
mAb14, a Monoclonal Antibody against Cell Surface PCNA: A Potential Tool for Sezary Syndrome Diagnosis and Targeted Immunotherapy
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of PBMCs from Patients with SS and Healthy Controls
2.2. Isolation and Culture of Primary Human Natural Killer (NK) Cells
2.3. Antibodies
2.4. Immunofluorescence Staining of csPCNA and nPCNA
2.5. Imaging Flow Cytometry for csPCNA and nPCNA
2.6. Flow Cytometry for csPCNA and nPCNA
2.7. Apoptosis Assay
2.8. Cell Cycle Analysis of Cells Expressing csPCNA (mAb14+)
2.9. G2/M Cell Cycle Arrest by Nocodazole
2.10. Flow Cytometry for csPCNA Detection in Human Blood Samples
2.11. Hematoxylin and Eosin (H&E) Staining of CD4+CD26- mAb14+ Peripheral Lymphocytes of Patients with SS
2.12. NK Stimulation and IFNγ Assay
2.13. NK-Cell Cytotoxic Activity
2.14. Statistical Analysis
3. Results
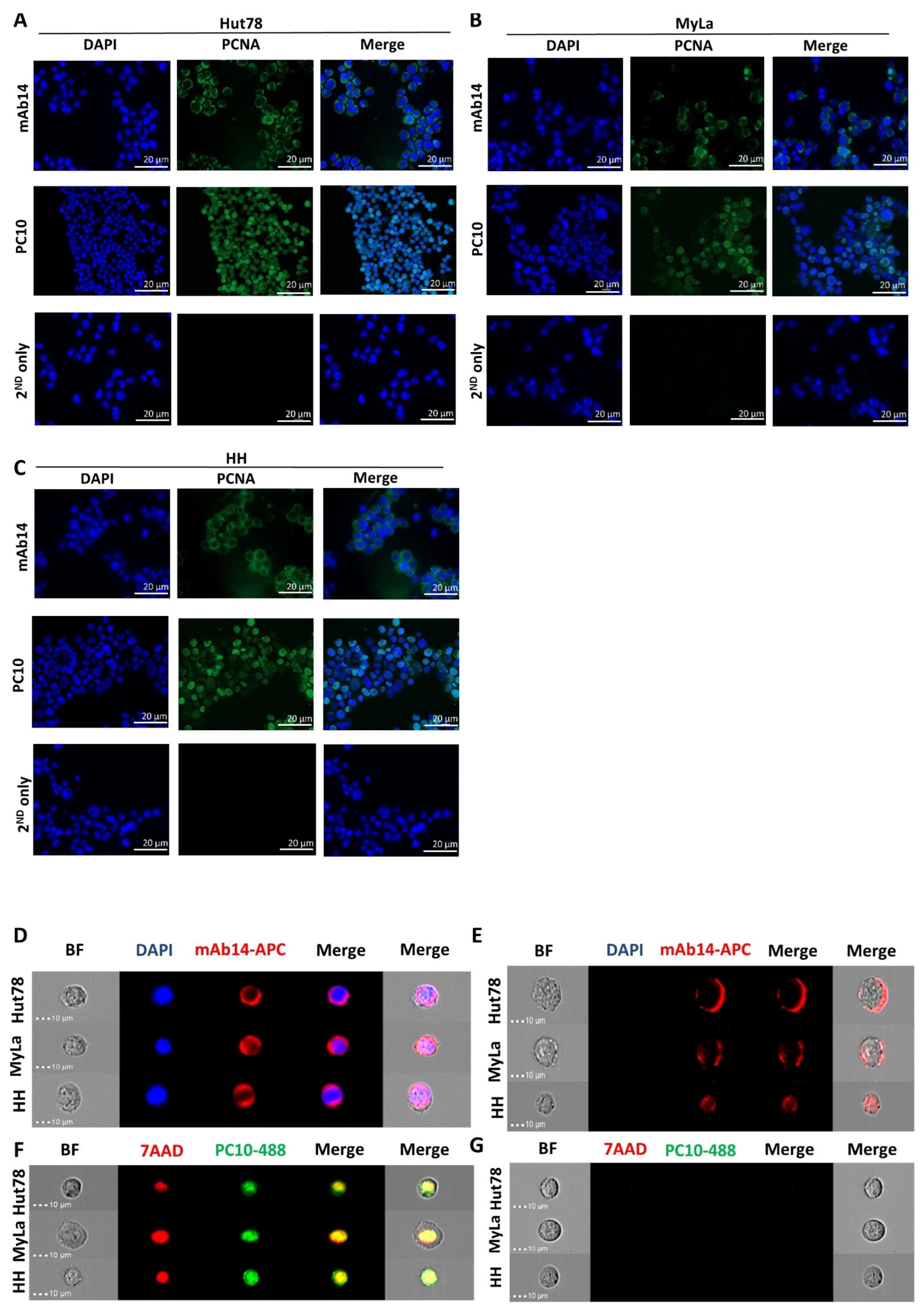
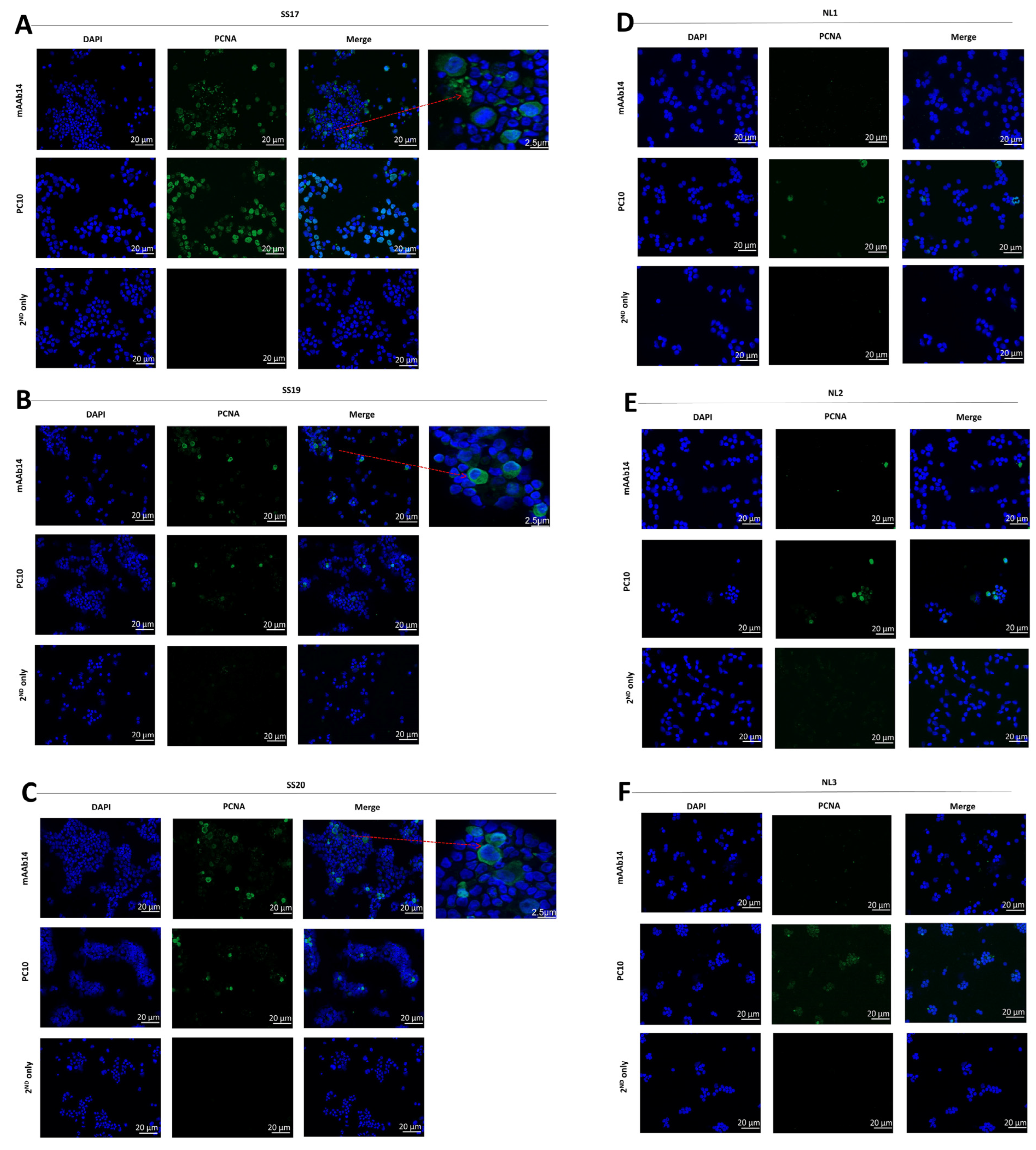
3.1. mAb14 Detects PCNA on the Membrane and in the Cytoplasm of CTCL Cell Lines
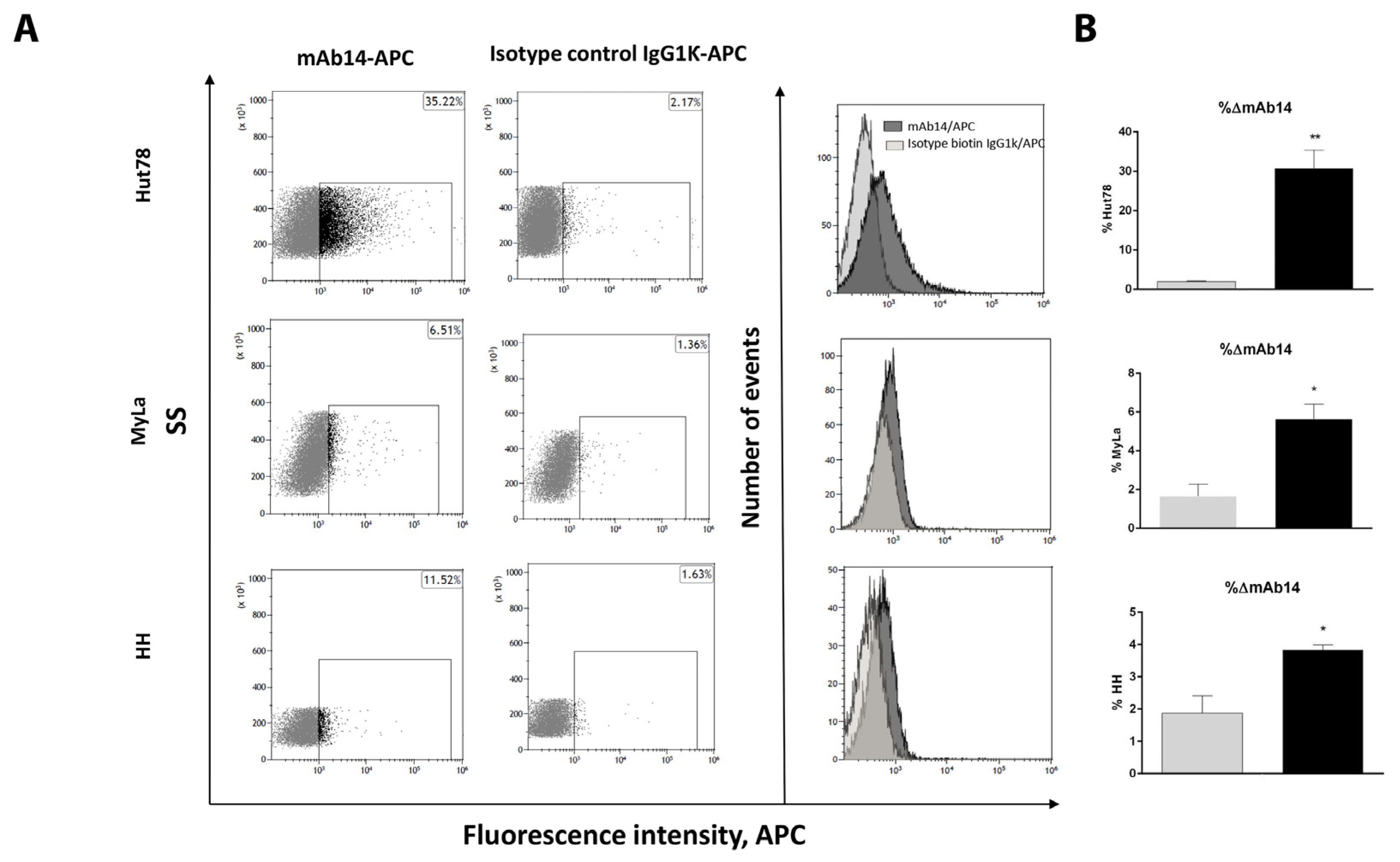
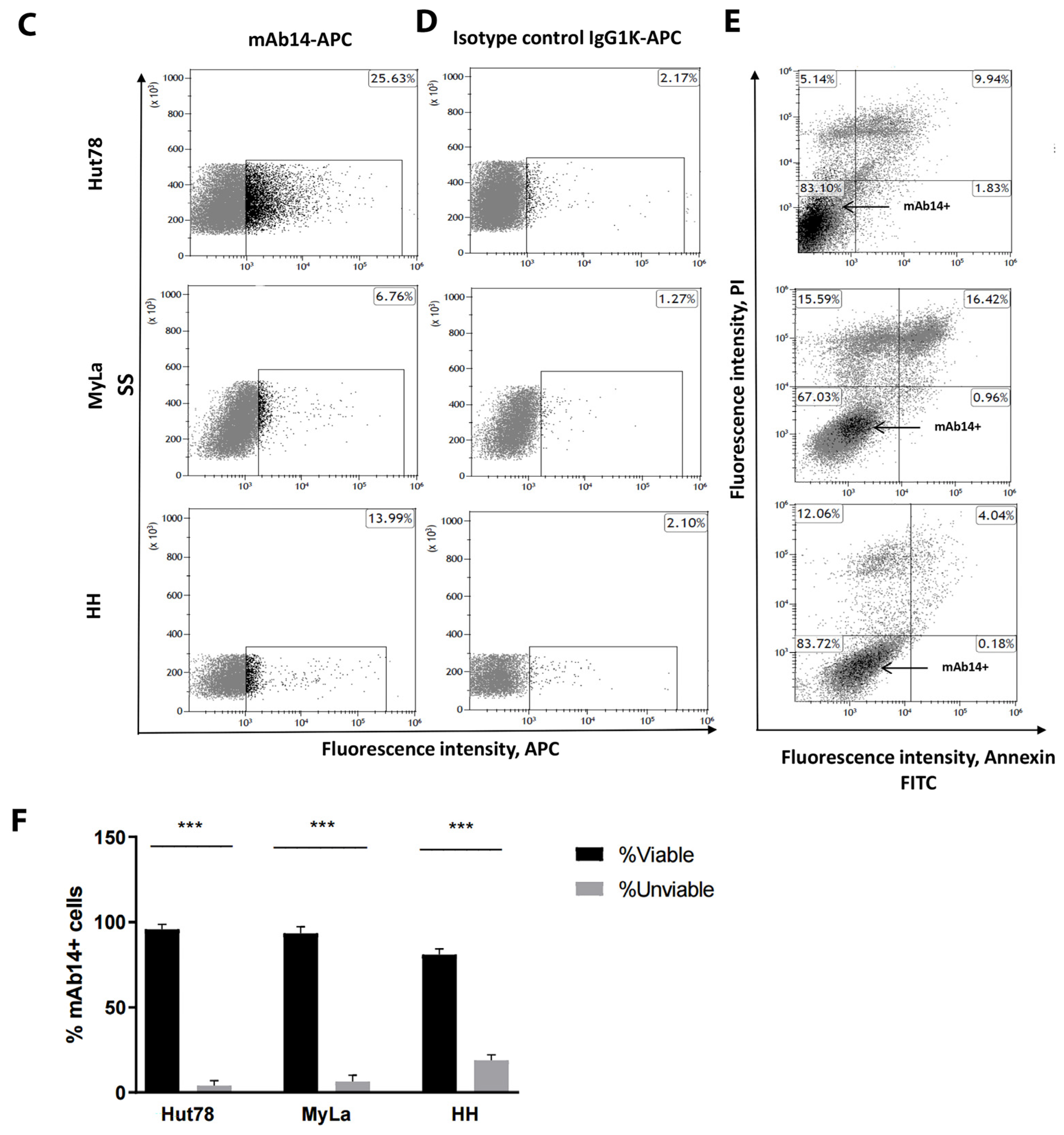
3.2. csPCNA Is Expressed in a Subpopulation of Viable CTCL Cell Lines whereas nPCNA Is Expressed in Most of the Cells
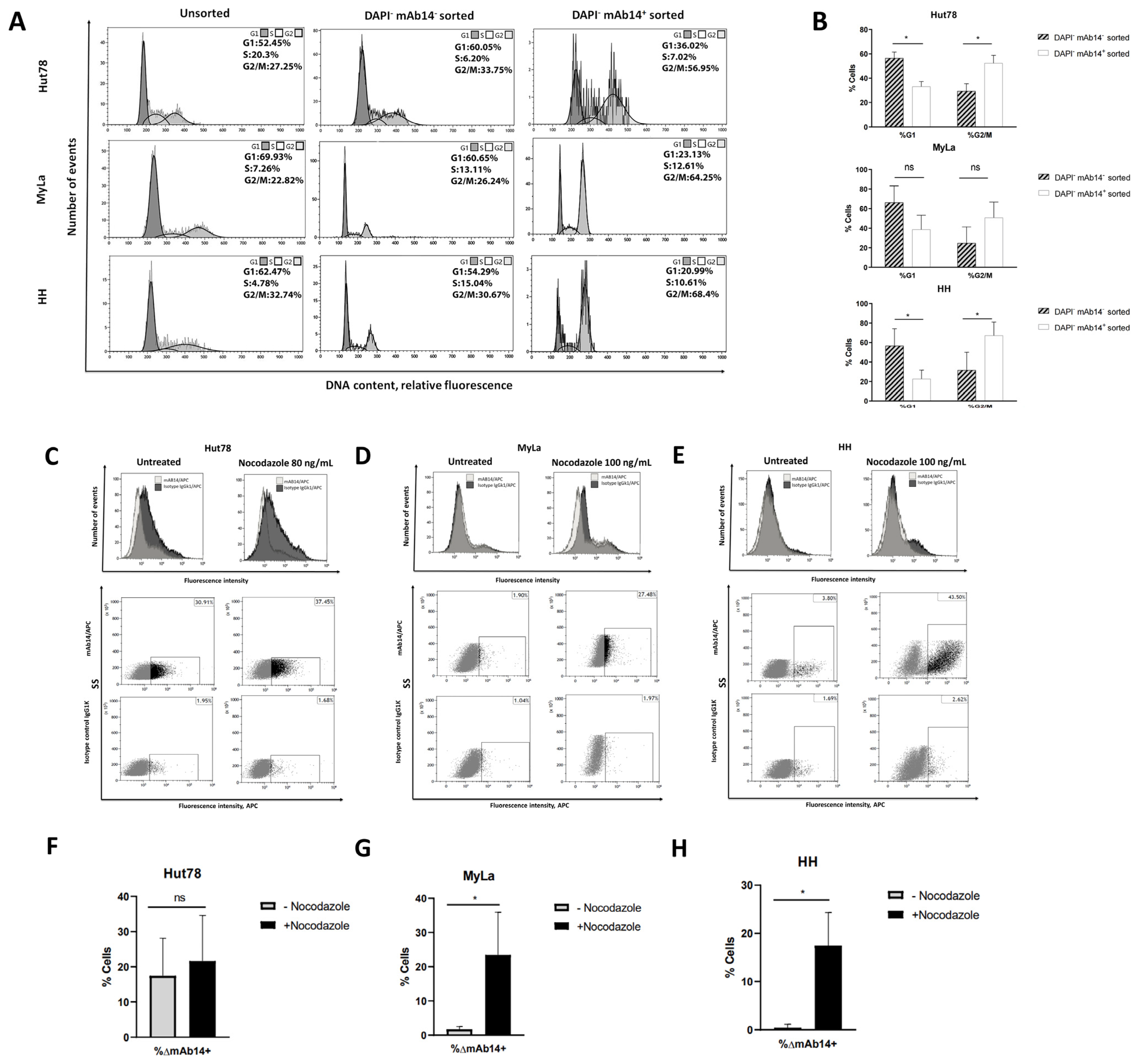
3.3. csPCNA Is Mostly Expressed in CTCL Cells at G2/M Phase
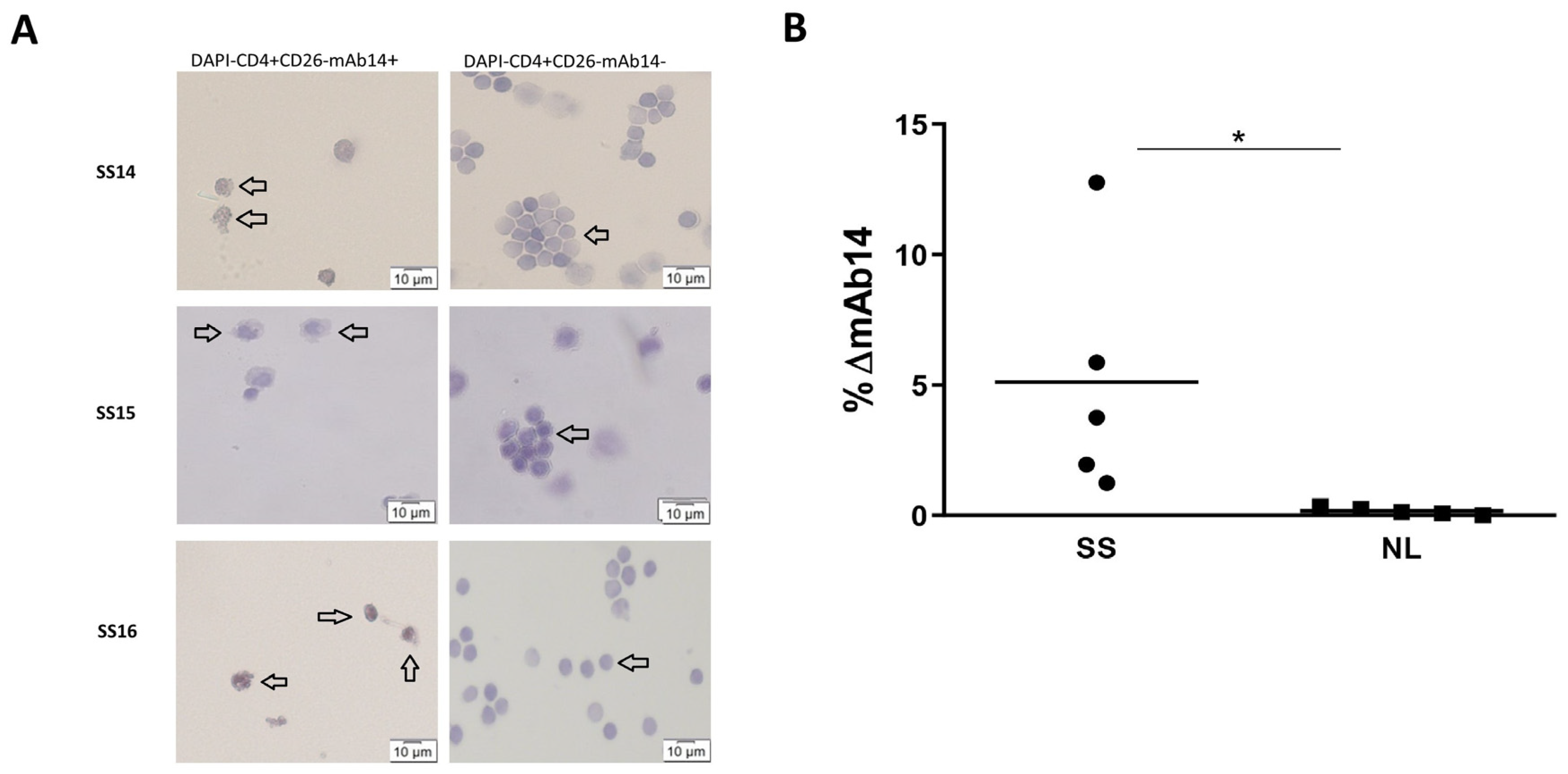
3.4. Viable CD4+CD26−mAb14+ Peripheral Lymphocytes of Patients with Sezary Exhibit Atypical Morphology
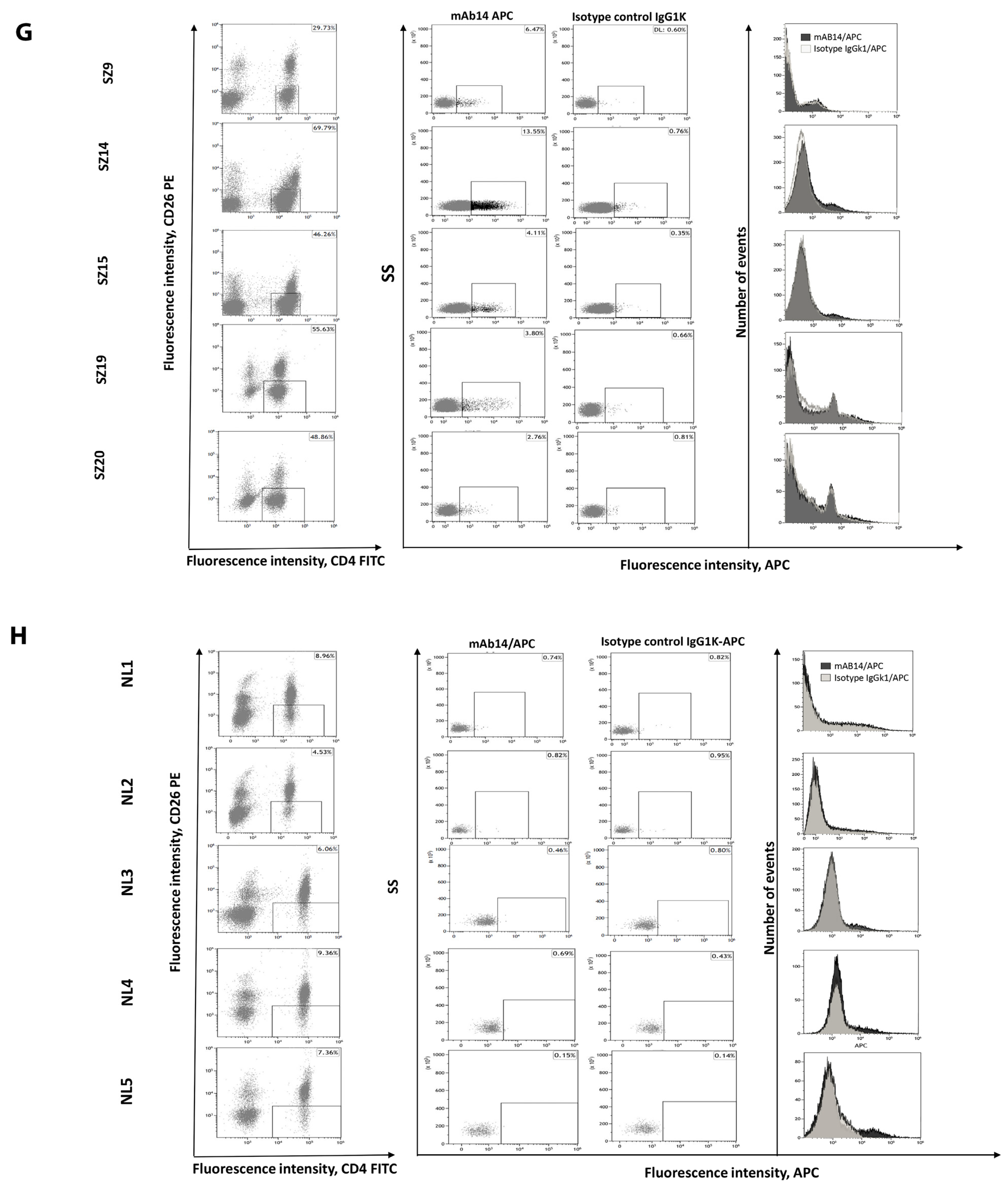
3.5. csPCNA Is Expressed in PBMCs from Patients with Sezary but Not PBMCs from Healthy Donors
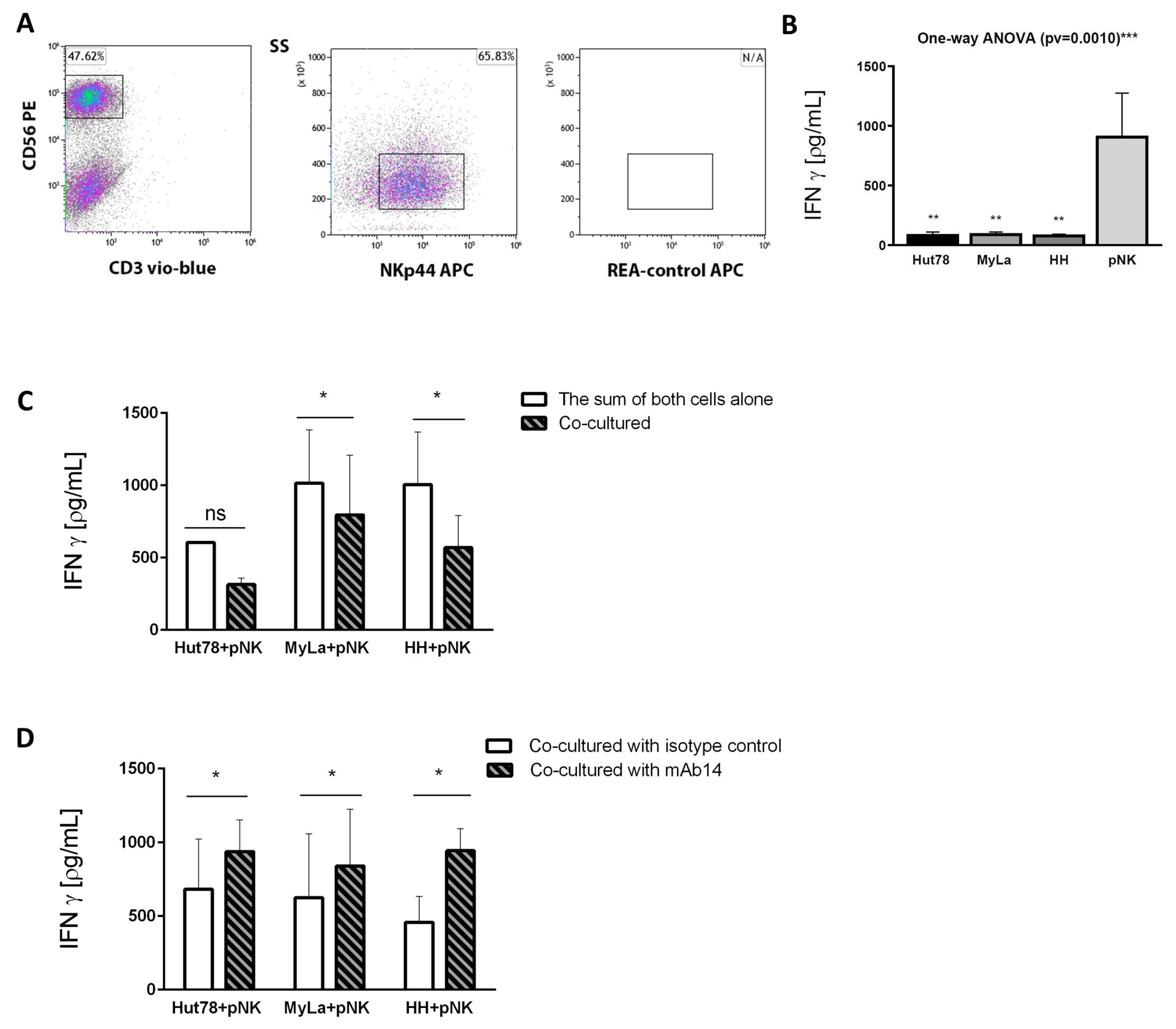
3.6. mAb14 Activates IFN-γ Secretion from Effector Primary NK Cells Co-Cultured with CTCL Cell Lines
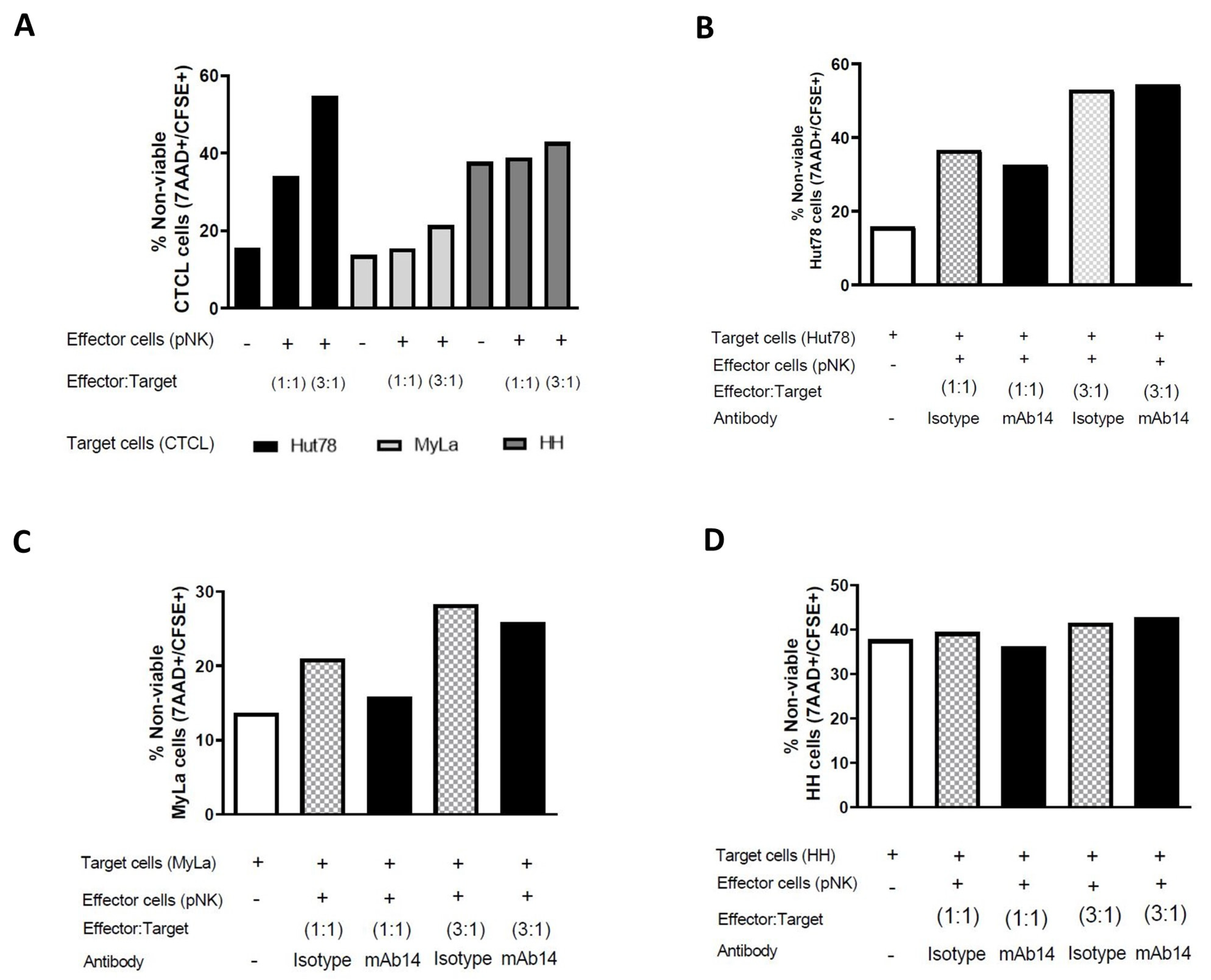
3.7. mAb14 Did Not Enhance the Cytotoxic Activity of pNK Cells against CTCL Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, J.J.; Clark, R.A.; Watanabe, R.; Kupper, T.S. Sezary syndrome and mycosis fungoides arise from distinct T-cell subsets: A biologic rationale for their distinct clinical behaviors. Blood 2010, 116, 767–771. [Google Scholar] [CrossRef]
- Rodd, A.L.; Ververis, K.; Karagiannis, T.C. Current and Emerging Therapeutics for Cutaneous T-Cell Lymphoma: Histone Deacetylase Inhibitors. Lymphoma 2012, 2012, 290685. [Google Scholar] [CrossRef]
- Sidiropoulos, K.G.; Martinez-Escala, M.E.; Yelamos, O.; Guitart, J.; Sidiropoulos, M. Primary cutaneous T-cell lymphomas: A review. J. Clin. Pathol. 2015, 68, 1003–1010. [Google Scholar] [CrossRef]
