
Differential Impacts on Tensional Homeostasis of Gastric Cancer Cells Due to Distinct Domain Variants of E-Cadherin

 , ,
, , 
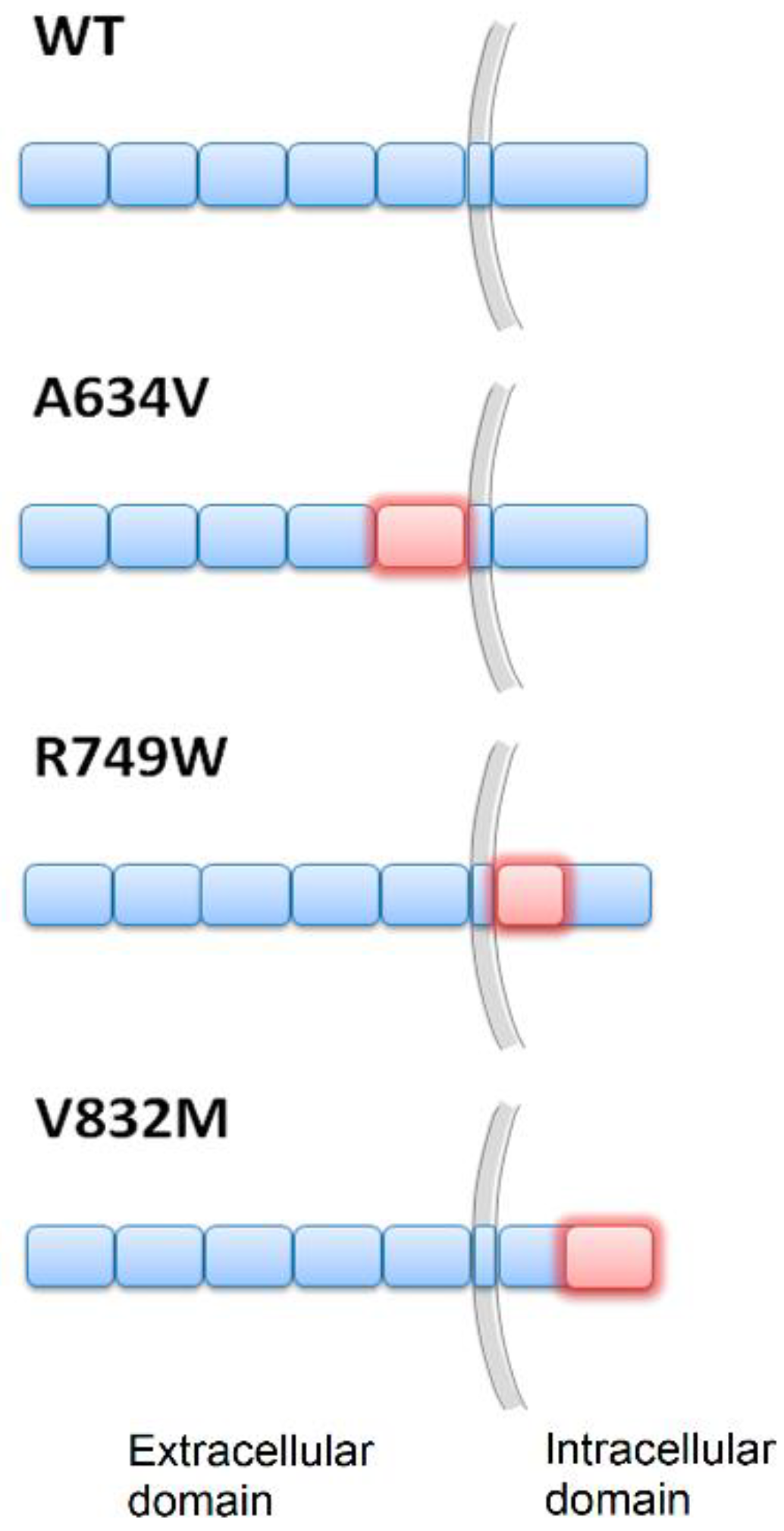{kind=link}
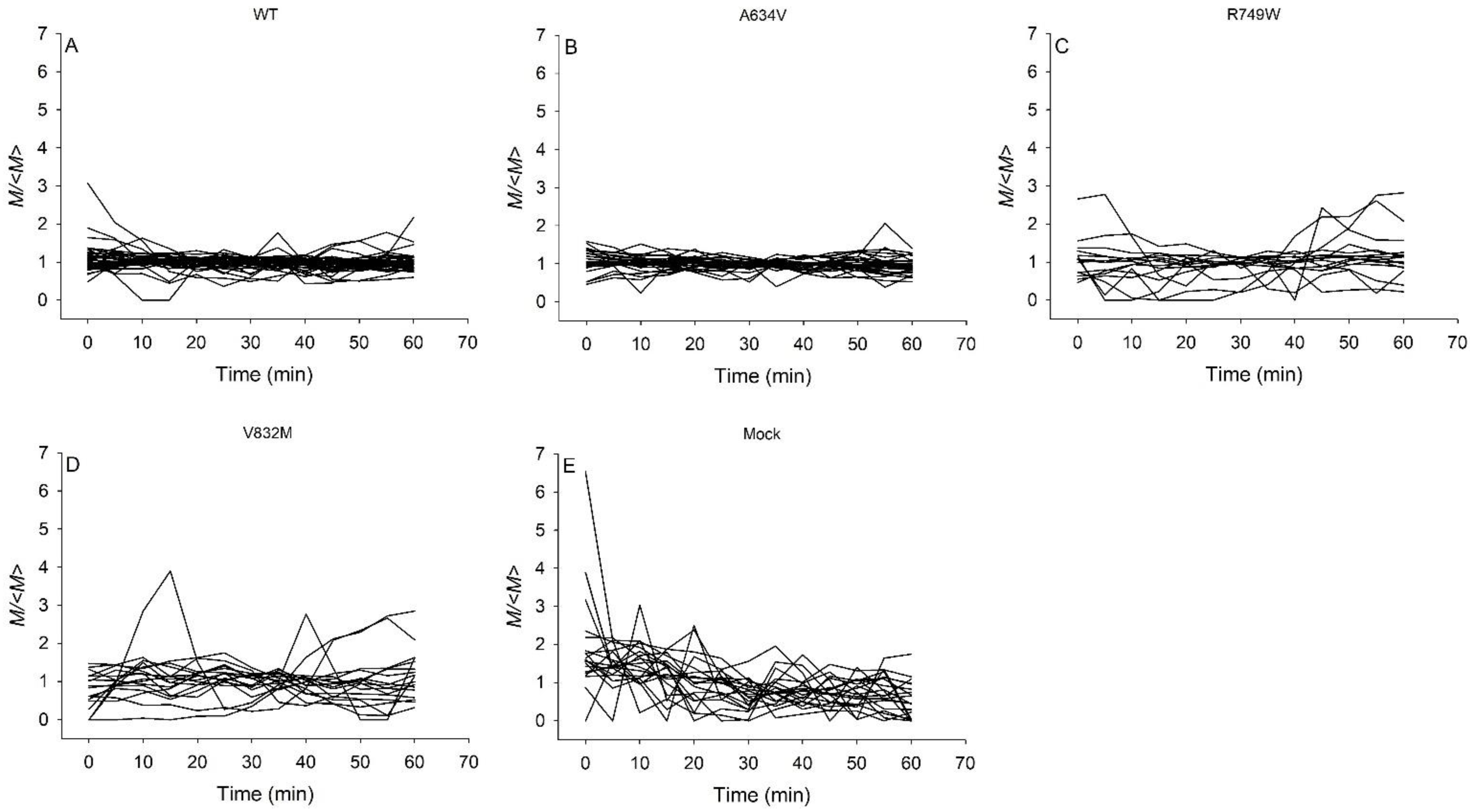{kind=link}
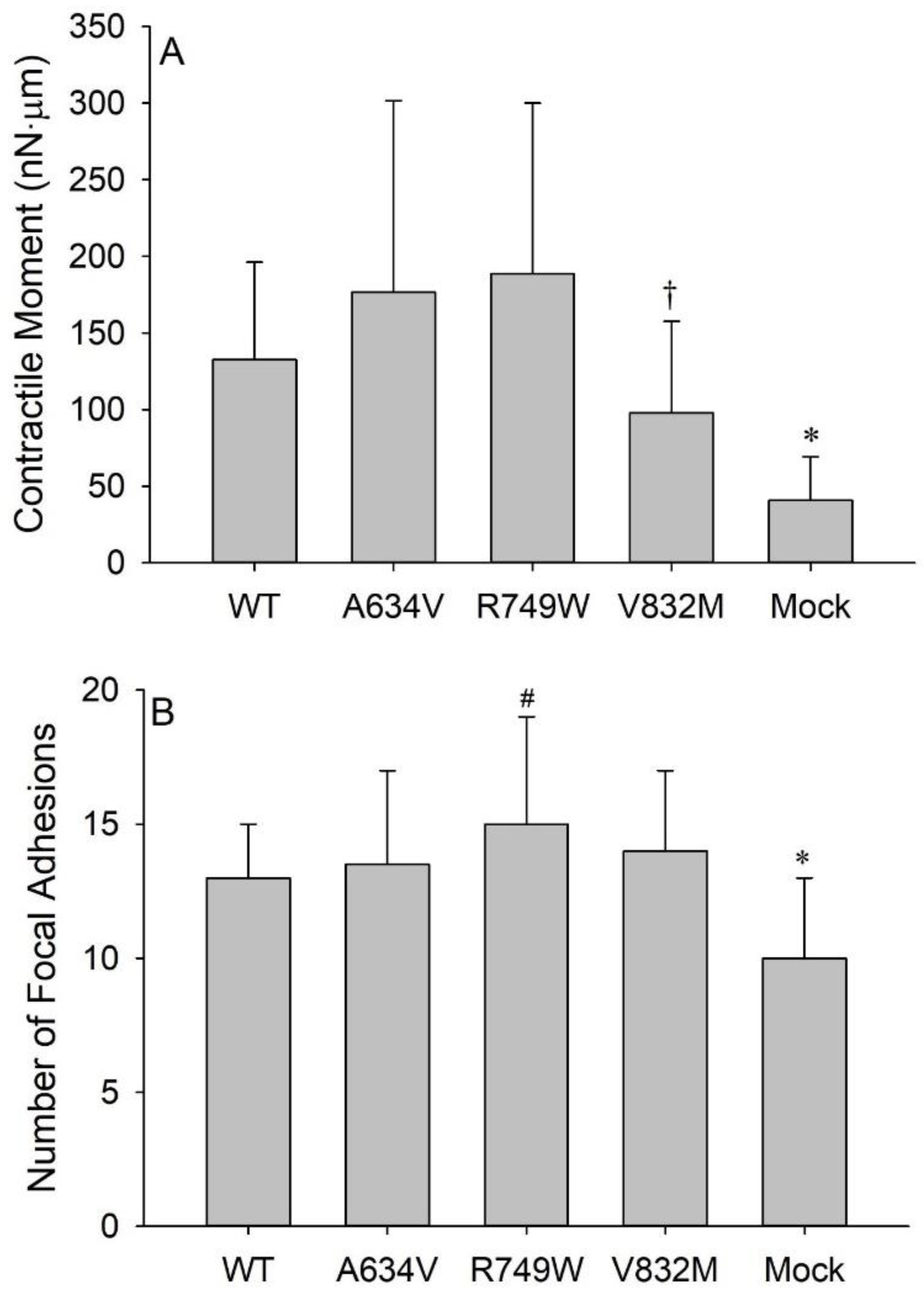{kind=link}
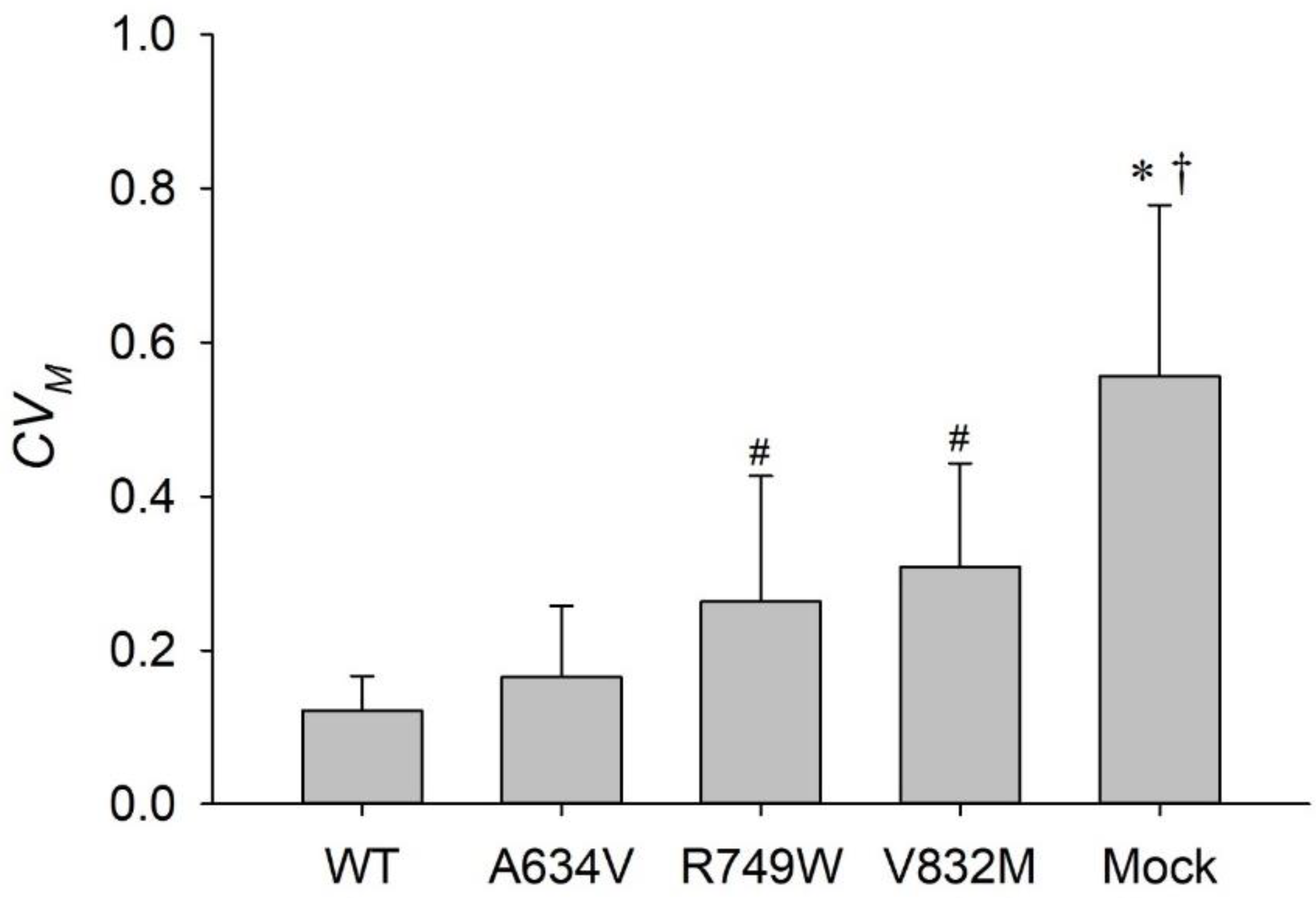{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Transfections
2.2. Micropattern Traction Microscopy
2.3. Contractile Moment and Tension
2.4. Data Analysis
2.5. Quantitative Metrics of Tensional Homeostasis
2.6. Statistical Analysis
3. Results
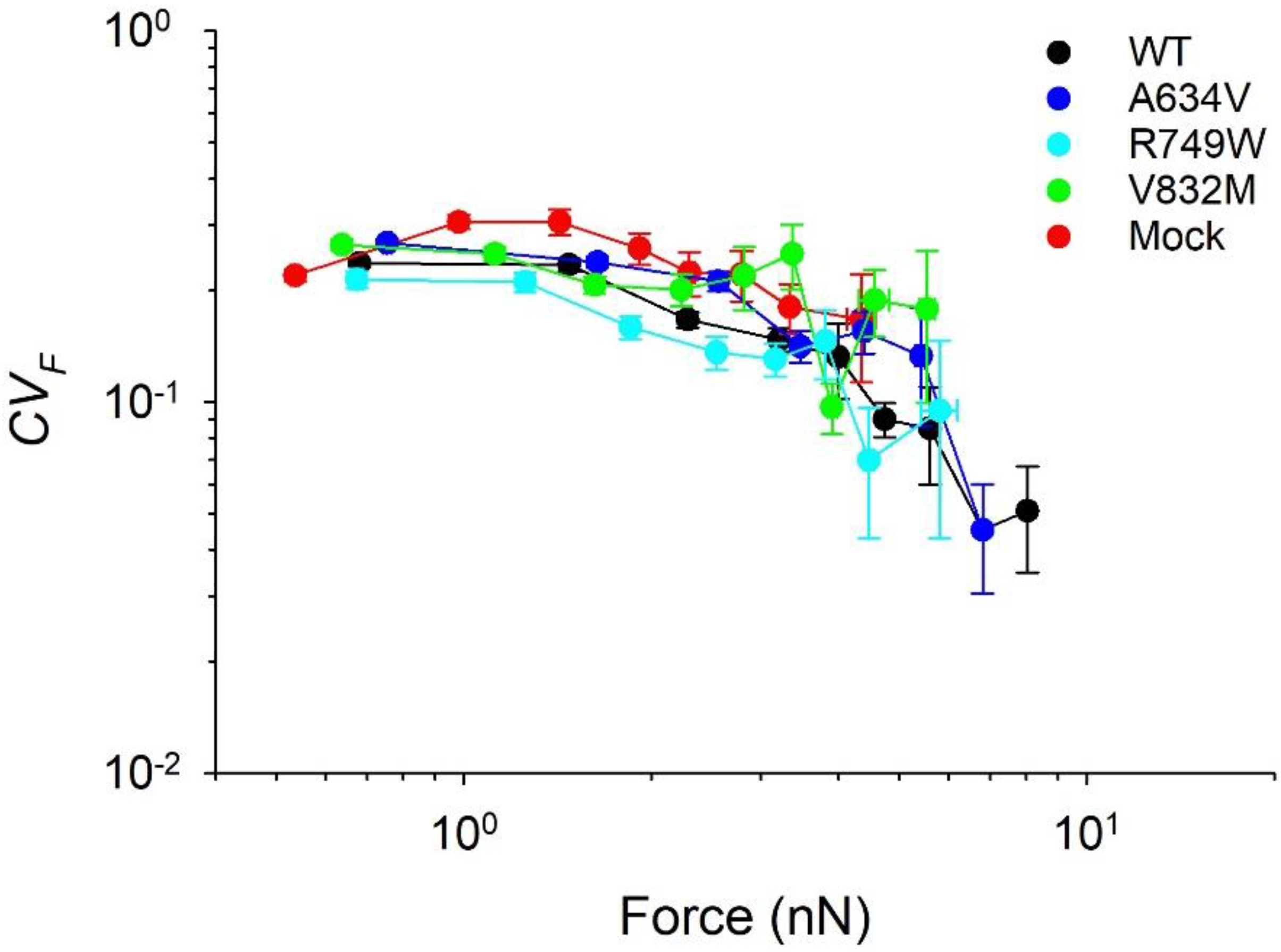
3.1. Contractile Moments of Juxtamembrane, Intracellular E-Cadherin Mutants, and Mock Cells Exhibit Greater Temporal Variability than Wild-Type Cells and Extracellular Mutants
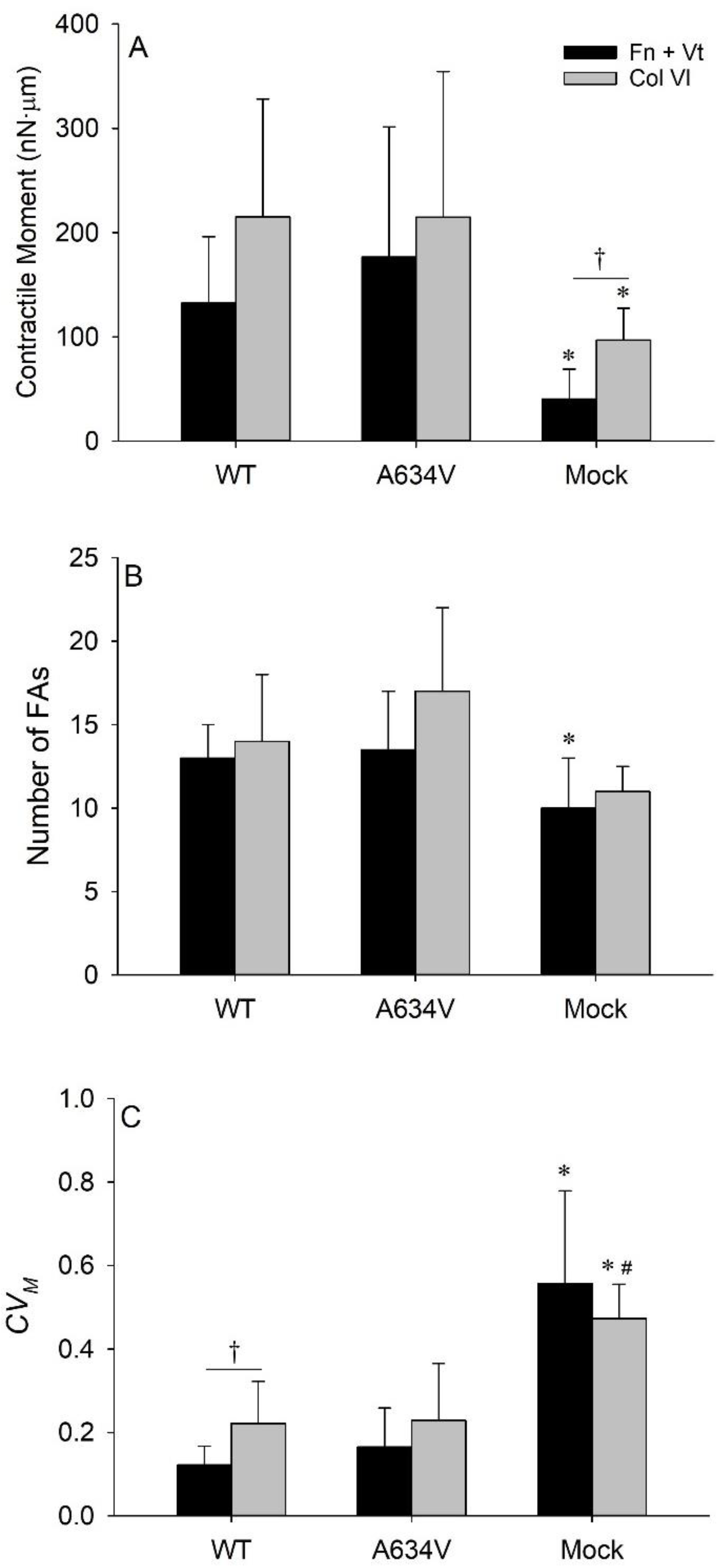
3.2. E-Cadherin Expression Promotes Cell Tension
3.3. Juxtamembrane and Intracellular E-Cadherin Mutants Compromise Tensional Homeostasis
3.4. Collagen VI Enhances Traction Field Magnitude and Fluctuations
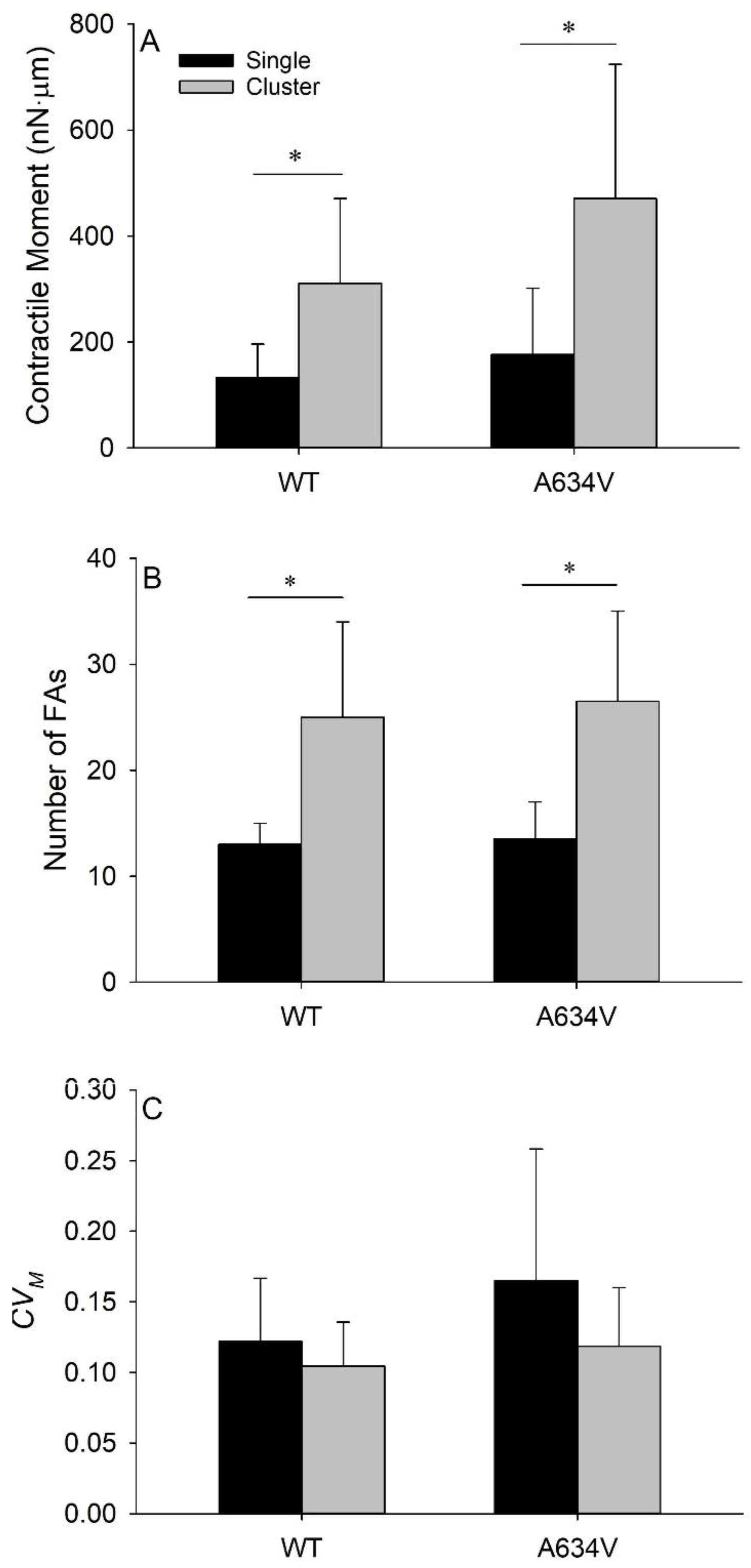
3.5. Extracellular Mutant of E-Cadherin Does Not Affect Tensional Homeostasis in Cell Clusters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chien, S. Mechanotransduction and endothelial cell homeostasis: The wisdom of the cell. