cells combined with tumor cells and α-galactosylceramide induce a potent, therapeutic and NK-cell dependent antitumor immunity in B cell lymphoma. J. Transl. Med. 2017, 15, 115. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; Wang, F.; Stieren, E.; Tong, Q. SIRT3, a Mitochondrial Sirtuin Deacetylase, Regulates Mitochondrial Function and Thermogenesis in Brown Adipocytes. J. Biol. Chem. 2005, 280, 13560–13567. [Google Scholar] [CrossRef] [Green Version]
- Alderliesten, M.C.; Klarenbeek, J.B.; van der Luit, A.H.; van Lummel, M.; Jones, D.R.; Zerp, S.; Divecha, N.; Verheij, M.; van Blitterswijk, W.J. Phosphoinositide phosphatase SHIP-1 regulates apoptosis induced by edelfosine, Fas ligation and DNA damage in mouse lymphoma cells. Biochem. J. 2011, 440, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Kume, M.; Kiyohara, E.; Matsumura, Y.; Koguchi-Yoshioka, H.; Tanemura, A.; Hanaoka, Y.; Taminato, M.; Tashima, H.; Tomita, K.; Kubo, T.; et al. Ganglioside GD3 May Suppress the Functional Activities of Benign Skin T Cells in Cutaneous T-Cell Lymphoma. Front. Immunol. 2021, 12, 1026. [Google Scholar] [CrossRef]
- Madigan, J.P.; Robey, R.W.; Poprawski, J.E.; Huang, H.; Clarke, C.J.; Gottesman, M.M.; Cabot, M.C.; Rosenberg, D.W. A role for ceramide glycosylation in resistance to oxaliplatin in colorectal cancer. Exp. Cell Res. 2020, 388, 111860. [Google Scholar] [CrossRef]
- Barth, B.M.; Gustafson, S.J.; Young, M.M.; Fox, T.E.; Shanmugavelandy, S.S.; Kaiser, J.M.; Cabot, M.C.; Kester, M.; Kuhn, T.B. Inhibition of NADPH oxidase by glucosylceramide confers chemoresistance. Cancer Biol. Ther. 2010, 10, 1126–1136. [Google Scholar] [CrossRef] [Green Version]
- Gouazé, V.; Yu, J.Y.; Bleicher, R.J.; Han, T.Y.; Liu, Y.Y.; Wang, H.; Gottesman, M.M.; Bitterman, A.; Giuliano, A.E.; Cabot, M.C. Overexpression of glucosylceramide synthase and P-glycoprotein in cancer cells selected for resistance to natural product chemotherapy. Mol. Cancer Ther. 2004, 3, 633–640. [Google Scholar]
- Morjani, H.; Aouali, N.; El Btaouri, H.; Dumontet, C.; Eddabra, L.; Malagarie-Cazenave, S.; Madoulet, C. Accumulation of lactosylceramide and overexpression of a PSC833-resistant P-glycoprotein in multidrug-resistant human sarcoma cells. Oncol. Rep. 2011, 25, 1161–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Y.; Gupta, V.; Patwardhan, G.A.; Bhinge, K.; Zhao, Y.; Bao, J.; Mehendale, H.; Cabot, M.C.; Li, Y.T.; Jazwinski, S.M. Glucosylceramide synthase upregulates MDR1 expression in the regulation of cancer drug resistance through cSrc and β-catenin signaling. Mol. Cancer 2010, 9, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.W.; Park, Y.; Roh, J.-L.; Cho, K.-J.; Choi, S.-H.; Nam, S.Y.; Kim, S.Y. Prognostic value of glucosylceramide synthase and P-glycoprotein expression in oral cavity cancer. Int. J. Clin. Oncol. 2016, 21, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-Y.; Patwardhan, G.A.; Bhinge, K.; Gupta, V.; Gu, X.; Jazwinski, S.M. Suppression of Glucosylceramide Synthase Restores p53-Dependent Apoptosis in Mutant p53 Cancer Cells. Cancer Res. 2011, 71, 2276–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, J.-L.; Kim, E.H.; Park, J.Y.; Kim, J.W. Inhibition of Glucosylceramide Synthase Sensitizes Head and Neck Cancer to Cisplatin. Mol. Cancer Ther. 2015, 14, 1907–1915. [Google Scholar] [CrossRef] [Green Version]
- Suchanski, J.; Grzegrzolka, J.; Owczarek, T.; Pasikowski, P.; Piotrowska, A.; Kocbach, B.; Nowak, A.; Dziegiel, P.; Wojnar, A.; Ugorski, M. Sulfatide decreases the resistance to stress-induced apoptosis and increases P-selectin-mediated adhesion: A two-edged sword in breast cancer progression. Breast Cancer Res. 2018, 20, 133. [Google Scholar] [CrossRef]
- Owczarek, T.B.; Suchanski, J.; Pula, B.; Kmiecik, A.M.; Chadalski, M.; Jethon, A.; Dziegiel, P.; Ugorski, M. Galactosylceramide Affects Tumorigenic and Metastatic Properties of Breast Cancer Cells as an Anti-Apoptotic Molecule. PLoS ONE 2013, 8, e84191. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.W.; Wang, R.; Cai, Q.Q.; Qi, B.; Wu, W.; Zhang, Y.H.; Wu, X.Z. Sulfatide epigenetically regulates miR-223 and promotes the migration of human hepatocellular carcinoma cells. J. Hepatol. 2014, 60, 792–801. [Google Scholar] [CrossRef]
- Grasso‡, C.; Field‡, C.S.; Tang, C.-W.; Ferguson, P.M.; Compton, B.J.; Anderson, R.J.; Painter, G.F.; Weinkove, R.; Hermans, I.F.; Berridge, M.V. Vaccines adjuvanted with an NKT cell agonist induce effective T-cell responses in models of CNS lymphoma. Immunotherapy 2020, 12, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-C.; Chuang, Y.-T.; Hsu, Y.-T.; Huang, J.-T.; Wu, T.-C.; Hung, C.-F.; Yang, Y.-C.; Chang, C.-L. Intra-Peritoneal Hyperthermia Combining α-Galactosylceramide in the Treatment of Ovarian Cancer. PLoS ONE 2013, 8, e69336. [Google Scholar] [CrossRef] [Green Version]
- Sainz, V.; Moura, L.; Peres, C.; de Matos, A.I.N.; Viana, A.; Wagner, A.M.; Ramirez, J.E.V.; Barata, T.S.; Gaspar, M.M.; Brocchini, S.; et al. α-Galactosylceramide and peptide-based nano-vaccine synergistically induced a strong tumor suppressive effect in melanoma. Acta Biomater. 2018, 76, 193–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bhave, M.S.; Yagita, H.; Cardell, S.L. Natural Killer T-Cell Agonist α-Galactosylceramide and PD-1 Blockade Synergize to Reduce Tumor Development in a Preclinical Model of Colon Cancer. Front. Immunol. 2020, 11, 581301. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, E.; Archer, J.; Wang, S.Z.; Dekker, N.; Aerts, J.; Karlsson, S.; Cox, T.M. Inhibition of UDP-glucosylceramide synthase in mice prevents Gaucher disease-associated B-cell malignancy. J. Pathol. 2015, 235, 113–124. [Google Scholar] [CrossRef]
