
Getting the Most Out of Enzyme Cascades: Strategies to Optimize In Vitro Multi-Enzymatic Reactions

 , and
, and
Abstract
:
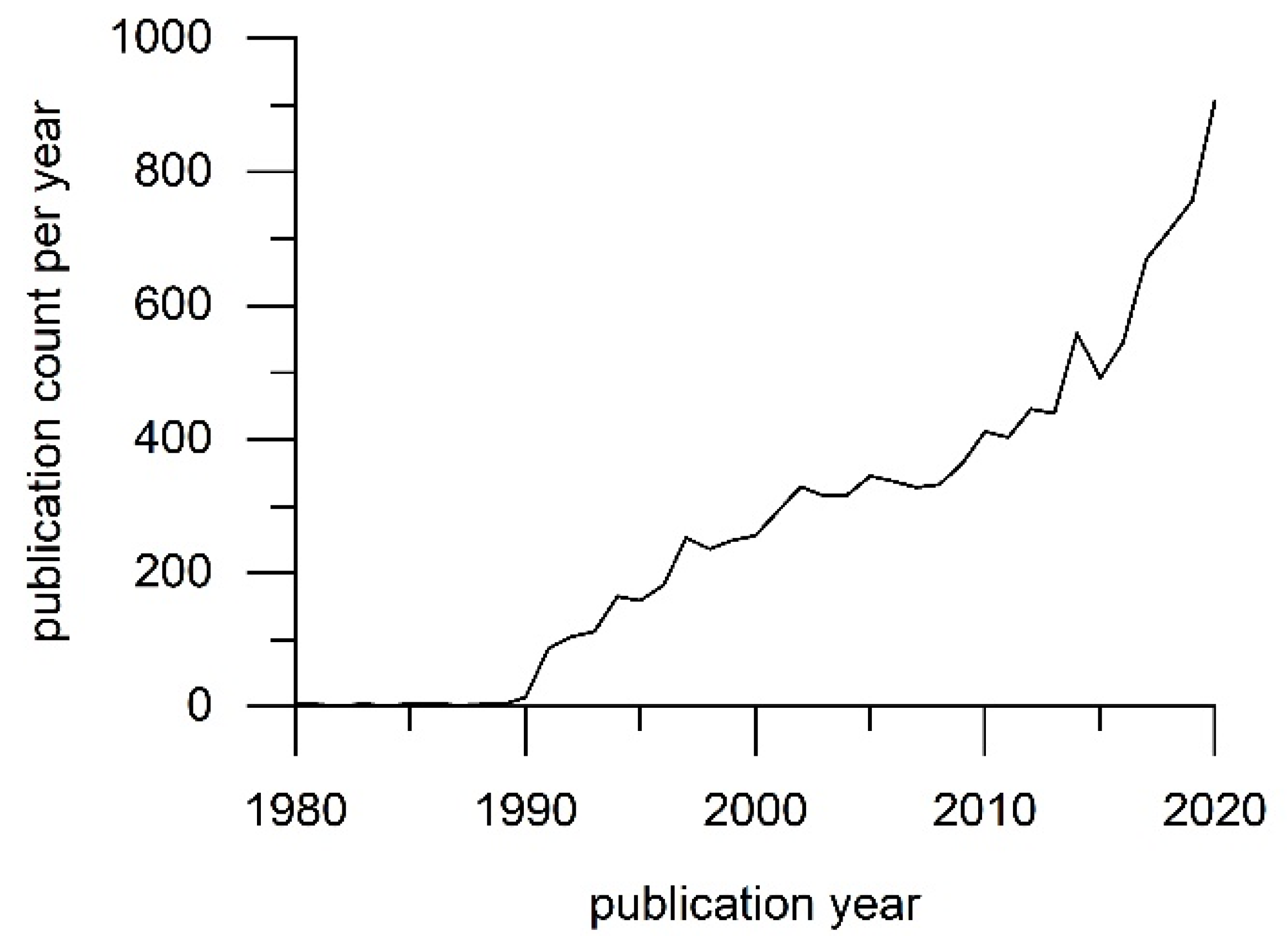
1. Introduction
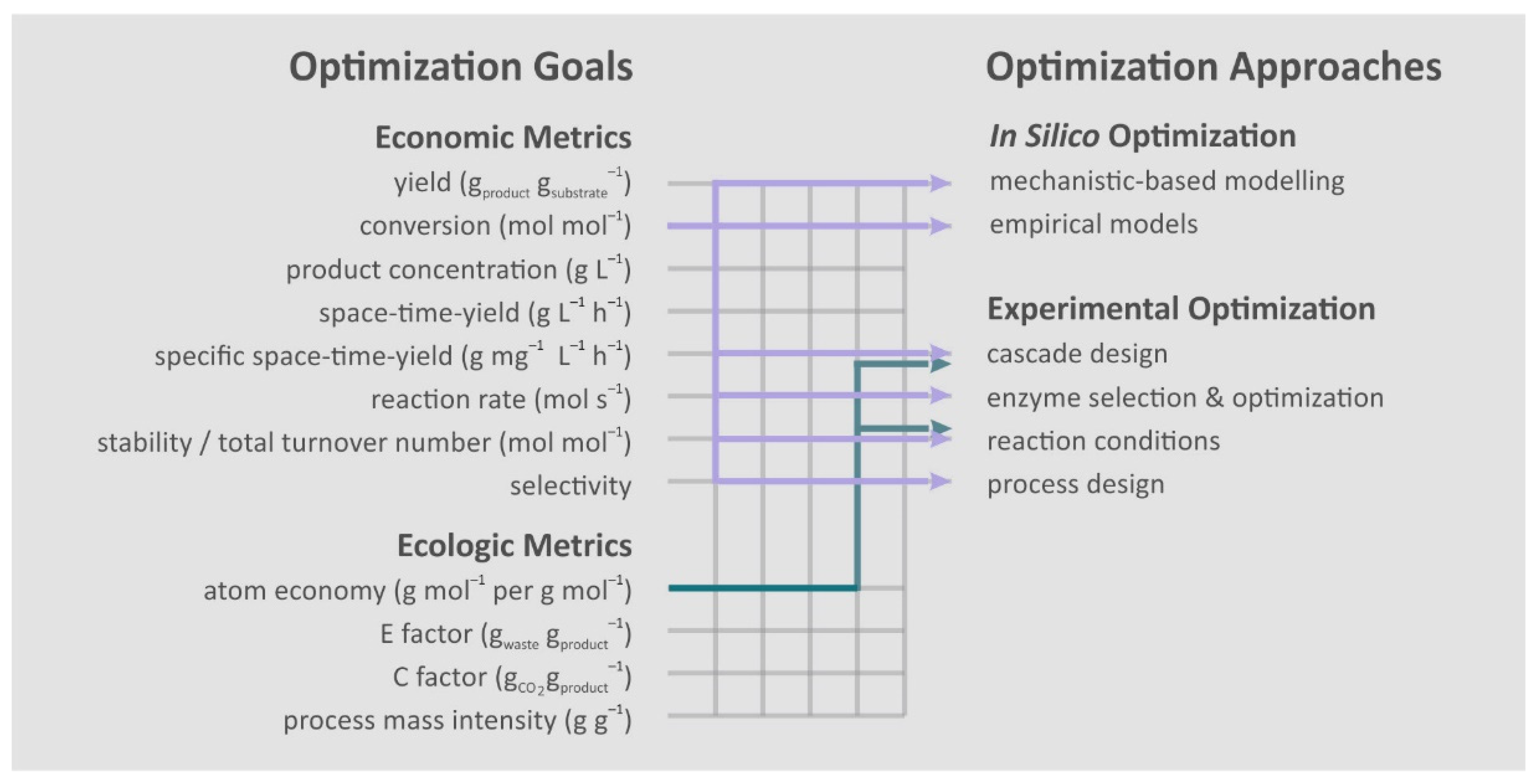
2. Optimization Goals
3. Optimization Approaches
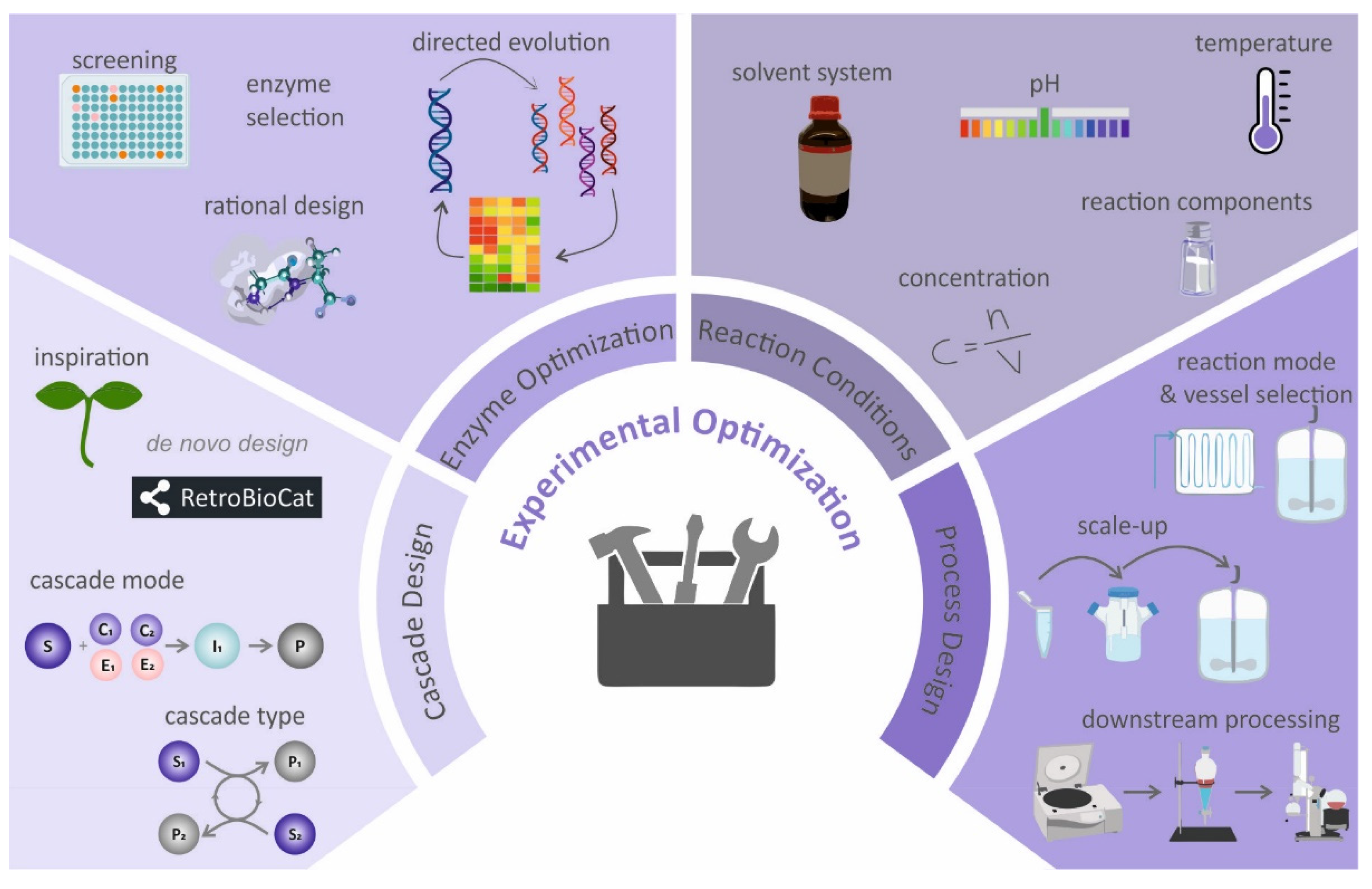
3.1. Experimental-Based Optimization of Enzyme Cascades
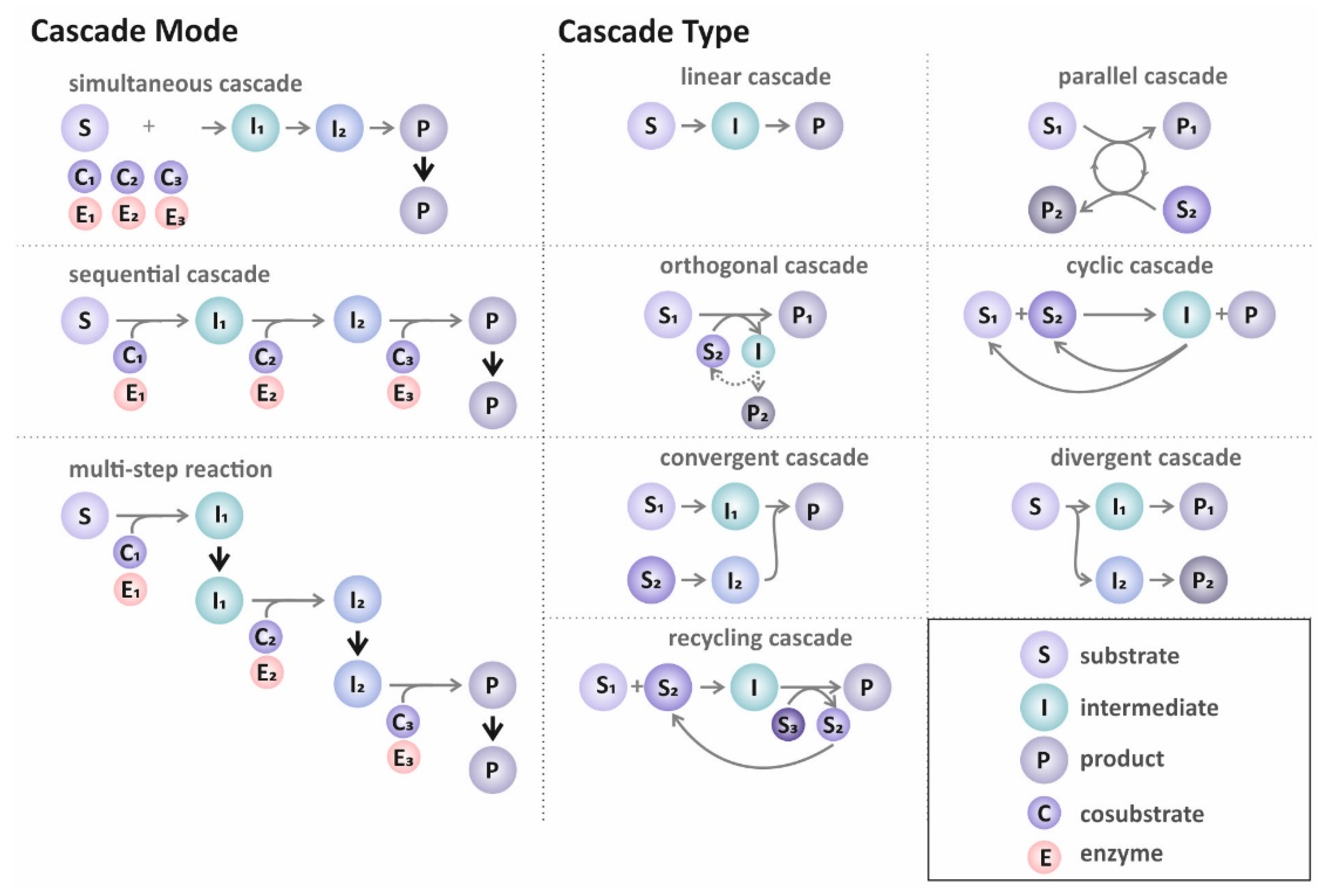
3.1.1. Cascade Design
Reaction Pathway
Cascade Mode
Cascade Type
3.1.2. Enzyme-Selection and -Optimization
3.1.3. Reaction Conditions
Solvent System
Reaction Temperature
Substrate and Cofactor Concentrations
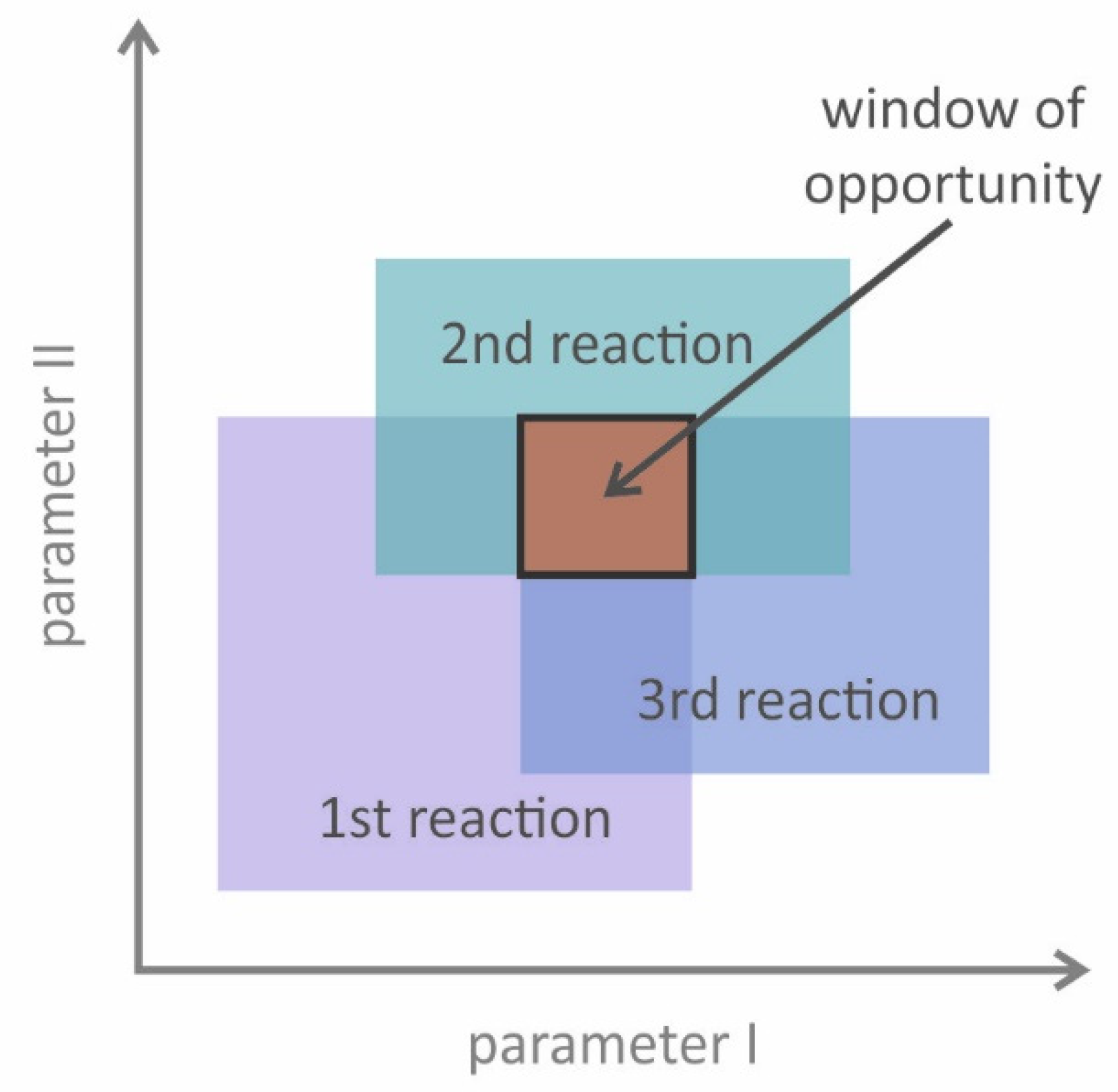
3.1.4. Process Design
3.2. In Silico-Based Optimization of Enzyme Cascades
3.2.1. Mechanistic Based Modelling of Enzyme Cascades
3.2.2. Empirical Models for Enzyme Cascade Optimization
4. Remaining Challenges and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Winkler, C.K.; Schrittwieser, J.H.; Kroutil, W. Power of Biocatalysis for Organic Synthesis. ACS Cent. Sci. 2021, 7, 55–71. [Google Scholar] [CrossRef]
- Abdelraheem, E.M.M.; Busch, H.; Hanefeld, U.; Tonin, F. Biocatalysis explained: From pharmaceutical to bulk chemical production. React. Chem. Eng. 2019, 4, 1878–1894. [Google Scholar] [CrossRef] [Green Version]
- Beigi, M.; Gauchenova, E.; Walter, L.; Waltzer, S.; Bonina, F.; Stillger, T.; Rother, D.; Pohl, M.; Müller, M. Regio- and Stereoselective Aliphatic–Aromatic Cross-Benzoin Reaction: Enzymatic Divergent Catalysis. Chem. Eur. J. 2016, 22, 13999–14005. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.J.; Nicholas, J.; Humphreys, L. Biocatalysis in Organic Synthesis: The Retrosynthesis Approach; Royal Society of Chemistry: Cambridge, UK, 2018; ISBN 978-1-78262-530-8. [Google Scholar]
- Ghaffari-Moghaddam, M.; Eslahi, H.; Aydin, Y.A.; Saloglu, D. Enzymatic processes in alternative reaction media: A mini review. J. Biol. Methods 2015, 2, e25. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Gallego, F.; Schmidt-Dannert, C. Multi-enzymatic synthesis. Curr. Opin. Chem. Biol. 2010, 14, 174–183. [Google Scholar] [CrossRef]
- Rosenthal, K.; Lütz, S. Recent developments and challenges of biocatalytic processes in the pharmaceutical industry. Curr. Opin. Green Sustain. Chem. 2018, 11, 58–64. [Google Scholar] [CrossRef]
- Bell, E.L.; Finnigan, W.; France, S.P.; Green, A.P.; Hayes, M.A.; Hepworth, L.J.; Lovelock, S.L.; Niikura, H.; Osuna, S.; Romero, E.; et al. Biocatalysis. Nat. Rev. Methods Prim. 2021, 1, 46. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, W.; Hepworth, L.J.; Flitsch, S.L.; Turner, N.J. RetroBioCat as a computer-aided synthesis planning tool for biocatalytic reactions and cascades. Nat. Catal. 2021, 4, 98–104. [Google Scholar] [CrossRef]
- García-Junceda, E.; Lavandera, I.; Rother, D.; Schrittwieser, J.H. (Chemo)enzymatic cascades—Nature’s synthetic strategy transferred to the laboratory. J. Mol. Catal. B Enzym. 2015, 114, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Claaßen, C.; Gerlach, T.; Rother, D. Stimulus-Responsive Regulation of Enzyme Activity for One-Step and Multi-Step Syntheses. Adv. Synth. Catal. 2019, 361, 2387–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdmann, V.; Lichman, B.R.; Zhao, J.; Simon, R.C.; Kroutil, W.; Ward, J.M.; Hailes, H.C.; Rother, D. Enzymatic and Chemoenzymatic Three-Step Cascades for the Synthesis of Stereochemically Complementary Trisubstituted Tetrahydroisoquinolines. Angew. Chem. 2017, 129, 12677–12681. [Google Scholar] [CrossRef] [Green Version]
- France, S.P.; Hepworth, L.J.; Turner, N.J.; Flitsch, S.L. Constructing Biocatalytic Cascades: In Vitro and in Vivo Approaches to de Novo Multi-Enzyme Pathways. ACS Catal. 2017, 7, 710–724. [Google Scholar] [CrossRef]
- Schrittwieser, J.H.; Velikogne, S.; Hall, M.; Kroutil, W. Artificial Biocatalytic Linear Cascades for Preparation of Organic Molecules. Chem. Rev. 2017, 118, 270–348. [Google Scholar] [CrossRef] [Green Version]