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H1209–H1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphrey, J.D. Vascular adaptation and mechanical homeostasis at tissue, cellular, and sub-cellular levels. Cell Biochem. Biophys. 2008, 50, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Stamenović, D.; Smith, M.L. Tensional homeostasis at different length scales. Soft Matter 2020, 16, 6946–6963. [Google Scholar] [CrossRef] [PubMed]
- Paszek, M.J.; Zahir, N.; Johnson, K.R.; Lakins, J.N.; Rozenberg, G.I.; Gefen, A.; Reinhart-King, C.A.; Margulies, S.S.; Dembo, M.; Boettiger, D.; et al. Tensional homeostasis and the malignant phenotype. Cancer Cell 2005, 8, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Butcher, D.T.; Alliston, T.; Weaver, V.M. A tense situation: Forcing tumour progression. Nat. Rev. Cancers 2009, 9, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Mayer, B.; Johnson, J.P.; Leitl, F.; Jauch, K.W.; Heiss, M.M.; Schildberg, F.W.; Birchmeier, W.; Funke, I. E-cadherin expression in primary and metastatic gastric cancer: Down-regulation correlates with cellular dedifferentiation and glandular disintegration. Cancer Res. 1993, 53, 1690–1695. [Google Scholar]
- Figueiredo, J.; Soderberg, O.; Simoes-Correia, J.; Grannas, K.; Suriano, G.; Seruca, R. The importance of E-cadherin binding partners to evaluate the pathogenicity of E-cadherin missense mutations associated to HDGC. Eur. J. Hum. Genet. 