
The C250T Mutation of TERTp Might Grant a Better Prognosis to Glioblastoma by Exerting Less Biological Effect on Telomeres and Chromosomes Than the C228T Mutation
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Patients
2.2. Molecular Analyses
2.3. Statistical Analyses
2.4. Ethics Statement
3. Results
3.1. Patients
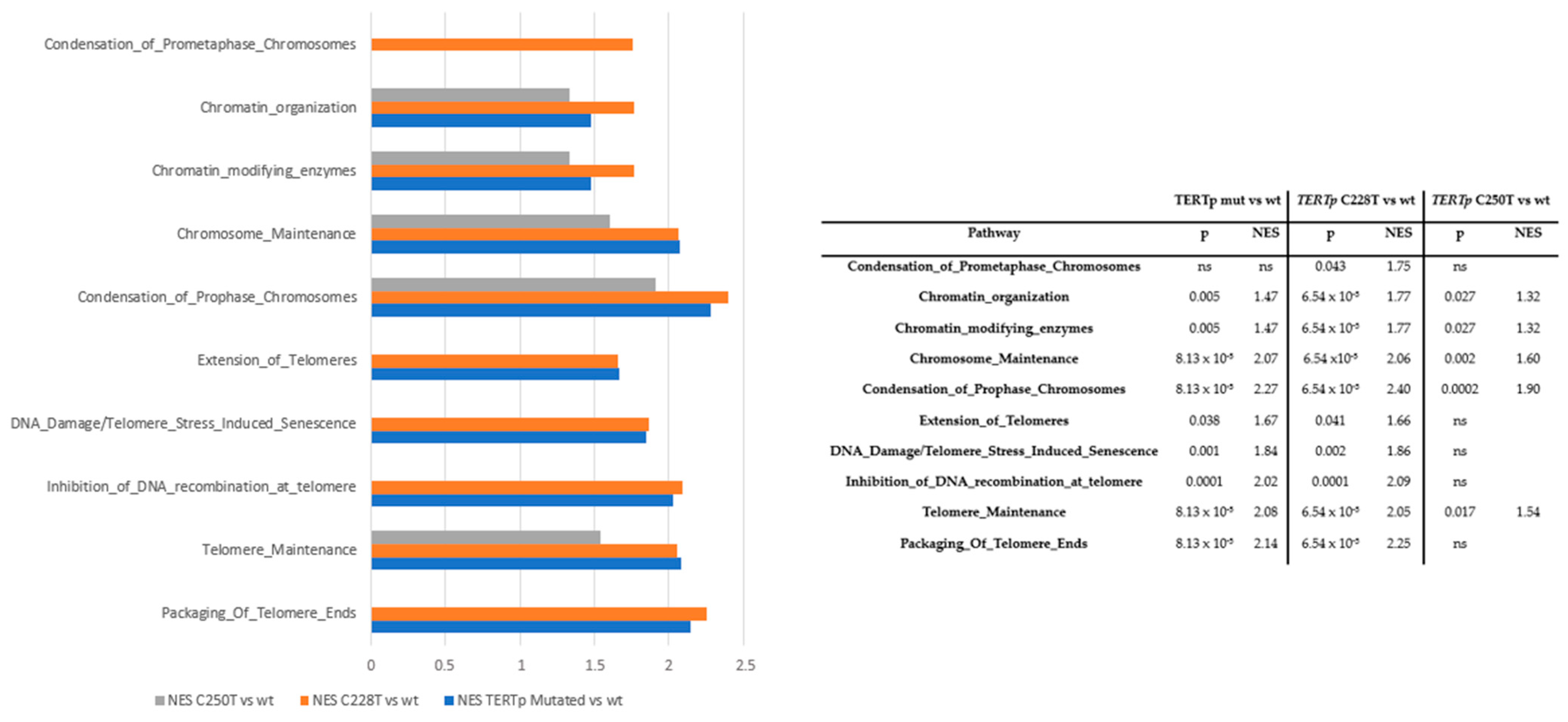
3.2. TERTp Mutations
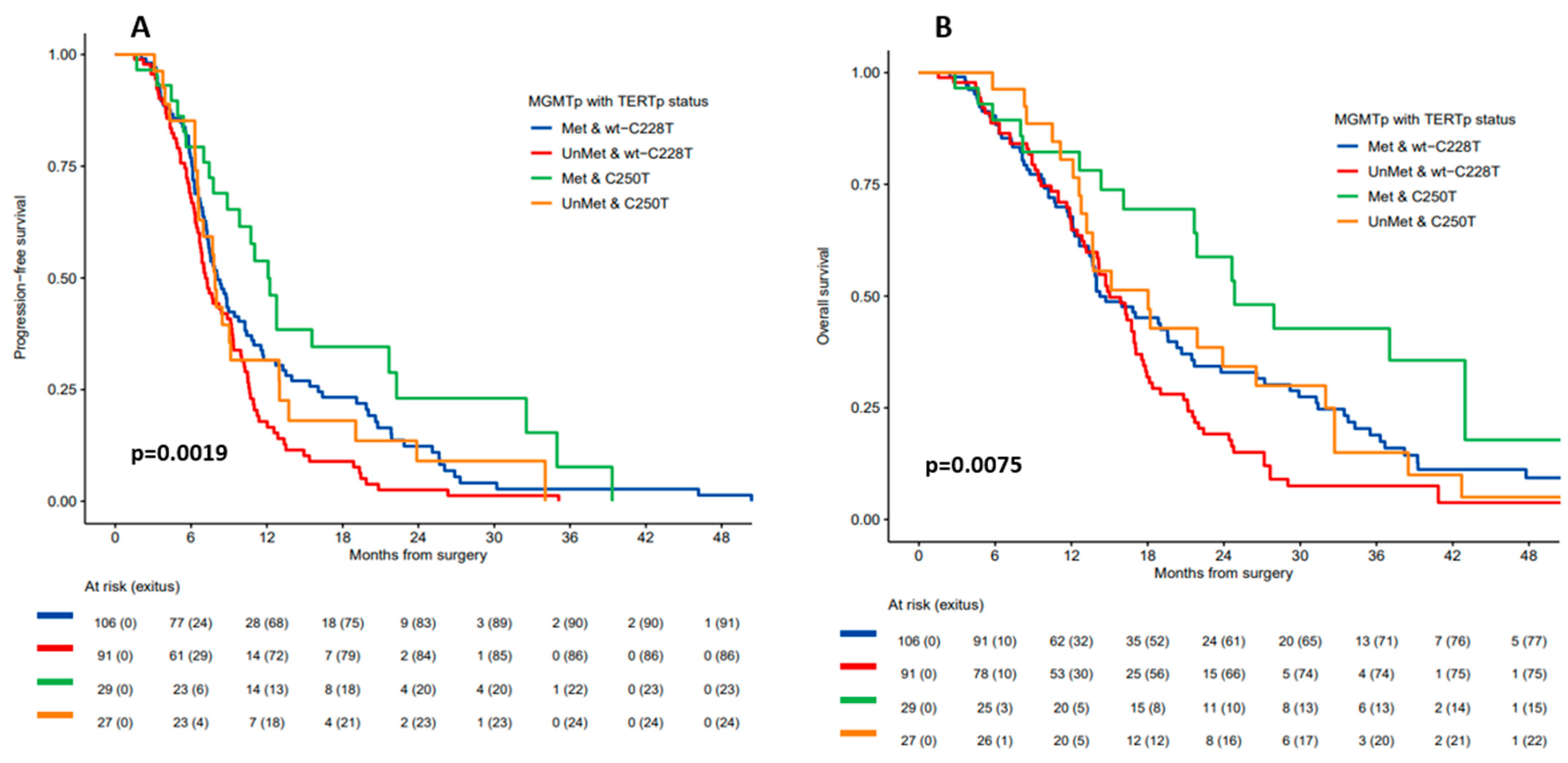
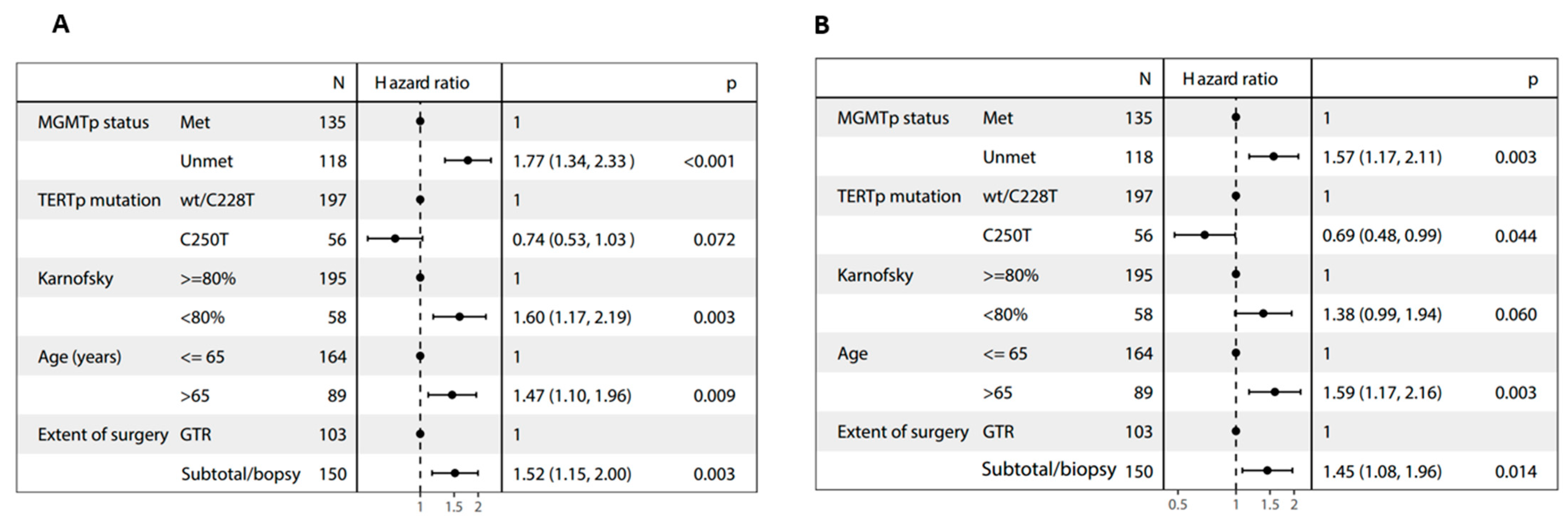
3.3. Outcomes

{kind=link}
{kind=link}
{kind=link}
| Comparisons | mPFS | mOS | |||
|---|---|---|---|---|---|
| Months (95% CI) | p | Months (95% CI) | p | ||
| All patients | 8 (7.4–9.1) | 16.3 (14.2–18) | |||
| TERTp wt vs. mut | wt | 7 (6.3–9) | 0.103 | 14.2 (11.9–22.4) | 0.801 |
| C228T + C250T | 8.4 (7.6–9.8) | 16.7 (14.7–18.4) | |||
| TERTp C250T vs. C228T vs. wt | C250T | 9.1 (7.8–12.8) | 0.048 | 21.9 (15.1–32.7) | 0.047 |
| C228T | 8.1 (7.3–9.4) | 16 (14–17.1) | |||
| wt | 7 (6.3–9) | 14.2 (11.9–22.4) | |||
| TERTp C250T vs. Wt + C228T | C250T | 9.1 (7.8–12.8) | 0.026 | 21.9 (15.1–32.7) | 0.016 |
| wt + C228T | 7.7 (7.1–8.9) | 15 (14–17.1) | |||
| MGMTp status & TERTp C250T vs. Wt + C228T | Met & C250T | 12.1 (9.8–22.3) | 0.002 | 24.8 (21.7–NR) | 0.007 |
| Met & wt + C228T | 8.1 (7.3–1.3) | 14.2 (13.4–2.7) | |||
| UnMet & wt + C228T | 7.1 (6.7–9.2) | 15 (14–17.1) | |||
| UnMet & C250T | 7.9 (6.6–13) | 18 (13.2–32.7) | |||

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ostrom, Q.T.; Patil, N.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2013-2017. Neuro Oncol. 2020, 22, iv1–iv96. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.; van den Bent, M.; Preusser, M.; Le Rhun, E.; Tonn, J.C.; Minniti, G.; Bendszus, M.; Balana, C.; Chinot, O.; Dirven, L.; et al. EANO guidelines on the diagnosis and treatment of diffuse gliomas of adulthood. Nat. Rev. Clin. Oncol. 2021, 18, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Mohile, N.A.; Messersmith, H.; Gatson, N.T.; Hottinger, A.F.; Lassman, A.; Morton, J.; Ney, D.; Nghiemphu, P.L.; Olar, A.; Olson, J.; et al. Therapy for Diffuse Astrocytic and Oligodendroglial Tumors in Adults: ASCO-SNO Guideline. J. Clin. Oncol. 2022, 40, 403–426. [Google Scholar] [CrossRef] [PubMed]