- Wątek, M.; Piktel, E.; Wollny, T.; Durnaś, B.; Fiedoruk, K.; Lech-Marańda, E.; Bucki, R. Defective Sphingolipids Metabolism and Tumor Associated Macrophages as the Possible Links Between Gaucher Disease and Blood Cancer Development. Int. J. Mol. Sci. 2019, 20, 843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouaze-Andersson, V.; Cabot, M.C. Sphingolipid metabolism and drug resistance in hematological malignancies. Anti-Cancer Agents Med. Chem. 2012, 11, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, P.; LeLeux, J.; Liu, J.; Roy, K. A simple, clinically relevant therapeutic vaccine shows long-term protection in an aggressive, delayed-treatment B lymphoma model. JCI Insight 2017, 2, e92522. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Y.; Yun, M.M.; Dong, X.; Xu, M.; Zhao, R.; Han, X.; Zhou, E.; Yun, F.; Su, W.; Liu, C.; et al. Combination of cytokine-induced killer and dendritic cells pulsed with antigenic α-1,3-galactosyl epitope–enhanced lymphoma cell membrane for effective B-cell lymphoma immunotherapy. Cytotherapy 2016, 18, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Robert, A.; Wiels, J. Shiga Toxins as Antitumor Tools. Toxins 2021, 13, 690. [Google Scholar] [CrossRef]
- Suchanowska, A.; Kaczmarek, R.; Duk, M.; Lukasiewicz, J.; Smolarek, D.; Majorczyk, E.; Jaskiewicz, E.; Laskowska, A.; Waśniowska, K.; Grodecka, M.; et al. A Single Point Mutation in the Gene Encoding Gb3/CD77 Synthase Causes a Rare Inherited Polyagglutination Syndrome. J. Biol. Chem. 2012, 287, 38220–38230. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Miyata, Y.; Matsuo, T.; Shida, Y.; Hakariya, T.; Ohba, K.; Taima, T.; Ito, A.; Suda, T.; Hakomori, S.-I.; et al. Stage-specific embryonic antigen-4 is a histological marker reflecting the malignant behavior of prostate cancer. Glycoconj. J. 2019, 36, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.-H.; Lin, R.-J.; Hung, J.-T.; Hsieh, C.-B.; Hung, T.-H.; Lo, F.-Y.; Ho, M.-Y.; Yeh, C.-T.; Huang, Y.-L.; Yu, J.; et al. High expression FUT1 and B3GALT5 is an independent predictor of postoperative recurrence and survival in hepatocellular carcinoma. Sci. Rep. 2017, 7, 10750. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.; Hung, J.; Wu, C.; Huang, Y.; Lee, C.; Yeh, C.; Chung, Y.; Lo, F.; Lai, L.; Tung, J.K.; et al. Globo H Is a Promising Theranostic Marker for Intrahepatic Cholangiocarcinoma. Hepatol. Commun. 2021, 6, 194–208. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-W.; Lee, C.H.; Lee, P.; Lin, J.; Hsu, C.-W.; Hung, J.-T.; Lin, J.-J.; Yu, J.-C.; Shao, L.-E.; Yu, J.; et al. Expression of Globo H and SSEA3 in breast cancer stem cells and the involvement of fucosyl transferases 1 and 2 in Globo H synthesis. Proc. Natl. Acad. Sci. USA 2008, 105, 11667–11672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.-P.; Yang, P.-S.; Chien, M.-N.; Chen, M.-J.; Lee, J.-J.; Liu, C.-L. Aberrant expression of tumor-associated carbohydrate antigen Globo H in thyroid carcinoma. J. Surg. Oncol. 2016, 114, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, N.; Suzuki, Y.; Takahashi, H.; Uemura, M.; Nishimura, J.; Hata, T.; Takemasa, I.; Mizushima, T.; Ishii, H.; Doki, Y.; et al. SSEA-3 as a novel amplifying cancer cell surface marker in colorectal cancers. Int. J. Oncol. 2013, 42, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stimmer, L.; Dehay, S.; Nemati, F.; Massonnet, G.; Richon, S.; Decaudin, D.; Klijanienko, J.; Johannes, L. Human breast cancer and lymph node metastases express Gb3 and can be targeted by STxB-vectorized chemotherapeutic compounds. BMC Cancer 2014, 14, 916. [Google Scholar] [CrossRef]
- Tyler, A.; Johansson, A.; Karlsson, T.; Gudey, S.K.; Brännström, T.; Grankvist, K.; Behnam-Motlagh, P. Targeting glucosylceramide synthase induction of cell surface globotriaosylceramide (Gb3) in acquired cisplatin-resistance of lung cancer and malignant pleural mesothelioma cells. Exp. Cell Res. 2015, 336, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Geyer, P.E.; Maak, M.; Nitsche, U.; Perl, M.; Novotny, A.; Slotta-Huspenina, J.; Dransart, E.; Holtorf, A.; Johannes, L.; Janssen, K.-P. Gastric Adenocarcinomas Express the Glycosphingolipid Gb3/CD77: Targeting of Gastric Cancer Cells with Shiga Toxin B-Subunit. Mol. Cancer Ther. 2016, 15, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Kwak, C.-Y.; Shayman, J.A.; Kim, J.H. Globoside promotes activation of ERK by interaction with the epidermal growth factor receptor. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Giussani, P.; Tringali, C.; Riboni, L.; Viani, P.; Venerando, B. Sphingolipids: Key Regulators of Apoptosis and Pivotal Players in Cancer Drug Resistance. Int. J. Mol. Sci. 2014, 15, 4356–4392. [Google Scholar] [CrossRef] [Green Version]
- Roy, K.R.; Uddin, M.B.; Roy, S.C.; Hill, R.A.; Marshall, J.; Li, Y.; Chamcheu, J.C.; Lu, H.; Liu, Y. Gb3-cSrc complex in glycosphingolipid-enriched microdomains contributes to the expression of p53 mutant protein and cancer drug resistance via β-catenin–activated RNA methylation. FASEB BioAdvances 2020, 2, 653–667. [Google Scholar] [CrossRef] [PubMed]