2013, 21, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Rübsam, M.; Mertz, A.F.; Kubo, A.; Marg, S.; Jüngst, C.; Goranci-Buzhala, G.; Schauss, A.C.; Horsley, V.; Dufresne, E.R.; Moser, M.; et al. E-cadherin integrates mechanotransduction and EGFR signaling to control junctional tissue polarization and tight junction positioning. Nat. Commun. 2017, 8, 1250. [Google Scholar] [CrossRef]
- DuFort, C.C.; Paszek, M.J.; Weaver, V.M. Balancing forces: Architectural control of mechanotransduction. Nat. Rev. Mol. Cell Biol. 2011, 12, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Lecuit, T.; Yap, A.S. E-cadherin junctions as active mechanical integrators in tissue dynamics. Nat. Cell Biol. 2015, 17, 533–539. [Google Scholar] [CrossRef]
- Paredes, J.; Figueiredo, J.; Albelgaria, A.; Oliviera, P.; Carvalho, J.; Ribeiro, A.S.; Caldeira, J.; Costa, A.M.; Simões-Correia, J.; Oliveira, M.J.; et al. Epithelial E- and P-cadherins: Role and clinical significance in cancer. Biochim. Biophys. Acta 2012, 1826, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodeling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Henke, E.; Nandigama, R.; Ergün, S. Extracellular matrix in the tumor environment and its impact on cancer therapy. Front. Mol. Biosci. 2020, 6, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, M.A.; Desai, R.; Solski, P.A.; Der, C.J.; Keely, P.J. ROCK-generated contractility regulates breast epithelial cell differentiation in response to the physical properties of a three-dimensional collagen