- Dratwa, M.; Wysoczanska, B.; Lacina, P.; Kubik, T.; Bogunia-Kubik, K. TERT-Regulation and Roles in Cancer Formation. Front. Immunol. 2020, 11, 589929. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, N.; Rachakonda, S.; Kumar, R. Telomeres and Telomere Length: A General Overview. Cancers 2020, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Chebly, A.; Ropio, J.; Peloponese, J.M.; Poglio, S.; Prochazkova-Carlotti, M.; Cherrier, F.; Ferrer, J.; Idrissi, Y.; Segal-Bendirdjian, E.; Chouery, E.; et al. Exploring hTERT promoter methylation in cutaneous T-cell lymphomas. Mol. Oncol. 2022, 16, 1931–1946. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.D.; Komosa, M.; Sudhaman, S.; Leao, R.; Zhang, C.H.; Apolonio, J.D.; Hermanns, T.; Wild, P.J.; Klocker, H.; Nassiri, F.; et al. Dual role of allele-specific DNA hypermethylation within the TERT promoter in cancer. J. Clin. Invest. 2021, 131, e146915. [Google Scholar] [CrossRef]
- Heaphy, C.M.; Subhawong, A.P.; Hong, S.M.; Goggins, M.G.; Montgomery, E.A.; Gabrielson, E.; Netto, G.J.; Epstein, J.I.; Lotan, T.L.; Westra, W.H.; et al. Prevalence of the alternative lengthening of telomeres telomere maintenance mechanism in human cancer subtypes. Am. J. Pathol. 2011, 179, 1608–1615. [Google Scholar] [CrossRef]
- Diplas, B.H.; He, X.; Brosnan-Cashman, J.A.; Liu, H.; Chen, L.H.; Wang, Z.; Moure, C.J.; Killela, P.J.; Loriaux, D.B.; Lipp, E.S.; et al. The genomic landscape of TERT promoter wildtype-IDH wildtype glioblastoma. Nat. Commun. 2018, 9, 2087. [Google Scholar] [CrossRef]
- Shay, J.W.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef]
- Huang, D.S.; Wang, Z.; He, X.J.; Diplas, B.H.; Yang, R.; Killela, P.J.; Meng, Q.; Ye, Z.Y.; Wang, W.; Jiang, X.T.; et al. Recurrent TERT promoter mutations identified in a large-scale study of multiple tumour types are associated with increased TERT expression and telomerase activation. Eur. J. Cancer 2015, 51, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Killela, P.J.; Pirozzi, C.J.; Healy, P.; Reitman, Z.J.; Lipp, E.; Rasheed, B.A.; Yang, R.; Diplas, B.H.; Wang, Z.; Greer, P.K.; et al. Mutations in IDH1, IDH2, and in the TERT promoter define clinically distinct subgroups of adult malignant gliomas. Oncotarget 2014, 5, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, K. TERT promoter mutation as a diagnostic marker for diffuse gliomas. Neuro Oncol. 