CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy


1
Molecular Sciences Institute, School of Chemistry, University of the Witwatersrand, Johannesburg 2050, South Africa
2
Department of Biotechnology, Delft University of Technology, 2628 BL Delft, The Netherlands
Catalysts 2019, 9(3), 261; https://0-doi-org.brum.beds.ac.uk/10.3390/catal9030261
Submission received: 27 February 2019
/
Revised: 4 March 2019
/
Accepted: 5 March 2019
/
Published: 14 March 2019
(This article belongs to the Special Issue Biocatalysts: Design and Application)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Biocatalysis has emerged in the last decade as a pre-eminent technology for enabling the envisaged transition to a more sustainable bio-based economy. For industrial viability it is essential that enzymes can be readily recovered and recycled by immobilization as solid, recyclable catalysts. One method to achieve this is via carrier-free immobilization as cross-linked enzyme aggregates (CLEAs). This methodology proved to be very effective with a broad selection of enzymes, in particular carbohydrate-converting enzymes. Methods for optimizing CLEA preparations by, for example, adding proteic feeders to promote cross-linking, and strategies for making the pores accessible for macromolecular substrates are critically reviewed and compared. Co-immobilization of two or more enzymes in combi-CLEAs enables the cost-effective use of multiple enzymes in biocatalytic cascade processes and the use of “smart” magnetic CLEAs to separate the immobilized enzyme from other solids has raised the CLEA technology to a new level of industrial and environmental relevance. Magnetic-CLEAs of polysaccharide-converting enzymes, for example, are eminently suitable for use in the conversion of first and second generation biomass.
1. Introduction
One of the great technological challenges of the 21st century is to implement the transition from an unsustainable fossil resources-based economy to a greener and more sustainable one based on renewable biomass and utilizing manufacturing processes that minimize, or preferably avoid, the generation of waste and the use of toxic and/or hazardous materials. Biocatalysis has the right credentials to achieve this [1].
- -
- Enzymes are produced from raw materials that are inexpensive, readily available and renewable, which means that prices are not subject to large fluctuations.
- -
- Enzymes are essentially non-hazardous, non-toxic, biocompatible and biodegradable. In contrast, serious environmental costs are associated with mining precious metal catalysts and large price fluctuations are the norm. Moreover, significant costs are associated with the removal of traces of noble metals from end-products as is required by regulatory agencies in, for example, the pharmaceutical industry.
- -
- Enzymatic reactions can be performed in conventional reactors, without any need for specialized equipment, under mild conditions (ambient temperature and pressure, physiological pH) in an environmentally acceptable solvent (water).
- -
- Enzymatic reactions involving multifunctional molecules can proceed with high regio- and stereoselectivity and generally without the need for functional group activation and protection and deprotection steps. This affords synthetic routes that, compared with conventional organic syntheses, are more step economic, more energy efficient, generate less waste and provide products in exquisite stereochemical purities that are difficult to compete with.
Consequently, in the last two decades biocatalysis has emerged as an important technology for meeting the growing demand for green and sustainable processing [2,3,4]. This embraces the whole gamut of chemicals manufacture, from the enantioselective synthesis of chiral drugs [5,6,7,8,9,10,11,12] to the conversion of renewable biomass to liquid fuels and commodity chemicals [13,14,15]. This raises the perennial question: if biocatalysis is so superior why has it not been extensively used in the past? The answer is simple: thanks to advances in molecular biology and biotechnology, biocatalysis has undergone a spectacular leap forward in the last two decades:
- -
- Many more enzymes have been identified through (meta)genome mining, that is the in silico analysis of publicly accessible genome sequence data bases that have been generated as a result of next generation genome sequencing [16].
- -
- Advances in gene synthesis have enabled the synthesis of identified genes, ready for cloning into a host production organism, in a few weeks at relatively low cost. This has significantly reduced the cost of development and subsequent production of enzymes at industrial scale.
- -
- Advances in protein engineering, using directed evolution techniques [17,18], have enabled optimization of enzyme performance under the challenging conditions encountered in industrial-scale processes, namely, high (stereo)selectivities, activities and space-time yields with non-natural substrates at high substrate concentrations, in the presence of organic solvents.
In short, more enzymes are available, the enzymes are better, and, thanks to recombinant DNA technology, they can be economically produced on an industrial scale, thus enabling the design of improved industrial biocatalysts [19]. Notwithstanding these numerous benefits of using enzymes, their industrial application is often hampered by a lack of long-term stability and difficult recovery and re-use. However, for both economic and environmental viability, these valuable catalysts need to be efficiently recovered and recycled and this is accomplished using immobilization technologies [20,21].
2. Enzyme Immobilization
The fact that enzymes are soluble in water means that recovery from aqueous waste streams is challenging. This hurdle can be overcome by immobilizing the enzyme as a solid, heterogeneous catalyst that can be separated by filtration or centrifugation. In addition to facilitating the recovery and re-use of enzymes, immobilization generally results in an increase in their storage and operational stability since it decreases the flexibility of the enzyme, reducing its propensity to unfold and denature under the influence of heat, organic solvents or by autolysis. Improved performance is reflected in a higher catalyst productivity (kg product/kg enzyme) and a lower enzyme cost contribution. Furthermore, immobilization can facilitate their use in continuous flow processing [22,23]. In short, immobilization can enable applications that would not be economically viable with the free enzyme [24,25,26,27,28,29,30,31]. Nonetheless, the majority of biocatalyst applications still involve a single use of water-soluble enzymes, i.e., on a throw away basis, which is not cost effective and not in line with the concept of a circular economy.
Since most biocatalytic processes are performed under roughly the same conditions of (ambient) temperature and pressure, multiple biocatalytic steps can be integrated into enzymatic cascade processes [32] and co-immobilization of two or more enzymes affords multifunctional solid catalysts capable of catalyzing such cascade processes [33].
Methods for immobilization can be conveniently divided into three types: (i) immobilization on a prefabricated support (carrier) typically an organic resin or silica, (ii) entrapment in a polymeric matrix that is generated in the presence of the enzyme and (iii) cross-linking of enzyme molecules. Binding to a carrier can involve simple adsorption, e.g., via hydrophobic or ionic interactions or actual covalent bonding [34]. The simplest method is to physically adsorb the enzyme on a carrier and can be successful in organic media [35]. However, the enzyme will readily be leached from the surface in the presence of water [35]. A serious drawback of carrier-bound enzymes [36] is their low productivities (kgs product per kg enzyme) owing to the large amount of non-catalytic ballast (generally 90 to 99% of the total mass). Moreover, the cost contribution of the carrier is often substantially higher than that of the enzyme. In contrast, immobilization by cross linking of enzyme molecules affords carrier-free immobilized enzymes with high productivities and avoids the costs of a carrier.
Usually some loss of activity is observed on immobilization but the increase in stability coupled with reusability, affording dramatic cost reductions compared with the soluble enzyme, more than compensates for this. Two of the most important industrial enzymes in use today are glucose isomerase and penicillin G amidase (PGA) [27]. Roughly 500 tons of immobilized glucose isomerase (EC 5.3.1.18) are used annually to produce ca. 106 tons of high fructose corn syrup (HFCS), corresponding to catalyst productivities of 10,000 kgs per kg. Similarly, PGA is used in the production of more than 20,000 tons per annum of 6-amino penicillanic acid (6-APA), by enzymatic hydrolysis of penicillin G, and the manufacture of more than 10,000 tons of semi-synthetic penicillin and cephalosporin antibiotics annually [37]. Its use in an immobilized form is an indispensable condition for commercial viability and productivities of 600 kg per kg or more are the norm [38]. Interestingly, the production of both immobilized enzymes involves a cross-linking step with glutaraldehyde. The latter is generally the cross-linker of choice as it is inexpensive, readily available and its use in cross-linking of proteins has GRAS (generally regarded as safe) status for use in, among other things, food and beverages processing [39].
Turner and coworkers [40] recently outlined the requirements that an immobilized enzyme should meet for commercial viability of a process in the pharmaceutical industry: (i) high enzyme loading (10 wt%), (ii) high activity recovery (50%), (iii) no leaching under reaction conditions, (iv) tolerance to organic solvents, (v) recyclable (20 cycles), (vi) good substrate mass transport and (vii) mechanically stable in batch and flow reactors and (viii) last but not least, it must result in a lower cost of goods compared with alternatives. The authors also pointed out that a method should have broad applicability over numerous enzyme classes and, as an absolute minimum, generality within the same enzyme class. However, we note that while generality may be a positive attribute it has no influence on the cost-effectiveness of a method.
3. Cross-Linked Enzyme Aggregates (CLEAs)
Cross-linked enzymes (CLEs) were already known in the 1960s. They were formed by mixing a solution of an enzyme in aqueous buffer with an aqueous solution of glutaraldehyde. They were difficult to handle, gelatinous materials exhibiting low activity, poor reproducibility and low stability and shelf life. Consequently, carrier-bound enzymes became the norm for the next three decades. In the early 1990s Altus Biologics introduced the use of cross-linked enzyme crystals (CLECs) as industrial biocatalysts [41]. The methodology was broadly applicable and CLECs exhibited excellent operational stability and controllable particle size coupled with high productivity and facile recovery and re-use, making them ideally suited for applications as industrial biocatalysts. However, they had one inherent limitation: the need to crystallize the enzyme, a laborious and costly procedure requiring enzyme of high purity.
We surmised that crystallization could be replaced by precipitation of the enzyme from aqueous buffer, a simpler and less expensive method not requiring high purity enzymes. It was well-known that the addition of salts, or water miscible organic solvents or non-ionic polymers, to aqueous solutions of proteins leads to their precipitation as physical aggregates that are held together by non-covalent bonding, without perturbation of their tertiary structure, and redissolve on the addition of water. We showed that addition of a bifunctional cross-linker rendered them permanently insoluble, as what is essentially a cross-linked polymer, while maintaining their pre-organized superstructure, and, hence their catalytic activity. The result was a new class of immobilized enzymes for which we coined the term cross-linked enzyme aggregates (CLEAs). The first publication [42] appeared in 2000 and the technology was commercialized by the industrial biotech company, CLEA Technologies B.V., founded in 2002 as a spin-out of the Biocatalysis and Organic Chemistry Group at Delft University of Technology.
Since selective precipitation with ammonium sulfate is commonly used to purify enzymes, the CLEA methodology essentially combines enzyme purification and immobilization into a single unit operation. This enables cost effective production of the CLEA from crude cell lysate obtained from fermentation broth, without the need for intermediate work-up or purification. The CLEA technology has very broad scope. It has been successfully applied to the immobilization of countless enzymes from the five enzyme classes—hydrolases, oxidoreductases, lyases, transferases and isomerases—used in chemicals manufacture and forms the subject of an increasing number of reviews [43,44,45,46].
3.1. Cross-Linked Enzyme Aggregate (CLEA) Preparation
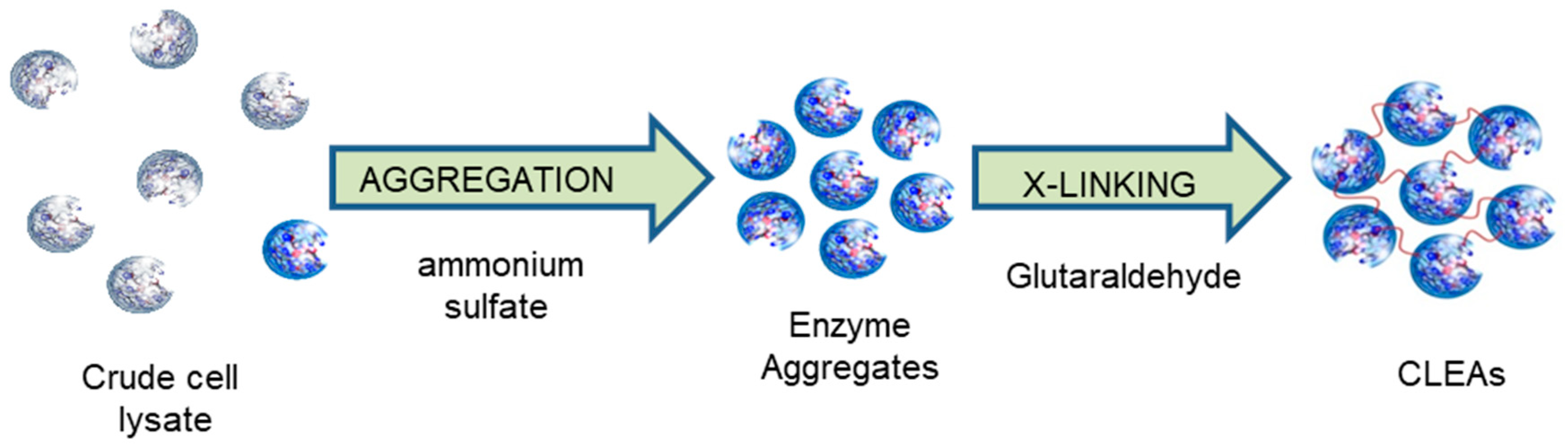
The preparation of CLEAs involves (a) precipitation as physical aggregates from a solution in aqueous buffer and (b) cross-linking of the physical aggregates with a bifunctional cross-linking agent (Figure 1). The precipitation step is instantaneous and much faster than the cross-linking step. Therefore, the precipitant (aggregant) and the cross-linker can be added simultaneously, combining the two steps into one operation.