- Chuang, P.K.; Hsiao, M.; Hsu, T.L.; Chang, C.F.; Wu, C.Y.; Chen, B.R.; Huang, H.W.; Liao, K.S.; Chen, C.C.; Chen, C.L.; et al. Signaling pathway of globo-series glycosphingolipids and β1,3-galactosyltransferase V (β3GalT5) in breast cancer. Proc. Natl. Acad. Sci. USA 2019, 116, 3518–3523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steelant, W.F.; Kawakami, Y.; Ito, A.; Handa, K.; Bruyneel, E.A.; Mareel, M.; Hakomori, S. Monosialyl-Gb5 organized with cSrc and FAK in GEM of human breast carcinoma MCF-7 cells defines their invasive properties. FEBS Lett. 2002, 531, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Van Slambrouck, S.; Steelant, W.F.A. Clustering of monosialyl-Gb5 initiates downstream signalling events leading to invasion of MCF-7 breast cancer cells. Biochem. J. 2007, 401, 689–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.Y.; Wang, S.H.; Lin, J.; Tsai, Y.C.; Yu, J.; Wu, J.C.; Hung, J.T.; Lin, J.J.; Wu, Y.Y.; Yeh, K.T.; et al. Globo-H Ceramide Shed from Cancer Cells Triggers Translin-Associated Factor X-Dependent Angiogenesis. Cancer Res. 2014, 74, 6856–6866. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, R.M.; Chamba, A.; Holder, M.J.; Challa, A.; Smith, D.C.; Hodgkin, M.N.; Lord, J.M.; Gordon, J. Dynamic interplay between the neutral glycosphingolipid CD77/Gb3 and the therapeutic antibody target CD20 within the lipid bilayer of model B lymphoma cells. Biochem. Biophys. Res. Commun. 2007, 355, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Maloney, M.D.; Binnington-Boyd, B.; Lingwood, C.A. Globotriaosyl ceramide modulates interferon-α-induced growth inhibition and CD19 expression in Burkitt’s lymphoma cells. Glycoconj. J. 1999, 16, 821–828. [Google Scholar] [CrossRef]
- Junqua, S.; Larsen, A.K.; Wils, P.; Mishal, Z.; Wiels, J.; Le Pecq, J.B. Decreased accessibility of globotriaosylceramide associated with decreased tumorigenicity in Burkitt’s lymphoma variants induced by immunoselection. Cancer Res. 1989, 49, 6480–6486. [Google Scholar]
- Jackson, T.; Van Exel, C.; Reagans, K.; Verret, R.; Maloney, M. Comparison of adhesion mechanisms and surface protein expression in CD77-positive and CD77-negative Burkitt’s lymphoma cells. Cell. Mol. Biol. 2001, 47, 1195–1200. [Google Scholar]
- Maloney, M.D.; A Lingwood, C. CD19 has a potential CD77 (globotriaosyl ceramide)-binding site with sequence similarity to verotoxin B-subunits: Implications of molecular mimicry for B cell adhesion and enterohemorrhagic Escherichia coli pathogenesis. J. Exp. Med. 1994, 180, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Mori, T.; Kiyokawa, N.; Katagiri, Y.U.; Taguchi, T.; Suzuki, T.; Sekino, T.; Sato, N.; Ohmi, K.; Nakajima, H.; Takeda, T.; et al. Globotriaosyl ceramide (CD77/Gb3) in the glycolipid-enriched membrane domain participates in B-cell receptor–mediated apoptosis by regulating Lyn kinase activity in human B cells. Exp. Hematol. 2000, 28, 1260–1268. [Google Scholar] [CrossRef]
- Tétaud, C.; Falguières, T.; Carlier, K.; Lécluse, Y.; Garibal, J.; Coulaud, D.; Busson, P.; Steffensen, R.; Clausen, H.; Johannes, L.; et al. Two Distinct Gb3/CD77 Signaling Pathways Leading to Apoptosis Are Triggered by Anti-Gb3/CD77 mAb and Verotoxin-1. J. Biol. Chem. 2003, 278, 45200–45208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pudymaitis, A.; Armstrong, G.; Lingwood, C. Verotoxin-resistant cell clones are deficient in the glycolipid globotriosylceramide: Differential basis of phenotype. Arch. Biochem. Biophys. 1991, 286, 448–452. [Google Scholar] [CrossRef]
- Đevenica, D.; Čulić, V.; Vuica, A.; Markotić, A. Biochemical, pathological and oncological relevance of Gb3Cer receptor. Med. Oncol. 2011, 28, 675–684. [Google Scholar] [CrossRef]
- Fujii, Y.; Dohmae, N.; Takio, K.; Kawsar, S.M.A.; Matsumoto, R.; Hasan, I.; Koide, Y.; Kanaly, R.A.; Yasumitsu, H.; Ogawa, Y.; et al. A Lectin from the Mussel Mytilus galloprovincialis Has a Highly Novel Primary Structure and Induces Glycan-mediated Cytotoxicity of Globotriaosylceramide-expressing Lymphoma Cells. J. Biol. Chem. 2012, 287, 44772–44783. [Google Scholar] [CrossRef] [Green Version]
- Hasan, I.; Sugawara, S.; Fujii, Y.; Koide, Y.; Terada, D.; Iimura, N.; Fujiwara, T.; Takahashi, K.G.; Kojima, N.; Rajia, S.; et al. MytiLec, a Mussel R-Type Lectin, Interacts with Surface Glycan Gb3 on Burkitt’s Lymphoma Cells to Trigger Apoptosis through Multiple Pathways. Mar. Drugs 2015, 13, 7377–7389. [Google Scholar] [CrossRef] [Green Version]
- Debernardi, J.; Hollville, E.; Lipinski, M.; Wiels, J.; Robert, A. Differential role of FL-BID and t-BID during verotoxin-1-induced apoptosis in Burkitt’s lymphoma cells. Oncogene 2018, 37, 2410–2421. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, G.; Capasso, S.; Sticco, L.; Russo, D. Glycosphingolipids: Synthesis and functions. FEBS J. 2013, 280, 6338–6353. [Google Scholar] [CrossRef]
- Iwabuchi, K. Gangliosides in the Immune System: Role of Glycosphingolipids and Glycosphingolipid-Enriched Lipid Rafts in Immunological Functions. In Gangliosides. Methods in Molcular Biology; Sonnino, S., Prinetti, A., Eds.; Humana Press: New York, NY, USA, 2018; Volume 1804, pp. 83–95. [Google Scholar] [CrossRef]
- Shihashi, G.; Yagi, T.; Suzuki, S.; Seki, M.; Kohashi, S.; Ueda, T.; Kameyama, K.; Kusunoki, S.; Nakajima, H.; Okamoto, S.; et al. Immune-mediated Neuropathy with Anti-disialosyl IgM Antibodies in Diffuse Large B-cell Lymphoma: A Case Report and Literature Review. Intern. Med. 2015, 54, 1647–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Péguet-Navarro, J.; Sportouch, M.; Popa, I.; Berthier, O.; Schmitt, D.; Portoukalian, J. Gangliosides from Human Melanoma Tumors Impair Dendritic Cell Differentiation from Monocytes and Induce Their Apoptosis. J. Immunol. 2003, 170, 3488–3494. [Google Scholar] [CrossRef] [Green Version]