2019, 21, 417–418. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, K.; Arita, H.; Satomi, K.; Yamasaki, K.; Matsushita, Y.; Nakamura, T.; Miyakita, Y.; Umehara, T.; Kobayashi, K.; Tamura, K.; et al. TERT promoter mutation status is necessary and sufficient to diagnose IDH-wildtype diffuse astrocytic glioma with molecular features of glioblastoma. Acta Neuropathol. 2021, 142, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Stichel, D.; Ebrahimi, A.; Reuss, D.; Schrimpf, D.; Ono, T.; Shirahata, M.; Reifenberger, G.; Weller, M.; Hanggi, D.; Wick, W.; et al. Distribution of EGFR amplification, combined chromosome 7 gain and chromosome 10 loss, and TERT promoter mutation in brain tumors and their potential for the reclassification of IDHwt astrocytoma to glioblastoma. Acta Neuropathol. 2018, 136, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Berzero, G.; Di Stefano, A.L.; Ronchi, S.; Bielle, F.; Villa, C.; Guillerm, E.; Capelle, L.; Mathon, B.; Laurenge, A.; Giry, M.; et al. IDH-wildtype lower-grade diffuse gliomas: The importance of histological grade and molecular assessment for prognostic stratification. Neuro Oncol. 2021, 23, 955–966. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly recurrent TERT promoter mutations in human melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Geng, P.; Zhao, X.; Ou, J.; Li, J.; Sa, R.; Liang, H. TERT Genetic Mutations as Prognostic Marker in Glioma. Mol. Neurobiol. 2017, 54, 3665–3669. [Google Scholar] [CrossRef]
- Malkki, H. Neuro-oncology: TERT promoter mutations could indicate poor prognosis in glioblastoma. Nat. Rev. Neurol. 2014, 10, 546. [Google Scholar]
- Simon, M.; Hosen, I.; Gousias, K.; Rachakonda, S.; Heidenreich, B.; Gessi, M.; Schramm, J.; Hemminki, K.; Waha, A.; Kumar, R. TERT promoter mutations: A novel independent prognostic factor in primary glioblastomas. Neuro Oncol. 2015, 17, 45–52. [Google Scholar] [CrossRef]
- Arita, H.; Ichimura, K. Prognostic significance of TERT promoter mutations in adult-type diffuse gliomas. Brain Tumor Pathol. 2022, 39, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Yamasaki, K.; Matsushita, Y.; Nakamura, T.; Shimokawa, A.; Takami, H.; Tanaka, S.; Mukasa, A.; Shirahata, M.; Shimizu, S.; et al. A combination of TERT promoter mutation and MGMT methylation status predicts clinically relevant subgroups of newly diagnosed glioblastomas. Acta Neuropathol. Commun. 