
Semi-Rational Design of Proteus mirabilis l-Amino Acid Deaminase for Expanding Its Substrate Specificity in α-Keto Acid Synthesis from l-Amino Acids


Abstract
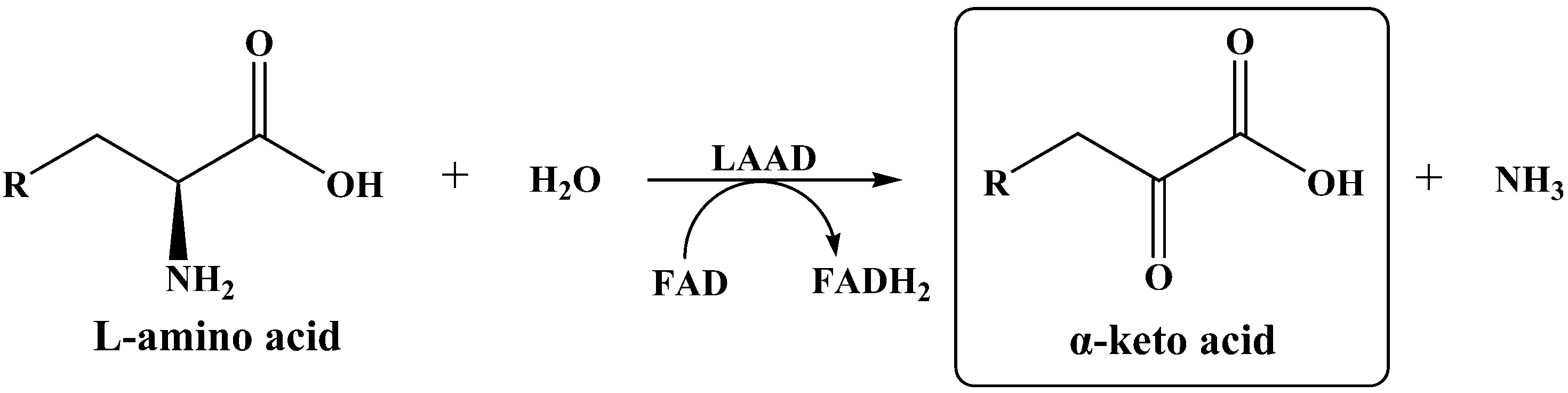
:1. Introduction
2. Results and Discussion
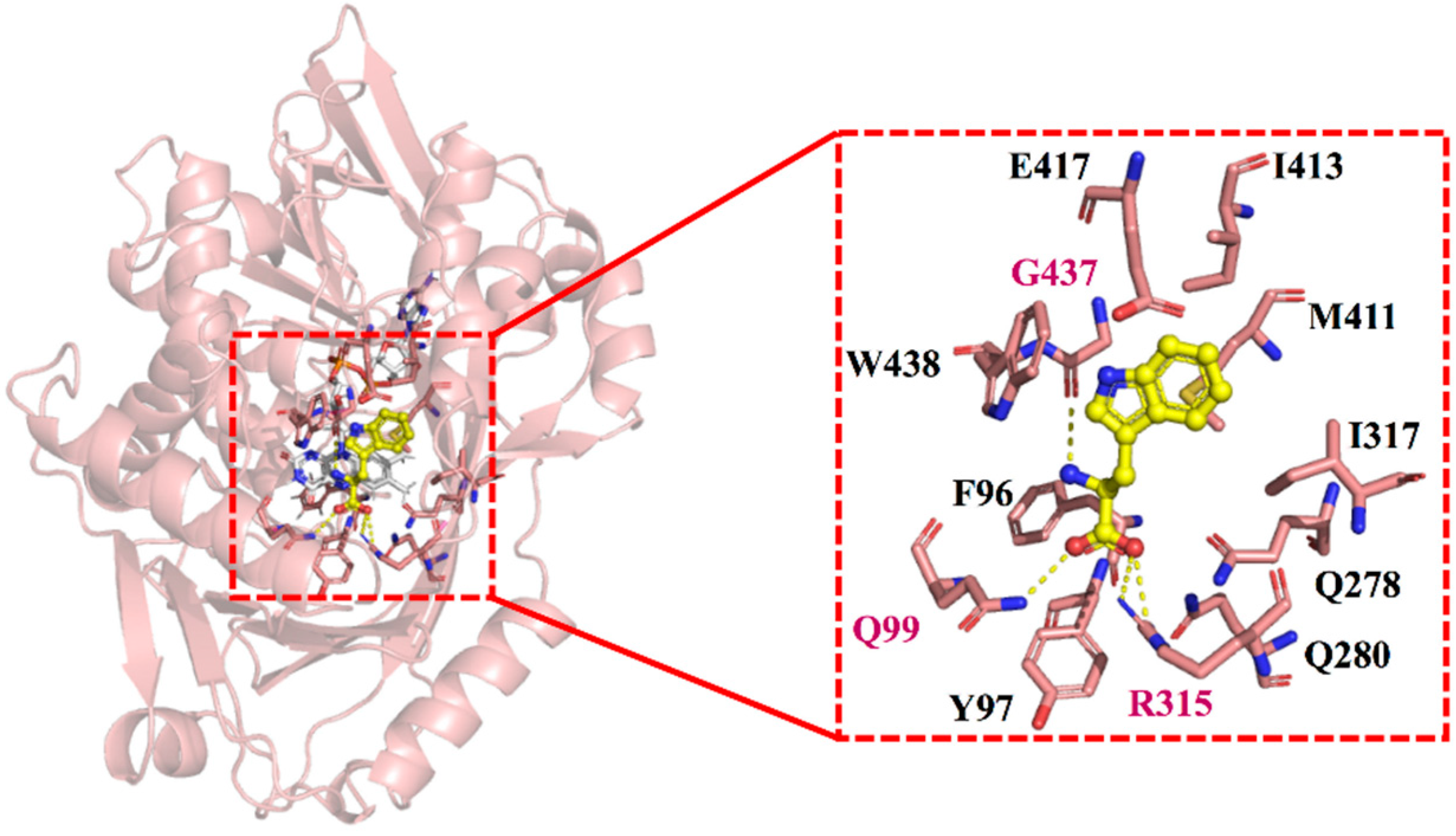
2.1. Homologous Modeling and Prediction of Functional Sites
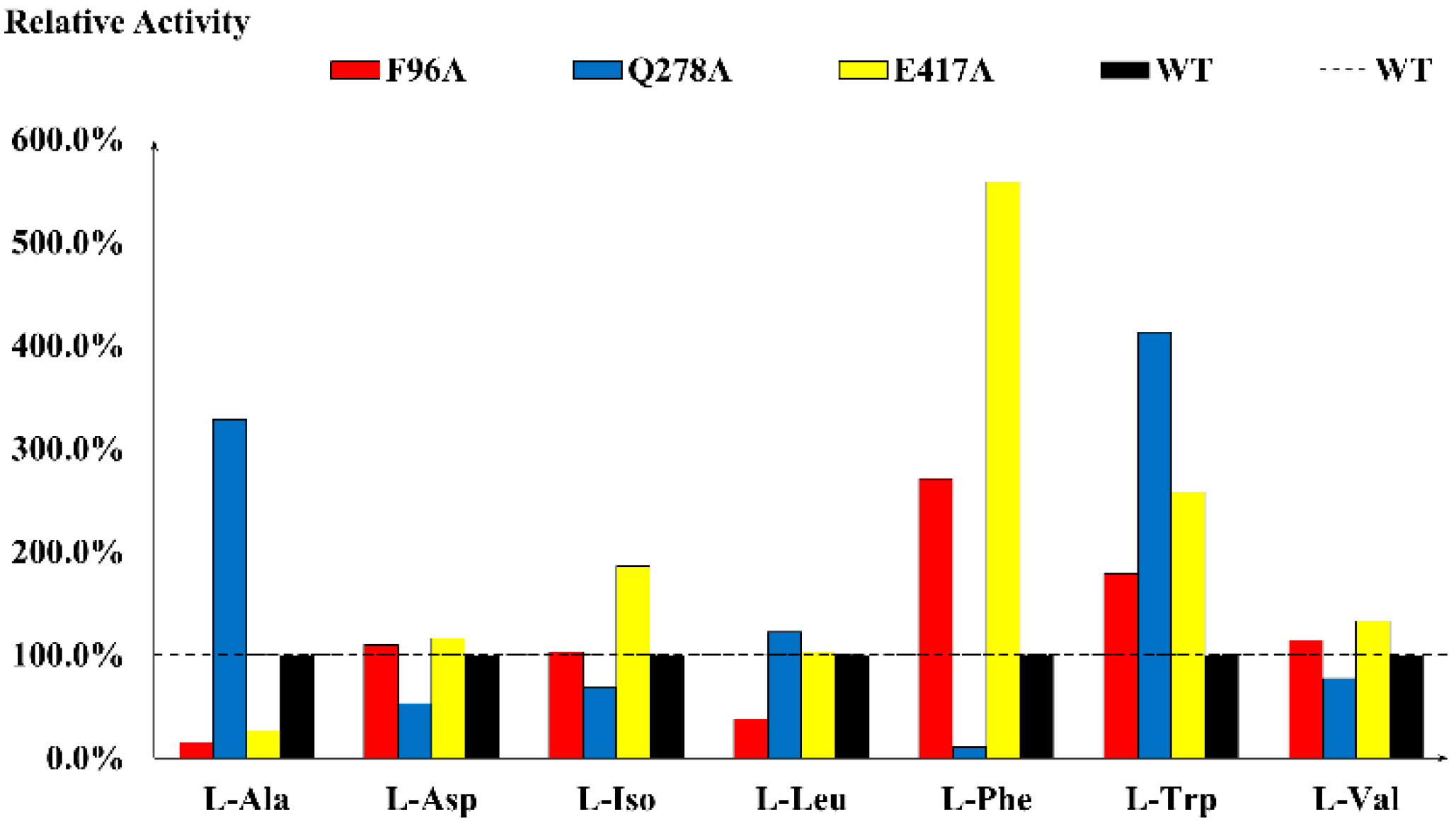
2.2. Determination of Key Residues via Alanine Scanning
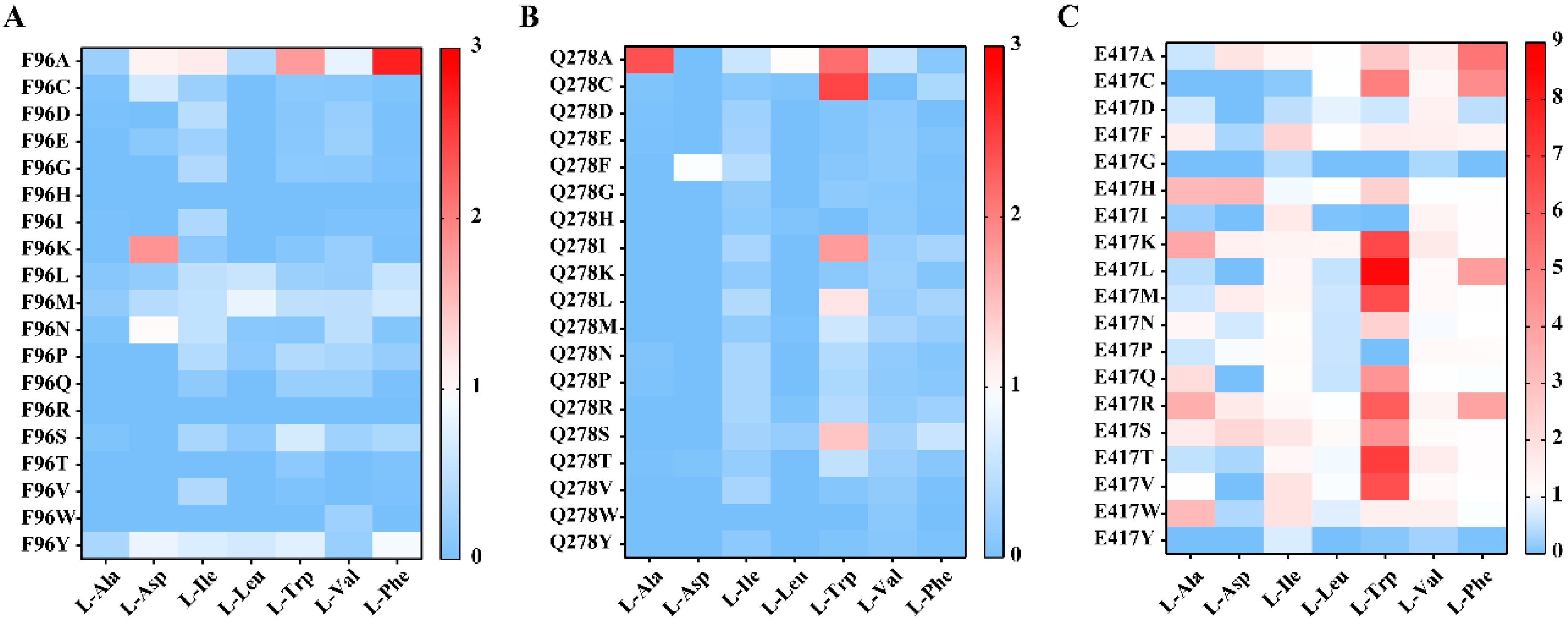
2.3. Site-Directed Saturation Mutagenesis at Potential Key Residues
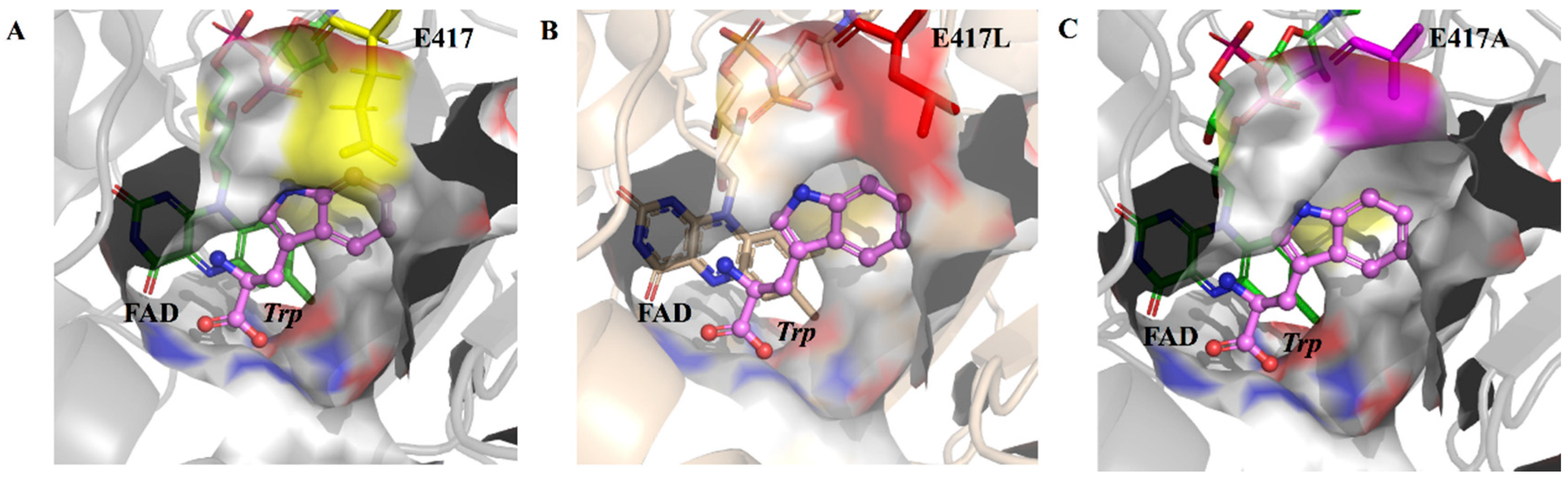
2.4. Structural Analysis of E417 and Its Mutations
3. Materials and Methods
3.1. Materials
3.2. Molecular Modeling and Docking Calculations
3.3. Alanine Scanning and Site-Directed Saturation Mutagenesis
3.4. Cell Culture and Protein Expression of PmiLAAD Mutants
3.5. Measurement of Enzymatic Activity
3.6. Determination of Kinetic Parameters
3.7. Analysis of Produced Keto Acids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Motta, P.; Molla, G.; Pollegioni, L.; Nardini, M. Structure-Function Relationships in l-Amino Acid Deaminase, a Flavoprotein Belonging to a Novel Class of Biotechnologically Relevant Enzymes. J. Biol. Chem. 2016, 291, 10457–10475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molla, G.; Melis, R.; Pollegioni, L. Breaking the mirror: l-Amino acid deaminase, a novel stereoselective biocatalyst. Biotechnol. Adv. 2017, 35, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, J.; Shin, H.-D.; Liu, L.; Du, G.; Chen, J. Biotechnological production of alpha-keto acids: Current status and perspectives. Bioresour. Technol. 