- Kirsch, I.; Watanabe, R.; O’Malley, J.T.; Williamson, D.W.; Scott, L.L.; Elco, C.P.; Teague, J.E.; Gehad, A.; Lowry, E.L.; LeBoeuf, N.R.; et al. TCR sequencing facilitates diagnosis and identifies mature T cells as the cell of origin in CTCL. Sci. Transl. Med. 2015, 7, 308ra158. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.F.; Newland, K.; McCormack, C.; Lade, S.; Prince, H.M. Mycosis fungoides and Sézary syndrome: Current challenges in assessment, management and prognostic markers. Australas. J. Dermatol. 2016, 57, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.T.; Kollnberger, S.D.; Wedderburn, L.R.; Bowness, P. Expansion and enhanced survival of natural killer cells expressing the killer immunoglobulin-like receptor KIR3DL2 in spondylarthritis. Arthritis Rheum. 2005, 52, 3586–3595. [Google Scholar] [CrossRef]
- Bahler, D.W.; Hartung, L.; Hill, S.; Bowen, G.M.; Vonderheid, E.C. CD158k/KIR3DL2 is a useful marker for identifying neoplastic T-cells in Sézary syndrome by flow cytometry. Cytom. Part B Clin. Cytom. 2008, 74, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Ortonne, N.; Le Gouvello, S.; Mansour, H.; Poillet, C.; Martin, N.; Delfau-Larue, M.H.; Leroy, K.; Farcet, J.P.; Bagot, M.; Bensussan, A. CD158K/KIR3DL2 transcript detection in lesional skin of patients with erythroderma is a tool for the diagnosis of Sezary syn-drome. J. Investig. Dermatol. 2008, 128, 465–472. [Google Scholar] [CrossRef]
- Wechsler, J.; Bagot, M.; Nikolova, M.; Parolini, S.; Martin-Garcia, N.; Boumsell, L.; Moretta, A.; Bensussan, A. Killer cell immunoglobu-lin-like receptor expression delineates in situ Sezary syndrome lymphocytes. J. Pathol. 2003, 199, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Ortonne, N.; Le Gouvello, S.; Tabak, R.; Marie-Cardine, A.; Setiao, J.; Berrehar, F.; Nghe-Tang, A.; Martin, N.; Bagot, M.; Bensussan, A. CD158k/KIR3DL2 and NKp46 are frequently expressed in transformed mycosis fungoides. Exp. Dermatol. 2012, 21, 461–463. [Google Scholar] [CrossRef]
- Bagot, M.; Moretta, A.; Sivori, S.; Biassoni, R.; Cantoni, C.; Bottino, C.; Boumsell, L.; Bensussan, A. CD4+ cutaneous T-cell lymphoma cells express the p140–killer cell immunoglobulin-like receptor. Blood 2001, 97, 1388–1391. [Google Scholar] [CrossRef]
- Poszepczynska-Guigné, E.; Schiavon, V.; D’Incan, M.; Echchakir, H.; Musette, P.; Ortonne, N.; Boumsell, L.; Moretta, A.; Bensussan, A.; Bagot, M. CD158k/KIR3DL2 is a new phenotypic marker of Sezary cells: Relevance for the diagnosis and follow-up of Sezary syndrome. J. Investig. Dermatol. 2004, 122, 820–823. [Google Scholar] [CrossRef]
- Battistella, M.; Leboeuf, C.; Ram-Wolff, C.; Hurabielle, C.; Bonnafous, C.; Sicard, H.; Bensussan, A.; Bagot, M.; Janin, A. KIR3DL2 expression in cutaneous T-cell lymphomas: Expanding the spectrum for KIR3DL2 targeting. Blood 2017, 130, 2900–2902. [Google Scholar] [CrossRef]
- Hurabielle, C.; Thonnart, N.; Ram-Wolff, C.; Sicard, H.; Bensussan, A.; Bagot, M.; Marie-Cardine, A. Usefulness of KIR3DL2 to Diagnose, Follow-Up, and Manage the Treatment of Patients with Sézary Syndrome. Clin. Cancer Res. 2017, 23, 3619–3627. [Google Scholar] [CrossRef]
- Fujii, K. New Therapies and Immunological Findings in Cutaneous T-Cell Lymphoma. Front. Oncol. 2018, 8, 198. [Google Scholar] [CrossRef]