- Sperl, J.M.; Sieber, V. Multienzyme Cascade Reactions—Status and Recent Advances. ACS Catal. 2018, 8, 2385–2396. [Google Scholar] [CrossRef]
- Shaeri, J.; Wright, I.; Rathbone, E.B.; Wohlgemuth, R.; Woodley, J.M. Characterization of enzymatic D-xylulose 5-phosphate synthesis. Biotechnol. Bioeng. 2008, 101, 761–767. [Google Scholar] [CrossRef]
- Woodley, J.M.; Titchener-Hooker, N.J. The use of windows of operation as a bioprocess design tool. Bioprocess Eng. 1996, 14, 263–268. [Google Scholar] [CrossRef]
- Schwander, T.; Von Borzyskowski, L.S.; Burgener, S.; Cortina, N.S.; Erb, T.J. A synthetic pathway for the fixation of carbon dioxide in vitro. Science 2016, 354, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Hold, C.; Billerbeck, S.; Panke, S. Forward design of a complex enzyme cascade reaction. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Gmelch, T.J.; Sperl, J.M.; Sieber, V. Optimization of a reduced enzymatic reaction cascade for the production of L-alanine. Sci. Rep. 2019, 9, 11754. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.H.; Sayar, A.; Kaulmann, U.; Dalby, P.A.; Ward, J.M.; Woodley, J.M. Reaction modelling and simulation to assess the integrated use of transketolase and ω-transaminase for the synthesis of an aminotriol. Biocatal. Biotransform. 2006, 24, 449–457. [Google Scholar] [CrossRef]
- Ardao, I.; Zeng, A.-P. In silico evaluation of a complex multi-enzymatic system using one-pot and modular approaches: Application to the high-yield production of hydrogen from a synthetic metabolic pathway. Chem. Eng. Sci. 2013, 87, 183–193. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, C.; Zou, R.; Zhou, K.; Stephanopoulos, G.; Too, H.P. Statistical experimental design guided optimization of a one-pot biphasic multienzyme total synthesis of amorpha-4,11-diene. PLoS ONE 2013, 8, e79650. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Kohlhaas, M.; Enoki, J.; Meier, R.; Schönenberger, B.; Wohlgemuth, R.; Kourist, R.; Niemeyer, F.; van Niekerk, D.; Bräsen, C.; et al. A combined experimental and modelling approach for the Weimberg pathway optimisation. Nat. Commun. 2020, 11, 1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, P.; Sinha, S. Industrial Biocatalysis: An Insight into Trends and Future Directions. Curr. Sustain. Energy Rep. 2020, 7, 66–72. [Google Scholar] [CrossRef]
- Truppo, M.D. Biocatalysis in the Pharmaceutical Industry: The Need for Speed. ACS Med. Chem. Lett. 2017, 8, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Dias Gomes, M.; Woodley, J.M. Considerations when Measuring Biocatalyst Performance. Molecules 2019, 24, 3573. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.; Lütz, S.; Rosenthal, K. Environmental assessment of enzyme production and purification. Molecules 2021, 26, 573. [Google Scholar] [CrossRef]
- Carro, J.; Fernández-Fueyo, E.; Fernández-Alonso, C.; Cañada, J.; Ullrich, R.; Hofrichter, M.; Alcalde, M.; Ferreira, P.; Martínez, A.T. Self-sustained enzymatic cascade for the production of 2,5-furandicarboxylic acid from 5-methoxymethylfurfural. Biotechnol. Biofuels 2018, 11, 86. [Google Scholar] [CrossRef] [Green Version]
- Baraibar, Á.G.; von Lieres, E.; Wiechert, W.; Pohl, M.; Rother, D. Effective Production of (S)-α-Hydroxy ketones: An Reaction Engineering Approach. Top. Catal. 2013, 57, 401–411. [Google Scholar] [CrossRef]
- Woodley, J.M. New frontiers in biocatalysis for sustainable synthesis. Curr. Opin. Green Sustain. Chem. 2020, 21, 22–26. [Google Scholar] [CrossRef]
- Korman, T.P.; Opgenorth, P.H.; Bowie, J.U. A synthetic biochemistry platform for cell free production of monoterpenes from glucose. Nat. Commun. 2017, 8, 15526. [Google Scholar] [CrossRef]
- Cheng, T.; Liu, H.; Zou, H.; Chen, N.; Shi, M.; Xie, C.; Zhao, G.; Xian, M. Enzymatic process optimization for the in vitro production of isoprene from mevalonate. Microb. Cell Fact. 2017, 16, 8. [Google Scholar] [CrossRef] [Green Version]