2016, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Eckel-Passow, J.E.; Lachance, D.H.; Molinaro, A.M.; Walsh, K.M.; Decker, P.A.; Sicotte, H.; Pekmezci, M.; Rice, T.; Kosel, M.L.; Smirnov, I.V.; et al. Glioma Groups Based on 1p/19q, IDH, and TERT Promoter Mutations in Tumors. N. Engl. J. Med. 2015, 372, 2499–2508. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Rachakonda, P.S.; Hosen, I.; Volz, F.; Hemminki, K.; Weyerbrock, A.; Kumar, R. TERT promoter mutations and telomere length in adult malignant gliomas and recurrences. Oncotarget 2015, 6, 10617–10633. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.N.; Lie, A.; Li, T.; Chowdhury, R.; Liu, F.; Ozer, B.; Wei, B.; Green, R.M.; Ellingson, B.M.; Wang, H.J.; et al. Human TERT promoter mutation enables survival advantage from MGMT promoter methylation in IDH1 wild-type primary glioblastoma treated by standard chemoradiotherapy. Neuro Oncol. 2017, 19, 394–404. [Google Scholar] [PubMed]
- Gramatzki, D.; Felsberg, J.; Hentschel, B.; Wolter, M.; Schackert, G.; Westphal, M.; Regli, L.; Thon, N.; Tatagiba, M.; Wick, W.; et al. Telomerase reverse transcriptase promoter mutation– and O6-methylguanine DNA methyltransferase promoter methylation–mediated sensitivity to temozolomide in isocitrate dehydrogenase–wild-type glioblastoma: Is there a link? Eur. J. Cancer 2021, 147, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Giunco, S.; Padovan, M.; Angelini, C.; Cavallin, F.; Cerretti, G.; Morello, M.; Caccese, M.; Rizzo, B.; d’Avella, D.; Puppa, A.D.; et al. Prognostic role and interaction of TERT promoter status, telomere length and MGMT promoter methylation in newly diagnosed IDH wild-type glioblastoma patients. ESMO Open 2023, 8, 101570. [Google Scholar] [CrossRef]
- Kessler, T.; Berberich, A.; Sadik, A.; Sahm, F.; Gorlia, T.; Meisner, C.; Hoffmann, D.C.; Wick, A.; Kickingereder, P.; Rubmann, P.; et al. Methylome analyses of three glioblastoma cohorts reveal chemotherapy sensitivity markers within DDR genes. Cancer Med. 2020, 9, 8373–8385. [Google Scholar] [CrossRef]