2016, 219, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Okino, S.; Suda, M.; Fujikura, K.; Inui, M.; Yukawa, H. Production of d-lactic acid by Corynebacterium glutamicum under oxygen deprivation. Appl. Microbiol. Biotechnol. 2008, 78, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ma, Q.; Zhu, H. Distribution, industrial applications, and enzymatic synthesis of d-amino acids. Appl. Microbiol. Biotechnol. 2015, 99, 3341–3349. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, B.; Pan, B. Biotransformation of phenylpyruvic acid to phenyllactic acid by growing and resting cells of a Lactobacillus sp. Biotechnol. Lett. 2007, 29, 593–597. [Google Scholar] [CrossRef]
- Subhi, A.L.; Diegelman, P.; Porter, C.W.; Tang, B.; Lu, Z.J.; Markham, G.D.; Kruger, W.D. Methylthioadenosine Phosphorylase Regulates Ornithine Decarboxylase by Production of Downstream Metabolites. J. Biol. Chem. 2003, 278, 49868–49873. [Google Scholar] [CrossRef] [Green Version]
- Dilger, R.; Kobler, C.; Weckbecker, C.; Hoehler, D.; Baker, D.H. 2-Keto-4-(Methylthio)Butyric Acid (Keto Analog of Methionine) Is a Safe and Efficacious Precursor of l-Methionine in Chicks. J. Nutr. 2007, 137, 1868–1873. [Google Scholar] [CrossRef]
- Ichikawa, T.; Maeda, S.; Araki, Y.; Ishido, Y. Synthetic studies of amino acids by the use of the copper complex. I. Syntheses of. beta.-hydroxy DL-amino acids by the use of the complex of copper(II) with the Schiff base derived from glycine and pyruvic acid. J. Am. Chem. Soc. 1970, 92, 5514–5516. [Google Scholar] [CrossRef]