- Patel, F.; Spassieva, S.D. Side Effects in Cancer Therapy: Are Sphingolipids to Blame? Adv Cancer Res. 2018, 140, 367–388. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-J.; Chung, T.-W.; Kang, S.-K.; Lee, Y.-C.; Ko, J.-H.; Kim, J.-G.; Kim, C.-H. Ganglioside GM3 modulates tumor suppressor PTEN-mediated cell cycle progression—transcriptional induction of p21WAF1 and p27kip1 by inhibition of PI-3K/AKT pathway. Glycobiology 2006, 16, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Tringali, C.; Lupo, B.; Cirillo, F.; Papini, N.; Anastasia, L.; Lamorte, G.; Colombi, P.; Bresciani, R.; Monti, E.; Tettamanti, G.; et al. Silencing of membrane-associated sialidase Neu3 diminishes apoptosis resistance and triggers megakaryocytic differentiation of chronic myeloid leukemic cells K562 through the increase of ganglioside GM3. Cell Death Differ. 2009, 16, 164–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, Y.; Izumoto, S.; Suzuki, T.; Kinoshita, M.; Kagawa, N.; Wada, K.; Hashimoto, N.; Maruno, M.; Nakatsuji, Y.; Yoshimine, T. Ganglioside GM3 inhibits proliferation and invasion of glioma. J. Neuro-Oncol. 2005, 71, 99–106. [Google Scholar] [CrossRef]
- Mitsuda, T.; Furukawa, K.; Fukumoto, S.; Miyazaki, H.; Urano, T.; Furukawa, K. Overexpression of Ganglioside GM1 Results in the Dispersion of Platelet-derived Growth Factor Receptor from Glycolipid-enriched Microdomains and in the Suppression of Cell Growth Signals. J. Biol. Chem. 2002, 277, 11239–11246. [Google Scholar] [CrossRef] [Green Version]
- Ono, M.; Handa, K.; Sonnino, S.; Withers, D.A.; Nagai, H.; Hakomori, S.-I. GM3 Ganglioside Inhibits CD9-Facilitated Haptotactic Cell Motility: Coexpression of GM3 and CD9 Is Essential in the Downregulation of Tumor Cell Motility and Malignancy. Biochemistry 2001, 40, 6414–6421. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Sun, P.; Paller, A. Ganglioside GM3 Inhibits Matrix Metalloproteinase-9 Activation and Disrupts Its Association with Integrin. J. Biol. Chem. 2003, 278, 25591–25599. [Google Scholar] [CrossRef] [Green Version]
- Prinetti, A.; Aureli, M.; Illuzzi, G.; Prioni, S.; Nocco, V.; Scandroglio, F.; Gagliano, N.; Tredici, G.; Rodriguez-Menendez, V.; Chigorno, V.; et al. GM3 synthase overexpression results in reduced cell motility and in caveolin-1 upregulation in human ovarian carcinoma cells. Glycobiology 2009, 20, 62–77. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.W.; Sun, P.; Yan, Q.; Paller, A.S.; Gerami, P.; Ho, N.; Vashi, N.; Le Poole, I.C.; Wang, X.Q. De-N-acetyl GM3 Promotes Melanoma Cell Migration and Invasion through Urokinase Plasminogen Activator Receptor Signaling-Dependent MMP-2 Activation. Cancer Res. 2009, 69, 8662–8669. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Sun, M.; Yu, M.; Fu, Q.; Jiang, H.; Yu, G.; Li, G. Gangliosides profiling in serum of breast cancer patient: GM3 as a potential diagnostic biomarker. Glycoconj. J. 2019, 36, 419–428. [Google Scholar] [CrossRef]
- Gu, Y.; Zhang, J.; Mi, W.; Yang, J.; Han, F.; Lu, X.; Yu, W. Silencing of GM3 synthase suppresses lung metastasis of murine breast cancer cells. Breast Cancer Res. 2008, 10, R1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonhard, V.; Alasino, R.V.; Pasqualini, M.E.; Cremonezzi, D.C.; García, N.H.; Beltramo, D.M. Monosialoganglioside GM1 reduces toxicity of Ptx and increase anti-metastasic effect in a murine mammary cancer model. Sci. Rep. 2020, 10, 10191. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Mohanty, K.C. Correlation of gangliosides GM2 and GM3 with metastatic potential to lungs of mouse B16 melanoma. J. Exp. Clin. Cancer Res. 2003, 22, 125–134. [Google Scholar] [PubMed]
- Noguchi, M.; Suzuki, T.; Kabayama, K.; Takahashi, H.; Chiba, H.; Shiratori, M.; Abe, S.; Watanabe, A.; Satoh, M.; Hasegawa, T.; et al. GM3 synthase gene is a novel biomarker for histological classification and drug sensitivity against epidermal growth factor receptor tyrosine kinase inhibitors in non-small cell lung cancer. Cancer Sci. 2007, 98, 1625–1632. [Google Scholar] [CrossRef]
- Noguchi, M.; Kabayama, K.; Uemura, S.; Kang, B.-W.; Saito, M.; Igarashi, Y.; Inokuchi, J.-I. Endogenously produced ganglioside GM3 endows etoposide and doxorubicin resistance by up-regulating Bcl-2 expression in 3LL Lewis lung carcinoma cells. Glycobiology 2006, 16, 641–650. [Google Scholar] [CrossRef]
- Tringali, C.A.; Lupo, B.; Silvestri, I.; Papini, N.; Anastasia, L.; Tettamanti, G.; Venerando, B. The Plasma Membrane Sialidase NEU3 Regulates the Malignancy of Renal Carcinoma Cells by Controlling β1 Integrin Internalization and Recycling. J. Biol. Chem. 2012, 287, 42835–42845. [Google Scholar] [CrossRef] [Green Version]
- Hyuga, S.; Kawasaki, N.; Hyuga, M.; Ohta, M.; Shibayama, R.; Kawanishi, T.; Yamagata, S.; Yamagata, T.; Hayakawa, T. Ganglioside GD1a inhibits HGF-induced motility and scattering of cancer cells through suppression of tyrosine phosphorylation of c-Met. Int. J. Cancer 2001, 94, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Takaku, S.; Wang, P.; Hu, D.; Hyuga, S.; Sato, T.; Yamagata, S.; Yamagata, T. Ganglioside GD1a regulation of caveolin-1 and Stim1 expression in mouse FBJ cells:Augmented expression of caveolin-1 and Stim1 in cells with increased GD1a content. Glycoconj. J. 2006, 23, 303–315. [Google Scholar] [CrossRef]
- Díaz, M.I.; Díaz, P.; Bennett, J.C.; Urra, H.; Ortiz, R.; Orellana, P.C.; Hetz, C.; Quest, A.F.G. Caveolin-1 suppresses tumor formation through the inhibition of the unfolded protein response. Cell Death Dis. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Mandal, C.; Sarkar, S.; Chatterjee, U.; Schwartz-Albiez, R.; Mandal, C. Disialoganglioside GD3-synthase over expression inhibits survival and angiogenesis of pancreatic cancer cells through cell cycle arrest at S-phase and disruption of integrin-β1-mediated anchorage. Int. J. Biochem. Cell Biol. 2014, 53, 162–173. [Google Scholar] [CrossRef]