matrix. J. Cell Biol. 2003, 163, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, P.P.; Keely, P.J. Mechanical signaling through the cytoskeleton regulates cell proliferation by coordinated focal adhesion and Rho GTPase signaling. J. Cell Sci. 2011, 124, 1195–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, A.M.; Pereira, J.; Melo, S.; Fernandes, M.S.; Carneiro, P.; Seruca, R.; Figueiredo, J. The extracellular matrix: An accomplice in gastric cancer development and progression. Cells 2020, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Webster, K.D.; Ng, W.P.; Fletcher, D.A. Tensional homeostasis in single fibroblasts. Biophys. J. 2014, 107, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Canović, E.P.; Zollinger, A.J.; Tam, S.N.; Smith, M.L.; Stamenović, D. Tensional homeostasis in endothelial cells is a multicellular phenomenon. Am. J. Physiol. Cell Physiol. 2016, 311, C528–C535. [Google Scholar] [CrossRef]
- Weng, S.; Shao, Y.; Chen, W.; Fu, J. Mechanosensitive subcellular rheostasis drives emergent single-cell mechanical homeostasis. Nat. Mater. 2016, 15, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Zollinger, A.J.; Xu, H.; Figueiredo, J.; Paredes, J.; Seruca, R.; Stamenović, D.; Smith, M.L. Dependence of tensional homeostasis on cell type and on cell-cell interactions. Cell. Mol. Bioeng. 2018, 11, 175–184. [Google Scholar] [CrossRef]