- Kerber, R.C.; Fernando, M.S. α-Oxocarboxylic Acids. J. Chem. Educ. 2010, 87, 1079–1084. [Google Scholar] [CrossRef]
- Escobar, J.; Frank, J.W.; Suryawan, A.; Nguyen, H.V.; Van Horn, C.G.; Hutson, S.M.; Davis, T.A. Leucine and α-Ketoisocaproic Acid, but Not Norleucine, Stimulate Skeletal Muscle Protein Synthesis in Neonatal Pigs. J. Nutr. 2010, 140, 1418–1424. [Google Scholar] [CrossRef] [Green Version]
- Mitch, W.E. Dietary therapy in uremia: The impact on nutrition and progressive renal failure. Kidney Int. 2000, 57, S38–S43. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hou, Y.; Liu, L.; Li, J.; Du, G.; Chen, J.; Wang, M. A new approach for efficient synthesis of phenyllactic acid from L-phenylalanine: Pathway design and cofactor engineering. J. Food Biochem. 2018, 42, e12584. [Google Scholar] [CrossRef]
- Ding, H.; Zhao, W.; Lü, C.; Huang, J.; Hu, S.; Yao, S.; Mei, L.; Wang, J.; Mei, J. Biosynthesis of 4-hydroxyphenylpyruvic acid from l-tyrosine using recombinant Escherichia coli cells expressing membrane bound l-amino acid deaminase. Chin. J. Chem. Eng. 2018, 26, 380–385. [Google Scholar] [CrossRef]
- Hossain, G.S.; Li, J.; Shin, H.-D.; Du, G.; Wang, M.; Liu, L.; Chen, J. One-Step Biosynthesis of α-Keto-γ-Methylthiobutyric Acid from L-Methionine by an Escherichia coli Whole-Cell Biocatalyst Expressing an Engineered L-Amino Acid Deaminase from Proteus vulgaris. PLoS ONE 2014, 9, e114291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, M. Origin, Microbiology, Nutrition, and Pharmacology of D-Amino Acids. Chem. Biodivers. 