- Colell, A.; Morales, A.; Fernández-Checa, J.C.; García-Ruiz, C. Ceramide generated by acidic sphingomyelinase contributes to tumor necrosis factor-α-mediated apoptosis in human colon HT-29 cells through glycosphingolipids formation: Possible role of ganglioside GD3. FEBS Lett. 2002, 526, 135–141. [Google Scholar] [CrossRef] [Green Version]
- García-Ruiz, C.; Colell, A.; París, R.; Fernández-Checa, J.C. Direct interaction of GD3 ganglioside with mitochondria generates reactive oxygen species followed by mitochondrial permeability transition, cytochrome c release, and caspase activation. FASEB J. 2000, 14, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lluis, J.M.; Llacuna, L.; Von Montfort, C.; Bárcena, C.; Enrich, C.; Morales, A.; Fernandez-Checa, J.C. GD3 Synthase Overexpression Sensitizes Hepatocarcinoma Cells to Hypoxia and Reduces Tumor Growth by Suppressing the cSrc/NF-κB Survival Pathway. PLoS ONE 2009, 4, e8059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, K.; Kambe, M.; Miyata, M.; Ohkawa, Y.; Tajima, O.; Furukawa, K. Ganglioside GD3 induces convergence and synergism of adhesion and hepatocyte growth factor/Met signals in melanomas. Cancer Sci. 2014, 105, 52–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, T.R.; Battula, V.L.; Werden, S.J.; Vijay, G.V.; Ramirez-Peña, E.Q.; Taube, J.H.; Chang, J.T.; Miura, N.; Porter, W.; Sphyris, N.; et al. GD3 synthase regulates epithelial–mesenchymal transition and metastasis in breast cancer. Oncogene 2014, 34, 2958–2967. [Google Scholar] [CrossRef] [Green Version]
- Ando, I.; Hoon, D.S.B.; Suzuki, Y.; Saxton, R.E.; Golub, S.H.; Irie, R.F. Ganglioside GM2 on the K562 cell line is recognized as a target structure by human natural killer cells. Int. J. Cancer 1987, 40, 12–17. [Google Scholar] [CrossRef]
- Kaucic, K.; Liu, Y.; Ladisch, S. Modulation of Growth Factor Signaling by Gangliosides: Positive Or Negative? Methods Enzymol. 2006, 417, 168–185. [Google Scholar] [CrossRef] [Green Version]
- Marconi, M.; Ascione, B.; Ciarlo, L.; Vona, R.; Garofalo, T.; Sorice, M.; Gianni, A.M.; Locatelli, S.L.; Carlo-Stella, C.; Malorni, W.; et al. Constitutive localization of DR4 in lipid rafts is mandatory for TRAIL-induced apoptosis in B-cell hematologic malignancies. Cell Death Dis. 2013, 4, e863. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, M.; Kurano, M.; Nitta, T.; Kanoh, H.; Inokuchi, J.-I.; Yatomi, Y. Serum GM3(d18:1-16:0) and GM3(d18:1-24:1) levels may be associated with lymphoma: An exploratory study with haematological diseases. Sci. Rep. 2019, 9, 6308. [Google Scholar] [CrossRef] [Green Version]
- Venkatanarayanan, S.; Nagarajan, B. Multiple marker validity of urinary hexosaminidase and polyamines in haematopoietic malignancy. Biochem. Int. 1989, 18, 301–309. [Google Scholar]
- Anh-Tuan, N.; Pick, J.; Mód, A.; Hollán, S. Gangliosides in acute myeloid leukaemia (AML) and non-hodgkin’s lymphoma (NHL). Eur. J. Cancer Clin. Oncol. 1986, 22, 1003–1007. [Google Scholar] [CrossRef]
- O’Boyle, K.P.; Freeman, K.; Kalisiak, A.; Agregado, A.; Scheinberg, D.A. Patterns of Ganglioside Expression in B Cell Neoplasms. Leuk. Lymphoma 1996, 21, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Büschenfelde, C.M.Z.; Feuerstacke, Y.; Götze, K.S.; Scholze, K.; Peschel, C. GM1 Expression of Non-Hodgkin’s Lymphoma Determines Susceptibility to Rituximab Treatment. Cancer Res. 2008, 68, 5414–5422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, O.; Tasaki, K.; Kusakabe, T.; Abe, M. UDP-GlcNAc2-epimerase regulates cell surface sialylation and ceramide-induced cell death in human malignant lymphoma. Int. J. Mol. Med. 2008, 22, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, O.; Nozawa, Y.; Abe, M. Regulatory roles of cell surface sialylation in susceptibility to sphingomyelinase in human diffuse large B cell lymphoma. Int. J. Oncol. 2005, 27, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, O.; Abe, M.; Hashimoto, Y. Sialylation by β-galactoside α-2,6-sialyltransferase and N-glycans regulate cell adhesion and invasion in human anaplastic large cell lymphoma. Int. J. Oncol. 2015, 46, 973–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, O.; Abe, M.; Hashimoto, Y. Sialylation and glycosylation modulate cell adhesion and invasion to extracellular matrix in human malignant lymphoma: Dependency on integrin and the Rho GTPase family. Int. J. Oncol. 2015, 47, 2091–2099. [Google Scholar] [CrossRef] [Green Version]
- Ballereau, S.; Levade, T.; Genisson, Y.; Andrieu-Abadie, N. Alteration of ceramide 1-O-functionalization as a promising approach for cancer therapy. Anti-Cancer Agents Med. Chem. 2012, 12, 316–328. [Google Scholar] [CrossRef]
- Watters, R.; Fox, T.E.; Tan, S.-F.; Shanmugavelandy, S.; Choby, J.; Broeg, K.; Liao, J.; Kester, M.; Cabot, M.C.; Loughran, T.P.; et al. Targeting glucosylceramide synthase synergizes with C6-ceramide nanoliposomes to induce apoptosis in natural killer cell leukemia. Leuk. Lymphoma 2012, 54, 1288–1296. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.; Costa-Pinheiro, P.; Patterson, L.; Drews, K.; Spiegel, S.; Kester, M. Novel Sphingolipid-Based Cancer Therapeutics in the Personalized Medicine Era. Adv. Cancer Res. 2018, 140, 327–366. [Google Scholar] [CrossRef]
- Canals, D.; Perry, D.M.; Jenkins, R.W.; Hannun, Y.A. Drug targeting of sphingolipid metabolism: Sphingomyelinases and ceramidases. Br. J. Pharmacol. 2011, 163, 694–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canals, D.; Hannun, Y.A. Novel Chemotherapeutic Drugs in Sphingolipid Cancer Research. Handb. Exp. Pharmacol. 2013, 215, 211–238. [Google Scholar] [CrossRef] [Green Version]