- Van de Donk, N.W.; Dhimolea, E. Brentuximab vedotin. mAbs 2012, 4, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Withers, D.R.; Gaspal, F.M.; Bekiaris, V.; McConnell, F.M.; Kim, M.; Anderson, G.; Lane, P.J. OX40 and CD30 signals in CD4+ T-cell effector and memory function: A distinct role for lymphoid tissue inducer cells in maintaining CD4+ T-cell memory but not effector function. Immunol. Rev. 2011, 244, 134–148. [Google Scholar] [CrossRef]
- Tchernev, G.; Cardoso, J.C.; Arseniev, L.; Okamoto, H. Extrinsic apoptotic pathways: A new potential “target” for more sufficient therapy in a case of cutaneous anaplastic large cd30+ alk-T-cell lymphoma. Indian J. Dermatol. 2021, 56, 87–91. [Google Scholar]
- Barberio, E.; Thomas, L.; Skowron, F.; Balme, B.; Dalle, S. Transformed mycosis fungoides: Clinicopathological features and outcome. Br. J. Dermatol. 2007, 157, 284–289. [Google Scholar] [CrossRef]
- Pulitzer, M.; Myskowski, P.L.; Horwitz, S.M.; Querfeld, C.; Connolly, B.; Li, J.; Murali, R. Mycosis fungoides with large cell transformation: Clinicopathological features and prognostic factors. Pathology 2014, 46, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Edinger, J.T.; Clark, B.Z.; Pucevich, B.E.; Geskin, L.J.; Swerdlow, S.H. CD30 expression and proliferative fraction in nontransformed mycosis fungoides. Am. J. Surg. Pathol. 2009, 33, 1860–1868. [Google Scholar] [CrossRef]
- Benner, M.F.; Jansen, P.M.; Vermeer, M.H.; Willemze, R. Prognostic factors in transformed mycosis fungoides: A retrospective analysis of 100 cases. Blood 2011, 119, 1643–1649. [Google Scholar] [CrossRef]
- Michel, L.; Jean-Louisc, F.; Begue, E.; Bensussan, A.; Bagot, M. Use of PLS3, Twist, CD158k/KIR3DL2, and NKp46 gene expression combination for reliable Sézary syndrome diagnosis. Blood 2013, 121, 8. [Google Scholar] [CrossRef] [PubMed]
- Aliahmad, P.; Kaye, J. Development of all CD4 T lineages requires nuclear factor TOX. J. Exp. Med. 2008, 205, 245–256. [Google Scholar] [CrossRef]
- Dulmage, B.O.; Akilov, O.; Vu, J.R.; Falo, L.D.; Geskin, L.J. Dysregulation of the TOX-RUNX3 pathway in cutaneous T-cell lymphoma. Oncotarget 2019, 10, 3104–3113. [Google Scholar] [CrossRef]
- Huang, Y.; Litvinov, I.V.; Wang, Y.; Su, M.W.; Tu, P.; Jiang, X.; Kupper, T.S.; Dutz, J.P.; Sasseville, D.; Zhou, Y. Thymocyte selection-associated high mobility group box gene (TOX) is aberrantly over-expressed in mycosis fungoides and correlates with poor prognosis. Oncotarget 2014, 5, 4418–4425. [Google Scholar] [CrossRef] [PubMed]
- Morimura, S.; Sugaya, M.; Suga, H.; Miyagaki, T.; Ohmatsu, H.; Fujita, H.; Asano, Y.; Tada, Y.; Kadono, T.; Sato, S. TOX expression in different subtypes of cutaneous lymphoma. Arch. Dermatol. Res. 2014, 306, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Booken, N.; Gratchev, A.; Utikal, J.; Weiß, C.; Yu, X.; Qadoumi, M.; Schmuth, M.; Sepp, N.; Nashan, D.; Rass, K.; et al. Sézary syndrome is a unique cutaneous T-cell lymphoma as identified by an expanded gene signature including diagnostic marker molecules CDO1 and DNM3. Leukemia 2008, 22, 393–399. [Google Scholar] [CrossRef]