2010, 7, 1491–1530. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, S.; Martínez-Gómez, A.I.; Rodríguez-Vico, F.; Clemente-Jiménez, J.M.; Las Hera-Vázquez, F.J. Natural Occurrence and Industrial Applications of D-Amino Acids: An Overview. Chem. Biodivers. 2010, 7, 1531–1548. [Google Scholar] [CrossRef]
- Parmeggiani, F.; Lovelock, S.L.; Weise, N.J.; Ahmed, S.T.; Turner, N.J. Synthesis ofD- andL-Phenylalanine Derivatives by Phenylalanine Ammonia Lyases: A Multienzymatic Cascade Process. Angew. Chem. Int. Ed. 2015, 54, 4608–4611. [Google Scholar] [CrossRef] [Green Version]
- Parmeggiani, F.; Ahmed, S.T.; Thompson, M.P.; Weise, N.J.; Galman, J.; Gahloth, D.; Dunstan, M.S.; Leys, D.; Turner, N.J. Single-Biocatalyst Synthesis of Enantiopured-Arylalanines Exploiting an Engineeredd-Amino Acid Dehydrogenase. Adv. Synth. Catal. 2016, 358, 3298–3306. [Google Scholar] [CrossRef]
- Zhang, D.; Jing, X.; Zhang, W.; Nie, Y.; Xu, Y. Highly selective synthesis of d-amino acids from readily available l-amino acids by a one-pot biocatalytic stereoinversion cascade. RSC Adv. 2019, 9, 29927–29935. [Google Scholar] [CrossRef] [Green Version]
- Walton, C.J.W.; Parmeggiani, F.; Barber, J.E.B.; McCann, J.L.; Turner, N.J.; Chica, R.A. Engineered Aminotransferase for the Production of d -Phenylalanine Derivatives Using Biocatalytic Cascades. ChemCatChem 2018, 10, 470–474. [Google Scholar] [CrossRef]