The precipitant can be a salt, such as ammonium sulfate or sodium sulfate, a water-miscible organic solvent or a non-ionic polymer, such as polyethylene glycol (PEG). In practice, ammonium sulfate works well with almost all enzymes and is generally the precipitant of choice. Water miscible organic solvents, on the other hand, are generally less environmentally attractive.
As with cross-linking of proteins in general, glutaraldehyde is generally the cross-linker of choice. Although the chemistry is complex and not fully understood, cross-linking involves reaction of free amino groups on the surface of neighbouring enzyme molecules, with oligomers or polymers of glutaraldehyde, and is usually performed at pH 7.0 to 9.0. With some enzymes, e.g., nitrilases, low or no activity recovery was observed with glutaraldehyde cross-linking. We reasoned that it could be caused by glutaraldehyde penetrating the interior of the protein and reacting with amino groups crucial for the activity of the enzyme. We found that cross-linking with dextran polyaldehyde, a bulky polyaldehyde that is not able to penetrate the interior of the enzyme, followed by reduction of the Schiff’s base moieties to form irreversible amine linkages, could be used in such cases [47]. Other cross-linkers, derived from readily available polysaccharides, have also been used, e.g., pectin dialdehyde [48]. More recently, the use of a readily available bis-epoxide, glycerol diglycidyl ether as a cross-linker was described [49].
The molar ratio of cross-linker to enzyme influences the activity, stability and particle size of the resulting CLEA. If the ratio is too low sufficient cross linking doesn’t occur, affording a CLEA that is too flexible and unstable towards leaching in water. If it is too high, excessive cross linking can cause a complete loss of the enzyme’s flexibility resulting in a low activity recovery. The rate of addition of the cross-linker and the cross-linking time also influence the activity recovery and particle size and morphology of CLEAs. The particle size of a CLEA can also increase on ageing [50] strongly suggesting that the cross-linking process was not complete.
3.2. Effect of Additives
As would be expected, the efficacy of cross-linking is influenced by the availability of free amino groups, usually of Lys residues, on the surface of the enzyme and is problematical with electronegative enzymes containing a paucity of free amino groups. CLEAs derived from such enzymes tend to be mechanically fragile and suffer from facile enzyme leaching [51]. The problem can generally be overcome by co-aggregation with polymers containing numerous amino groups. For example, co-aggregation with polyethyleneimine (PEI) affords CLEAs with high activity recoveries and excellent operational stabilities [52,53,54]. Similarly, pentaethylene-hexamine [55], and polylysine [56] have been used to good effect. Alternatively, co-aggregation with so-called proteic feeders rich in Lys residues, such as bovine serum albumin (BSA), led to remakable improvements in activity recovery and operational stabilities of a lipase and penicillin amidase [57,58]. Hen egg white was also an effective proteic feeder [59] and, more recently, feather meal, an inexpensive poultry byproduct rich in protein, was used as a novel proteic feeder in the preparation of combi-CLEAs of a mixture of pectinase, pectin lyase, pectine methyl esterase and polygalacturonase, for use in grape juice clarification [60].
4. Scope of CLEA Technology
CLEA technology has broad scope and, over the last two decades, an increasingly wide selection of hydrolases, oxidoreductases, lyases, transferases and isomerases has been successfully used in organic synthesis in general [61], and asymmetric synthesis in particular [62].
4.1. Hydrolase CLEAs
The majority of CLEAs described in the literature involve hydrolases, probably because they are the enzymes that have the most industrial applications and are the simplest enzymes to work with. These include CLEAs from lipases [63], proteases [64], esterases [65,66], amidases [67], nitrilases [68], epoxide hydrolases [69], and glycosidases.
CLEAs have been prepared from numerous lipases, including Candida Antarctica lipase B (CaLB) [70,71], Thermomyces lanuginose [72], Penicillium expansum [73], Penicillium notatum [74] and Candida rugosa [75], to name but a few. A CLEA was also produced from a perhydrolase [76] and used to catalyze the in situ generation of a peracid in a Baeyer–Villiger oxidation.
CLEAs have been prepared from a variety of proteases. In particular, the CLEA of alcalase (E.C. 3.4.21.62), the inexpensive alkaline protease from Bacillus licheniformis (also known as subtilisin Carlsberg) used in laundry detergents, has been widely used in resolutions of amino acid esters and amines and peptide synthesis [77]. Penicillin G amidase (PGA) CLEAs were shown to be extremely effective catalysts, with high volumetric productivities, for the hydrolysis of penicillin G and the synthesis of semi-synthetic penicillins [78] and cephalosporins [79] such as ampicillin and cephalexin, respectively.
In our experience, over almost two decades CLEAs have been exceptionally suited to carbohydrate conversion processes. The number of enzymes acting on carbohydrates is vast with numerous commercially important applications, not only in food and beverages processing [80] but also in pharmaceuticals, nutraceuticals, cosmetic ingredients, and biomass conversion. The latter is of increasing importance in the context of the bio-based economy (see later).
CLEAs have been successfully prepared from a variety of glycosidases (E.C.3.2.1.) which catalyze the formation of ether bonds between two carbohydrate molecules or a carbohydrate and an aglycon. For example, an Aspergillus oryzae β-galactosidase CLEA was applied successfully to the synthesis of galacto-oligosaccharides (GOS) [81], commercially important prebiotic food additives. More recently, a CLEA of a novel β-galactosidase from a marine metagenomic library was described [82]. In comparison with the free enzyme, the CLEA displayed a slightly higher GOS yield, a much higher thermostability, good storage stability, better galactose tolerance and 82% activity retention after 10 cycles.
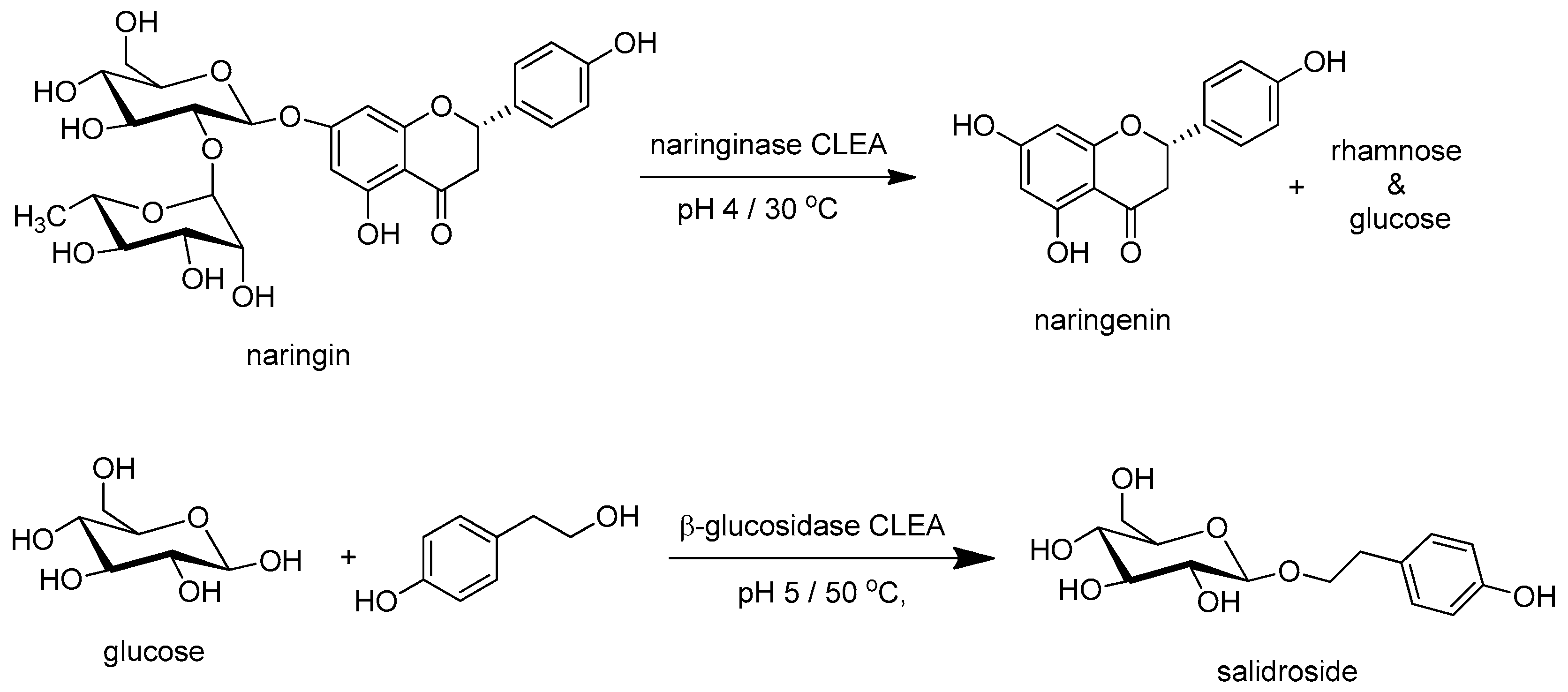
Glycoside CLEAs have also been used in the hydrolysis and synthesis of glycosides with aglycons. Thus, a CLEA of Penicillium decumbens naringinase (E.C. 3.2.1.40) catalyzed the hydrolysis of the glycoside naringin into naringenin (Figure 2 [83]). A CLEA of the β-glucosidase (E.C. 3.2.1.20) from Prunus domestica seeds catalyzed the synthesis of p-hydroxyphenethyl α-d-glucopyranoside, salidroside that is used in Chinese traditional medicine. The CLEA had a much higher productivity and improved thermal stability compared to the free enzyme (Figure 2) [84].