- Carrato, C.; Alameda, F.; Esteve-Codina, A.; Pineda, E.; Arpi, O.; Martinez-Garcia, M.; Mallo, M.; Gut, M.; Lopez-Martos, R.; Barco, S.D.; et al. Glioblastoma TCGA Mesenchymal and IGS 23 Tumors are Identifiable by IHC and have an Immune-phenotype Indicating a Potential Benefit from Immunotherapy. Clin. Cancer Res. 2020, 26, 6600–6609. [Google Scholar] [CrossRef]
- Estival, A.; Sanz, C.; Ramirez, J.-L.; Maria Velarde, J.; Domenech, M.; Carrato, C.; de las Penas, R.; Gil-Gil, M.; Sepulveda, J.; Armengol, R.; et al. Pyrosequencing versus methylation-specific PCR for assessment of MGMT methylation in tumor and blood samples of glioblastoma patients. Sci. Rep. 2019, 9, 11125. [Google Scholar] [CrossRef]
- Esteve-Codina, A.; Alameda, F.; Carrato, C.; Pineda, E.; Arpi, O.; Martinez Garcia, M.; Mallo, M.; Gut, M.; Dabad, M.; Tortosa, A.; et al. RNA-Sequencing and immunohistochemistry reveal ZFN7 as a stronger marker of survival than molecular subtypes in G-CIMP-negative glioblastoma. Clin. Cancer Res. 2020, 27, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Sergushichev, A.A. An algorithm for fast preranked gene set enrichment analysis using cumulative statistic calculation. bioRxiv 2016, 60012, 1–9. [Google Scholar]
- Korotkevich, G.; Sukhov, V.; Budin, N.; Shpak, B.; Artyomov, M.N.; Sergushichev, A. Fast gene set enrichment analysis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Boeva, V.; Popova, T.; Bleakley, K.; Chiche, P.; Cappo, J.; Schleiermacher, G.; Janoueix-Lerosey, I.; Delattre, O.; Barillot, E. Control-FREEC: A tool for assessing copy number and allelic content using next-generation sequencing data. Bioinformatics 2012, 28, 423–425. [Google Scholar] [CrossRef]
- Boscolo-Rizzo, P.; Tirelli, G.; Polesel, J.; Sia, E.; Phillips, V.; Borsetto, D.; De Rossi, A.; Giunco, S. TERT promoter mutations in head and neck squamous cell carcinoma: A systematic review and meta-analysis on prevalence and prognostic significance. Oral. Oncol. 2023, 140, 106398. [Google Scholar] [CrossRef]
- You, H.; Wu, Y.; Chang, K.; Shi, X.; Chen, X.D.; Yan, W.; Li, R. Paradoxical prognostic impact of TERT promoter mutations in gliomas depends on different histological and genetic backgrounds. CNS Neurosci. Ther. 2017, 23, 790–797. [Google Scholar] [CrossRef]