- Wu, L.; Guo, X.; Wu, G.; Liu, P.; Liu, Z. Efficient enzymatic synthesis of α-keto acids by redesigned substrate-binding pocket of the l-amino acid deaminase (PmiLAAD). Enzym. Microb. Technol. 2020, 132, 109393. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, J.; Shin, H.-D.; Liu, L.; Du, G.; Guocheng, D. Tuning the transcription and translation of L-amino acid deaminase in Escherichia coli improves α-ketoisocaproate production from L-leucine. PLoS ONE 2017, 12, e0179229. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Sakir, H.G.; Li, J.; Shin, H.-D.; Du, G.; Chen, J.; Liu, L. Rational molecular engineering of l-amino acid deaminase for production of α-ketoisovaleric acid from l-valine by Escherichia coli. RSC Adv. 2017, 7, 6615–6621. [Google Scholar] [CrossRef] [Green Version]
- Melis, R.; Rosini, E.; Pirillo, V.; Pollegioni, L.; Molla, G. In vitro evolution of an l-amino acid deaminase active on l-1-naphthylalanine. Catal. Sci. Technol. 2018, 8, 5359–5367. [Google Scholar] [CrossRef]
- Ju, Y.; Tong, S.; Gao, Y.; Zhao, W.; Liu, Q.; Gu, Q.; Xu, J.; Niu, L.; Teng, M.; Zhou, H. Crystal structure of a membrane-bound l-amino acid deaminase from Proteus vulgaris. J. Struct. Biol. 2016, 195, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Morrison, K.L.; Weiss, G.A. Combinatorial alanine-scanning. Curr. Opin. Chem. Biol. 2001, 5, 302–307. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, S.; Song, W.; Liu, J.; Chen, X.; Hu, G.; Zhou, Y.; Liu, L.; Wu, J. Enhanced Catalytic Efficiency of L-amino Acid Deaminase Achieved by a Shorter Hydride Transfer Distance. ChemCatChem 2021, 13, 4557–4566. [Google Scholar] [CrossRef]