- Xu, H.; Donegan, S.; Dreher, J.M.; Stark, A.J.; Canović, E.P.; Stamenović, D.; Smith, M.L. Focal adhesion displacement magnitude is a unifying feature of tensional homeostasis. Acta Biomater. 2020, 113, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.; Ferreira, R.M.; Xu, H.; Gonçalves, M.; Barros-Carvalho, A.; Cravo, J.; Maia, A.F.; Carneiro, P.; Figueiredo, C.; Smith, M.L.; et al. Integrin β1 orchestrates the abnormal cell-matrix attachment and invasive behaviour of E-cadherin dysfunctional cells. Gastric Cancer 2022, 25, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Suriano, G.; Oliveira, C.; Ferreira, P.; Machado, J.C.; Brodin, M.C.; De Wever, O.; Bruyneel, E.A.; Moguilevsky, N.; Grehan, N.; Porter, T.R.; et al. Identification of CDH1 germline missense mutations associated with functional inactivation of the E-cadherin protein in young gastric cancer probands. Hum. Mol. Genet. 2003, 12, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Kaurah, P.; MacMillan, A.; Boyd, N.; Senz, J.; De Luca, A.; Chun, N.; Suriano, G.; Zaor, S.; Van Manen, L.; Gilpin, C.; et al. Founder and recurrent CDH1 mutations in families with hereditary diffuse gastric cancer. JAMA 2007, 297, 2360–2372. [Google Scholar] [CrossRef] [Green Version]
- More, H.; Humar, B.; Weber, W.; Ward, R.; Christian, A.; Lintott, C.; Graziano, F.; Ruzzo, A.-M.; Acosta, E.; Boman, B.; et al. Identification of seven novel germline mutations in the human E-cadherin (CDH1) gene. Hum. Mutat. 2007, 28, 203. [Google Scholar] [CrossRef]