- Honda, K.; Kimura, K.; Ninh, P.H.; Taniguchi, H.; Okano, K.; Ohtake, H. In vitro bioconversion of chitin to pyruvate with thermophilic enzymes. J. Biosci. Bioeng. 2017, 124, 296–301. [Google Scholar] [CrossRef]
- Huffman, M.A.; Fryszkowska, A.; Alvizo, O.; Borra-Garske, M.; Campos, K.R.; Canada, K.A.; Devine, P.N.; Duan, D.; Forstater, J.H.; Grosser, S.T.; et al. Design of an in vitro biocatalytic cascade for the manufacture of islatravir. Science 2019, 366, 1255–1259. [Google Scholar] [CrossRef]
- Babich, L.; van Hemert, L.J.C.; Bury, A.; Hartog, A.F.; Falcicchio, P.; van der Oost, J.; van Herk, T.; Wever, R.; Rutjes, F.P.J.T. Synthesis of non-natural carbohydrates from glycerol and aldehydes in a one-pot four-enzyme cascade reaction. Green Chem. 2011, 13, 2895–2900. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Kelly, R.M.; Han, Y. Simultaneous biosynthesis of (R)-acetoin and ethylene glycol from D-xylose through in vitro metabolic engineering. Metab. Eng. Commun. 2018, 7, e00074. [Google Scholar] [CrossRef] [PubMed]
- Sehl, T.; Hailes, H.C.; Ward, J.M.; Wardenga, R.; Von Lieres, E.; Offermann, H.; Westphal, R.; Pohl, M.; Rother, D. Two steps in one pot: Enzyme cascade for the synthesis of nor(pseudo)ephedrine from inexpensive starting materials. Angew. Chem. Int. Ed. 2013, 52, 6772–6775. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, A. Topics in Enzyme and Fermentation Biotechnology; Horwood: Chichester, UK, 1977; ISBN 9780470988961. [Google Scholar]
- Moradian, A.; Benner, S.A. A biomimetic biotechnological process for converting starch to fructose: Thermodynamic and evolutionary considerations in applied enzymology. J. Am. Chem. Soc. 2002, 114, 6980–6987. [Google Scholar] [CrossRef]
- Cassimjee, K.E.; Branneby, C.; Abedi, V.; Wells, A.; Berglund, P. Transaminations with isopropyl amine: Equilibrium displacement with yeast alcohol dehydrogenase coupled to in situ cofactor regeneration. Chem. Commun. 2010, 46, 5569–5571. [Google Scholar] [CrossRef] [PubMed]
- Bisogno, F.R.; Lavandera, I.; Kroutil, W.; Gotor, V. Tandem Concurrent Processes: One-Pot Single-Catalyst Biohydrogen Transfer for the Simultaneous Preparation of Enantiopure Secondary Alcohols. J. Org. Chem. 2009, 74, 1730–1732. [Google Scholar] [CrossRef] [Green Version]
- Jakoblinnert, A.; Rother, D. A two-step biocatalytic cascade in micro-aqueous medium: Using whole cells to obtain high concentrations of a vicinal diol. Green Chem. 2014, 16, 3472–3482. [Google Scholar] [CrossRef] [Green Version]
- Oeggl, R.; Maßmann, T.; Jupke, A.; Rother, D. Four Atom Efficient Enzyme Cascades for All 4-Methoxyphenyl-1,2-propanediol Isomers Including Product Crystallization Targeting High Product Concentrations and Excellent E-Factors. ACS Sustain. Chem. Eng. 2018, 6, 11819–11826. [Google Scholar] [CrossRef]
- Molla, G.S.; Wohlgemuth, R.; Liese, A. One-pot enzymatic reaction sequence for the syntheses of d-glyceraldehyde 3-phosphate and l-glycerol 3-phosphate. J. Mol. Catal. B Enzym. 2016, 124, 77–82. [Google Scholar] [CrossRef]
- Dirkmann, M.; Nowack, J.; Schulz, F. An In Vitro Biosynthesis of Sesquiterpenes Starting from Acetic Acid. ChemBioChem 2018, 19, 2146–2151. [Google Scholar] [CrossRef]
- Rolf, J.; Julsing, M.K.; Rosenthal, K.; Lütz, S. A Gram-Scale Limonene Production Process with Engineered Escherichia coli. Molecules 2020, 25, 1881. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Honda, K.; Sakai, T.; Okano, K.; Omasa, T.; Hirota, R.; Kuroda, A.; Ohtake, H. Synthetic metabolic engineering-a novel, simple technology for designing a chimeric metabolic pathway. Microb. Cell Fact. 2012, 11, 120. [Google Scholar] [CrossRef] [Green Version]