- Li, G.; Yao, P.; Gong, R.; Li, J.; Liu, P.; Lonsdale, R.; Wu, Q.; Lin, J.; Zhu, D.; Reetz, M.T. Simultaneous engineering of an enzyme’s entrance tunnel and active site: The case of monoamine oxidase MAO-N. Chem. Sci. 2017, 8, 4093–4099. [Google Scholar] [CrossRef] [Green Version]
- Hossain, G.S.; Li, J.; Shin, H.D.; Liu, L.; Wang, M.; Du, G.; Chen, J. Improved production of alpha-ketoglutaric acid (alpha-KG) by a Bacillus subtilis whole-cell biocatalyst via engineering of l-amino acid deaminase and deletion of the alpha-KG utilization pathway. J. Biotechnol. 2014, 187, 71–77. [Google Scholar] [CrossRef]
- Wang, X.; Nie, Y.; Xu, Y. Improvement of the Activity and Stability of Starch-Debranching Pullulanase from Bacillus naganoensis via Tailoring of the Active Sites Lining the Catalytic Pocket. J. Agric. Food Chem. 2018, 66, 13236–13242. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, G.; Han, R.; Dong, J.; Zhang, W.; Zhang, R.; Ni, Y. Carbonyl group-dependent high-throughput screening and enzymatic characterization of diaromatic ketone reductase. Catal. Sci. Technol. 2016, 6, 6320–6327. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Active Sites for Mutation | Reference |
|---|---|---|
| l-Ala | M411 | This study |
| l-Val | Y97, Q278, Q280, M411 | This study |
| l-Ile | Q278, I317, M411 | This study |
| l-Leu | Q278, I317, M411, I413, W438 | This study |
| l-Phe | Q278, I317, M411, E417 | This study |
| l-Trp | Y97, Q99, Q278, Q280, R315, I317I, M411, I413, E417, G437, W438 | This study |
| l-Asp | F96, Y97, G98, Q99, Q278, Q280, M411, G437, W438 | This study |
| l-Phe | F93, F184, P186, M394 (Proteus mirabilis) | [22] |
