A Novel Thermostable Cytochrome P450 from Sequence-Based Metagenomics of Binh Chau Hot Spring as a Promising Catalyst for Testosterone Conversion

 ,
,
Abstract
:1. Introduction
2. Results
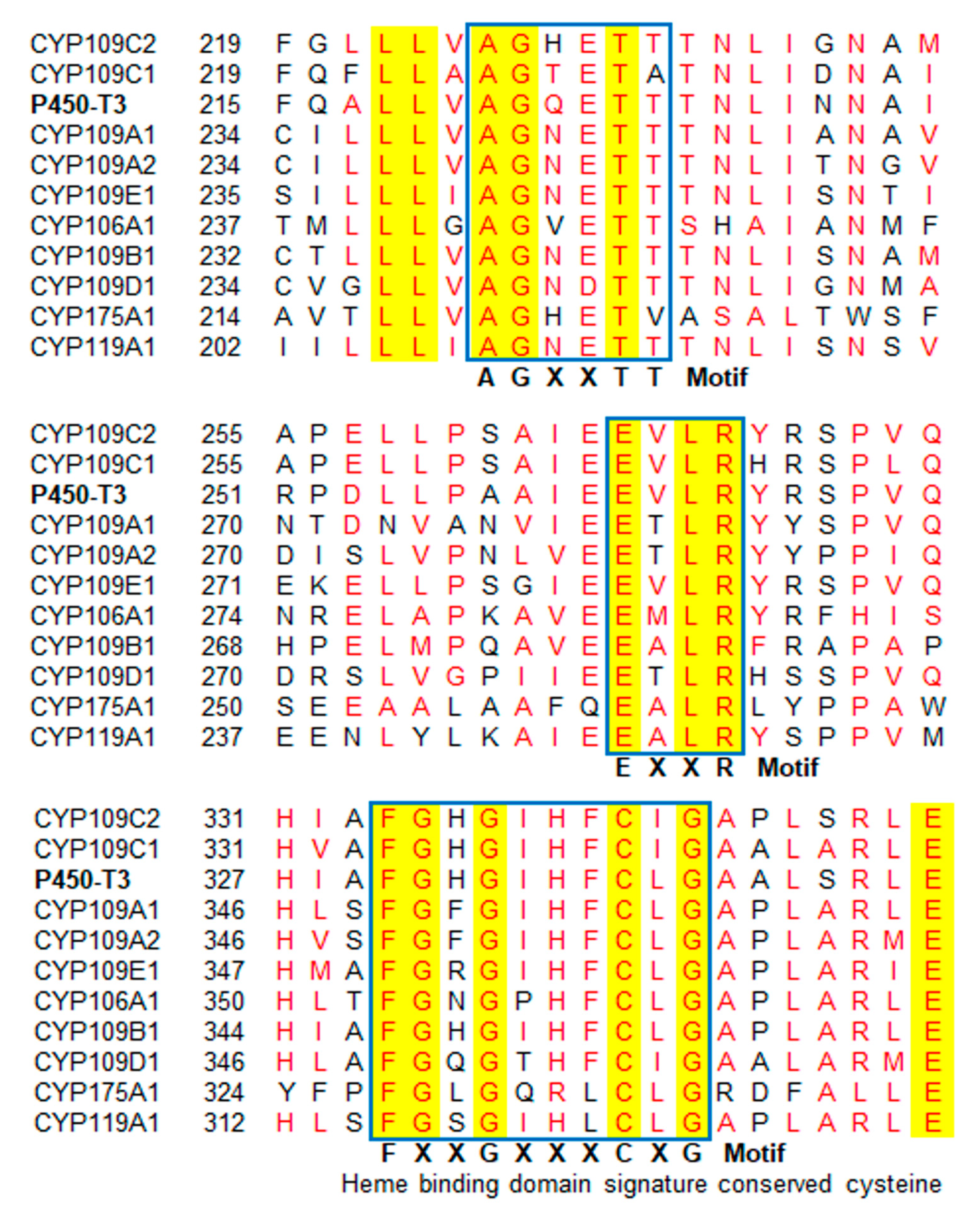
2.1. Identification and Bioinformatic Analyses of P450-T3
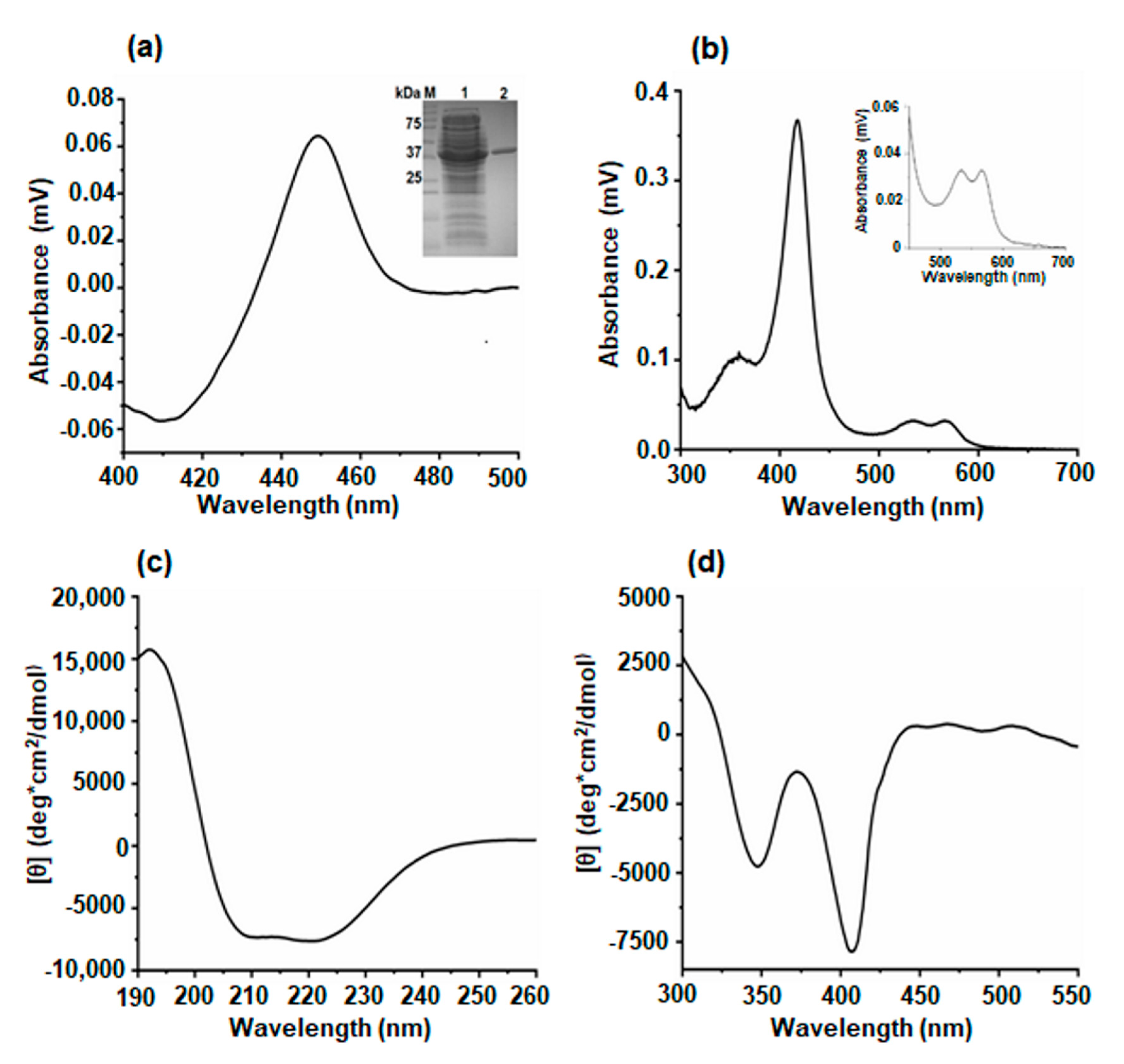
2.2. Production and Purification of P450-T3
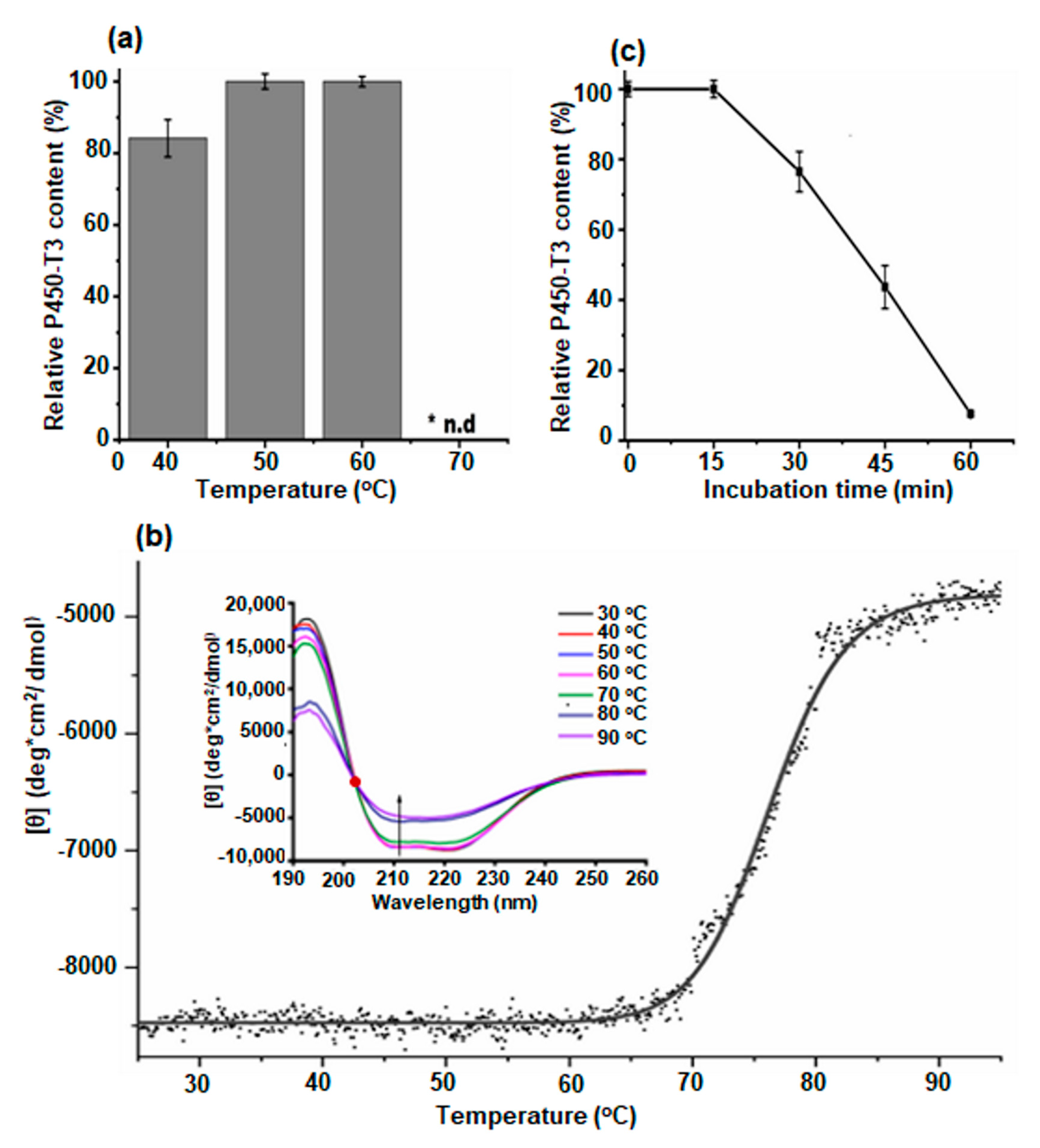
2.3. Thermal Stability of P450-T3
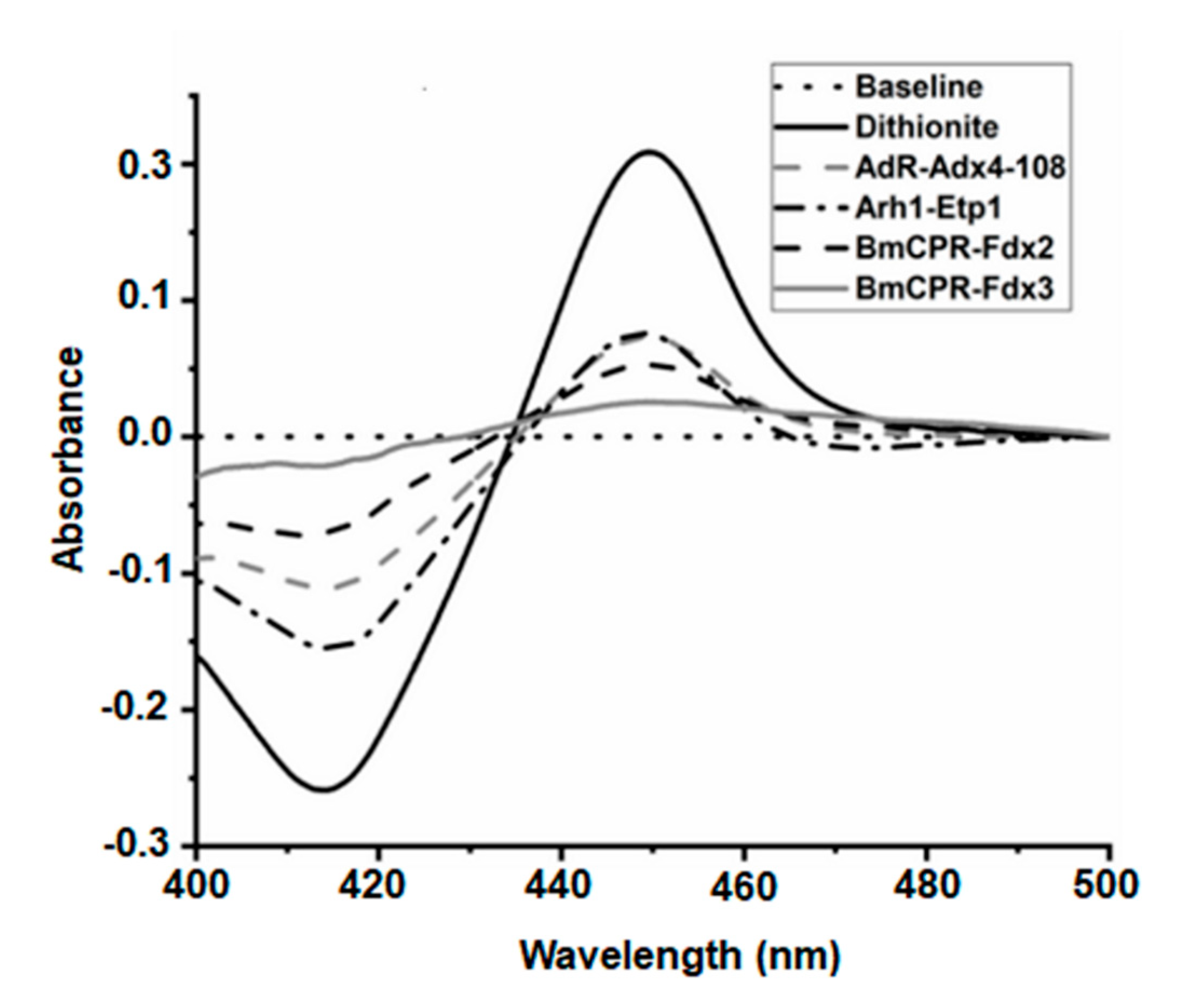
2.4. Identification of Electron Transfer Partners
2.5. Substrate Screening for P450-T3
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. Bioinformatics Analysis
4.2.2. Production and Purification of P450-T3
4.2.3. Circular Dichroism Spectroscopy
4.2.4. Thermal Stability
4.2.5. Investigation of Electron Transfer Partners
4.2.6. Monitoring of Substrate Binding
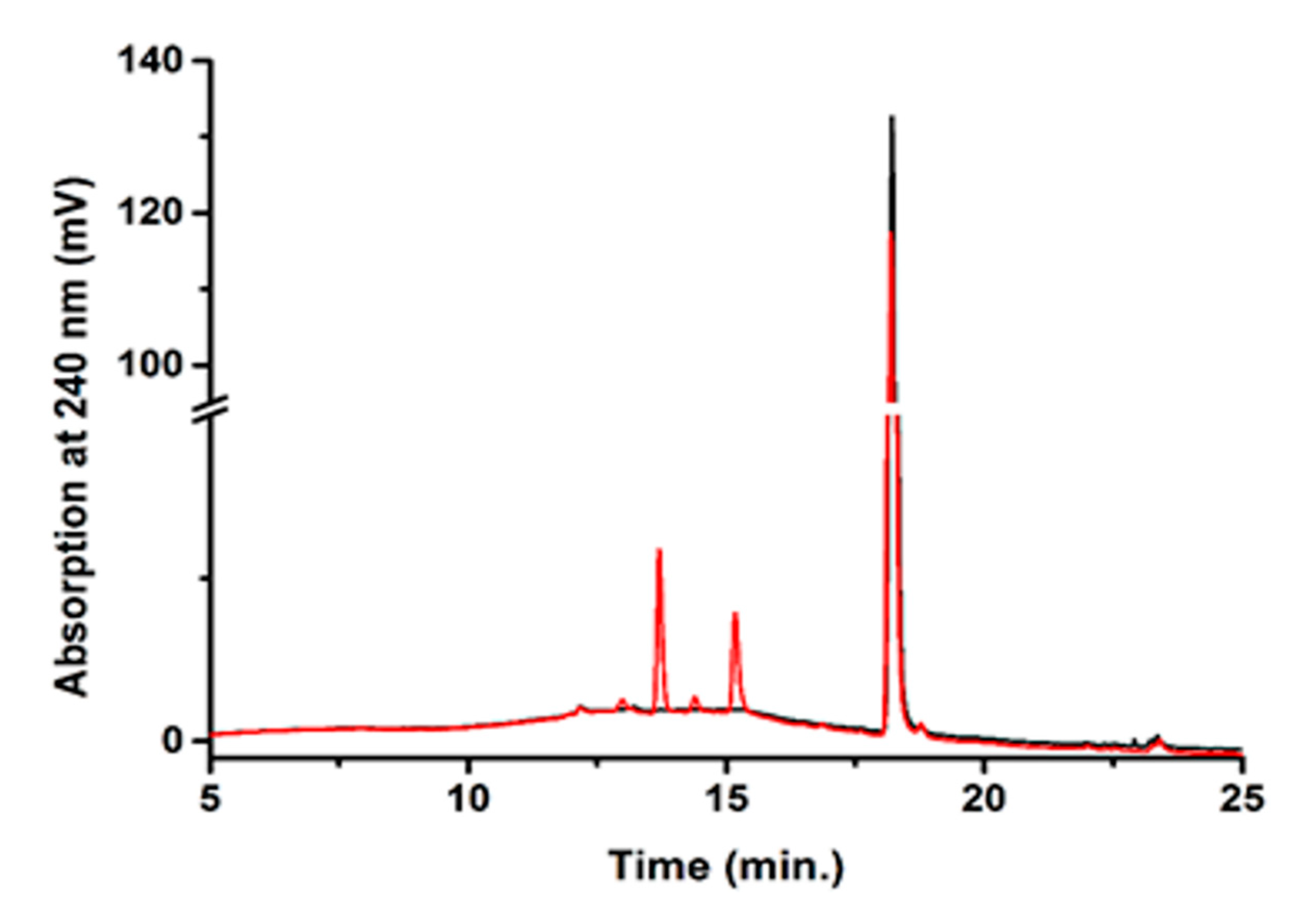
4.2.7. In Vitro Conversion of Testosterone and HPLC Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Werck-Reichhart, D.; Feyereisen, R. Cytochromes P450: A success story. Genome Biol. 2000, 1, reviews3003.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, R. Cytochromes P450 as versatile biocatalysts. J. Biotechnol. 2006, 124, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Urlacher, V.; Schmid, R.D. Biotransformations using prokaryotic P450 monooxygenases. Curr. Opin. Biotechnol. 2002, 13, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Niehaus, F.; Bertoldo, C.; Kähler, M.; Antranikian, G. Extremophiles as a source of novel enzymes for industrial application. Appl. Microbiol. Biotechnol. 1999, 51, 711–729. [Google Scholar] [CrossRef]
- O’Reilly, E.; Köhler, V.; Flitsch, S.L.; Turner, N.J. Cytochromes P450 as useful biocatalysts: Addressing the limitations. Chem. Commun. 2011, 47, 2490–2501. [Google Scholar] [CrossRef]
- Bernhardt, R.; Urlacher, V.B. Cytochromes P450 as promising catalysts for biotechnological application: Chances and limitations. Appl. Microbiol. Biotechnol. 2014, 98, 6185–6203. [Google Scholar] [CrossRef]
- Syed, K.; Shale, K.; Nazir, K.H.M.N.H.; Krasevec, N.; Mashele, S.S.; Pagadala, N.S. Genome-wide identification, annotation and characterization of novel thermostable cytochrome P450 monooxygenases from the thermophilic biomass-degrading fungi Thielavia terrestris and Myceliophthora thermophila. Genes Genom. 2014, 36, 321–333. [Google Scholar] [CrossRef]
- Wright, R.L.; Harris, K.; Solow, B.; White, R.H.; Kennelly, P.J. Cloning of a potential cytochrome P450 from the archaeon Sulfolobus solfataricus. FEBS Lett. 1996, 384, 235–239. [Google Scholar] [CrossRef] [Green Version]