- Hameetman, L.; Van der Fits, L.; Zoutman, W.H.; Jacoba, J.; Luiting, O.; Siegal, G.; Iwan JP de Esch Vermeer, M.; Tensen, C.P. EPHA4 is overexpressed but not functionally active in Sézary syndrome. Oncotarget 2015, 6, 31868. [Google Scholar] [CrossRef] [PubMed]
- Wysocka, M.; Kossenkov, A.; Benoit, B.M.; Troxel, A.B.; Singer, E.; Schaffer, A.; Kim, B.; Dentchev, T.; Nagata, S.; Ise, T.; et al. CD164 and FCRL3 are highly expressed on CD4+CD26-T cells in Sézary syndrome patients. J. Investig. Dermatol. 2014, 134, 229–236. [Google Scholar] [CrossRef]
- Gibson, J.F.; Huang, J.; Liu, K.J.; Carlson, K.R.; Foss, F.; Choi, J.; Edelson, R.; Hussong, J.W.; Mohl, R.; Hill, S.; et al. Cutaneous T-cell lymphoma (CTCL): Current practices in blood assessment and the utility of T-cell receptor (TCR)-Vβ chain restriction. J. Am. Acad. Dermatol. 2016, 74, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulos, A.; Thorpe, S.D.; Cortes, E.; Lachowski, D.; Rice, A.J.; Mykuliak, V.V.; Róg, T.; Lee, D.A.; Hytönen, V.P.; del Río Hernández, A.E. Syndecan-4 tunes cell mechanics by activating the kindlin-integrin-RhoA pathway. Nat. Mater. 2020, 19, 669–678. [Google Scholar] [CrossRef]
- Chung, J.S.; Shiue, L.H.; Duvic, M.; Pandya, A.; Cruz, P.D., Jr.; Ariizumi, K. Sézary syndrome cells overexpress syndecan-4 bearing distinct heparan sulfate moieties that suppress T-cell activation by binding DC-HIL and trapping TGF-β on the cell surface. Blood 2011, 117, 3382–3390. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gao, J.; Zhang, S. Functional and Clinical Characteristics of Cell Adhesion Molecule CADM1 in Cancer. Front. Cell Dev. Biol. 2021, 9, 714298. [Google Scholar] [CrossRef]
- Sasaki, H.; Nishikata, I.; Shiraga, T.; Akamatsu, E.; Fukami, T.; Hidaka, T.; Kubuki, Y.; Okayama, A.; Hamada, K.; Okabe, H.; et al. Overexpression of a cell adhesion molecule, TSLC1, as a possible molecular marker for acute-type adult T-cell leukemia. Blood 2005, 105, 1204–1213. [Google Scholar] [CrossRef]
- Yuki, A.; Shinkuma, S.; Hayashi, R.; Fujikawa, H.; Kato, T.; Homma, E.; Hamade, Y.; Onodera, O.; Matsuoka, M.; Shimizu, H.; et al. CADM1 is a diagnostic marker in early-stage mycosis fungoides: Multicenter study of 58 cases. J. Am. Acad. Dermatol. 2018, 79, 1039–1046. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Morizane, S.; Hamada, T.; Miyake, T.; Sugaya, M.; Iwata, H.; Fujii, K.; Haramoto-Shiratsuki, R.; Nakagawa, Y.; Miura, M.; et al. The expression of cell adhesion molecule 1 and its splicing variants in Sézary cells and cell lines from cutaneous T-cell lymphoma. J. Dermatol. 2019, 46, 967–977. [Google Scholar] [CrossRef]