- Skácel, J.; Slusher, B.S.; Tsukamoto, T. Small Molecule Inhibitors Targeting Biosynthesis of Ceramide, the Central Hub of the Sphingolipid Network. J. Med. Chem. 2021, 64, 279–297. [Google Scholar] [CrossRef]
- Adan-Gokbulut, A.; Kartal-Yandim, M.; Iskender, G.; Baran, Y. Novel Agents Targeting Bioactive Sphingolipids for the Treatment of Cancer. Curr. Med. Chem. 2012, 20, 108–122. [Google Scholar] [CrossRef]
- Liu, J.; Beckman, B.S.; Foroozesh, M. A review of ceramide analogs as potential anticancer agents. Futur. Med. Chem. 2013, 5, 1405–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metkar, S.S.; Anand, M.; Manna, P.P.; Naresh, K.; Nadkarni, J. Ceramide-Induced Apoptosis in Fas-Resistant Hodgkin’s Disease Cell Lines Is Caspase Independent. Exp. Cell Res. 2000, 255, 18–29. [Google Scholar] [CrossRef]
- Kuroki, J.; Hirokawa, M.; Kitabayashi, A.; Lee, M.; Horiuchi, T.; Kawabata, Y.; Miura, A.B. Cell-permeable ceramide inhibits the growth of B lymphoma Raji cells lacking TNF-α-receptors by inducing G0/G1 arrest but not apoptosis: A new model for dissecting cell-cycle arrest and apoptosis. Leukemia 1996, 10, 1950–1958. [Google Scholar]
- Gustafsson, K.; Christensson, B.; Sander, B.; Flygare, J. Cannabinoid Receptor-Mediated Apoptosis Induced byR(+)-Methanandamide and Win55,212-2 Is Associated with Ceramide Accumulation and p38 Activation in Mantle Cell Lymphoma. Mol. Pharmacol. 2006, 70, 1612–1620. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, M.; Kantarjian, H.; Ravandi, F. Acute promyelocytic leukemia current treatment algorithms. Blood Cancer J. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Bernasconi, E.; Gaudio, E.; Kwee, I.; Rinaldi, A.; Cascione, L.; Tarantelli, C.; Mensah, A.A.; Stathis, A.; Zucca, E.; Vesci, L.; et al. The novel atypical retinoid ST5589 down-regulates Aurora Kinase A and has anti-tumour activity in lymphoma pre-clinical models. Br. J. Haematol. 2015, 171, 378–386. [Google Scholar] [CrossRef]
- Karam, L.; Houshaymi, B.; Abdel-Samad, R.; Jaafar, M.; Halloum, I.; Pisano, C.; Neipel, F.; Darwiche, N.; Merhi, R.A. Antitumor activity of the synthetic retinoid ST1926 on primary effusion lymphoma in vitro and in vivo models. Oncol. Rep. 2017, 39, 721–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghandour, B.; Pisano, C.; Darwiche, N.; Dbaibo, G. Restoration of ceramide de novo synthesis by the synthetic retinoid ST1926 as it induces adult T-cell leukemia cell death. Biosci. Rep. 2020, 40, 10. [Google Scholar] [CrossRef] [PubMed]
- Dbaibo, G.S.; Kfoury, Y.; Darwiche, N.; Panjarian, S.; Kozhaya, L.; Nasr, R.; Abdallah, M.; Hermine, O.; El-Sabban, M.; De Thé, H.; et al. Arsenic trioxide induces accumulation of cytotoxic levels of ceramide in acute promyelocytic leukemia and adult T-cell leukemia/lymphoma cells through de novo ceramide synthesis and inhibition of glucosylceramide synthase activity. Haematologica 2007, 92, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barna, G.; Sebestyén, A.; Weischede, S.; Peták, I.; Mihalik, R.; Formelli, F.; Kopper, L. Different ways to induce apoptosis by fenretinide and all-trans-retinoic acid in human B lymphoma cells. Anticancer Res. 2005, 25, 4179–4185. [Google Scholar]
- Hsieh, T.-C.; Wu, J.M. Apoptosis and Restriction of G1/S Cell Cycle by Fenretinide in Burkitt’s Lymphoma Mutu I Cell Line Accessed with bcl-6 Down-Regulation. Biochem. Biophys. Res. Commun. 2000, 276, 1295–1301. [Google Scholar] [CrossRef]
- Darwiche, N.; Abou-Lteif, G.; Bazarbachi, A. Reactive oxygen species mediate N-(4-hydroxyphenyl)retinamide-induced cell death in malignant T cells and are inhibited by the HTLV-I oncoprotein Tax. Leukemia 2007, 21, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Cowan, A.J.; Frayo, S.L.; Press, O.; Palanca-Wessels, M.C.; Pagel, J.M.; Green, D.J.; Gopal, A.K. Bortezomib and fenretinide induce synergistic cytotoxicity in mantle cell lymphoma through apoptosis, cell-cycle dysregulation, and IκBα kinase downregulation. Anticancer Drugs 2015, 26, 974–983. [Google Scholar] [CrossRef]
- Makena, M.R.; Koneru, B.; Nguyen, T.; Kang, M.H.; Reynolds, C.P. Reactive Oxygen Species–Mediated Synergism of Fenretinide and Romidepsin in Preclinical Models of T-cell Lymphoid Malignancies. Mol. Cancer Ther. 2017, 16, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Makena, M.R.; Nguyen, T.H.; Koneru, B.; Hindle, A.; Chen, W.-H.; Verlekar, D.U.; Kang, M.H.; Reynolds, C.P. Vorinostat and fenretinide synergize in preclinical models of T-cell lymphoid malignancies. Anticancer Drugs 2020, 32, 34–43. [Google Scholar] [CrossRef]
- Gopal, A.K.; Pagel, J.M.; Hedin, N.; Press, O.W. Fenretinide enhances rituximab-induced cytotoxicity against B-cell lymphoma xenografts through a caspase-dependent mechanism. Blood 2004, 103, 3516–3520. [Google Scholar] [CrossRef]