- McLean, M.A.; Maves, S.A.; Weiss, K.E.; Krepich, S.; Sligar, S.G. Characterization of a Cytochrome P450 from the Acidothermophilic Archaea Sulfolobus solfataricus. Biochem. Biophys. Res. Commun. 1998, 252, 166–172. [Google Scholar] [CrossRef]
- Koo, L.S.; Tschirret-Guth, R.A.; Straub, W.E.; Moënne-Loccoz, P.; Loehr, T.M.; Ortiz de Montellano, P.R. The active site of the thermophilic CYP119 from Sulfolobus solfataricus. J. Biol. Chem. 2000, 275, 14112–14123. [Google Scholar] [CrossRef] [Green Version]
- Koo, L.S.; Immoos, C.E.; Cohen, M.S.; Farmer, P.J.; Ortiz de Montellano, P.R. Enhanced electron transfer and lauric acid hydroxylation by site-directed mutagenesis of CYP119. J. Am. Chem. Soc. 2002, 124, 5684–5691. [Google Scholar] [CrossRef] [PubMed]
- Blasco, F.; Kauffmann, I.; Schmid, R.D. CYP175A1 from Thermus thermophilus HB27, the first beta-carotene hydroxylase of the P450 superfamily. Appl. Microbiol. Biotechnol. 2004, 64, 671–674. [Google Scholar] [CrossRef]
- Yano, J.K.; Blasco, F.; Li, H.; Schmid, R.D.; Henne, A.; Poulos, T.L. Preliminary characterization and crystal structure of a thermostable cytochrome P450 from Thermus thermophilus. J. Biol. Chem. 2003, 278, 608–616. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.L.; Thomson, R.E.S.; Strohmaier, S.J.; Gumulya, Y.; Gillam, E.M.J. Determinants of thermostability in the cytochrome P450 fold. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 97–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavanti, M.; Porter, J.L.; Sabatini, S.; Turner, N.J.; Flitsch, S.L. Panel of New Thermostable CYP116B Self-Sufficient Cytochrome P450 Monooxygenases that Catalyze C−H Activation with a Diverse Substrate Scope. ChemCatChem 2018, 10, 1042–1051. [Google Scholar] [CrossRef] [Green Version]
- Gumulya, Y.; Baek, J.-M.; Wun, S.-J.; Thomson, R.E.S.; Harris, K.L.; Hunter, D.J.B.; Behrendorff, J.B.Y.H.; Kulig, J.; Zheng, S.; Wu, X.; et al. Engineering highly functional thermostable proteins using ancestral sequence reconstruction. Nat. Catal. 2018, 1, 878–888. [Google Scholar] [CrossRef]
- Kubota, M.; Nodate, M.; Yasumoto-Hirose, M.; Uchiyama, T.; Kagami, O.; Shizuri, Y.; Misawa, N. Isolation and functional analysis of cytochrome P450 CYP153A genes from various environments. Biosci. Biotechnol. Biochem. 2005, 69, 2421–2430. [Google Scholar] [CrossRef]