- Legler, D.F.; Loetscher, M.; Roos, R.S.; Clark-Lewis, I.; Baggiolini, M.; Moser, B. B Cell–attracting Chemokine 1, a Human CXC Chemokine Expressed in Lymphoid Tissues, Selectively Attracts B Lymphocytes via BLR1/CXCR5. J. Exp. Med. 1998, 187, 655–660. [Google Scholar] [CrossRef]
- Wang, L.; Rocas, D.; Dalle, S.; Sako, N.; Pelletier, L.; Martin, N.; Dupuy, A.; Tazi, N.; Balme, B.; Vergier, B.; et al. Primary cutaneous peripheral T-cell lymphomas with a T-follicular helper phenotype: An integrative clinical, pathological and molecular case series study. Br. J. Dermatol. 2022, 187, 970–980. [Google Scholar] [CrossRef]
- Picchio, M.C.; Scala, E.; Pomponi, D.; Caprini, E.; Frontani, M.; Angelucci, I.; Mangoni, A.; Lazzeri, C.; Perez, M.; Remotti, D.; et al. CXCL13 is highly produced by Sezary cells and enhances their migratory ability via a synergistic mechanism involving CCL19 and CCL21 chemokines. Cancer Res. 2008, 68, 7137–7146. [Google Scholar] [CrossRef]
- Bosisio, F.M.; Cerroni, L. Expression of T-follicular helper markers in sequential biopsies of progressive mycosis fungoides and other primary cutaneous T-Cell Lymphomas. Am. J. Dermatopathol. 2015, 37, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Olsen, E.A.; Whittaker, S.; Willemze, R.; Pinter-Brown, L.; Foss, F.; Geskin, L.; Schwartz, L.; Horwitz, S.; Guitart, J.; Zic, J.; et al. Primary cu-taneous lymphoma: Recommendations for clinical trial design and staging update from the ISCL, USCLC, and EORTC. Blood 2022, 140, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Rosental, B.; Brusilovsky, M.; Hadad, U.; Oz, D.; Appel, M.Y.; Afergan, F.; Yossef, R.; Rosenberg, L.A.; Aharoni, A.; Cerwenka, A.; et al. Proliferating cell nuclear antigen is a novel inhibitory ligand for the natural cytotoxicity receptor NKp44. J. Immunol. 2011, 187, 5693–5702. [Google Scholar] [CrossRef] [PubMed]
- Krishna, T.S.; Kong, X.-P.; Gary, S.; Burgers, P.M.; Kuriyan, J. Crystal structure of the eukaryotic DNA polymerase processivity factor PCNA. Cell 1994, 79, 1233–1243. [Google Scholar] [CrossRef]
- Moretta, A.; Biassoni, R.; Bottino, C.; Mingari, M.C.; Moretta, L. Natural cytotoxicity receptors that trigger human NK-cell-mediated cytolysis. Immunol. Today 2000, 21, 228–234. [Google Scholar] [CrossRef]
- Koch, J.; Steinle, A.; Watzl, C.; Mandelboim, O. Activating natural cytotoxicity receptors of natural killer cells in cancer and infection. Trends Immunol. 2013, 34, 182–191. [Google Scholar] [CrossRef]
- Kruse, P.H.; Matta, J.; Ugolini, S.; Vivier, E. Natural cytotoxicity receptors and their ligands. Immunol. Cell Biol. 2013, 92, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non–major histocompatibility complex–restricted Tumor Cell Lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef]
- Hershkovitz, O.; Jivov, S.; Bloushtain, N.; Zilka, A.; Landau, G.; Bar-Ilan, A.; Lichtenstein, R.G.; Campbell, K.S.; van Kuppevelt, T.H.; Porgador, A. Characterization of the Recognition of Tumor Cells by the Natural Cytotoxicity Receptor, NKp44. Biochemistry 2007, 46, 7426–7436. [Google Scholar] [CrossRef]