- Cowan, A.J.; Cowan, A.J.; Stevenson, P.A.; Stevenson, P.A.; Gooley, T.A.; Gooley, T.A.; Frayo, S.L.; Frayo, S.L.; Oliveira, G.R.; Oliveira, G.R.; et al. Results of a phase I-II study of fenretinide and rituximab for patients with indolent B-cell lymphoma and mantle cell lymphoma. Br. J. Haematol. 2017, 176, 583–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohrbacher, A.M.; Yang, A.S.; Groshen, S.; Kummar, S.; Gutierrez, M.E.; Kang, M.H.; Tsao-Wei, D.; Reynolds, C.P.; Newman, E.M.; Maurer, B.J. Phase I Study of Fenretinide Delivered Intravenously in Patients with Relapsed or Refractory Hematologic Malignancies: A California Cancer Consortium Trial. Clin. Cancer Res. 2017, 23, 4550–4555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.-F.; Liu, X.; Fox, T.E.; Barth, B.; Sharma, A.; Turner, S.; Awwad, A.; Dewey, A.; Doi, K.; Spitzer, B.; et al. Acid ceramidase is upregulated in AML and represents a novel therapeutic target. Oncotarget 2016, 7, 83208–83222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmura, S.J.; Nodzenski, E.; Kharbanda, S.; Pandey, P.; Quintáns, J.; Kufe, D.W.; Weichselbaum, R.R. Down-Regulation of Ceramide Production Abrogates Ionizing Radiation-Induced CytochromecRelease and Apoptosis. Mol. Pharmacol. 2000, 57, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Vethakanraj, H.S.; Sesurajan, B.P.; Padmanaban, V.P.; Jayaprakasam, M.; Murali, S.; Sekar, A.K. Anticancer effect of acid ceramidase inhibitor ceranib-2 in human breast cancer cell lines MCF-7, MDA MB-231 by the activation of SAPK/JNK, p38 MAPK apoptotic pathways, inhibition of the Akt pathway, downregulation of ERα. Anticancer Drugs 2018, 29, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Adibhatla, R.M.; Hatcher, J.F.; Gusain, A. Tricyclodecan-9-yl-Xanthogenate (D609) Mechanism of Actions: A Mini-Review of Literature. Neurochem. Res. 2011, 37, 671–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schick, H.; Amtmann, E.; Berdel, W.; Danhauser-Riedl, S.; Reichert, A.; Steinhauser, G.; Rastetter, J.; Sauer, G. Antitumoral activity of a xanthate compound I. Cytotoxicity studies with neoplastic cell lines in vitro. Cancer Lett. 1989, 46, 143–147. [Google Scholar] [CrossRef]
- Amtmann, E.; Zöller, M. Stimulation of CD95-induced apoptosis in T-cells by a subtype specific neutral sphingomyelinase inhibitor. Biochem. Pharmacol. 2005, 69, 1141–1148. [Google Scholar] [CrossRef]
- Chen, J.-J.; Long, Z.-J.; Xu, D.-F.; Xiao, R.-Z.; Liu, L.-L.; Xu, Z.-F.; Qiu, S.X.; Lin, D.-J.; Liu, Q. Inhibition of autophagy augments the anticancer activity of α-mangostin in chronic myeloid leukemia cells. Leuk. Lymphoma 2013, 55, 628–638. [Google Scholar] [CrossRef]
- Van Der Luit, A.H.; Vink, S.R.; Klarenbeek, J.B.; Perrissoud, D.; Solary, E.; Verheij, M.; Van Blitterswijk, W.J. A new class of anticancer alkylphospholipids uses lipid rafts as membrane gateways to induce apoptosis in lymphoma cells. Mol. Cancer Ther. 2007, 6, 2337–2345. [Google Scholar] [CrossRef] [Green Version]
- Alonso-Alonso, R.; Mondéjar, R.; Martínez, N.; García-Diaz, N.; Pérez, C.; Merino, D.; Rodríguez, M.; Esteve-Codina, A.; Fuste, B.; Gut, M.; et al. Identification of tipifarnib sensitivity biomarkers in T-cell acute lymphoblastic leukemia and T-cell lymphoma. Sci. Rep. 2020, 10, 6721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Shaheen, S.S.; Dixit, D.; Sen, E. Farnesyltransferase Inhibitor Manumycin Targets IL1β-Ras-HIF-1α Axis in Tumor Cells of Diverse Origin. Inflammation 2012, 35, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Carey, G.B.; Roy, S.K.; Daino, H. The natural tumorcide Manumycin-A targets protein phosphatase 1α and reduces hydrogen peroxide to induce lymphoma apoptosis. Exp. Cell Res. 2015, 332, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.R.; da Silva-Júnior, E.F. Inhibiting the “Undruggable” RAS/Farnesyltransferase (FTase) Cancer Target by Manumycin-related Natural Products. Curr. Med. Chem. 2021, 28, 189–211. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, E.; Yap, M.C.; Iyer, A.; Perinpanayagam, M.A.; Gamma, J.M.; Vincent, K.M.; Lakshmanan, M.; Raju, A.; Tergaonkar, V.; Tan, S.Y.; et al. Targeting N-myristoylation for therapy of B-cell lymphomas. Nat. Commun. 2020, 11, 5348. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Taiyab, A.; Hussain, A.; Alajmi, M.; Islam, A.; Hassan, I. Targeting the Sphingosine Kinase/Sphingosine-1-Phosphate Signaling Axis in Drug Discovery for Cancer Therapy. Cancers 2021, 13, 1898. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sibley, C.D.; Kharel, Y.; Huang, T.; Brown, A.M.; Wonilowicz, L.G.; Bevan, D.R.; Lynch, K.R.; Santos, W.L. Lipophilic tail modifications of 2-(hydroxymethyl)pyrrolidine scaffold reveal dual sphingosine kinase 1 and 2 inhibitors. Bioorganic Med. Chem. 2020, 30, 115941. [Google Scholar] [CrossRef]
- Pitman, M.R.; Powell, J.; Coolen, C.; Moretti, P.A.; Zebol, J.R.; Pham, D.H.; Finnie, J.W.; Don, A.S.; Ebert, L.M.; Bonder, C.S.; et al. A selective ATP-competitive sphingosine kinase inhibitor demonstrates anti-cancer properties. Oncotarget 2015, 6, 7065–7083. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.-B.; Ling, L.-U.; Bunte, R.M.; Chng, W.-J.; Chiu, G.N. In vivo efficacy of a novel liposomal formulation of safingol in the treatment of acute myeloid leukemia. J. Control. Release 2012, 160, 290–298. [Google Scholar] [CrossRef]
- Coward, J.; Ambrosini, G.; Musi, E.; Truman, J.-P.; Haimovitz-Friedman, A.; Allegood, J.C.; Wang, E.; Merrill, J.A.H.; Schwartz, G.K. Safingol (l-threo-sphinganine) induces autophagy in solid tumor cells through inhibition of PKC and the PI3-kinase pathway. Autophagy 2009, 5, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, N.; Morris, E.A.; Kharel, Y.; Raje, M.; Gao, M.; Tomsig, J.L.; Lynch, K.R.; Santos, W. Structure−Activity Relationship Studies and in Vivo Activity of Guanidine-Based Sphingosine Kinase Inhibitors: Discovery of SphK1- and SphK2-Selective Inhibitors. J. Med. Chem. 2015, 58, 1879–1899. [Google Scholar] [CrossRef] [PubMed]