- Blaß, L.K.; Weyler, C.; Heinzle, E. Network design and analysis for multi-enzyme biocatalysis. BMC Bioinform. 2017, 18, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnigan, W.; Flitsch, S.L.; Hepworth, L.J.; Turner, N.J. Enzyme Cascade Design: Retrosynthesis Approach. In Enzyme Cascade Design and Modelling; Springer: Cham, Switzerland, 2021; pp. 7–30. [Google Scholar]
- Carbonell, P.; Wong, J.; Swainston, N.; Takano, E.; Turner, N.J.; Scrutton, N.S.; Kell, D.B.; Breitling, R.; Faulon, J.L. Selenzyme: Enzyme selection tool for pathway design. Bioinformatics 2018, 34, 2153–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Kamp, A.; Klamt, S. MEMO: A Method for Computing Metabolic Modules for Cell-Free Production Systems. ACS Synth. Biol. 2020, 9, 556–566. [Google Scholar] [CrossRef] [Green Version]
- Birmingham, W.R.; Starbird, C.A.; Panosian, T.D.; Nannemann, D.P.; Iverson, T.M.; Bachmann, B.O. Bioretrosynthetic construction of a didanosine biosynthetic pathway. Nat. Chem. Biol. 2014, 10, 392–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2017, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-Enzymatic Cascade Reactions: Overview and Perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Cutlan, R.; De Rose, S.; Isupov, M.N.; Littlechild, J.A.; Harmer, N.J. Using enzyme cascades in biocatalysis: Highlight on transaminases and carboxylic acid reductases. Biochim. Biophys. Acta-Proteins Proteom. 2020, 1868, 140322. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Biocatalysis-key to sustainable industrial chemistry. Curr. Opin. Biotechnol. 2010, 21, 713–724. [Google Scholar] [CrossRef]
- Núñez-López, G.; Morel, S.; Hernández, L.; Musacchio, A.; Amaya-Delgado, L.; Gschaedler, A.; Remaud-Simeon, M.; Arrizon, J. One-pot bi-enzymatic cascade synthesis of puerarin polyfructosides. Carbohydr. Polym. 2020, 247, 116710. [Google Scholar] [CrossRef]
- Fogg, D.E.; Dos Santos, E.N. Tandem catalysis: A taxonomy and illustrative review. Coord. Chem. Rev. 2004, 248, 2365–2379. [Google Scholar] [CrossRef]
- Wasilke, J.-C.; Obrey, S.J.; Tom Baker, R.; Bazan, G.C. Concurrent Tandem Catalysis. Chem. Rev. 2005, 105, 1001–1020. [Google Scholar] [CrossRef]
- Mayer, S.F.; Kroutil, W.; Faber, K. Enzyme-initiated domino (cascade) reactions. Chem. Soc. Rev. 2001, 30, 332–339. [Google Scholar] [CrossRef]
- You, C.; Zhang, Y.-H.P. Biomanufacturing by in vitro biosystems containing complex enzyme mixtures. Process Biochem. 2017, 52, 106–114. [Google Scholar] [CrossRef]
- Todd, A.E.; Orengo, C.A.; Thornton, J.M. Evolution of function in protein superfamilies, from a structural perspective. J. Mol. Biol. 2001, 307, 1113–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolf, J.; Siedentop, R.; Lütz, S.; Rosenthal, K. Screening and Identification of Novel cGAS Homologues Using a Combination of in Vitro and In Vivo Protein Synthesis. Int. J. Mol. Sci. 2019, 21, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcicchio, P.; Levisson, M.; Kengen, S.W.M.; Koutsopoulos, S.; van der Oost, J. (Hyper)thermophilic Enzymes: Production and Purification. Methods Mol. Biol. 2014, 1129, 487–496. [Google Scholar] [CrossRef] [PubMed]
- You, C.; Shi, T.; Li, Y.; Han, P.; Zhou, X.; Zhang, Y.-H.P. An in vitro synthetic biology platform for the industrial biomanufacturing of myo-inositol from starch. Biotechnol. Bioeng. 2017, 114, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Ishqi, H.M.; Husain, Q. Enzyme engineering: Reshaping the biocatalytic functions. Biotechnol. Bioeng. 2020, 117, 1877–1894. [Google Scholar] [CrossRef] [PubMed]
- Woodley, J.M. Integrating protein engineering with process design for biocatalysis. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2018, 376, 20170062. [Google Scholar] [CrossRef] [Green Version]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Improved protein structure prediction using potentials from deep learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef]
- Gao, W.; Mahajan, S.P.; Sulam, J.; Gray, J.J. Deep Learning in Protein Structural Modeling and Design. Patterns 2020, 1, 100142. [Google Scholar] [CrossRef]