- Kim, B.S.; Kim, S.Y.; Park, J.; Park, W.; Hwang, K.Y.; Yoon, Y.J.; Oh, W.K.; Kim, B.Y.; Ahn, J.S. Sequence-based screening for self-sufficient P450 monooxygenase from a metagenome library. J. Appl. Microbiol. 2007, 102, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- López-López, O.; Cerdán, M.E.; González-Siso, M.I. Hot spring metagenomics. Life 2013, 3, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Tung, N.V.; Hoang, N.H.; Thoa, N.K. Mining cytochrome p450 genes through next generation sequencing and metagenomic analysis from Binh Chau hot spring. Acad. J. Biol. 2019, 41. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.E.; Kraševec, N.; Mullins, J.; Nelson, D.R. The CYPome (Cytochrome P450 complement) of Aspergillus nidulans. Fungal Genet. Biol. 2009, 46, S53–S61. [Google Scholar] [CrossRef] [PubMed]
- Martinis, S.A.; Atkins, W.M.; Stayton, P.S.; Sligar, S.G. A conserved residue of cytochrome P-450 is involved in heme-oxygen stability and activation. J. Am. Chem. Soc. 1989, 111, 9252–9253. [Google Scholar] [CrossRef]
- Hasemann, C.A.; Kurumbail, R.G.; Boddupalli, S.S.; Peterson, J.A.; Deisenhofer, J. Structure and function of cytochromes P450: A comparative analysis of three crystal structures. Structure 1995, 3, 41–62. [Google Scholar] [CrossRef]
- Syed, K.; Mashele, S.S. Comparative analysis of P450 signature motifs EXXR and CXG in the Large and diverse kingdom of fungi: Identification of evolutionarily conserved amino acid patterns characteristic of P450 family. PLoS ONE 2014, 9, e95616. [Google Scholar] [CrossRef] [Green Version]
- Omura, T.; Sato, R. The carbon monoxide-binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J. Biol. Chem. 1964, 239, 2370–2378. [Google Scholar]
- Ravichandran, K.G.; Boddupalli, S.S.; Hasermann, C.A.; Peterson, J.A.; Deisenhofer, J. Crystal structure of hemoprotein domain of P450BM-3, a prototype for microsomal P450’s. Science 1993, 261, 731–736. [Google Scholar] [CrossRef]
- Luthra, A.; Denisov, I.G.; Sligar, S.G. Spectroscopic features of cytochrome P450 reaction intermediates. Arch. Biochem. Biophys. 2011, 507, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Mak, P.J.; Denisov, I.G. Spectroscopic studies of the cytochrome P450 reaction mechanisms. Biochim. Biophys. Acta 2018, 1866, 178–204. [Google Scholar] [CrossRef]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Milhim, M.; Gerber, A.; Neunzig, J.; Hannemann, F.; Bernhardt, R. A Novel NADPH-dependent flavoprotein reductase from Bacillus megaterium acts as an efficient cytochrome P450 reductase. J. Biotechnol. 2016, 231, 83–94. [Google Scholar] [CrossRef]