- Huse, J.T. TERT promoter mutation designates biologically aggressive primary glioblastoma. Neuro Oncol. 2015, 17, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.G.; Nguyen, T.Q.; Ngo, T.N.M.; Nguyen, H.C.; Fung, K.M.; Dunn, I.F. The interaction between TERT promoter mutation and MGMT promoter methylation on overall survival of glioma patients: A meta-analysis. BMC Cancer 2020, 20, 897. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Larsson, C.; Xu, D. Mechanisms underlying the activation of TERT transcription and telomerase activity in human cancer: Old actors and new players. Oncogene 2019, 38, 6172–6183. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Kumar, R. TERT promoter mutations in telomere biology. Mutat. Res. Rev. Mutat. Res. 2017, 771, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.S.; Wen, J.; Bacchetti, S. The human telomerase catalytic subunit hTERT: Organization of the gene and characterization of the promoter. Hum. Mol. Genet. 1999, 8, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Q.L.; Sun, W.; Chandrasekharan, P.; Cheng, H.S.; Ying, Z.; Lakshmanan, M.; Raju, A.; Tenen, D.G.; Cheng, S.Y.; et al. Non-canonical NF-kappaB signalling and ETS1/2 cooperatively drive C250T mutant TERT promoter activation. Nat. Cell Biol. 2015, 17, 1327–1338. [Google Scholar] [CrossRef]
- Ropio, J.; Prochazkova-Carlotti, M.; Batista, R.; Pestana, A.; Chebly, A.; Ferrer, J.; Idrissi, Y.; Cappellen, D.; Duraes, C.; Boaventura, P.; et al. Spotlight on hTERT Complex Regulation in Cutaneous T-Cell Lymphomas. Genes 2023, 14, 439. [Google Scholar] [CrossRef]
- Mavrogiannou, E.; Strati, A.; Stathopoulou, A.; Tsaroucha, E.G.; Kaklamanis, L.; Lianidou, E.S. Real-time RT-PCR quantification of human telomerase reverse transcriptase splice variants in tumor cell lines and non-small cell lung cancer. Clin. Chem. 2007, 53, 53–61. [Google Scholar] [CrossRef]
- Salgado, C.; Roelse, C.; Nell, R.; Gruis, N.; van Doorn, R.; van der Velden, P. Interplay between TERT promoter mutations and methylation culminates in chromatin accessibility and TERT expression. PLoS ONE 2020, 15, e0231418. [Google Scholar] [CrossRef]
- Ohba, S.; Kuwahara, K.; Yamada, S.; Abe, M.; Hirose, Y. Correlation between IDH, ATRX, and TERT promoter mutations in glioma. Brain Tumor Pathol. 2020, 37, 33–40. [Google Scholar] [CrossRef]