| l-Leu | Tuning the transcription and translation levels (Proteus vulgaris) | [23] |
| l-Met | K104, A337 (Proteus vulgaris) | [15] |
| l-Val | N100,Q276,R316,F318 (Proteus myxofaciens) | [24] |
| l-1-naphthylalanine | F318A, V412A, V438P (Proteus myxofaciens) | [25] |
| Substrate | Biocatalyst | Km (mM) | Vmax (μM·mg−1·min−1)) | kcat (s−1) | kcat/Km (mM−1·s−1) |
|---|---|---|---|---|---|
| l-Trp | E417A | 7.92 ± 1.59 | 3.25 ± 0.14 | 2.78 ± 0.12 | 0.35 |
| E417L | 7.33 ± 1.76 | 8.53 ± 0.36 | 7.31 ± 0.31 | 1.00 | |
| WT | 6.01 ± 0.92 | 1.76 ± 0.05 | 1.51 ± 0.04 | 0.25 | |
| l-Asp | E417K | 11.61 ± 2.11 | 0.94 ± 0.06 | 0.81 ± 0.05 | 0.07 |
| WT | 21.88 ± 5.20 | 1.17 ± 0.13 | 1.00 ± 0.11 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, A.; Wang, Z.; Qu, H.; Nie, Y.; Xu, Y. Semi-Rational Design of Proteus mirabilis l-Amino Acid Deaminase for Expanding Its Substrate Specificity in α-Keto Acid Synthesis from l-Amino Acids. Catalysts 2022, 12, 175. https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020175
Fan A, Wang Z, Qu H, Nie Y, Xu Y. Semi-Rational Design of Proteus mirabilis l-Amino Acid Deaminase for Expanding Its Substrate Specificity in α-Keto Acid Synthesis from l-Amino Acids. Catalysts. 2022; 12(2):175. https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020175
Chicago/Turabian StyleFan, Anwen, Ziyao Wang, Haojie Qu, Yao Nie, and Yan Xu. 2022. "Semi-Rational Design of Proteus mirabilis l-Amino Acid Deaminase for Expanding Its Substrate Specificity in α-Keto Acid Synthesis from l-Amino Acids" Catalysts 12, no. 2: 175. https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020175