- Milhim, M.; Putkaradze, N.; Abdulmughni, A.; Kern, F.; Hartz, P.; Bernhardt, R. Identification of a new plasmid-encoded cytochrome P450 CYP107DY1 from Bacillus megaterium with a catalytic activity towards mevastatin. J. Biotechnol. 2016, 240, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.J.; Hannemann, F.; Schiffler, B.; Ewen, K.M.; Kappl, R.; Heinemann, U.; Bernhardt, R. Structural and thermodynamic characterization of the adrenodoxin-like domain of the electron-transfer protein Etp1 from Schizosaccharomyces pombe. J. Inorg. Biochem. 2011, 105, 957–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleser, M.; Hannemann, F.; Hutter, M.; Zapp, J.; Bernhardt, R. CYP105A1 mediated 3-hydroxylation of glimepiride and glibenclamide using a recombinant Bacillus megaterium whole-cell catalyst. J. Biotechnol. 2012, 157, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Janocha, S.; Bernhardt, R. Design and characterization of an efficient CYP105A1-based whole-cell biocatalyst for the conversion of resin acid diterpenoids in permeabilized Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 7639–7649. [Google Scholar] [CrossRef]
- Ringle, M.; Khatri, Y.; Zapp, J.; Hannemann, F.; Bernhardt, R. Application of a new versatile electron transfer system for cytochrome P450-based Escherichia coli whole-cell bioconversions. Appl. Microbiol. Biotechnol. 2013, 97, 7741–7754. [Google Scholar] [CrossRef]
- Kern, F.; Khatri, Y.; Litzenburger, M.; Bernhardt, R. CYP267A1 and CYP267B1 from Sorangium cellulosum So ce56 are highly versatile drug metabolizers. Drug Metab. Dispos. 2016, 44, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Khatri, Y.; Hannemann, F.; Girhard, M.; Kappl, R.; Même, A.; Ringle, M.; Janocha, S.; Leize-Wagner, E.; Urlacher, V.B.; Bernhardt, R. Novel family members of CYP109 from Sorangium cellulosum So ce56 exhibit characteristic biochemical and biophysical properties. Biotechnol. Appl. Biochem. 2013, 60, 18–29. [Google Scholar] [CrossRef]
- Abdulmughni, A.; Jóźwik, I.K.; Brill, E.; Hannemann, F.; Thunnissen, A.-M.W.H.; Bernhardt, R. Biochemical and structural characterization of CYP109A2, a vitamin D3 25-hydroxylase from Bacillus megaterium. FEBS J. 2017, 284, 3881–3894. [Google Scholar] [CrossRef] [Green Version]
- Khatri, Y.; Girhard, M.; Romankiewicz, A.; Ringle, M.; Hannemann, F.; Urlacher, V.B.; Hutter, M.C.; Bernhardt, R. Regioselective hydroxylation of norisoprenoids by CYP109D1 from Sorangium cellulosum So ce56. Appl. Microbiol. Biotechnol. 2010, 88, 485–495. [Google Scholar] [CrossRef]
- Jóźwik, I.K.; Kiss, F.M.; Gricman, Ł.; Abdulmughni, A.; Brill, E.; Zapp, J.; Pleiss, J.; Bernhardt, R.; Thunnissen, A.-M.W.H. Structural basis of steroid binding and oxidation by the cytochrome P450 CYP109E1 from Bacillus megaterium. FEBS J. 2016, 283, 4128–4148. [Google Scholar] [CrossRef] [PubMed]