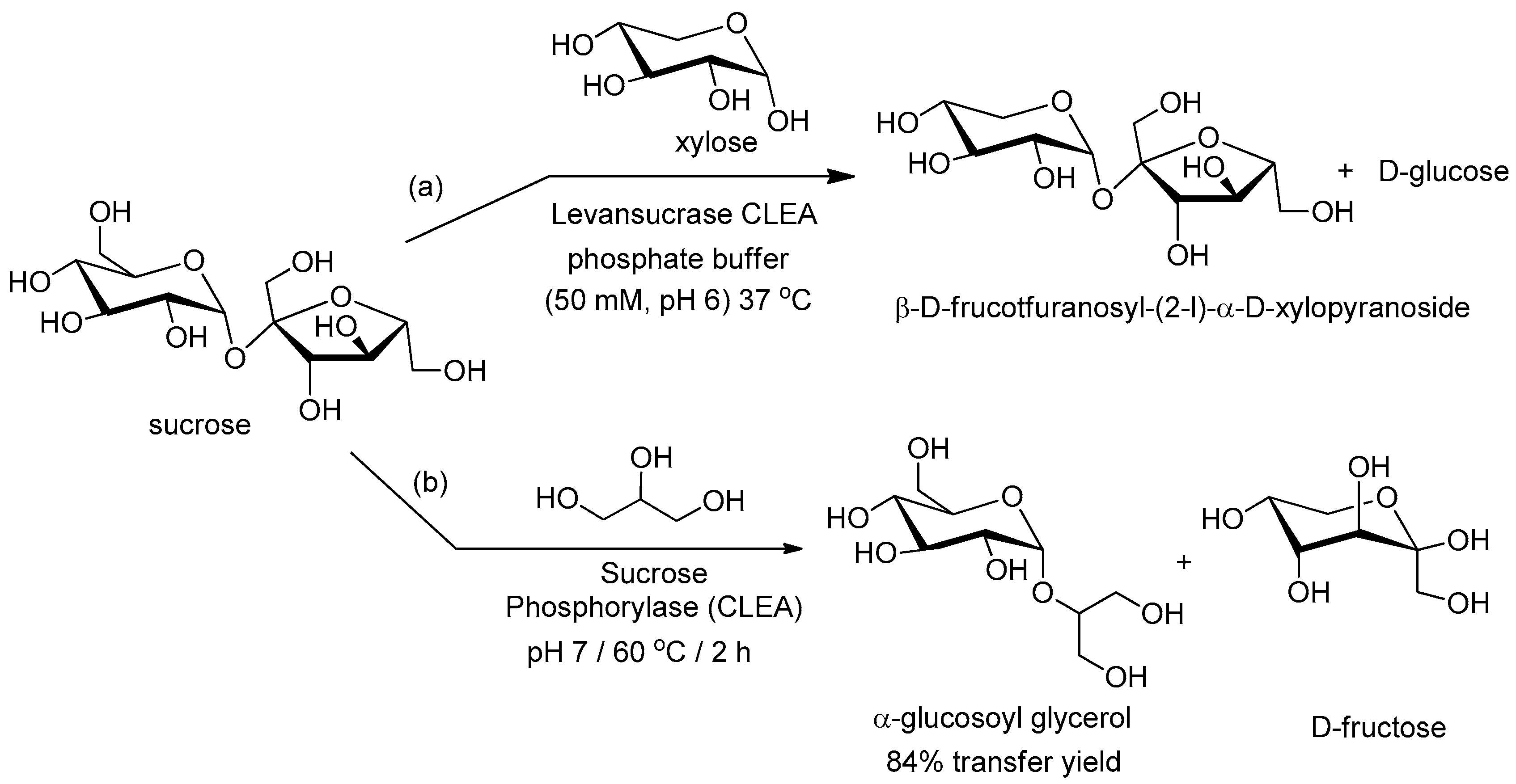
Levansucrase (E.C. 2.4.1.10) catalyzes the synthesis of levan from sucrose and the transfer of fructosyl moieties from sucrose to other acceptor molecules such as xylose (Figure 3a). A levansucrase-CLEA catalyzed the synthesis of the oligofructosides and transfructosylations as efficiently as the free enzyme and showed the highest reported specific activity for fructosyl- and glucosyl-transferases [85].
Sucrose phosphorylase (E.C. 2.4.1.7) catalyzes the reversible phosphorylation of sucrose into a mixture of fructose and d-glucose-1-phosphate (G1P). Soetaert and coworkers [86] showed that a CLEA of sucrose phosphorylase from Bifidobacterium adolescentis had a temperature optimum of 75 °C compared with 58 °C for the free enzyme. This was accompanied by an increase in the operational stability of the CLEA which retained full activity after 1 week at 60 °C compared with 20% loss of activity in 16 h with the free enzyme. Furthermore, the CLEA was recycled at least 5–10 times without loss of activity. Sucrose phosphorylase also catalyzes transglucosylations of carbohydrates and non-carbohydrate molecules such as glycerol (Figure 3b) [87]. The CLEA was extremely stable and eminently suitable for operation at 60 °C.
Similarly, a Saccharomyces cerevisiae invertase-CLEA exhibited an optimal operational temperature of 70 °C, compared with 40 °C for the free enzyme, in the hydrolysis of sucrose to a mixture of fructose and glucose (inverted sugar syrup) [88].
4.2. Oxidoreductase and Lyase CLEAs
CLEAs have been prepared from a variety of oxidoreductases. These include oxidases such as glucose oxidase (GOX), galactose oxidase (GOase), laccase [89,90], monoamine oxidase (MAO) [91] peroxidases [55,92,93,94] and dehydrogenases such as alcohol dehydrogenases/ketoreductases (KREDs), and formate dehydrogenase [95]. Laccase-CLEAs have been extensively studied because of the interest in using them for the removal of dyes, pharmaceutical residues and endocrine disruptors such as bisphenol A, from waste water [96,97,98].
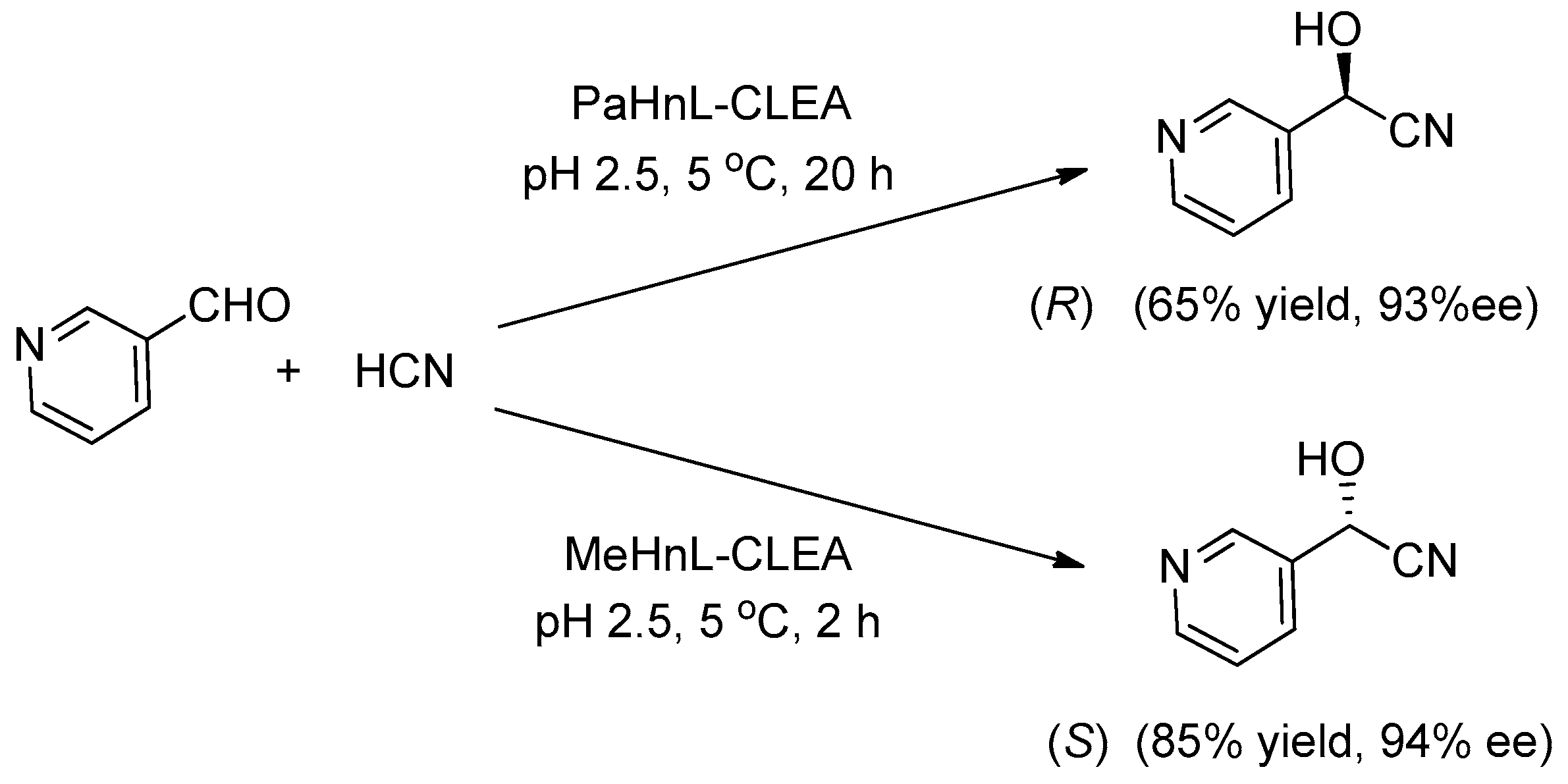
Hydroxynitrile lyases (HNLs; EC 4.1.2.10) [99] are good examples of lyases that have been successfully ‘cleated’. HNL-CLEAs are robust, highly effective catalysts for enantioselective hydrocyanation of aldehydes under microaqueous conditions and can be recycled several times without loss of activity. They have been prepared from inter alia, Prunus amygdalis [47], Prunus dulcis [100,101], Hevea brasiliensis [102], Manihot esculenta [103] and Linus usitatissimum (LuHNL, E.C. 4.1.2.46) [104]. HNL-CLEAs perform exceptionally well in organic solvents, essentially completely suppressing competing non-enzymatic hydrocyanation and can afford enantioselectivities that are not possible with the free enzyme (Figure 4) [105].
Because of the industrial importance of HNLs, they are in constant demand and when new HNLs are discovered they are usually ‘cleated’ to render them suitable for operation in organic solvents. For example, new HNLs from Davallia tyermannii [106] and Baliospernum montanum [107] were converted to CLEAs to afford robust, recyclable catalysts for the synthesis of chiral cyanohydrins.
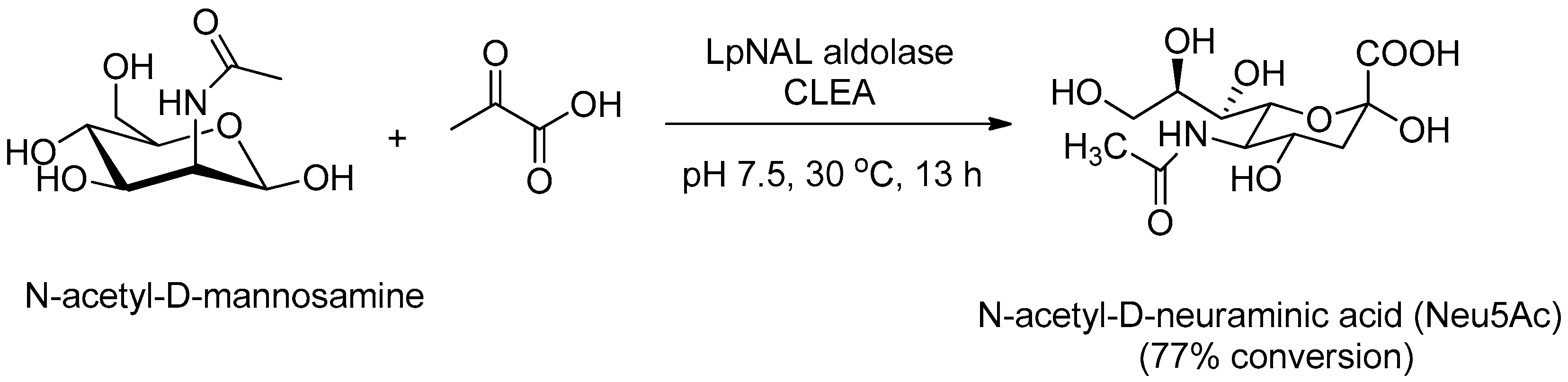
N-acetyl-d-neuraminic acid aldolase (NAL, E.C. 4.1.3.3) is used industrially at basic pH for the condensation of pyruvate and N-acetyl-d-mannosamine (ManNAc) into N-acetyl-d-neuraminic acid (Neu5Ac), an advanced intermediate for GSK’s antiviral drug Relenza® (zanamivir). Garcia-Garcia et al. [108,109] prepared a robust CLEA from the N-acetyl-d-neuraminic acid aldolase from Lactobacillus plantarum (LPNAL, E.C. 4.1.3.3) by ammonium sulfate co-precipitation with BSA and cross-linking with glutaraldehyde and applied it in the chemo-enzymatic synthesis of Neu5Ac (Figure 5), which could ultimately be made from the inexpensive N-acetyl-d-glucosamine. The CLEA exhibited good activity and operational stability at alkaline pH and at least 10 cycles were possible without any significant loss of activity.