- Stelling, A.; Wu, C.-T.; Bertram, K.; Hashwah, H.; Theocharides, A.; Manz, M.; Tzankov, A.; Müller, A. Pharmacological DNA demethylation restores SMAD1 expression and tumor suppressive signaling in diffuse large B-cell lymphoma. Blood Adv. 2019, 3, 3020–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Zang, Y.Q.; Lu, Y.; Dong, Q.P.; Dong, K.T.; Zhou, H.F. Chlorpromazine hydrochloride plays a tumor suppressive role in diffuse large B lymphoma by promoting the expression of S1PR2. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi 2021, 39, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Deng, J.; Wang, L.; Lee, H.; Armstrong, B.; Scuto, A.; Kowolik, C.; Weiss, L.M.; Forman, S.; Yu, H. S1PR1 is an effective target to block STAT3 signaling in activated B cell–like diffuse large B-cell lymphoma. Blood 2012, 120, 1458–1465. [Google Scholar] [CrossRef]
- Gao, Y.; Ding, X. miR-145-5p exerts anti-tumor effects in diffuse large B-cell lymphoma by regulating S1PR1/STAT3/AKT pathway. Leuk. Lymphoma 2021, 62, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Alshaker, H.; Cooper, C.; Winkler, M.; Pchejetski, D. The emerging role of FTY720 (Fingolimod) in cancer treatment. Oncotarget 2016, 7, 23106–23127. [Google Scholar] [CrossRef] [Green Version]
- Alinari, L.; Baiocchi, R.A.; Prætorius-Ibba, M. FTY720-induced blockage of autophagy enhances anticancer efficacy of milatuzumab in mantle cell lymphoma: Is FTY720 the next autophagy-blocking agent in lymphoma treatment? Autophagy 2012, 8, 416–417. [Google Scholar] [CrossRef] [Green Version]
- Kawai, H.; Matsushita, H.; Akashi, H.; Furuya, D.; Kawakami, S.; Suzuki, R.; Moriuchi, M.; Ogawa, Y.; Kawada, H.; Nakamura, N.; et al. Peripheral T-cell lymphomas as fingolimod-associated lymphoproliferative disorder for patients with multiple sclerosis-case report with literature review. Leuk. Lymphoma 2019, 61, 959–962. [Google Scholar] [CrossRef]
- Cesbron, E.; Monfort, J.B.; Giannesini, C.; Duriez, P.; Moguelet, P.; Senet, P.; Francès, C.; Barbaud, A.; Chasset, F. Primary cutaneous CD30+ T-cell lymphoproliferation during treatment with fingolimod: Case report and literature review. Ann. Dermatol. Venereol. 2018, 145, 433–438. [Google Scholar] [CrossRef]
- Roy, R.; Alotaibi, A.A.; Freedman, M.S. Sphingosine 1-Phosphate Receptor Modulators for Multiple Sclerosis. CNS Drugs 2021, 35, 385–402. [Google Scholar] [CrossRef]
- Khine, A.-A.; Firtel, M.; Lingwood, C.A. CD77-dependent retrograde transport of CD19 to the nuclear membrane: Functional relationship between CD77 and CD19 during germinal center B-cell apoptosis. J. Cell. Physiol. 1998, 176, 281–292. [Google Scholar] [CrossRef]
- Jennemann, R.; Volz, M.; Bestvater, F.; Schmidt, C.; Richter, K.; Kaden, S.; Müthing, J.; Gröne, H.-J.; Sandhoff, R. Blockade of Glycosphingolipid Synthesis Inhibits Cell Cycle and Spheroid Growth of Colon Cancer Cells In Vitro and Experimental Colon Cancer Incidence In Vivo. Int. J. Mol. Sci. 2021, 22, 10539. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, E.; Deroma, L.; Bembi, B.; Deegan, P.; Hollak, C.; Weinreb, N.J.; Cox, T.M. Enzyme replacement and substrate reduction therapy for Gaucher disease. Cochrane Database Syst. Rev. 2015, 2015, CD010324. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, D.; Kikkawa, H.; Ogino, K.; Hirabayashi, Y.; Oku, N.; Taki, T. GD1α-replica peptides functionally mimic GD1α, an adhesion molecule of metastatic tumor cells, and suppress the tumor metastasis. FEBS Lett. 1998, 441, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Berois, N.; Pittini, A.; Osinaga, E. Targeting Tumor Glycans for Cancer Therapy: Successes, Limitations, and Perspectives. Cancers 2022, 14, 645. [Google Scholar] [CrossRef]
- Doronin, I.I.; Vishnyakova, P.A.; Kholodenko, I.V.; Ponomarev, E.D.; Ryazantsev, D.Y.; Molotkovskaya, I.M.; Kholodenko, R.V. Ganglioside GD2 in reception and transduction of cell death signal in tumor cells. BMC Cancer 2014, 14, 295. [Google Scholar] [CrossRef] [Green Version]




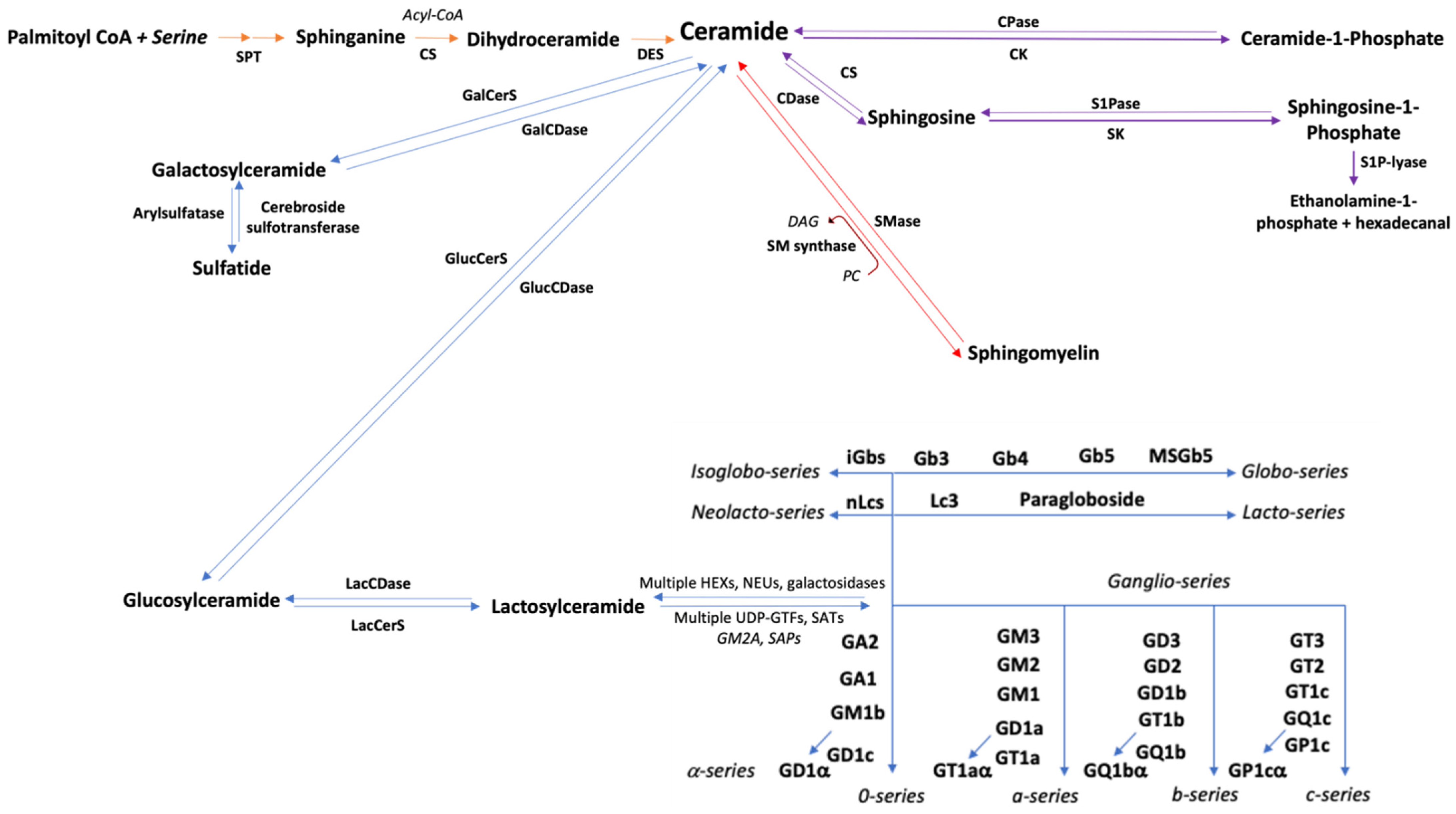{kind=link}
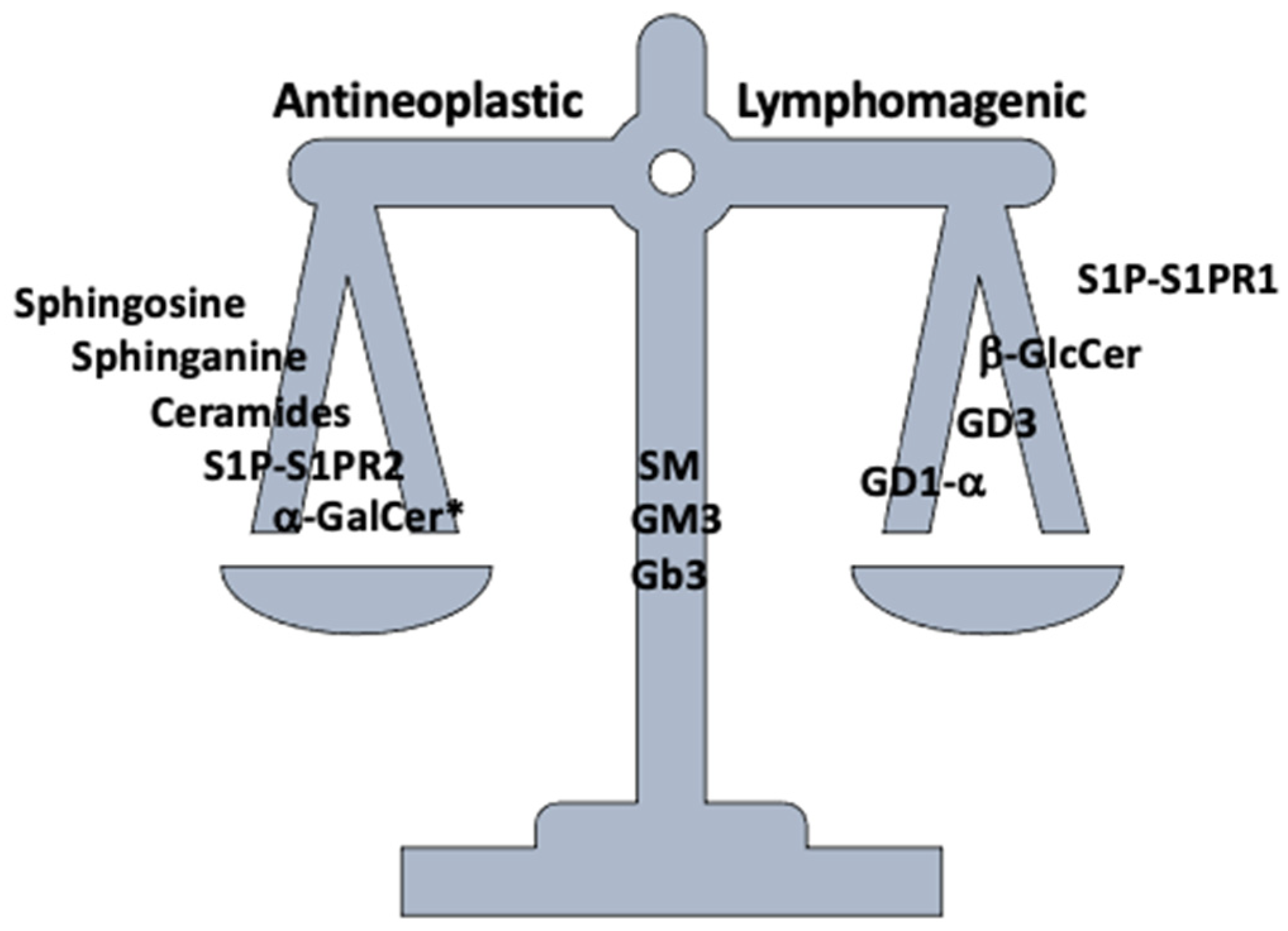{kind=link}
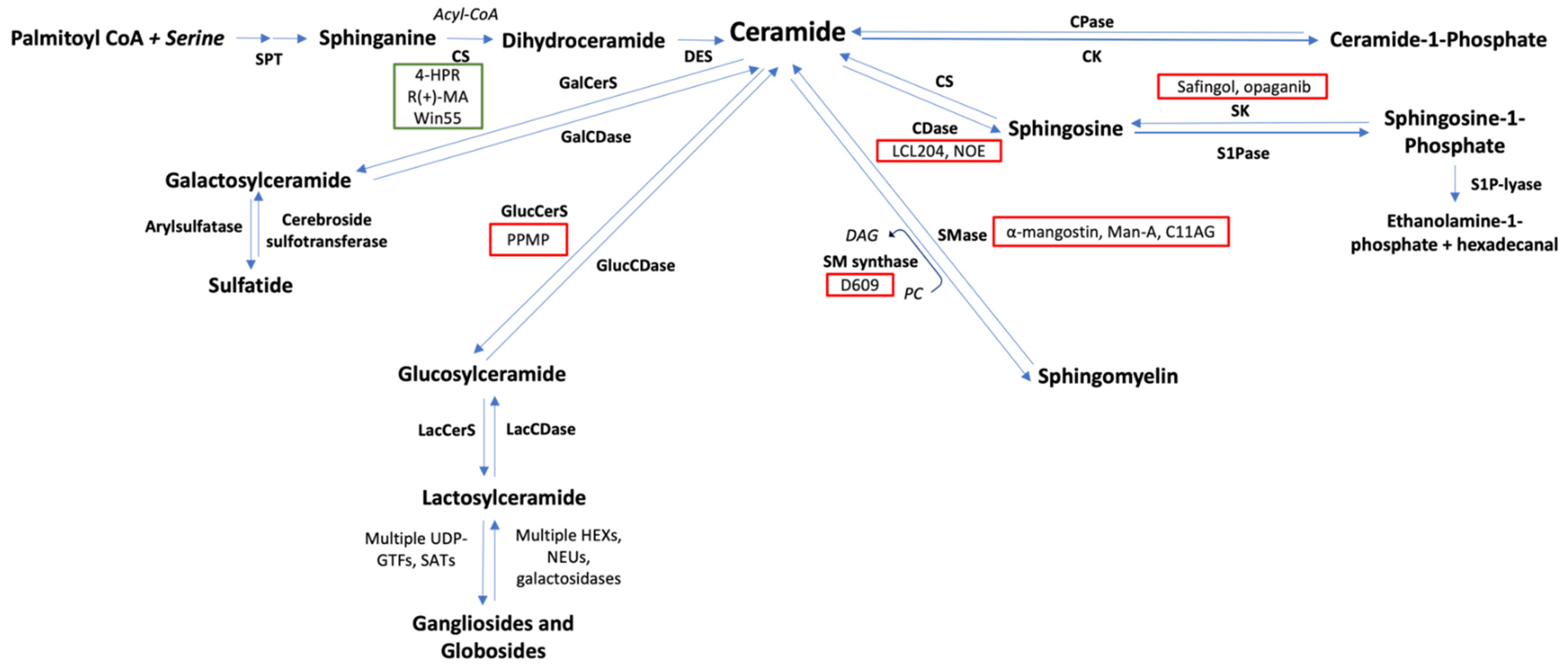{kind=link}
| Sphingolipid | Possible Mechanism | Reference |
|---|---|---|
| Predominantly Antineoplastic | ||
| Sphingoid bases | ↑ ROS ↑ p38, JNK ↑ cPARP ↑ AIF, Bak ↑ Lck ↓ PKC | [55,56,59,197,198] |
| Sphingosine-1-phosphate (S1PR2) | ↓ CXCL12 mediated migration ↓ PI3K-Akt-mTOR ↑ TGF-β/SMAD1/S1PR2 | [17,114,117] |
| Ceramide | ↑ Cytochrome c release ↑ Fas/FasL ↑ ROS ↑ CCL3, RHOB, KLF6, THBS1 ↑ JNK, ERK, p38, p21, p27 ↓ IAP3 ↓ PKC | [123,124,143,150,156] |
| α-galactosylceramide * | ↑ CD1d-mediated immune cytotoxicity | [199] |
| Predominantly Lymphomagenic | ||
| Sphingosine-1-phosphate (S1PR1) | ↓ CD1d-mediated immune cytotoxicity ↑ VEGF-independent angiogenesis ↑ PI3K mediated migration | [96,98,200] |
| Sphingomyelin | ↑ PI3K-Akt ↑ NFκB ↑ ICAM ↑ Transferrin endocytosis | [22,175,176,201] |
| GD3 | ↓ IL-17 | [202] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pherez-Farah, A.; López-Sánchez, R.d.C.; Villela-Martínez, L.M.; Ortiz-López, R.; Beltrán, B.E.; Hernández-Hernández, J.A. Sphingolipids and Lymphomas: A Double-Edged Sword. Cancers 2022, 14, 2051. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14092051
Pherez-Farah A, López-Sánchez RdC, Villela-Martínez LM, Ortiz-López R, Beltrán BE, Hernández-Hernández JA. Sphingolipids and Lymphomas: A Double-Edged Sword. Cancers. 2022; 14(9):2051. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14092051
Chicago/Turabian StylePherez-Farah, Alfredo, Rosa del Carmen López-Sánchez, Luis Mario Villela-Martínez, Rocío Ortiz-López, Brady E. Beltrán, and José Ascención Hernández-Hernández. 2022. "Sphingolipids and Lymphomas: A Double-Edged Sword" Cancers 14, no. 9: 2051. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14092051