- Girhard, M.; Klaus, T.; Khatri, Y.; Bernhardt, R.; Urlacher, V.B. Characterization of the versatile monooxygenase CYP109B1 from Bacillus subtilis. Appl. Microbiol. Biotechnol. 2010, 87, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Zhang, T.; Hall, E.A.; Hutchinson, S.; Cryle, M.J.; Wong, L.-L.; Zhou, W.; Bell, S.G. The crystal structure of the versatile cytochrome P450 enzyme CYP109B1 from Bacillus subtilis. Mol. BioSyst. 2015, 11, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Putkaradze, N.; Litzenburger, M.; Abdulmughni, A.; Milhim, M.; Brill, E.; Hannemann, F.; Bernhardt, R. CYP109E1 is a novel versatile statin and terpene oxidase from Bacillus megaterium. Appl. Microbiol. Biotechnol. 2017, 101, 8379–8393. [Google Scholar] [CrossRef] [PubMed]
- Putkaradze, N.; Litzenburger, M.; Hutter, M.C.; Bernhardt, R. CYP109E1 from Bacillus megaterium acts as a 24- and 25-Hydroxylase for cholesterol. ChemBioChem 2019, 20, 655–658. [Google Scholar] [CrossRef]
- Park, S.-Y.; Yamane, K.; Adachi, S.; Shiro, Y.; Weiss, K.E.; Maves, S.A.; Sligar, S.G. Thermophilic cytochrome P450 (CYP119) from Sulfolobus solfataricus: High resolution structure and functional properties. J. Inorg. Biochem. 2002, 91, 491–501. [Google Scholar] [CrossRef]
- Matsumura, H.; Matsuda, K.; Nakamura, N.; Ohtaki, A.; Yoshida, H.; Kamitori, S.; Yohda, M.; Ohno, H. Monooxygenation by a thermophilic cytochrome P450via direct electron donation from NADH. Metallomics 2011, 3, 389–395. [Google Scholar] [CrossRef]
- Schallmey, A.; den Besten, G.; Teune, I.G.P.; Kembaren, R.F.; Janssen, D.B. Characterization of cytochrome P450 monooxygenase CYP154H1 from the thermophilic soil bacterium Thermobifida fusca. Appl. Microbiol. Biotechnol. 2011, 89, 1475–1485. [Google Scholar] [CrossRef] [Green Version]
- Ho, W.W.; Li, H.; Nishida, C.R.; Ortiz de Montellano, P.R.; Poulos, T.L. Crystal structure and properties of CYP231A2 from the Thermoacidophilic archaeon Picrophilus torridus. Biochemistry 2008, 47, 2071–2079. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, Y.; Guengerich, F.P.; Ma, L.; Li, S.; Zhang, W. Engineering cytochrome P450 enzyme systems for biomedical and biotechnological applications. J. Biol. Chem. 2020, 295, 833–849. [Google Scholar] [CrossRef] [Green Version]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol. Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Xia, Y.; Qu, H.; Li, A.-D.; Liu, R.; Wang, Y.; Zhang, T. Discovery of new cellulases from the metagenome by a metagenomics-guided strategy. Biotechnol. Biofuels 2016, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henne, A.; Schmitz, R.A.; Bömeke, M.; Gottschalk, G.; Daniel, R. Screening of Environmental dna libraries for the presence of genes conferring lipolytic activity on Escherichia coli. Appl. Environ. Microbiol. 