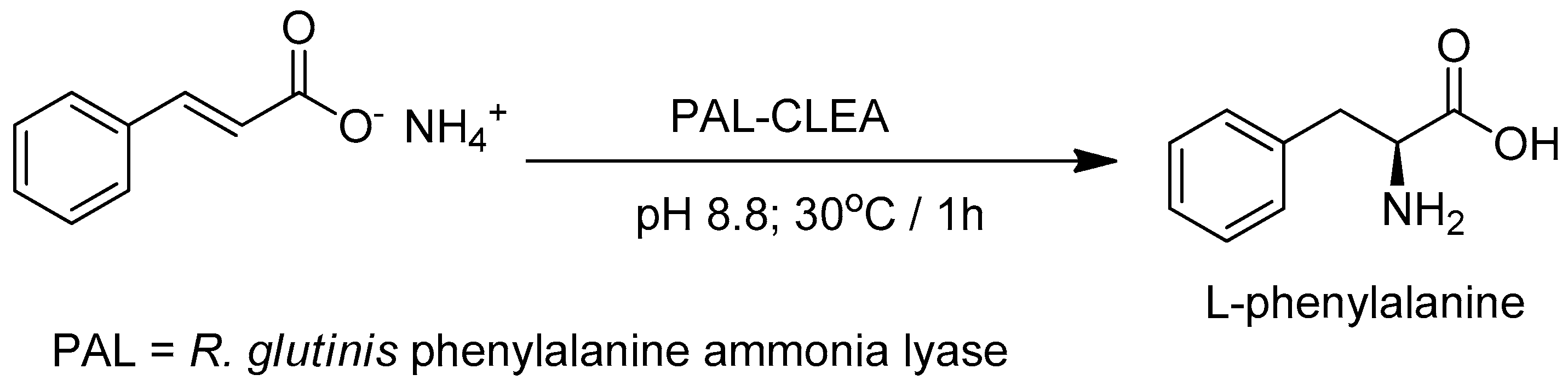
Cui and coworkers [110] prepared CLEAs of phenylalanine ammonia lyase (PAL; EC 4.3.1.5) from R. glutinis which catalyzes the industrially important formation of l-phenylalanine by asymmetric addition of ammonia to cinnamic acid (Figure 6). Compared to the free enzyme the PAL-CLEA had increased storage and operational stability and was recycled 12 times without appreciable loss of activity. More recently, the same group [111] prepared PAL-CLEA by coaggregation with bovine serum albumin and obtained CLEAs that were even more stable towards heat and organic solvents.
5. Advantages, Limitations and Optimization of CLEAs
An important advantage of immobilized enzymes in general, certainly from a cost-effectiveness viewpoint, is their ease of recovery and re-use, as heterogeneous catalysts, by filtration, centrifugation or, alternatively, in a fixed bed reactor. CLEAs are eminently recyclable. For example, the (R)-hydroxynitrile lyase (HNL)-CLEA from Prunus amygdalis which catalyzed the enantioselective hydrocyanation of o-chlorobenzaldehyde was recycled 10 times with neglible loss in activity [114].
In addition to facilitating its recovery and reuse, another important reason for immobilizing an enzyme is the significant enhancement of operational stability towards heat and organic solvents that is generally observed. Indeed, immobilization of enzymes as cross-linked enzyme aggregates can lead to dramatic increases in storage and operational stability. For example, the operational stability of a papain-CLEA, was dramatically enhanced compared to the limited thermal stability of free papain [45].
Many enzymes consist of multimers that are characterized by limited thermal stability outside the cell owing to their facile dissociation with concomitant loss of activity. CLEA formation can stabilize multimers by suppressing this dissociation through cross-linking. For example, CLEA preparation was used successfully to stabilize two tetrameric catalases [115]. The industrially important nitrile hydratases (NHases) are also multimers and notoriously unstable outside the cell. Consequently, NHase mediated industrial biotransformations, such as acrylamide and nicotinamide manufacture from the corresponding nitriles, are conducted as whole cell processes. In contrast, immobilization of a nitrile hydratase from an alkaliphilic bacterium, Nitriliruptor akaliphilus, as a CLEA led to a dramatic increase in storage stability compared to the free enzyme [113]. In the hydration of n-hexyl cyanide, the NHase-CLEA was recycled 36 times with a minor loss in activity that was largely attributed to mechanical losses that inevitably occurred on pipetting the supernatant from the solid CLEA. The industrially important enzyme, halohydrin dehalogenase (HHDH) is a homotetramer and is active only as the tetramer. It was recently shown that immobilization of HHDH as a CLEA occurred with 90% activity recovery and the resulting HHDH-CLEA exhibited a strong tolerance to organic solvents and retained 70% activity after 10 recycles [116].
Since the enzyme molecules in CLEAs are bound by covalent bonds, there is no leaching of enzyme observed in aqueous media, even under drastic conditions such as in the presence of surfactants. A further advantage, of immobilized enzymes in general, and CLEAs in particular, is that they exhibit no or low allergenicity since they cannot easily penetrate the skin.
Another important cost benefit is the fact that CLEAs can be prepared from enzyme samples of low purity such as crude cell lysate, obtained from fermentation broth. Furthermore, because they consist mainly of active enzyme, CLEAs exhibit high catalyst productivities (kg product per kg of enzyme) compared to carrier-bound enzymes and the costs of the carrier ballast are avoided.
The CLEA technology is broadly applicable but, since every enzyme is a different molecule, the protocol has to be optimized for every enzyme. This can be seen as a limitation but, in practice, optimizing the aggregation plus cross-linking protocol is not a lengthy procedure and can be readily automated [117]. The response surface methodology (RSM) has been successfully used, for example, in the optimization of the preparation of a CaLB-CLEA 68, a lipase CLEA from cocoa pod husk [118] and a protease CLEA [64]. The optimization of CLEA preparation has been reviewed by several authors [119,120,121].
Particle size is an important property of any heterogeneous catalyst since it directly affects mass transfer and filterability under operational conditions. Obviously, the rate of diffusion is influenced by the particle size and this is determined by, inter alia, the cross linker/enzyme ratio. A typical particle size of CLEAs is 5–50 μm and their filterability is generally sufficient for batch operation. CLEAs are highly porous materials and diffusional limitations are generally not observed when using them in biocatalytic transformations commonly used in organic synthesis. Optimum rates are observed with smaller particles but practical considerations, e.g., ease of filtration, necessitate the use of larger particles and in practice a compromise between good activity and good filterability has to be accepted.
During CLEA formation the enzyme molecules are packed together in a small volume which results in a relatively small pore size that can cause diffusion limitations with macromolecular substrates, such as polysaccharides and proteins. Moreover, washing and centrifugation can lead to further compression of the particles. Consequently, so-called porous CLEAs (p-CLEAs), with increased porosity, were developed and shown to have better internal mass transfer [122,123,124]. It involved adding starch to the enzyme solution prior to precipitation and cross-linking. After glutaraldehyde cross-linking α-amylase is added to hydrolyze the starch. The resulting p-CLEAs exhibited improved mass transfer. In a variation on this theme, porous β-mannanase CLEAs were prepared by using linear dextran polyaldehyde as a macromolecular cross-linker [125]. An alternative approach for circumventing compression of CLEAs during centrifugation or filtration is to use ‘smart’ magnetic CLEAs and separate them with a permanent magnet (see later). Moreover, the use of magnetic CLEAs enables the use of small, highly active particles without compromising the ability to recover and reuse the CLEA (see later).
In a novel refinement on the cross-linking theme, Brady and coworkers [126,127] prepared Spherezymes by adding cross-linking agents to a water-in-oil emulsion of an aqueous solution of P. fluorescens lipase. The spherezyme particles exhibited good activity in organic solvents.
5.1. Bioimprinting
In the early days of the CLEA technology we showed [128] that the addition of crown ethers or surfactants to solutions of lipases, prior to aggregation and cross-linking, could result in the formation of CLEAs with a higher activity than the free enzyme. This was consistent with the idea that the surfactant or crown ether induced the lipase to adopt a different conformation that could be more favourable. Subsequent cross-linking could then ‘lock‘ the enzyme in this favourable conformation and the additive could simply be removed by washing. This additive effect was later extended to the idea of using molecular “bioimprinting“ to influence the activity and/or substrate specificity of CLEAs. For example, addition of butanone to an HNL prior to aggregation and cross-linking led to the formation of an HNL-CLEA with improved synthetic activity towards butanone as substrate [104]. More recently, it was shown that the specificity of of sucrose phosphorylase towards α-glucosyl glycerol as substrate increased two-fold by prior imprinting with the substrate [87].
5.2. Reactor Configurations
The choice of reactor configuration is intimately connected with downstream processing and is largely influenced by the industry segment and characteristic product volumes. In the pharmaceutical and fine chemical industries product volumes are relatively small and processes are generally conducted in batch in multi-purpose stirred tank reactors (STRs), equipped with a propeller stirrer. The catalyst is recovered by filtration or centrifugation. A problem that is generally encountered in the use of STRs is mechanical attrition of the immobilized enzyme, resulting from shear forces caused by the propeller stirrer, leading to the formation of difficult to handle pulverized particles.
Several concepts have been employed to address the attrition problem. For example, in a membrane slurry reactor (MSR) the immobilized enzyme is retained inside the reactor because it is too large to pass through the pores of a membrane patch in the reactor wall. This enables the use of a broad range of catalyst particle sizes including the relatively small particles of CLEAs. The reaction and biocatalyst separation are combined into a single operation. High catalyst loadings, longer catalyst life-times owing to reduced mechanical stress, and higher volumetric and catalyst productivities are some of the many advantages of an MSR. Its practical utility was demonstrated in the industrially important PGA-CLEA catalyzed hydrolysis of penicillin G to 6-APA [129].
The perfusion basket reactor (BR) is a refinement of the ‘tea bag’ concept, whereby the immobilized biocatalyst is contained in a filtration membrane-like module suspended in the reactor. It was used successfully, for example, in the degradation of endocrine-disrupting chemicals in aqueous effluents catalyzed by laccase-CLEAs [130]. The rotating flow cell reactor is a novel variation on the basket reactor, developed by SpinChem (Umeå, Sweden). It involves a catalyst-containing compartment attached to a propeller stirrer. In addition to protecting the biocatalyst from shear forces caused by the stirrer, rates of mass transfer are significantly increased thus enabling process intensification. Yet another alternative is to use a bubble column reactor (BCR). This was used, for example, in the production of highly viscous fatty acid esters of polyols which was not feasible in conventional STRs or fixed bed reactors (FBRs) [131]. The water formed in the reaction was removed by entrainment with pressurized air and the latter also serves to mix the reactants without causing any significant attrition of the catalyst.
Continuous processing with immobilized enzymes in fixed bed reactors (FBRs) is the norm in the oils and fats processing industry. The relatively small particle size of CLEAs is an issue as it can lead to large pressure drops over the column. The problem can be alleviated by mixing the CLEA with an inert, less compressible solid, such as controlled microporous glass or perlite. Littlechild and coworkers [132], for example, prepared CLEAs from a thermophilic l-aminoacylase from Thermococcus litorali and subsequently mixed them with controlled pore glass before packing them in a capillary reactor fitted with a silica frit to contain them in the reactor. Much attention has been focused in recent years on process intensification through the use of microchannel reactors because of the many advantages compared with traditional batch processing: rapid mass and heat transfer and large surface area to volume ratios.
Yet another alternative is to use magnetic CLEAs that are contained in a column reactor with a magnetic field. This is discussed in Section 6.
5.3. Multi- and Combi-CLEAs
The union of the CLEA technology with multipurpose and cascade biocatalysis has proven to be particularly attractive, producing many interesting progeny [133]. Multipurpose CLEAs can be prepared from crude enzyme extracts containing multiple enzymes. For example, Gupta and coworkers [134] prepared a multi-CLEA exhibiting lipase, phospholipase and α-amylase activity from porcine pancreatic acetone powder. Similarly, the same group prepared a multi-CLEA from a commercial enzyme preparation, Pectinex™ Ultra SP-L, exhibiting pectinase, xylanase and cellulase activity [135]. More recently, Rodrigues and coworkers [136] prepared multi-CLEAs, with and without BSA as a feeder, from pectinases and cellulases in crude enzyme preparations and demonstrated their efficacy in grape juice clarification. The multi-CLEA and multi-CLEA-BSA were recycled 4 and 6 times, respectively, with 100% retention of activity.
A multi-oxidoreductase CLEA, containing laccase, versatile peroxidase and glucose oxidase was used for the elimination of pharmaceuticals from aqueous waste streams [137]. Multi-CLEAs can also be produced by combining different enzymes from the same (sub)class. For example, a multi-CLEA containing a fungal laccase having an acidic pH optimum and a bacterial laccase having an alkaline pH optimum afforded a robust biocatalyst, active at both acidic and alkaline pH, for removal of phenolic compounds contaminating wastewater [138]. This concept of creating multi CLEAs with broader activities can also be applied to other enzymes. For example, it has been successfully applied to commercial preparations containing mixtures of ferulic acid esterases (FAEs) to afford a multi-FAE-CLEA with activity and stability superior to the individual enzymes in the preparations [139]. Similarly, one can easily envisage application of this concept to lipases, where individual enzymes often have very different specificities.