- Wood, G.S.; Dubiel, C.; Mueller, C.; Abel, E.A.; Hoppe, R.T.; Edinger, A.; Weissman, I.; A Warnke, R. Most CD8+ cells in skin lesions of CD3+ CD4+ mycosis fungoides are CD3+ T cells that lack CD11b, CD16, CD56, CD57, and human Hanukah factor mRNA. Am. J. Pathol. 1991, 138, 1545–1552. [Google Scholar]
- Bouaziz, J.-D.; Ortonne, N.; Giustiniani, J.; Schiavon, V.; Huet, D.; Bagot, M.; Bensussan, A. Circulating Natural Killer Lymphocytes Are Potential Cytotoxic Effectors Against Autologous Malignant Cells in Sezary Syndrome Patients. J. Investig. Dermatol. 2005, 125, 1273–1278. [Google Scholar] [CrossRef] [PubMed]
- Laroche, L.; Papiernik, M.; Bach, J.F. Sezary syndrome. I. Specific skin-directed migration of peripheral blood lymphocytes. J. Immunol. 1983, 130, 2467–2470. [Google Scholar] [CrossRef] [PubMed]
- Kundu, K.; Ghosh, S.; Sarkar, R.; Edri, A.; Brusilovsky, M.; Gershoni-Yahalom, O.; Yossef, R.; Shemesh, A.; Soria, J.-C.; Lazar, V.; et al. Inhibition of the NKp44-PCNA Immune Checkpoint Using a mAb to PCNA. Cancer Immunol. Res. 2019, 7, 1120–1134. [Google Scholar] [CrossRef]
- Woods, A.L.; Hall, P.A.; Shepherd Hanby, A.M.; Waseem, N.H.; Lane, D.P.; Levison, D.A. The assessment of proliferating cell nuclear antigen (PCNA) immunostaining in primary gastrointestinal lymphomas and its relationship to histological grade, S G2 M phase fraction (flow cytometric analysis) and prognosis. Histopathology 1991, 19, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Li, Z.; Huang, J.; Miao, Z.; Zhang, J.; Chunyang, L.; Hao, X.; Huimian, X. Prognostic value and clinicopathological significance of proliferating cell nuclear antigen expressionin gastric cancer: A systematic review and meta-analysis. OncoTargets Ther. 2017, 10, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Naryzhny, S.N.; Lee, H. Proliferating cell nuclear antigen in the cytoplasm interacts with components of glycolysis and cancer. FEBS Lett. 2010, 584, 4292–4298. [Google Scholar] [CrossRef]
- Rosental, B.; Hadad, U.; Brusilovsky, M.; Campbell, K.S.; Porgador, A. A novel mechanism for cancer cells to evade immune attack by NK cells. OncoImmunology 2012, 1, 572–574. [Google Scholar] [CrossRef]
- Stetson, D.B.; Mohrs, M.; Reinhardt, R.L.; Baron, J.L.; Wang, Z.E.; Gapin, L.; Kronenberg, M.; Locksley, R.M. Constitutive cytokine mRNAs mark natural killer (NK) and NK T cells poised for rapid effector function. J. Exp. Med. 2003, 198, 1069–1076. [Google Scholar] [CrossRef]
- Campbell, K.S.; Yusa, S.-I.; Kikuchi-Maki, A.; Catina, T.L. NKp44 Triggers NK Cell activation through DAP12 association that is not influenced by a putative cytoplasmic inhibitory sequence. Perspect. Surg. 2004, 172, 899–906. [Google Scholar] [CrossRef]
- Babiychuk, E.B.; Draeger, A. Annexins in cell membrane dynamics. Ca2+-regulated association of lipid microdomains. J. Cell Biol. 2000, 150, 1113–1124. [Google Scholar] [CrossRef]