- Becker, M.; Nikel, P.; Andexer, J.N.; Lütz, S.; Rosenthal, K. A Multi-Enzyme Cascade Reaction for the Production of 2′3′-cGAMP. Biomolecules 2021, 11, 590. [Google Scholar] [CrossRef]
- Claassens, N.J.; Burgener, S.; Vögeli, B.; Erb, T.J.; Bar-Even, A. A critical comparison of cellular and cell-free bioproduction systems. Curr. Opin. Biotechnol. 2019, 60, 221–229. [Google Scholar] [CrossRef]
- van Schie, M.M.C.H.; Spöring, J.D.; Bocola, M.; Domínguez de María, P.; Rother, D. Applied biocatalysis beyond just buffers—From aqueous to unconventional media. Options and guidelines. Green Chem. 2021, 23, 3191–3206. [Google Scholar] [CrossRef]
- de Gonzalo, G.; Lavandera, I.; Faber, K.; Kroutil, W. Enzymatic Reduction of Ketones in “Micro-aqueous” Media Catalyzed by ADH-A from Rhodococcus ruber. Org. Lett. 2007, 9, 2163–2166. [Google Scholar] [CrossRef]
- Savile, C.K.; Janey, J.M.; Mundorff, E.C.; Moore, J.C.; Tam, S.; Jarvis, W.R.; Colbeck, J.C.; Krebber, A.; Fleitz, F.J.; Brands, J.; et al. Biocatalytic asymmetric synthesis of chiral amines from ketones applied to sitagliptin manufacture. Science 2010, 329, 305–309. [Google Scholar] [CrossRef] [Green Version]
- Carucci, C.; Bruen, L.; Gascón, V.; Paradisi, F.; Magner, E. Significant Enhancement of Structural Stability of the Hyperhalophilic ADH from Haloferax volcanii via Entrapment on Metal Organic Framework Support. Langmuir 2018, 34, 8274–8280. [Google Scholar] [CrossRef]
- Mahour, R.; Klapproth, J.; Rexer, T.F.T.; Schildbach, A.; Klamt, S.; Pietzsch, M.; Rapp, E.; Reichl, U. Establishment of a five-enzyme cell-free cascade for the synthesis of uridine diphosphate N-acetylglucosamine. J. Biotechnol. 2018, 283, 120–129. [Google Scholar] [CrossRef] [Green Version]
- Robescu, M.S.; Serra, I.; Terreni, M.; Ubiali, D.; Bavaro, T. A Multi-Enzymatic Cascade Reaction for the Synthesis of Vidarabine 5′-Monophosphate. Catalysts 2020, 10, 60. [Google Scholar] [CrossRef] [Green Version]
- Littlechild, J.A. Enzymes from Extreme Environments and their Industrial Applications. Front. Bioeng. Biotechnol. 2015, 3, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kihumbu, D.; Stillger, T.; Hummel, W.; Liese, A. Enzymatic synthesis of all stereoisomers of 1-phenylpropane-1,2-diol. Tetrahedron Asymmetry 2002, 13, 1069–1072. [Google Scholar] [CrossRef]
- Wachtmeister, J.; Jakoblinnert, A.; Rother, D. Stereoselective Two-Step Biocatalysis in Organic Solvent: Toward All Stereoisomers of a 1,2-Diol at High Product Concentrations. Org. Process Res. Dev. 2016, 20, 1744–1753. [Google Scholar] [CrossRef] [Green Version]
- Kulig, J.; Frese, A.; Kroutil, W.; Pohl, M.; Rother, D. Biochemical characterization of an alcohol dehydrogenase from Ralstonia sp. Biotechnol. Bioeng. 2013, 110, 1838–1848. [Google Scholar] [CrossRef] [PubMed]
- Elias, M.; Wieczorek, G.; Rosenne, S.; Tawfik, D.S. The universality of enzymatic rate–temperature dependency. Trends Biochem. Sci. 2014, 39, 1–7. [Google Scholar] [CrossRef]
- Li, Z.; Li, F.; Cai, L.; Chen, Z.; Qin, L.; Gao, X.-D. One-Pot Multienzyme Synthesis of Rare Ketoses from Glycerol. J. Agric. Food Chem. 2020, 68, 1347–1353. [Google Scholar] [CrossRef]
- Korman, T.P.; Sahachartsiri, B.; Li, D.; Vinokur, J.M.; Eisenberg, D.; Bowie, J.U. A synthetic biochemistry system for the in vitro production of isoprene from glycolysis intermediates. Protein Sci. 2014, 23, 576–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mordhorst, S.; Andexer, J.N. Round, round we go—Strategies for enzymatic cofactor regeneration. Nat. Prod. Rep. 2020, 37, 1316–1333. [Google Scholar] [CrossRef] [PubMed]
- Mordhorst, S.; Siegrist, J.; Müller, M.; Richter, M.; Andexer, J.N. Catalytic Alkylation Using a Cyclic S-Adenosylmethionine Regeneration System. Angew. Chem. Int. Ed. 2017, 56, 4037–4041. [Google Scholar] [CrossRef]