Recently, a novel lipase/protease multi-CLEA was reported [140]. Conducting reactions with a mixture of a protease and a lipase is not feasible with the free enzymes as the protease would break down the lipase but it is possible when the two enzymes are co-immobilized in a multi-CLEA.
We generally use the term combi-CLEA for deliberate co-immobilization of two or more enzymes in a single CLEA for the sole purpose of performing two or more sequential biotransformations. Multi-enzyme cascade processes [141,142,143,144,145] that telescope the individual reactions into one-pot cascade processes have several economic and environmental benefits: fewer unit operations, less solvent, and reactor, shorter cycle times, higher volumetric and space time yields and less waste. Furthermore, coupling of reactions can be used to drive equilibria towards product and, since individual biocatalytic steps generally proceed in water at ambient temperature and pressure, they can be readily integrated into one-pot processes. However, different enzymes are not always compatible and nature solves this problem by compartmentalizing enzymes in different parts of the cell. Hence, compartmentalization via immobilization can be the solution in enzymatic cascades.
In addition, co-immobilization of enzymes in combi-CLEAs can significantly increase the rates of sequential biocatalytic cascades by simulating the close proximity extant in microbial cells, what has become known as sequential channeling of substrates [146]. In our initial studies of combi-CLEAs we prepared combi-CLEAs of glucose oxidase or galactose oxidase with catalase [117]. Oxidases catalyze the aerobic oxidations of various substrates, generally with concomitant production of an equivalent of hydrogen peroxide. The latter can cause oxidative degradation of the enzyme and, in vivo, oxidases generally occur together with catalase which catalyzes the spontaneous decomposition of hydrogen peroxide to oxygen and water. We found that combi-CLEAs, containing co-immobilized oxidase with catalase, could not only be recycled without significant loss of activity but also exhibited significantly better activities and stabilities compared to a mixture of the two separate CLEAs.
A combi-CLEA consisting of glucose oxidase and horse radish peroxidase (HRP) can be used for the in situ production of hydrogen peroxide by aerobic oxidation of glucose followed by HRP catalyzed oxidations with the hydrogen peroxide [147]. Similarly, a combi-CLEA consisting of glucose oxidase and versatile peroxide (VP), with added glucose, catalyzed the elimination of endocrine disruptors from industrial wastewater streams [148].
The combi-CLEA concept has also been used to advantage in oxidation reactions requiring cofactor regeneration. A pertinent example is the ketoreductase (KRED)/glucose dehydrogenase (GDH) combi-CLEA that was used in the enantioselective reduction of a prochiral ketone with cofactor regeneration with glucose/GDH [149,150]. The combi-CLEA exhibited increased thermal and pH stability and high substrate tolerance compared with the free enzymes, coupled with high volumetric productivity, good operational stability and reusability. The authors predicted wide application of this combi-CLEA strategy in the production of various chiral alcohols on the basis of its high efficiency coupled with low cost.
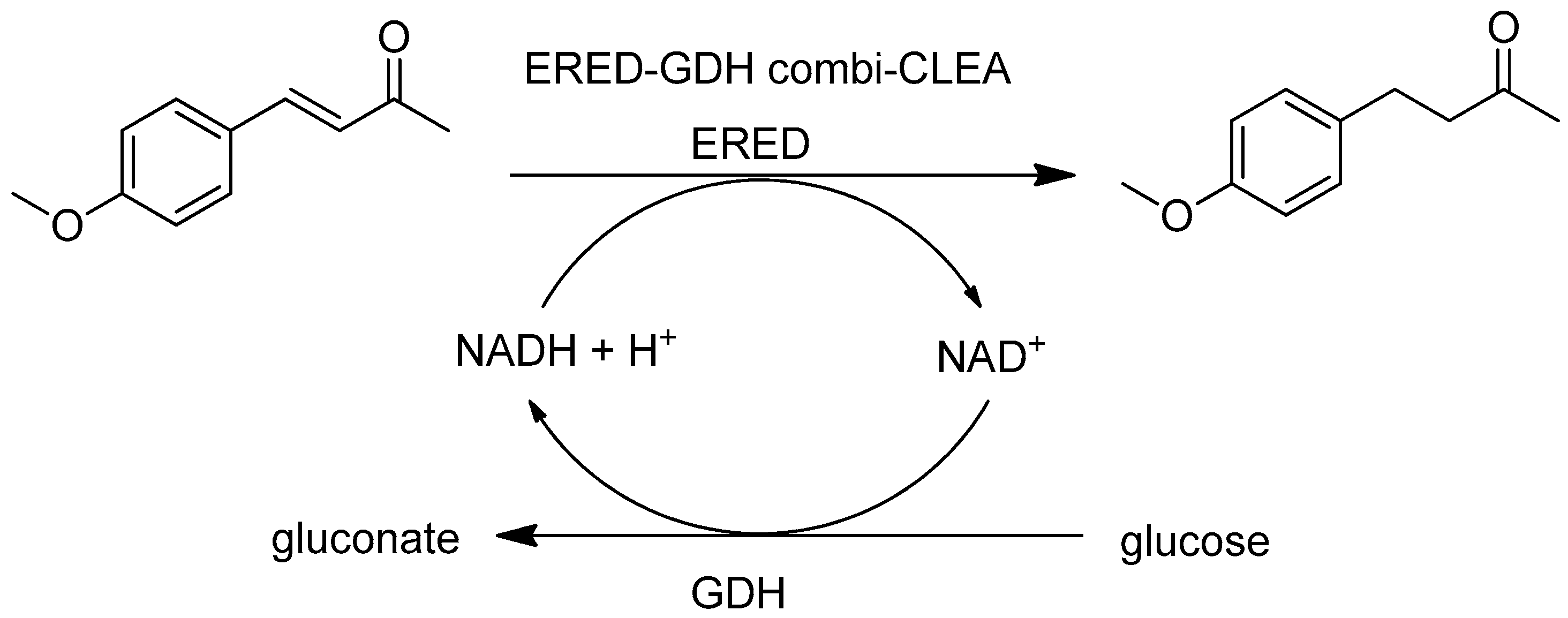
Another example involving GDH for cofactor recycle is the enoate reductase (ERED)/GDH combi-CLEA. The combi-CLEA exhibited improved thermal stability and 110% retention of the initial activity after 14 recycles (Figure 7) [151].
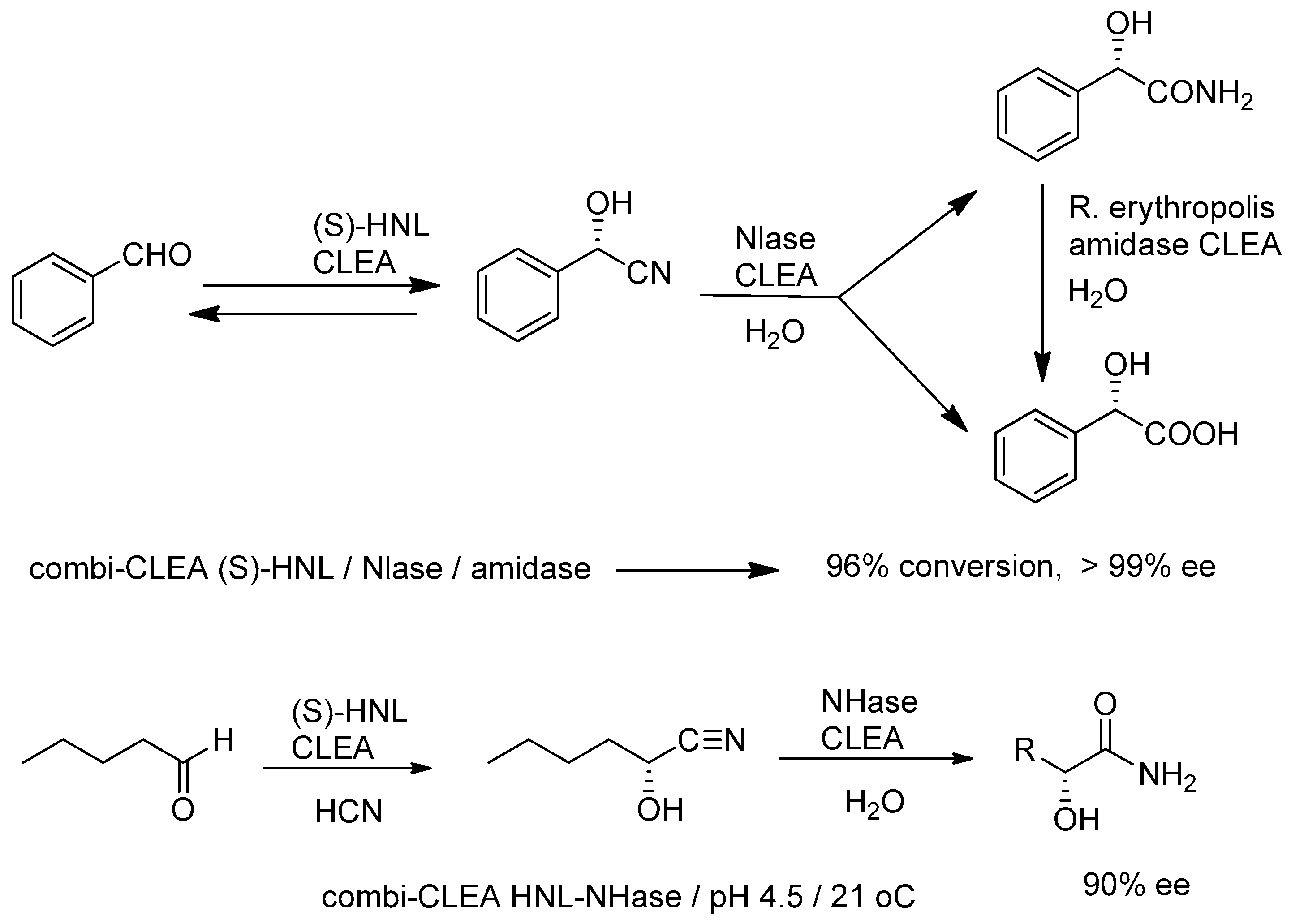
A trienzymatic combi-CLEA, consisting of a (S)-hydroxynitrile lyase from Manihot esculenta, a non-selective nitrilase from Pseudomonas fluotrescens EBC 191 and an amidase from Rhodococcus erythropolis, catalyzed the enantioselective conversion of benzaldehyde to (S)-mandelic acid in 90% yield and 99% ee (Figure 8) [152,153]. It was originally developed [33] as a hydroxynitrile lyase/nitrilase bienzymatic combi-CLEA but large amounts of the corresponding amide were formed as a byproduct of the nitrilase-catalyzed hydrolysis and an amidase was needed to convert the amide side product to the acid. Rates were higher than with mixtures of the respective CLEAs. Similarly, a combi CLEA comprising the manihot esculenta HNL and the alkaliphilic NHase from Nitriliruptor akaliphilus (EC 4.2.1.84) catalyzed the one-pot conversion of aldehydes to (S)-α-hydroxycarboxylic acid amides (Figure 8) [154].
Proline-specific X-prolyl-dipeptidyl aminopeptidase (PepX) and aminopeptidase N (PepN) are used in cheese-making, baking and meat tenderization. Enzymatic hydrolysis of casein, for example, can cause an undesirable bitter taste but this can be significantly reduced using a combination of PepX and PepN. Fischer and coworkers [155] showed that by using a PepX/PepN combi-CLEA the degree of hydrolysis of casein was increased by 52%.