- Pierini, T.; Nardelli, C.; Lema Fernandez, A.G.; Pierini, V.; Pellanera, F.; Nofrini, V.; Gorello, P.; Moretti, M.; Arniani, S.; Roti, G.; et al. New somatic TERT promoter variants enhance the Telomerase activity in Glioblastoma. Acta Neuropathol. Commun. 2020, 8, 145. [Google Scholar] [CrossRef]
- Williams, E.A.; Miller, J.J.; Tummala, S.S.; Penson, T.; Iafrate, A.J.; Juratli, T.A.; Cahill, D.P. TERT promoter wild-type glioblastomas show distinct clinical features and frequent PI3K pathway mutations. Acta Neuropathol. Commun. 2018, 6, 106. [Google Scholar] [CrossRef]


| Characteristic | All Patients n = 257 a | TERTp C228T n = 145 | TERTp C250T n = 57 | TERTp Wild-Type n = 55 | p b |
|---|---|---|---|---|---|
| Sex | 0.787 | ||||
| Male | 163 (63.4%) | 93 (64.1%) | 34 (59.6%) | 36 (65.5%) | |
| Female | 94 (36.6%) | 52 (35.9%) | 23 (40.4%) | 19 (34.5%) | |
| Age, yrs—median (range) | 60.6 (19–81) | 61.5 (31–81) | 61.5 (32–81) | 57.2 (19–78) | 0.052 |
| ≤65 | 167 (65.0%) | 89 (61.4%) | 35 (61.4%) | 43 (78.2%) | 0.069 |
| >65 | 90 (35.0%) | 56 (38.6%) | 22 (38.6%) | 12 (21.8%) | |
| KPS | 0.344 | ||||
| ≥80% | 198 (77.0%) | 109 (75.2%) | 48 (84.2%) | 41 (74.5%) | |
| <80% | 59 (23.0%) | 36 (24.8%) | 9 (15.8%) | 14 (25.5%) | |
| Extent of surgery | 0.200 | ||||
| Gross total resection | 103 (40.1%) | 57 (39.3%) | 28 (49.1%) | 18 (32.7%) | |
| Subtotal/biopsy | 154 (59.9%) | 88 (60.7%) | 29 (50.9%) | 37 (67.3%) | |
| MGMTp status | 0.686 | ||||
| Methylated | 135 (52.5%) | 81 (55.9%) | 29 (50.9%) | 25 (45.5%) | |
| Unmethylated | 118 (45.9%) | 62 (42.8%) | 27 (47.4%) | 29 (52.7%) | |
| Unknown | 4 (1.6%) | 2 (1.4%) | 1 (1.8%) | 1 (1.8%) |
| Molecular Alteration | Status | All Patients n = 92 | TERTp C228T n = 57 | TERTp C250T n = 18 | TERTp-wt n = 17 | p a |
|---|---|---|---|---|---|---|
| EGFR mutation | No | 79 (85.9%) | 50 (87.7%) | 13 (72.2%) | 16 (94.1%) | 0.216 |
| Yes | 13 (14.1%) | 7 (12.3%) | 5 (27.8%) | 1 (5.88%) | ||
| EGFR amplification | No | 49 (53.3%) | 27 (47.4%) | 12 (66.7%) | 10 (58.8%) | 0.316 |
| Yes | 43 (46.7%) | 30 (52.6%) | 6 (33.3%) | 7 (41.2%) | ||
| P53 mutation | No | 73 (79.3%) | 47 (82.5%) | 13 (72.2%) | 13 (76.5%) | 0.586 |
| Yes | 19 (20.7%) | 10 (17.5%) | 5 (27.8%) | 4 (23.5%) | ||
| PTEN mutation | No | 63 (68.5%) | 40 (70.2) | 11 (61.1%) | 12 (70.6%) | 0.754 |
| Yes | 29 (31.5%) | 17 (29.8) | 7 (38.9) | 5 (29.4%) | ||
| BRAF mutation | No | 89 (96.7%) | 57 (100%) | 16 (88.9%) | 16 (94.1%) | 0.052 |