- Strohmeier, G.A.; Eiteljörg, I.C.; Schwarz, A.; Winkler, M. Enzymatic One-Step Reduction of Carboxylates to Aldehydes with Cell-Free Regeneration of ATP and NADPH. Chem. Eur. J. 2019, 25, 6119–6123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardao, I.; Hwang, E.T.; Zeng, A.-P. In Vitro Multienzymatic Reaction Systems for Biosynthesis. Adv. Biochem. Eng. Biotechnol. 2013, 137, 153–184. [Google Scholar] [CrossRef]
- Woodley, J.M. Enzyme Cascade Process Design and Modelling. Enzym. Cascade Des. Model. 2021, 125–139. [Google Scholar] [CrossRef]
- Quin, M.B.; Wallin, K.K.; Zhang, G.; Schmidt-Dannert, C. Spatial organization of multi-enzyme biocatalytic cascades. Org. Biomol. Chem. 2017, 15, 4260–4271. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Wang, B.; Tan, J.; Zhu, L.; Li, L. Immobilized multienzymatic systems for catalysis of cascade reactions. Process Biochem. 2016, 51, 1193–1203. [Google Scholar] [CrossRef]
- Jia, F.; Narasimhan, B.; Mallapragada, S. Materials-based strategies for multi-enzyme immobilization and co-localization: A review. Biotechnol. Bioeng. 2014, 111, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Lozano, S.; López-Gallego, F. Wiring step-wise reactions with immobilized multi-enzyme systems. Biocatal. Biotransform. 2018, 36, 184–194. [Google Scholar] [CrossRef]
- Wiltschi, B.; Cernava, T.; Dennig, A.; Galindo Casas, M.; Geier, M.; Gruber, S.; Haberbauer, M.; Heidinger, P.; Herrero Acero, E.; Kratzer, R.; et al. Enzymes revolutionize the bioproduction of value-added compounds: From enzyme discovery to special applications. Biotechnol. Adv. 2020, 40, 107520. [Google Scholar] [CrossRef] [PubMed]
- Scherkus, C.; Schmidt, S.; Bornscheuer, U.T.; Gröger, H.; Kara, S.; Liese, A. A Fed-Batch Synthetic Strategy for a Three-Step Enzymatic Synthesis of Poly-ϵ-caprolactone. ChemCatChem 2016, 8, 3446–3452. [Google Scholar] [CrossRef]
- Mack, K.; Doeker, M.; Grabowski, L.; Jupke, A.; Rother, D. Extractive in situ product removal for the application of naturally produced L-alanine as an amine donor in enzymatic metaraminol production. Green Chem. 2021, 23, 4892–4901. [Google Scholar] [CrossRef]
- Tufvesson, P.; Fu, W.; Jensen, J.S.; Woodley, J.M. Process considerations for the scale-up and implementation of biocatalysis. Food Bioprod. Process. 2010, 88, 3–11. [Google Scholar] [CrossRef]
- Engelmann, C.; Johannsen, J.; Waluga, T.; Fieg, G.; Liese, A.; Bubenheim, P. A Multi-Enzyme Cascade for the Production of High-Value Aromatic Compounds. Catalysts 2020, 10, 1216. [Google Scholar] [CrossRef]
- Woodley, J.M. Scale-Up and Development of Enzyme-Based Processes for Large-Scale Synthesis Applications. In Biocatalysis in Organic Synthesis; Thieme Verlag: Stuttgart, Germany, 2015; ISBN 9783131746610. [Google Scholar]
- Schmidt-Traub, H.; Schulte, M.; Seidel-Morgenstern, A. Preparative Chromatography, 2nd ed.; Wiley-VCH: Weinheim, Germany, 2013. [Google Scholar]
- Mahour, R.; Marichal-Gallardo, P.A.; Rexer, T.F.T.; Reichl, U. Multi-enzyme Cascades for the In Vitro Synthesis of Guanosine Diphosphate L-Fucose. ChemCatChem 2021, 13, 1981–1989. [Google Scholar] [CrossRef]
- Česnik, M.; Sudar, M.; Hernández, K.; Charnock, S.; Vasić-Rački, D.; Clapés, P.; Blažević, Z.F. Cascade enzymatic synthesis of L-homoserine-mathematical modelling as a tool for process optimisation and design. React. Chem. Eng. 2020, 5, 747–759. [Google Scholar] [CrossRef]
- Zhong, C.; Wei, P.; Zhang, Y.-H.P. A kinetic model of one-pot rapid biotransformation of cellobiose from sucrose catalyzed by three thermophilic enzymes. Chem. Eng. Sci. 2017, 161, 159–166. [Google Scholar] [CrossRef]
- Dvorak, P.; Kurumbang, N.P.; Bendl, J.; Brezovsky, J.; Prokop, Z.; Damborsky, J. Maximizing the Efficiency of Multienzyme Process by Stoichiometry Optimization. ChemBioChem 2014, 15, 1891–1895. [Google Scholar] [CrossRef]