- Feng, J.-G.; Liu, Q.; Qin, X.; Geng, Y.-H.; Zheng, S.-T.; Liu, T.; Sheyhidin, I.; Lu, X.-M. Clinicopathological pattern and Annexin A2 and Cdc42 status in patients presenting with differentiation and lymphnode metastasis of esophageal squamous cell carcinomas. Mol. Biol. Rep. 2012, 39, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, S.; Liu, Q.; Liu, T.; Liang, M.; Gao, X.; Lu, M.; Sheyhidin, I.; Lu, X. Under-expression of annexin A2 is associated with Kazakh’s esophageal squamous cell carcinoma. Mol. Carcinog. 2015, 54, 779–788. [Google Scholar] [CrossRef]
- Marrufo, A.M.; Mathew, S.O.; Chaudhary, P.; Malaer, J.D.; Ahmed, N.; Vishwanatha, J.K.; Mathew, P.A. Blocking PCNA interaction with NKp44 enhances primary natural killer cell-mediated lysis of triple-negative breast cancer cells. Am. J. Cancer Res. 2023, 13, 1082–1090. [Google Scholar]
- Moyal, L.; Arkin, C.; Gorovitz-Haris, B.; Querfeld, C.; Rosen, S.; Knaneh, J.; Amitay-Laish, I.; Prag-Naveh, H.; Jacob-Hirsch, J.; Hodak, E. Mycosis fungoides-derived exosomes promote cell motility, are enriched with miR-155 and miR-1246, and their plasma cell-free expression may serve as potential biomarkers for disease burden. Br. J. Dermatol. 2021, 185, 999–1012. [Google Scholar] [CrossRef]
- Moyal, L.; Yehezkel, S.; Gorovitz, B.; Keren, A.; Gilhar, A.; Lubin, I.; Sherman, S.; Hodak, E. Oncogenic role of microRNA-155 in mycosis fungoides: An in vitro and xenograft mouse model study. Br. J. Dermatol. 2017, 177, 791–800. [Google Scholar] [CrossRef]
- Vonderheid, E.C. On the diagnosis of erythrodermic cutaneous T-cell lymphoma. J. Cutan. Pathol. 2006, 33, 27–42. [Google Scholar] [CrossRef]
- Harro, C.M.; Sprenger, K.B.; Chaurio, R.A.; Powers, J.J.; Innamarato, P.; Anadon, C.M.; Zhang, Y.; Biswas, S.; Mandal, G.; Mine, J.A.; et al. Sézary syndrome originates from heavily mutated hematopoietic progenitors. Blood Adv. 2023. [Google Scholar] [CrossRef]
- Bouayad, D.; Pederzoli-Ribeil, M.; Mocek, J.; Candalh, C.; Arlet, J.-B.; Hermine, O.; Reuter, N.; Davezac, N.; Witko-Sarsat, V. Nuclear-to-cytoplasmic relocalization of the proliferating cell nuclear antigen (PCNA) during differentiation involves a chromosome region maintenance 1 (CRM1)-dependent export and is a prerequisite for PCNA antiapoptotic activity in mature neutrophils. J. Biol. Chem. 2012, 287, 33812–33825. [Google Scholar] [CrossRef] [PubMed]
- Gaydosik, A.M.; Tabib, T.; Geskin, L.J.; Bayan, C.-A.; Conway, J.F.; Lafyatis, R.; Fuschiotti, P. Single-cell lymphocyte heterogeneity in advanced cutaneous T-cell lymphoma skin tumors. Clin. Cancer Res. 2019, 25, 4443–4454. [Google Scholar] [CrossRef]
- Vidulich, K.A.; Talpur, R.; Bassett, R.L.; Duvic, M. Overall survival in erythrodermic cutaneous T-cell lymphoma: An analysis of prognostic factors in a cohort of patients with erythrodermic cutaneous T-cell lymphoma. Int. J. Dermatol. 2009, 48, 243–252. [Google Scholar] [CrossRef]