- Simões-Correia, J.; Figueiredo, J.; Oliveira, C.; van Hengel, J.; Seruca, R.; van Roy, F.; Suriano, G. Endoplasmic reticulum quality control: A new mechanism of E-cadherin regulation and its implication in cancer. Hum. Mol. Genet. 2008, 17, 3566–3576. [Google Scholar] [CrossRef] [Green Version]
- Polio, S.R.; Rothenberg, K.E.; Stamenović, D.; Smith, M.L. A micropatterning and image processing approach to simplify measurement of cellular traction forces. Acta Biomater. 2012, 8, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Polio, S.R.; Parameswaran, H.; Canović, E.P.; Gaut, C.M.; Aksyonova, D.; Stamenović, D.; Smith, M.L. Topographical control of multiple cell adhesion molecules for traction force microscopy. Integr. Biol. 2014, 6, 357–365. [Google Scholar] [CrossRef]
- Maloney, J.M.; Walton, E.B.; Bruce, C.M.; Van Vliet, K.J. Influence of finite thickness and stiffness on cellular adhesion-induced deformation of compliant substrata. Phys. Rev. E 2008, 78, 041923. [Google Scholar] [CrossRef] [Green Version]
- Butler, J.P.; Tolić-Nørrelykke, I.M.; Fabry, B.; Fredberg, J.J. Traction fields, moments, and strain energy that cells exert on their surroundings. Am. J. Physiol. Cell Physiol. 2002, 282, C595–C605. [Google Scholar] [CrossRef] [Green Version]