| Yes | 3 (3.26%) | 0 (0.00%) | 2 (11.1%) | 1 (5.88%) | ||
| CDKN2AB Loss | No loss | 41 (44.6%) | 26 (45.6%) | 8 (44.4%) | 7 (41.2%) | 0.949 |
| Loss | 51 (55.4%) | 31 (54.4%) | 10 (55.6%) | 10 (58.8%) | ||
| PIK3 family mutation | No | 60 (65.2%) | 39 (68.4%) | 10 (55.6%) | 11 (64.7%) | 0.125 |
| Yes | 32 (34.8%) | 18 (31.6%) | 8 (44.4%) | 6 (35.3%) | ||
| MYC mutation | No | 89 (96.7%) | 56 (98.2%) | 18 (100%) | 15 (88.2%) | 0.114 |
| Yes | 3 (3.26%) | 1 (1.75%) | 0 (0.00%) | 2 (11.8%) | ||
| DAXX mutation | No | 90 (97.8%) | 56 (98.2%) | 17 (94.4%) | 17 (100%) | 0.619 |
| Yes | 2 (2.17%) | 1 (1.75%) | 1 (5.56%) | 0 (0.00%) | ||
| SMARCA family mutation | No | 77 (83.7) | 49 (86.0%) | 14 (77.8%) | 14 (82.4) | 0.249 |
| Yes | 15 (16.3) | 8 (14.0) | 4 (22.2) | 3 (17.6) | ||
| ATRX mutation | No | 86 (93.5%) | 52 (91.2%) | 17 (94.4%) | 17 (100%) | 0.724 |
| Yes | 6 (6.52%) | 5 (8.77%) | 1 (5.56%) | 0 (0.00%) | ||
| TERC LncRNA | Differential expression | No differences in the expression of TERC LncRNA | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorria, T.; Crous, C.; Pineda, E.; Hernandez, A.; Domenech, M.; Sanz, C.; Jares, P.; Muñoz-Mármol, A.M.; Arpí-Llucía, O.; Melendez, B.; et al. The C250T Mutation of TERTp Might Grant a Better Prognosis to Glioblastoma by Exerting Less Biological Effect on Telomeres and Chromosomes Than the C228T Mutation. Cancers 2024, 16, 735. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16040735
Gorria T, Crous C, Pineda E, Hernandez A, Domenech M, Sanz C, Jares P, Muñoz-Mármol AM, Arpí-Llucía O, Melendez B, et al. The C250T Mutation of TERTp Might Grant a Better Prognosis to Glioblastoma by Exerting Less Biological Effect on Telomeres and Chromosomes Than the C228T Mutation. Cancers. 2024; 16(4):735. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16040735
Chicago/Turabian StyleGorria, Teresa, Carme Crous, Estela Pineda, Ainhoa Hernandez, Marta Domenech, Carolina Sanz, Pedro Jares, Ana María Muñoz-Mármol, Oriol Arpí-Llucía, Bárbara Melendez, and et al. 2024. "The C250T Mutation of TERTp Might Grant a Better Prognosis to Glioblastoma by Exerting Less Biological Effect on Telomeres and Chromosomes Than the C228T Mutation" Cancers 16, no. 4: 735. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers16040735