- Rollin, J.A.; Del Campo, J.M.; Myung, S.; Sun, F.; You, C.; Bakovic, A.; Castro, R.; Chandrayan, S.K.; Wu, C.H.; Adams, M.W.W.; et al. High-yield hydrogen production from biomass by in vitro metabolic engineering: Mixed sugars coutilization and kinetic modeling. Proc. Natl. Acad. Sci. USA 2015, 112, 4964–4969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaraja, A.A.; Charton, P.; Cadet, X.F.; Fontaine, N.; Delsaut, M.; Wiltschi, B.; Voit, A.; Offmann, B.; Damour, C.; Grondin-Perez, B.; et al. A machine learning approach for efficient selection of enzyme concentrations and its application for flux optimization. Catalysts 2020, 10, 291. [Google Scholar] [CrossRef] [Green Version]
- Finnigan, W.; Cutlan, R.; Snajdrova, R.; Adams, J.P.; Littlechild, J.A.; Harmer, N.J. Engineering a seven enzyme biotransformation using mathematical modelling and characterized enzyme parts. ChemCatChem 2019, 11, 3474–3489. [Google Scholar] [CrossRef] [Green Version]
- Bujara, M.; Schümperli, M.; Pellaux, R.; Heinemann, M.; Panke, S. Optimization of a blueprint for in vitro glycolysis by metabolic real-time analysis. Nat. Chem. Biol. 2011, 7, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erb, T.J.; Jones, P.R.; Bar-Even, A. Synthetic metabolism: Metabolic engineering meets enzyme design. Curr. Opin. Chem. Biol. 2017, 37, 56–62. [Google Scholar] [CrossRef]
- Petroll, K.; Kopp, D.; Care, A.; Bergquist, P.L.; Sunna, A. Tools and strategies for constructing cell-free enzyme pathways. Biotechnol. Adv. 2019, 37, 91–108. [Google Scholar] [CrossRef]
- Fontaine, N.; Grondin-Perez, B.; Cadet, F.; Offmann, B. Modeling of a Cell-Free Synthetic System for Biohydrogen Production. J. Comput. Sci. Syst. Biol. 2015, 8, 132–139. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Involved Reaction Steps | Substrate | Product | Main Optimization Strategy | Optimization Target | Optimization Result | Reference |
|---|---|---|---|---|---|---|
| 13 | CO2 | Malate | Pathway design, metabolic proofreading, enzyme engineering | CO2 fixation efficiency, malate production rate | 5 nmol CO2 L−1·mg−1, 1080 µM fixated CO2; 530 µM malate (20-fold improvement) | [19] |
| 5 | Mevalonate | Isoprene | Optimization of reaction conditions and balancing of enzyme levels | Flux, conversion efficiency, production rate | 6323.5 µmol·L−1·h−1 (430 mg·L−1·h−1) isoprene in 2 mL; 302 mg·L−1 isoprene in 40 h in 50 mL (83-fold improvement) | [34] |
| 12 | Chitin | Pyruvate | Flux tuning (enzyme concentrations), cofactor regeneration | Pyruvate production (titer, rate) | 2.1 mM pyruvate in 5 h; 0.42 µmol·mL−1·h−1 pyruvate (3-fold improvement) | [35] |
| 6 | d-Glucose | l-Alanine | Kinetic analysis for enzyme ratios, cosubstrate and buffer optimization | l-Alanine yield | 0.17 g l-alanine L−1·h−1 in 12 h, yield of >95% | [21] |
| 3 | 2-Ethynylglycerol | Islatravir | Retrosynthetic pathway design, directed evolution | Islatravir yield | Yield of 51% islatravir | [36] |
| 4 | Glycerol | Chiral carbohydrate precursor | Optimization of reaction conditions | Conversion | Conversion of 100% in 8 h (Starting with 42%) | [37] |
| 7 | d-Xylose | (R)-Acetoin, ethylene glycol | Optimization of reaction conditions | (R)-Acetoin production | 2.03 mM and 1.02 mM·h−1 (R)-acetoin, 3.45 mM and 1.73 mM·h−1 ethylene glycol | [38] |
| 2 | Benzaldehyde | (R)-Phenylacetyl-carbinol, (1R,2S)-norephedrine | Switch from simultaneous to sequential mode and recycling of by-product (cascade design) | Conversion, atom economy | Increase from 2% to 78% conversion | [39] |
| 3 | Benzaldehyde | Tetrahydrosiochinolines | Switch from simultaneous to multi-step reaction mode (cascade design) | Conversion | Conversion of 88% | [13] |
| 4 | Starch | Fructose | Equilibrium shift by implementation of irreversible step (cascade design) | Yield | Yield of 62% (previous work: 42% [40]) | [41] |
| 2 | Acetophenone derivatives | (S)-1-Phenylethylamine derivatives | Equilibrium shift by implementation of irreversible consumption of by-product (cascade design) | Conversion | Increase from 63% to 99% conversion | [42] |