- Canović, E.P.; Seidl, D.T.; Polio, S.R.; Oberai, A.A.; Barbone, P.E.; Stamenović, D.; Smith, M.L. Biomechanical imaging of cell stiffness and prestress with subcellular resolution. Biomech. Model. Mechanobiol. 2014, 13, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Torday, J.S. Homeostasis as the mechanism of evolution. Biology 2015, 4, 573–590. [Google Scholar] [CrossRef] [PubMed]
- Canel, M.; Serrels, A.; Frame, M.C.; Brunton, V.G. E-cadherin-integrin crosstalk in cancer invasion and metastasis. J. Cell Sci. 2013, 126, 393–401. [Google Scholar] [CrossRef] [Green Version]
- Mangold, S.; Norwood, S.J.; Yasp, A.S.; Collins, B.M. The juxtamembrane domain of the E-cadherin cytoplasmic tail contributes to its interaction with myosin VI. Bioarchitecture 2012, 2, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, S.; Correia, J.; Feng, Y.; Figueiredo, J.; Sun, S.X.; Longmore, G.D.; Suriano, G.; Wirtz, D. α-Catenin mediates initial E-cadherin-dependent cell-cell recognition and subsequent bond strengthening. Proc. Natl. Acad. Sci. USA 2008, 105, 18331–18336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suriano, G.; Oliveira, M.J.; Huntsman, D.; Mateus, A.R.; Ferreira, P.; Casares, F.; Oliveira, C.; Carneiro, F.; Machado, J.C.; Mareel, M.; et al. E-cadherin germline missense mutations and cell phenotype: Evidence for the independence of cell invasion on the motile capabilities of the cells. Hum. Mol. Genet. 2003, 12, 3007–3016. [Google Scholar] [CrossRef]