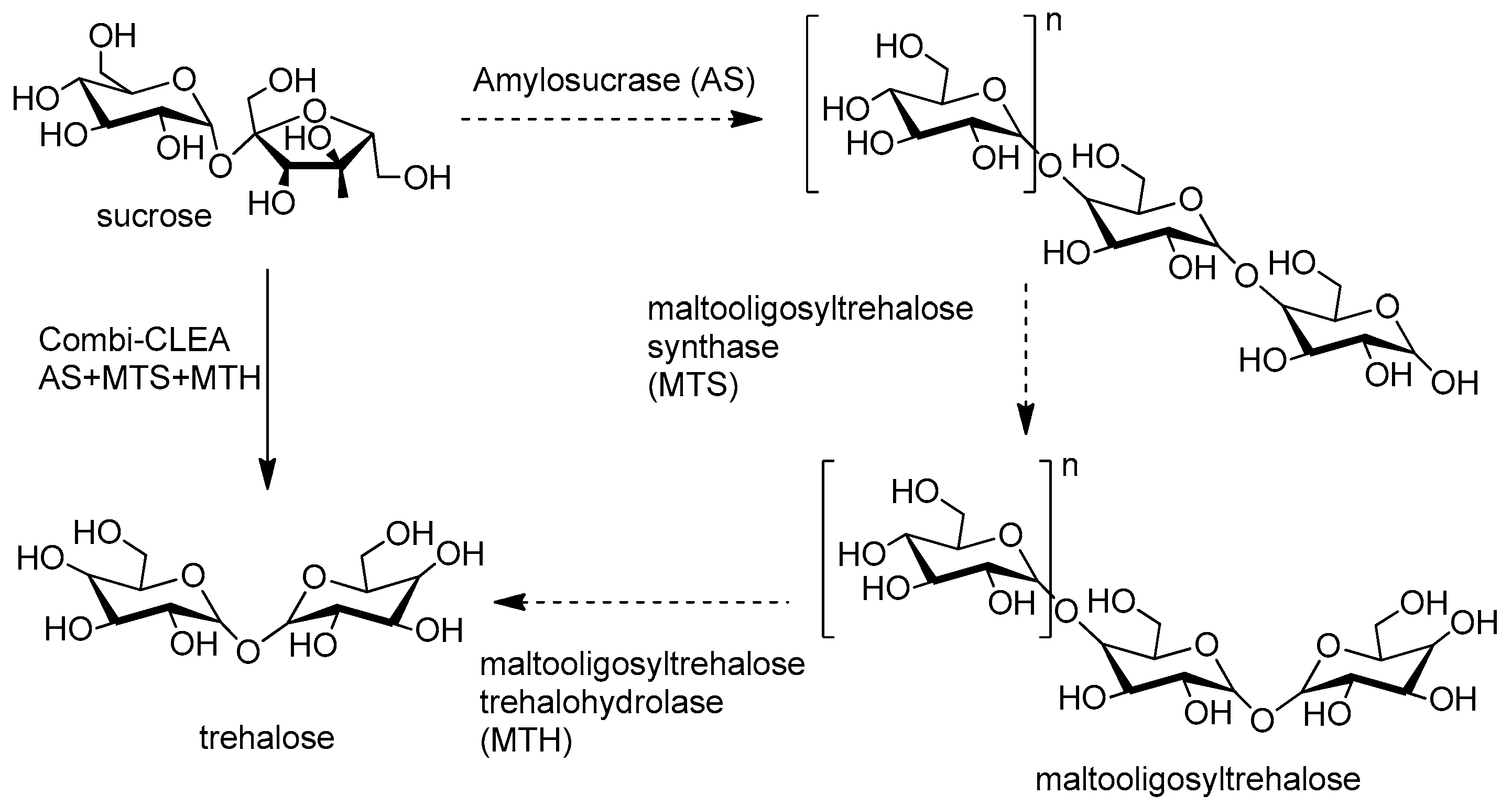
An area where combi-CLEAs have made significant inroads is in conversion of carbohydrates in general, and polysacharides in particular. A pertinent example is the use of a trienzymatic combi-CLEA of amylosucrase (AS; EC 2.4.1.4), maltooligosyltrehalose synthase (MTS; EC 5.4.99.15) and maltooligosyltrehalose terhalohydrolase (MTH; EC 3.2.1.141) in a one-pot conversion of sucrose to trehalose (Figure 9) [156]. The CLEA was recycled five times.
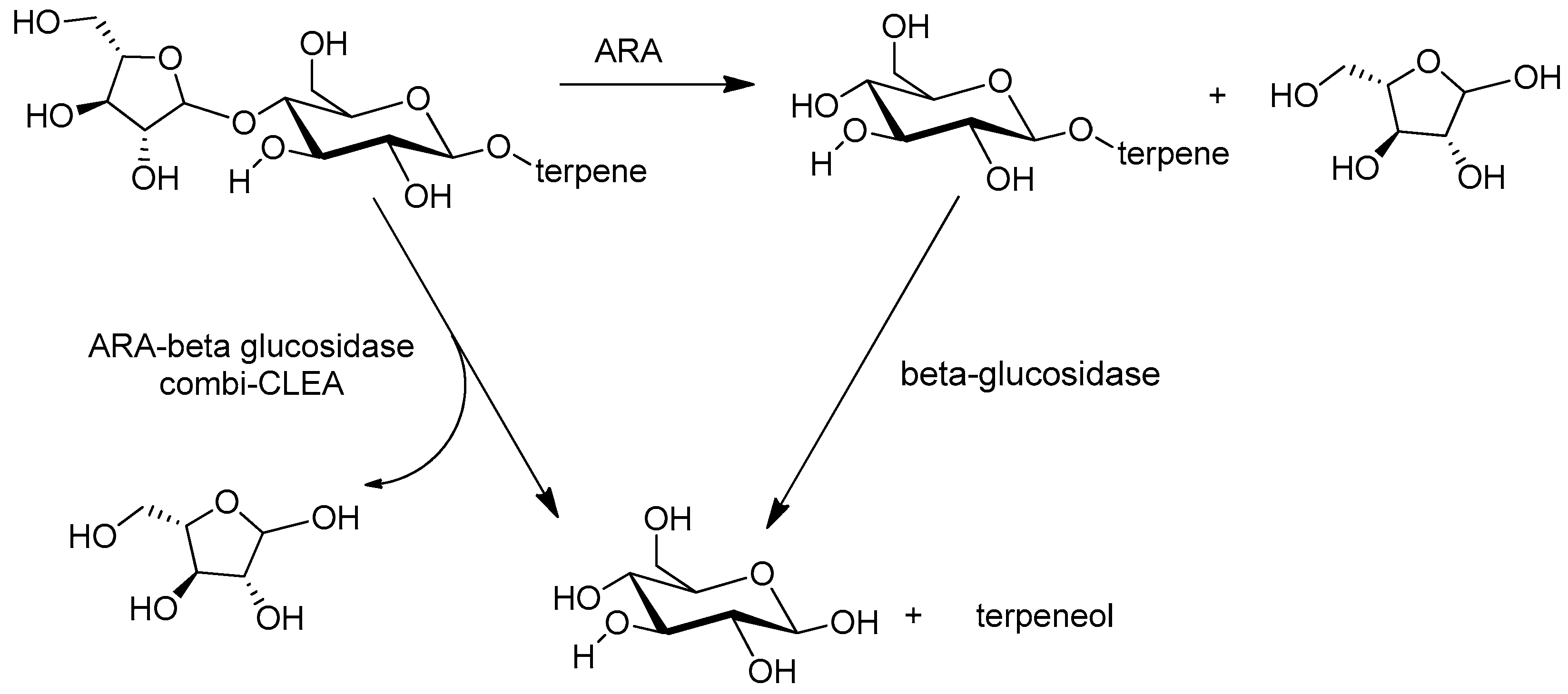
A combi-CLEA of α-L-arabinose (ARA) and β-glucosidase (βG) prepared from a commercial enzyme preparation, Rapidase AR 2000, was used for aroma enhancement in wine [157]. ARA catalyzes the formation of terpenyl-β-glucosides and βG catalyzes their subsequent hydrolysis to release terpenyl alcohols that enhance the wine aroma (Figure 10). All grape varieties comprise such precursors but Muscat grapes in particular have the highest concentrations. Wine treated with the combi-CLEA exhibited the highest concentration of total terpenes (18% higher) and of the individual terpenes—linalool (20% higher), nerol (20% higher) and geraniol (100% higher)—which determine the typical Muscat aroma. The stability of the enzymes in the combi-CLEA was an order of magnitude higher than that of the free enzymes, thus facilitating their reuse and reducing the impact of enzyme costs on the total costs of the winemaking.
Hydrolysis of starch, cellulose and other polysaccharides are important reactions in many industries including food and beverages processing. Furthermore, their industrial importance is rapidly increasing with the impending transition to a biobased economy, in which oil refining will, be largely superseded by polysaccharide refining for the production of basic chemicals and liquid fuels. In this scenario the enzymatic conversion of starch and waste lignocellulose, as first and second generation renewable biomass, respectively, to fermentable sugars will take centre stage [14].
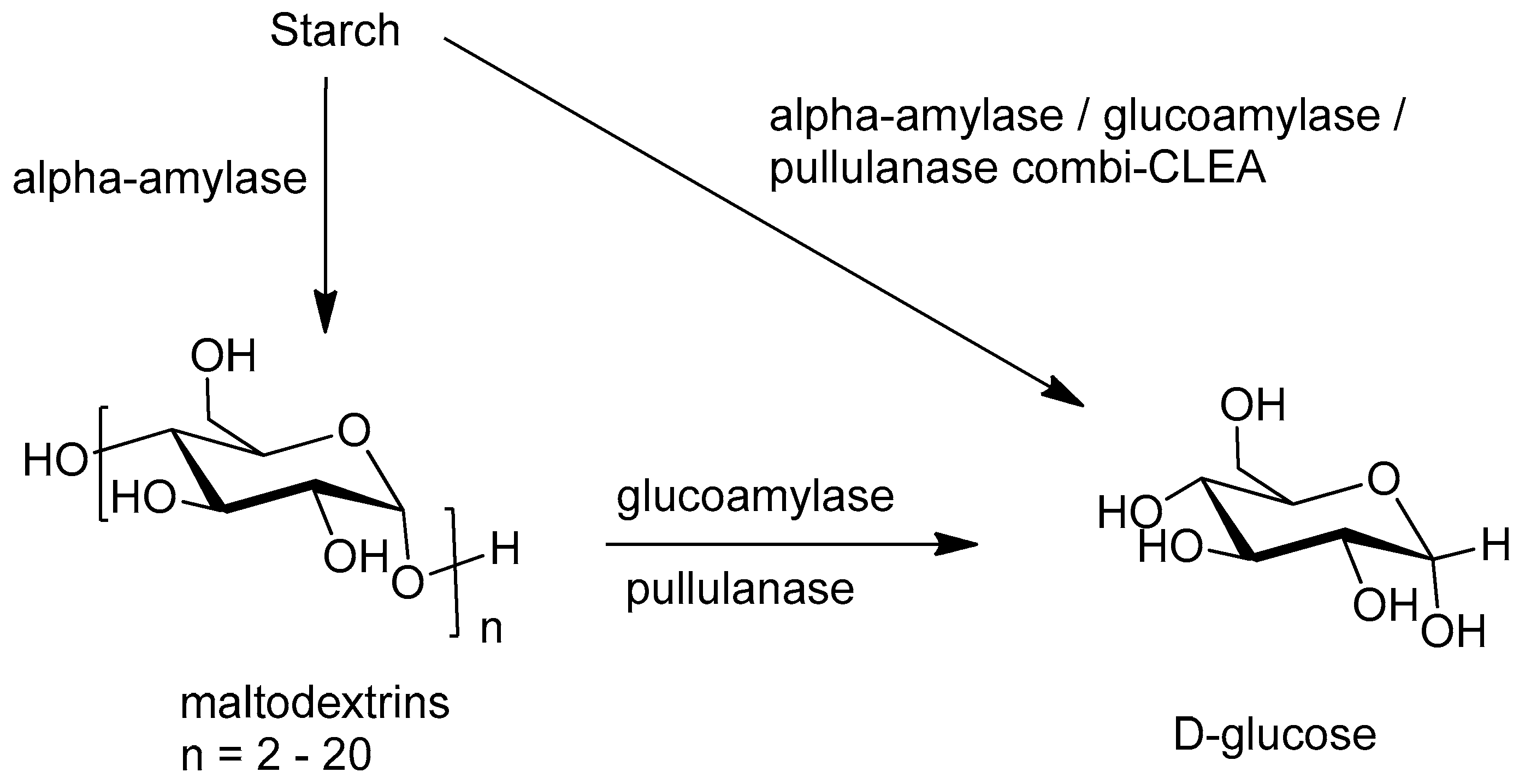
Enzymatic hydrolysis of starch involves two consecutive steps: liquefaction and saccharification. Liquefaction is catalyzed by α-amylase (E.C. 3.2.1.1) at 90 °C and pH 7 and hydrolyzes α-(1-4) glycosidic bonds. This is followed by glucoamylase (E.C. 3.2.1.3) catalyzed saccharification at 60 °C and pH 5, involving hydrolysis of both α-(1-4) and α-(1-6) glycosidic bonds. CLEAs have been successfully prepared from both α-amylase [158,159,160] and glucoamylase [161]. In some cases [161], macromolecular cross-linkers were used to produce more porous CLEAs with better activities.