2000, 66, 3113–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wu, H.; Wang, A.; Du, P.; Pei, X.; Li, H.; Yin, X.; Huang, L.; Xiong, X. Prospecting metagenomic enzyme subfamily genes for dna family shuffling by a novel pcr-based approach. J. Biol. Chem. 2010, 285, 41509–41516. [Google Scholar] [CrossRef] [Green Version]
- Yun, J.; Kang, S.; Park, S.; Yoon, H.; Kim, M.-J.; Heu, S.; Ryu, S. Characterization of a novel amylolytic enzyme encoded by a gene from a soil-derived metagenomic library. Appl. Environ. Microbiol. 2004, 70, 7229–7235. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, T.; Miyazaki, K. Product-induced gene expression, a product-responsive reporter assay used to screen metagenomic libraries for enzyme-encoding genes. Appl. Environ. Microbiol. 2010, 76, 7029–7035. [Google Scholar] [CrossRef] [Green Version]
- Knietsch, A.; Waschkowitz, T.; Bowien, S.; Henne, A.; Daniel, R. Construction and screening of metagenomic libraries derived from enrichment cultures: Generation of a Gene Bank for genes conferring alcohol oxidoreductase activity on Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 1408–1416. [Google Scholar] [CrossRef] [Green Version]
- Knietsch, A.; Bowien, S.; Whited, G.; Gottschalk, G.; Daniel, R. Identification and characterization of coenzyme B12-dependent glycerol dehydratase- and diol dehydratase-encoding genes from metagenomic DNA libraries derived from enrichment cultures. Appl. Environ. Microbiol. 2003, 69, 3048–3060. [Google Scholar] [CrossRef] [Green Version]
- Musumeci, M.A.; Lozada, M.; Rial, D.V.; Mac Cormack, W.P.; Jansson, J.K.; Sjöling, S.; Carroll, J.; Dionisi, H.M. Prospecting biotechnologically-relevant monooxygenases from cold sediment metagenomes: An in silico approach. Mar. Drugs 2017, 15, 114. [Google Scholar] [CrossRef]
- Gabor, E.M.; Alkema, W.B.L.; Janssen, D.B. Quantifying the accessibility of the metagenome by random expression cloning techniques. Environ. Microbiol. 2004, 6, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Tirawongsaroj, P.; Sriprang, R.; Harnpicharnchai, P.; Thongaram, T.; Champreda, V.; Tanapongpipat, S.; Pootanakit, K.; Eurwilaichitr, L. Novel thermophilic and thermostable lipolytic enzymes from a Thailand hot spring metagenomic library. J. Biotechnol. 2008, 133, 42–49. [Google Scholar] [CrossRef]