- Novelli, M.; Fava, P.; Sarda, C.; Ponti, R.; Osella-Abate, S.; Savoia, P.; Bergallo, M.; Lisa, F.; Teresa Fierro, M.; Quaglino, P. Blood flow cy-tometry in Se’zary syndrome: New insights on prognostic relevance and immunophenotypic changes during follow-up. Am. J. Clin. Pathol. 2015, 143, 57–69. [Google Scholar] [CrossRef]
- Pichardo, D.A.; Querfeld, C.; Guitart, J.; Kuzel, T.M.; Rosen, S.T. Cutaneous T-cell lymphoma: A paradigm for biological therapies. Leuk. Lymphoma 2004, 45, 1755–1765. [Google Scholar] [CrossRef]
- Netchiporouk, E.; Gantchev, J.; Tsang, M.; Thibault, P.; Watters, A.K.; Matthew Hughes, J.D.; Ghazawi, F.M.; Woetmann, A.; Ødum, N.; Sasseville, D.; et al. Analysis of CTCL cell lines reveals important differences between mycosis fungoides/Sézary sy drome vs. HTLV-1+ leukemic cell lines. Oncotarget 2017, 8, 95981–95998. [Google Scholar] [CrossRef] [PubMed]
- Manfrere, K.C.; Torrealba, M.P.; Miyashiro, D.R.; Pereira, N.Z.; Yoshikawa, F.S.; Oliveira, L.d.M.; Cury-Martins, J.; Duarte, A.J.; Sanches, J.A.; Sato, M.N. Profile of differentially expressed Toll-like receptor signaling genes in the natural killer cells of patients with Sézary syndrome. Oncotarget 2017, 8, 92183–92194. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Herrero, S.; Lopez-Soto, A.; Sordo-Bahamonde, C.; Gonzalez-Rodriguez, A.P.; Vitale, M.; Gonzalez, S. NK cell-based im-munotherapy in cancer metastasis. Cancers 2019, 11, 29. [Google Scholar] [CrossRef]
- Horton, N.C.; Mathew, S.O.; Mathew, P.A. Novel Interaction between Proliferating Cell Nuclear Antigen and HLA I on the Surface of Tumor Cells Inhibits NK Cell Function through NKp44. PLoS ONE 2013, 8, e59552. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knaneh, J.; Hodak, E.; Fedida-Metula, S.; Edri, A.; Eren, R.; Yoffe, Y.; Amitay-Laish, I.; Prag Naveh, H.; Lubin, I.; Porgador, A.; et al. mAb14, a Monoclonal Antibody against Cell Surface PCNA: A Potential Tool for Sezary Syndrome Diagnosis and Targeted Immunotherapy. Cancers 2023, 15, 4421. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers15174421
Knaneh J, Hodak E, Fedida-Metula S, Edri A, Eren R, Yoffe Y, Amitay-Laish I, Prag Naveh H, Lubin I, Porgador A, et al. mAb14, a Monoclonal Antibody against Cell Surface PCNA: A Potential Tool for Sezary Syndrome Diagnosis and Targeted Immunotherapy. Cancers. 2023; 15(17):4421. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers15174421
Chicago/Turabian StyleKnaneh, Jamal, Emmilia Hodak, Shlomit Fedida-Metula, Avishay Edri, Rachel Eren, Yael Yoffe, Iris Amitay-Laish, Hadas Prag Naveh, Ido Lubin, Angel Porgador, and et al. 2023. "mAb14, a Monoclonal Antibody against Cell Surface PCNA: A Potential Tool for Sezary Syndrome Diagnosis and Targeted Immunotherapy" Cancers 15, no. 17: 4421. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers15174421