The glucoamylase catalyzed hydrolysis of α-(1,6) branches is relatively slow and pullulanase (E.C. 3.2.1.41), is sometimes added to facilitate this process. Co-immobilization of the two enzymes in a combi-CLEA resulted in a shift in optimal pH (from 5 to 7) and temperature (from 60 to 70 °C) [162]. The combi-CLEA gave 100% conversion after 3 h compared with 30% with the free enzymes and 80% with a mixture of the two individual CLEAs. The combi-CLEA exhibited 90% and 85% retention of the glucoamylase and pullulanase activity, respectively, after 8 recycles.
More recently, the same group prepared a tri-enzyme combi-CLEA containing α-amylase, glucoamylase and pullulanase from commercially available enzyme preparations [163]. The thermal stability of all three enzymes was increased compared to the free enzymes. In a one-pot starch hydrolysis (Figure 11) 100%, 60% and 40% conversions were observed with the combi-CLEA, a mixture of the separate CLEAs and a mixture of the free enzymes, respectively, and the catalytic performance was maintained for up to 5 cycles.
The hydrolysis of lignocellulose is currently the focus of attention in connection with second generation biofuels production from waste lignocellulose streams. Lignocellulose has a complex structure consisting of lignin (15–25%), cellulose (30–45%) and hemi-cellulose (25–40%) that requires a complex cocktail of cellulolytic and hemi-cellulolytic enzymes to catalyze hydrolysis to its constituent sugars [164]. Cellulose hydrolysis involves at least five enzymes: exo-1,4-β-glucanase (EC 3.2.1.91), endo-,1,4-β-glucanase (EC 3.2.1.4), cellobiohydrolase (EC 3.2.1.176) and β-glucosidase (EC 3.2.1.21) and the more recently discovered, copper-dependent lytic polysaccharide monooxygenases (LPMOs).
Hemi-cellulose has a more complicated structure than cellulose and requires a diverse suite of enzymes to affect its hydrolysis to its constituent sugars, mainly xylose and mannose. They can be divided into two groups: core enzymes catalyzing cleavage of the polysaccharide backbone and ancilliary enzymes involved in the removal of functional groups. Examples of the former are endo-β-1,4-xylanase (EC 3.2.1.8), xylan-1,4-β-xylosidase (EC 3.2.1.37), endo-1,4-β-mannanase (EC 3.2.1.78) and β-1,4-mannosidase (EC 3.2.1.25). Ancilliary enzymes include β-glucuronidase (EC 3.2.1.139), acetylxylan esterase (EC 3.2.1.55), ferulic acid esterase (EC 3.1.1.73) and p-coumaric acid esterase (EC 3.1.1). Cellulolytic fungi and bacteria produce so-called cellulosomes, containing multi-enzyme complexes that, because of the close proximity of the enzymes, are more effective than simple mixtures of the free enzymes. One can envisage that this proximity effect can be mimicked in combi-CLEAs without having many of the disadvantages of cellulosomes [165].
For example, a xylanase-mannanase combi-CLEA was successfully used in the conversion of lime-treated sugar cane bagasse and milled corn stover [166] leading the authors to conclude that the efficiency of combi-CLEAs makes them ideal candidates for achieving cost-effective application of lignocellulytic enzymes. Similarly, co-immobilization of xylanase, β-1,3-glucanase and cellulase gave a combi-CLEA which was more thermally stable than the free enzymes and retained more than 97% of its activity on storing at 4 °C for 11 weeks, compared to 65% for the free enzymes [167]. The combi-CLEA was successfully used in the hydrolysis of ammonia-cooked sugar cane bagasse and could be recycled 6 times.
6. Magnetic CLEAs: The New Frontier
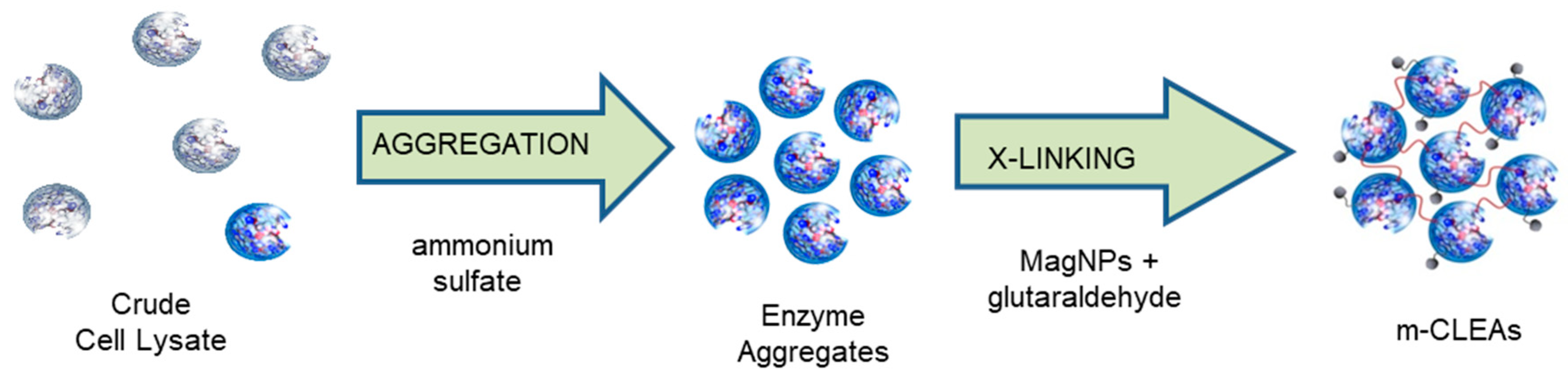
Notwithstanding the many advantages of CLEAs, the relatively small particle size and lack of mechanical robustness remain issues in many cases. The particle size can be increased by, for example, modifying the cross-linking protocol but inevitably a compromise has to be reached between the ease of processing of large particles and the lower mass transfer limitations and, hence, higher activities of small particles. Moreover, the processing of polysaccharides, in particular lignocellulosic feedstocks, is fraught with another difficulty: recovery and reuse of the solid catalyst often requires its separation from other solid materials present in the crude mixtures. In order to rectify these shortcomings, the CLEA technology has been raised to a new level of sophistication and industrial relevance with the invention of “smart” magnetically recoverable CLEAs (m-CLEAs) by conducting the cross-linking in the presence of magnetic (nano)particles (Figure 12).
m-CLEAs have a relatively small particle size, and high activity, combined with ease of separation on an industrial scale using standard commercial equipment for magnetic separation with permanent magnets, affording novel combinations of bioconversions and downstream processing. The m-CLEAs are particularly useful for processes in which they have to be separated from other solids, e.g., in slurry processes.
6.1. Preparation of m-CLEAs
Our original development of m-CLEAs [168] involved conducting the cross-linking step in the presence of amino functionalized particles of magnetite (Fe3O4) obtained by reaction of (3-aminopropyl)triethoxysilane (APTES) with surface hydroxyl groups. The magnetic particles are incorporated in the CLEA superstructure through reaction of the cross-linker with both the enzyme aggregates and the NH2 groups on the surface of the magnetic particles. The use of ferromagnetic catalysts, such as Raney nickel, on an industrial scale is well established [169]. In combination with commercially available magnetic separation equipment, e.g., from the Eclipse Magnets Automag series (see www.eclipsemagnetics.com), very high recoveries can be obtained at flow rates that are acceptable for industrial scale production. Subsequently, we and others [170] found that robust, stable mCLEAs could be produced in a more cost-effective way using non-functionalized magnetite partcles.
However, we found that leaching of iron occurs at acidic pH and is, moreover, accelerated in the presence of free carboxylic acids. These are precisely the conditions encountered in the hydrolysis of starch and, especially lignocellulose. We attribute the fact that iron leaching has not been observed by other authors to the fact that it becomes apparent only after long residence times and several recovery cycles. This obviously poses serious problems for their use in industrial processes. Another limitation of the conventional magnetite-based mCLEAs is the relatively low saturation magnetizability of magnetite which makes magnetic recovery in large scale processes challenging. In order to overcome these shortcomings of the conventional magnetite-based m-CLEAs, we developed mCLEAs based on non-functionalized magnetizable particles of zerovalent iron, so-called carbonyl iron particles (CIP) produced by thermal decomposition of iron pentacrabonyl [171]. This well-established method produces spherical iron particles with particle sizes of a few micrometers. These particles were then covered by a protected coating of silica with a thickness of a few nanometers silica in order to maintain their physical properties and protect them from corrosion under reaction conditions.
The m-CLEAs typically have particle sizes in the range 1–15 μm. Iron leaching was not observed with these m-CLEAs, even under relatively harsh conditions. The saturation magnetizations of these CIP-m-CLEAs were surprisingly high—up to 200 emu⸱g−1—compared to the range of 10–40 emu⸱g−1 observed with magnetite-based m-CLEAs. Using this new cost-effective methodology to prepare m-CLEAs with very high magnetic susceptibility adds little cost to regular CLEAs and not many recycles are required to achieve cost reductions. The technique was demonstrated with glucoamylase and glutaminase.
6.2. Applications of m-CLEAs and m-Combi-CLEAs
m-CLEAs have been prepared from a variety of enzymes. However, interest in industrial application of m-CLEAs and m-combi-CLEAs has primarily been motivated by the possibility of using them in the complex conversions of polysaccharides such as starch and, particularly, lignocellulose in connection with first- and second-generation biofuels and food and beverages.
For example, Talekar and coworkers [172] prepared m-CLEAs of α-amylase, for use in starch hydrolysis. The activity recovery was 100% with improved thermal and storage stability compared to the free enzyme and 100% activity retention after 6 recycles. A m-CLEA of glucoamylase was prepared using pectin dialdehyde as the cross linker with 95% activity recovery compared to 85% using glutaraldehyde as the cross linker [173]. The authors attributed the higher activity recovery to better mass transfer with macromolecular substrates in the more open porous structure. m-CLEAs of glucoamylase from Aspergillus niger with enhanced thermal and storage stability and reusability and 93% activity recovery were prepared by Gupta and coworkers [174]. More recently, Tardioli and coworkers [175] prepared m-glucoamylase CLEAs by co-aggregation with amino functionalized magnetic nanoparticles and using PEI as an aid in the cross-linking with glutaraldehyde. The m-CLEA exhibited similar behavior to the free enzyme in starch saccharification under typical industrial conditions (55 °C and pH 4.5). We found (see above) that m-glucoamylase CLEAs based on magnetite were not stable towards leaching at acidic pH.
Probably the most exciting and challenging development is the application of the m-CLEA technology to the complex chemistry of lignocellulose hydrolysis in connection with the production of second-generation biofuels and commodity chemicals from waste biomass [176,177]. Various groups have reported [178,179,180] the preparation of m-CLEAs with improved stability from a cellulase cocktail. However, its activity was measured only in the hydrolysis of the water soluble carboxymethyl cellulose. Bhattachariya and Pletschke [166] prepared m-CLEAs of a bacterial xylanase and observed that incorporation of Ca2+ ions in the CLEA led to increased thermal stability. The Ca-m-CLEA exhibited 35% more activity than the free enzyme and a 9 fold higher sugar release from ammonia pretreated sugar cane bagasse. Illias and coworkers [181] prepared m-CLEAs of a recombinant xylanase from Trichoderma reesei using maghemite (γ-Fe2O3) rather than the more usual magnetite (Fe3O4) nanoparticles. Second-generation biofuels are an obvious application for m-CLEAs of lignocellulolytic enzymes but applications in food and beverages processing are also envisaged.
The enzyme costs per kg product are crucial for the economic viability of these processes, particularly in the case of lignocellulosic feedstocks. In both 1G and 2G biofuels and commodity chemicals production, in order to be more cost-effective the enzyme catalyzed hydrolysis of the feedstock—starch or lignocellulose—is often combined with the subsequent fermentation step in so-called simultaneous saccharification and fermentation (SSF) processes [182]. Multiple recycling of the enzyme(s), using an immobilized, solid enzyme which can be easily recovered and re-used, represents a clear opportunity to reduce the enzyme costs and drive competitiveness and sustainability. However, the need to separate the solid immobilized enzyme from the vast amount of other solids such as fibers and/or yeasts (in SSF processes) suspended in the reaction mixture precludes the use of standard immobilized enzymes.
Up till now, attention has been focused on starch and lignocellulose hydrolysis but the m-CLEA technology can also be applied in the hydrolysis of other polysaccharides in processes for their conversion to biofuels and commodity chemicals. For example, pectins (galacturonans) and inulins (fructans) can be hydrolyzed using pectinase and inulinase, respectively. For example, a trienzyme magnetic combi-CLEA consisting of an α-amylase, pectinase and cellulase was used for clarification of fruit juices [183].