- Sharma, P.K.; Singh, K.; Singh, R.; Capalash, N.; Ali, A.; Mohammad, O.; Kaur, J. Characterization of a thermostable lipase showing loss of secondary structure at ambient temperature. Mol. Biol. Rep. 2012, 39, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.-X.; Wang, Y.-B.; Pan, Y.-J.; Li, W.-F. Differences in amino acids composition and coupling patterns between mesophilic and thermophilic proteins. Amino Acids. 2008, 34, 25–33. [Google Scholar] [CrossRef]
- Lee, C.-W.; Wang, H.-J.; Hwang, J.-K.; Tseng, C.-P. Protein thermal stability enhancement by designing salt bridges: A combined computational and experimental study. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Argos, P.; Rossmann, M.G.; Grau, U.M.; Zuber, H.; Frank, G.; Tratschin, J.D. Thermal stability and protein structure. Biochemistry 1979, 18, 5698–5703. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tsai, C.J.; Nussinov, R. Factors enhancing protein thermostability. Protein Eng. 2000, 13, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pack, S.P.; Yoo, Y.J. Packing-based difference of structural features between thermophilic and mesophilic proteins. Int. J. Biol. Macromol. 2005, 35, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Saab-Rincón, G.; Alwaseem, H.; Guzmán-Luna, V.; Olvera, L.; Fasan, R. Stabilization of the reductase domain in the catalytically self-sufficient cytochrome P450BM3 by consensus-guided mutagenesis. ChemBioChem 2018, 19, 622–632. [Google Scholar] [CrossRef]
- Eiben, S.; Bartelmäs, H.; Urlacher, V.B. Construction of a thermostable cytochrome P450 chimera derived from self-sufficient mesophilic parents. Appl. Microbiol. Biotechnol. 2007, 75, 1055–1061. [Google Scholar] [CrossRef]
- Goyal, S.; Banerjee, S.; Mazumdar, S. Oxygenation of monoenoic fatty acids by CYP175A1, an orphan cytochrome P450 from Thermus thermophilus HB27. Biochemistry 2012, 51, 7880–7890. [Google Scholar] [CrossRef]
- Salamanca-Pinzon, S.G.; Khatri, Y.; Carius, Y.; Keller, L.; Müller, R.; Lancaster, C.R.D.; Bernhardt, R. Structure-function analysis for the hydroxylation of Δ4 C21-steroids by the myxobacterial CYP260B1. FEBS Lett. 2016, 590, 1838–1851. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Ge, S.; Gao, M.; Jian, R.; Chen, X.; Xu, X.; Li, D.; Zhang, K.; Chen, W.-H. Synthesis and biological activity of embelin and its derivatives: An overview. Mini Rev. Med. Chem. 2020, 20, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.; Blanc, D.; Cunliffe, W.J. 13-cis retinoic acid and acne. Lancet 1980, 316, 1048–1049. [Google Scholar] [CrossRef]
- Schroeder, M.; Zouboulis, C.C. All-trans-retinoic acid and 13-cis-retinoic acid: Pharmacokinetics and biological activity in different cell culture models of human keratinocytes. Horm. Metab. Res. 2007, 39, 136–140. [Google Scholar] [CrossRef]