- Caldeira, J.; Figueiredo, J.; Brás-Pereira, C.; Carneiro, P.; Moreira, A.M.; Pinto, M.T.; Relvas, J.B.; Carneiro, F.; Barbosa, M.; Casares, F.; et al. E-cadherin-defective gastric cancer cells depend on laminin to survive and invade. Hum. Mol. Genet. 2015, 24, 5891–5900. [Google Scholar] [CrossRef] [Green Version]
- Roca-Cusachs, P.; Gauthier, N.C.; del Rio, A.; Sheetz, M.P. Clustering of α5β1integrins determines adhesion strength whereas αvβ3 and talin enable mechanotransduction. Proc. Natl. Acad. Sci. USA 2009, 106, 16245–16250. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Bunde, K.A.; Figueiredo, J.; Seruca, R.; Smith, M.L.; Stamenović, D. Differential Impacts on Tensional Homeostasis of Gastric Cancer Cells Due to Distinct Domain Variants of E-Cadherin. Cancers 2022, 14, 2690. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14112690
Xu H, Bunde KA, Figueiredo J, Seruca R, Smith ML, Stamenović D. Differential Impacts on Tensional Homeostasis of Gastric Cancer Cells Due to Distinct Domain Variants of E-Cadherin. Cancers. 2022; 14(11):2690. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14112690
Chicago/Turabian StyleXu, Han, Katie A. Bunde, Joana Figueiredo, Raquel Seruca, Michael L. Smith, and Dimitrije Stamenović. 2022. "Differential Impacts on Tensional Homeostasis of Gastric Cancer Cells Due to Distinct Domain Variants of E-Cadherin" Cancers 14, no. 11: 2690. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers14112690