| 2 | Racemic sec-alcohol, prochiral ketone | Enantiopure sec-alcohols | Switch to orthogonal cascade (cascade design) | Cofactor demand, atom efficiency | No additional cofactor recycling necessary | [43] |
| 2 | Acetaldehyde, benzaldehyde | 1-Phenylpropane-1,2-diol | Switch to unconventional media | Product concentration, downstream processing | Space–time yield 327 g·L−1·d−1; E factor 21.3 kgwaste·kgproduct−1 (1600-fold increase) | [44] |
| 2 | 4-Methoxy benzaldehyde, acetaldehyde | 4-Methoxyphenyl-1,2-propanediol isomers | Setting up self-sufficient recycling cascade (cascade design) | Atom economy, E factor, downstream processing | Atom economy 99.9%, E factor close to 1 kgwaste·kgproduct−1, space–time yield 165 g·L−1·d−1, isolated yield 38%, product purity 99.9% | [45] |
| Number of Involved Reaction Steps | Substrate | Product | Optimization Method | Variable Parameters | Optimization Result | Reference |
|---|---|---|---|---|---|---|
| 27 | Glucose | Terpene | Kinetic model | Enzyme concentrations | 95% yield, 15 g·L−1 titer | [33] |
| 2 | l-Alanine, 4-hydroxy-2-oxobutanoate | l-Homoserine | Kinetic model | Enzyme concentrations | 3.2 g·L−1·h−1 (18% improvement), 80.1 g·L−1 (100% improvement) | [104] |
| 5 | Xylose | α–Ketoglutarate | Kinetic model | Enzyme ratio | 98% yield (two-fold improvement) | [25] |
| 10 | Glucose | Dihydroxyacetone phosphate | Kinetic model | Enzyme ratio | 88% increase in product concentration, decrease in cofactor concentration to one-fourth | [20] |
| 3 | Sucrose | Cellobiose | Kinetic model | Enzyme ratio, enzyme concentrations | 62% yield in 10 h (ten-fold reduction in reaction time), 2.4-fold reduction of enzyme concentrations | [105] |
| 2 | d-Fructose 1,6-bisphosphate | d-Glyceraldehyde 3-phosphate, l-glycerol 3-phosphate | Kinetic model | Reactor type | 100% conversion, 10.56 g·L−1·day−1 | [46] |
| 5 | 1,2,3-Trichloropropane | Glycerol | Dynamic simulations based on Michaelis-Menten kinetics | Enzyme ratio, enzyme variants | 56% decreased enzyme loading | [106] |
| 2 | β-Hydroxypyruvate, glycolaldehyde | Erythrulose-aminotriol | Combined kinetic model and empirical model for process characterization | Enzyme ratio | 100% yield | [22] |
| 7 | Mevalonic acid | Amorpha-4,11-diene | Statistical experimental design using Taguchi orthogonal array design | Enzyme ratio | 100% yield (five-fold improvement) | [24] |
| 13 | Cellobiose | Hydrogen, carbondioxide | Multi-objective genetic algorithm | Enzyme ratio, temperature and cross-over inhibition of phosphate | 87% yield, 355 mmol·L−1·h−1 (eight-fold improvement) | [23] |
| 10 | Glucose-6-phosphate | Hydrogen | Artificial neural networks, non-linear kinetic model fitted with a genetic algorithm | Enzyme loading, temperature | 54 mmol H2 L−1·h−1 (67-fold improvement) | [107] |
| 7 | Glucose | Dihydroxyacetone phosphate | Artificial neural network | Enzyme ratio | 63% flux improvement | [108] |
| 7 | Methyl-4-toluate | 4-Tolyl alcohol | Genetic algorithm | Enzyme concentrations | 90% yield (two-fold improvement) | [109] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siedentop, R.; Claaßen, C.; Rother, D.; Lütz, S.; Rosenthal, K. Getting the Most Out of Enzyme Cascades: Strategies to Optimize In Vitro Multi-Enzymatic Reactions. Catalysts 2021, 11, 1183. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11101183
Siedentop R, Claaßen C, Rother D, Lütz S, Rosenthal K. Getting the Most Out of Enzyme Cascades: Strategies to Optimize In Vitro Multi-Enzymatic Reactions. Catalysts. 2021; 11(10):1183. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11101183
Chicago/Turabian StyleSiedentop, Regine, Christiane Claaßen, Dörte Rother, Stephan Lütz, and Katrin Rosenthal. 2021. "Getting the Most Out of Enzyme Cascades: Strategies to Optimize In Vitro Multi-Enzymatic Reactions" Catalysts 11, no. 10: 1183. https://0-doi-org.brum.beds.ac.uk/10.3390/catal11101183