- Zasada, M.; Budzisz, E. Retinoids: Active molecules influencing skin structure formation in cosmetic and dermatological treatments. Postepy Dermatol. Alergol. 2019, 36, 392–397. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Peng, Y.; Zhao, J.; Li, Q.; Yu, X.; Acevedo-Rocha, C.G.; Li, A. Bacterial cytochrome P450-catalyzed regio- and stereoselective steroid hydroxylation enabled by directed evolution and rational design. Bioresour. Bioprocess. 2020, 7, 2. [Google Scholar] [CrossRef]
- Nelson, D.R. The cytochrome p450 homepage. Hum. Genom. 2009, 4, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 1999, 292, 195–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.-F.; Yang, G.-Y.; Zhang, Y.; Xie, Y.; Withers, S.G.; Feng, Y. A general and efficient strategy for generating the stable enzymes. Sci. Rep. 2016, 6, 33797. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Chhatpar, H.S. Purification, characterization and thermodynamics of antifungal protease from Streptomyces sp. A6. J. Basic Microbiol. 2011, 51, 424–432. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Shift from Low-Spin to High-Spin |
|---|---|
| Lauric acid | + |
| Palmitic acid | + |
| Embelin | + |
| Retinoic acid (all-trans) | + |
| Retinoic acid (13-cis) | + |
| 11-Deoxycorticosterone (DOC) | - |
| 11-Deoxycortisol (RSS) | - |
| Progesterone | - |
| Testosterone | + |
| Nootkatone | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, K.-T.; Nguyen, N.-L.; Tung, N.V.; Nguyen, H.H.; Milhim, M.; Le, T.-T.-X.; Lai, T.-H.-N.; Phan, T.-T.-M.; Bernhardt, R. A Novel Thermostable Cytochrome P450 from Sequence-Based Metagenomics of Binh Chau Hot Spring as a Promising Catalyst for Testosterone Conversion. Catalysts 2020, 10, 1083. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10091083
Nguyen K-T, Nguyen N-L, Tung NV, Nguyen HH, Milhim M, Le T-T-X, Lai T-H-N, Phan T-T-M, Bernhardt R. A Novel Thermostable Cytochrome P450 from Sequence-Based Metagenomics of Binh Chau Hot Spring as a Promising Catalyst for Testosterone Conversion. Catalysts. 2020; 10(9):1083. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10091083
Chicago/Turabian StyleNguyen, Kim-Thoa, Ngọc-Lan Nguyen, Nguyen Van Tung, Huy Hoang Nguyen, Mohammed Milhim, Thi-Thanh-Xuan Le, Thi-Hong-Nhung Lai, Thi-Tuyet-Minh Phan, and Rita Bernhardt. 2020. "A Novel Thermostable Cytochrome P450 from Sequence-Based Metagenomics of Binh Chau Hot Spring as a Promising Catalyst for Testosterone Conversion" Catalysts 10, no. 9: 1083. https://0-doi-org.brum.beds.ac.uk/10.3390/catal10091083