The growing trend towards the utilization of food supply chain waste (FSCW) [184], most of which goes to landfill and has a negative value, is also stimulating the application of biocatalysis, e.g., proteases, carbohydrases, and lipases, in the processing of waste proteins, carbohydrates and oils and fats, respectively.
6.3. Lipase-m-CLEAs
Lipase-m-CLEAs have significant commercial potential in the processing of non-edible and waste oils and fats to 2G biodiesel. Conventional chemical processes generally involve catalysis by bases such as sodium methoxide and are energy-intensive operations with high waste treatment costs. Moreover, the feedstock must have a low free fatty acid content since any free fatty acids will neutralize the basic catalyst, resulting in catalyst losses and the formation of soap emulsions. In contrast, enzymatic transesterification is characterized by mild reaction conditions, low energy demand, low waste treatment costs and more flexibility with regard to free fatty acids in the feedstock, as they are also esterified to biodiesel. However, current lipase-based methods suffer from high enzyme cost contributions and inactivation of the lipase by the methanol and/or impurities in the feedstock. The use of recyclable m-CLEAs of the lipase catalysts could provide the necessary reduction in enzyme costs to enable commercial viability.
m-CLEAs of CaLB were used in the production of biodiesel by transesterification of sunflower oil [185], non-edible vegetable oils, such as jatropha and cameline, and waste frying oils [186] and microalgae oil [187]. Similarly, surfactant activated m-CLEAs of Thermomyces lanuginosus lipase (TLL) [188] and Rhizomucor miehei lipase (RML) [189] were used in the production of biodiesel from jatropha oil. The m-CLEAs could be readily recovered and recycled for several cycles.
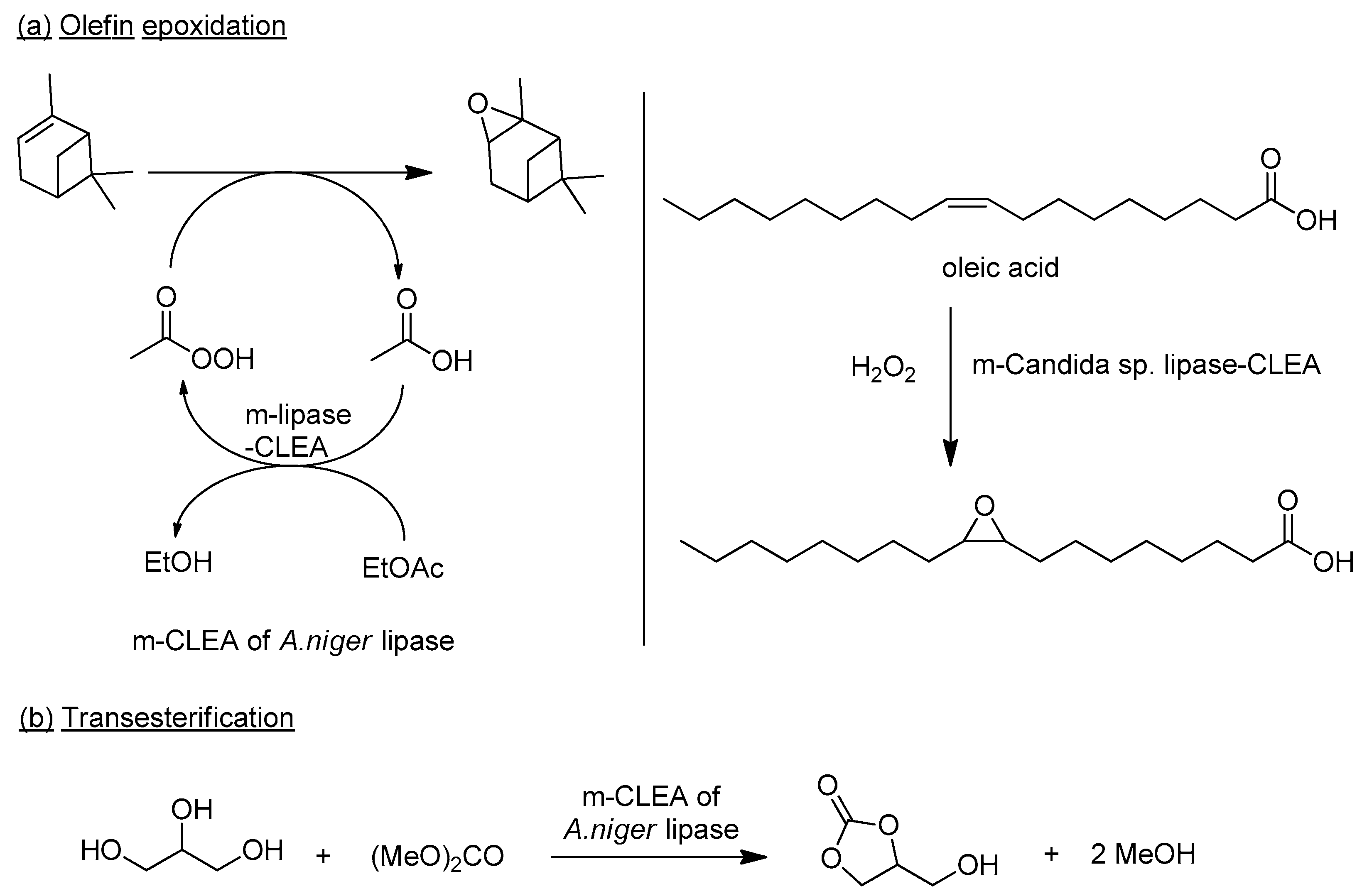
A m-CLEA of a Candida sp. lipase was used to catalyze the epoxidation of oleic acid with hydrogen peroxide, via in situ lipase catalyzed formation of peroleic acid (Figure 13) [190]. The m-CLEA exhibited higher thermal and storage stability and better tolerance of hydrogen peroxide compared to the free enzyme and the standard CLEA. Similarly, a m-CLEA of an Aspergillus niger lipase was used [191] to catalyze the epoxidation of α-pinene with hydrogen peroxide in ethyl acetate as solvent and acetate-source (Figure 13) and the synthesis of glycerol carbonate by transesterification with dimethyl carbonate [192].
m-CLEAs of Rhizopus arrhizus lipase were used for the production of butyl oleate, a widely used diesel additive, in a novel microreactor consisting of a column containing the m-CLEA, and sieve plates through which solvents and products can pass, and surrounded by a rotating magnetic field [193]. Reactants were fed from the bottom of the column as in a fluidized bed.
6.4. Ligninolytic Enzymes and Waste Water Treatment
White rot fungi produce a range of extracellular enzymes—laccases, lignin peroxidases and manganese peroxidases—which are involved in the recycling of organic material in vivo and have broad applications in bioremediation. Immobilization of these enzymes, for example as CLEAs or combi-CLEAs is necessary in order to be cost-effective in waste water treatment. Although it is difficult to achieve the very low enzyme costs required for commercial viability in waste water treatment in general there may very well be niche opportunities in the case of emerging micropollutants that are difficult to remove by other means For example, laccase-m-CLEAs [197,198] and a combi-m-CLEA of horseradish peroxidase and glucose oxidase [199] have been used for the decolorization of recalcitritant dyes in waste water and various reactor concepts have been considered. A laccase-m-CLEA was also used for removal of pharmaceuticals from waste water [200] and an acrylamidase m-CLEA was used to catalyze the biodegradation of acrylamide in industrial waste water [201].
6.5. Synthesis of Semi-Synthetic Penicillin and Cephalosporin Antibiotics
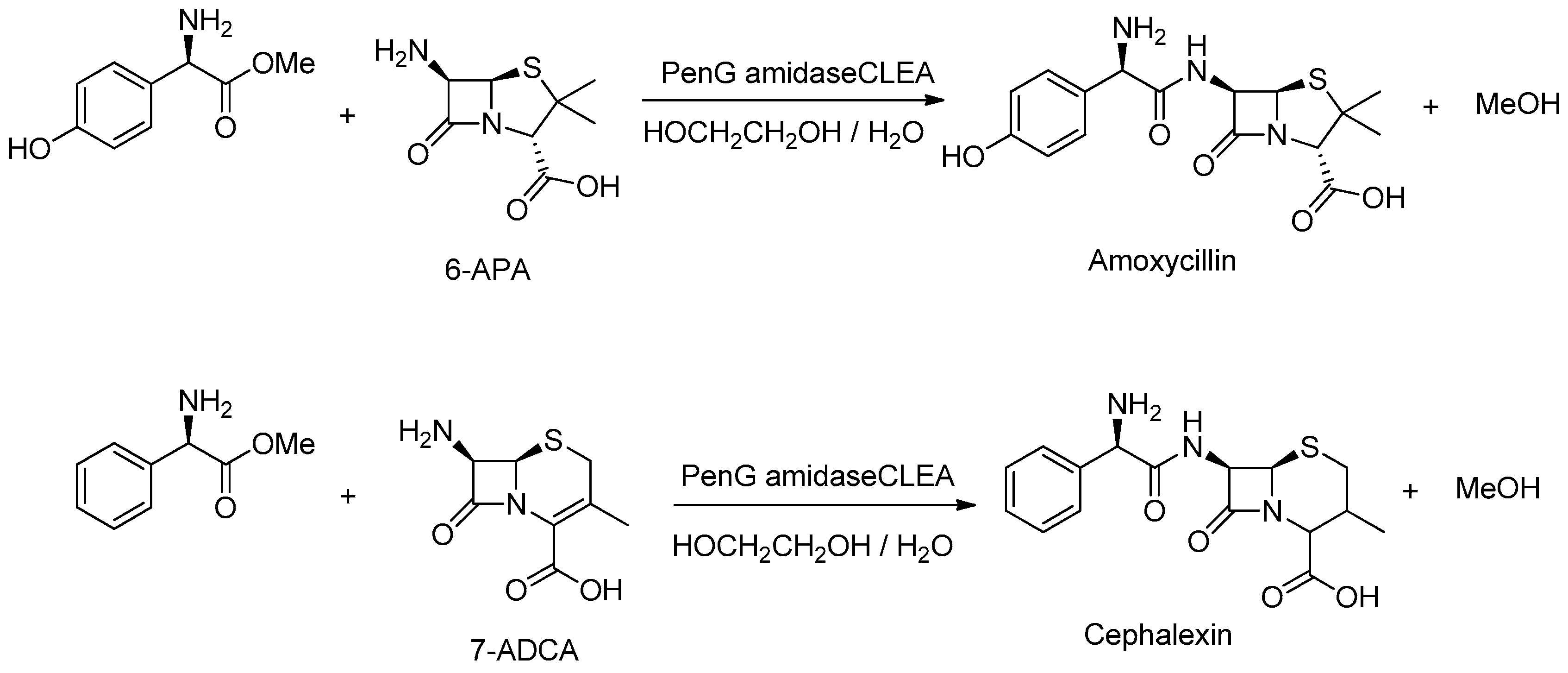
Semi-synthetic penicillins and cephalosporins are produced on an industrial scale by PGA catalyzed coupling of the side-chain with the 6-aminopenicillanic acid (6-APA) or 7-aminodesacetoxycephalosporanic acid (7-ADCA) nucleus (Figure 14). The coupling is conducted in water and the product crystallizes out of the reaction mixture. This is essential as otherwise the enzyme would catalyze the thermodynamically favored hydrolysis of the product. However, the enzyme needs to be immobilized to enable its reuse but then it has to be separated from the solid product. This can be achieved using a sieve bottom reactor in which the crystals of product pass through the sieve while the larger particles of immobilized enzyme are retained in the reactor [202]. Alternatively, PGA m-CLEA was shown to catalyze the hydrolysis of penicillin G [203] and it could be used to produce semi-synthetic penicillins [204] and cephalosporins [205].
6.6. Other Potential Applications
Magnetic CLEAs of other enzymes have been described. For example, m-combi-CLEAs have been prepared from ketoreductases (KREDs) and glucose dehydrogenase for cofactor regeneration with glucose and used in the synthesis of chiral alcohols [149,206,207]. Cui and coworkers [170] prepared m-CLEAs of phenyl alanine ammonia lyase (PAL EC 4.3.1.24) and showed that storage and operational stabilities were superior to those of the simple CLEA. Bovine carbonic anhydrase, of interest in connection with processes for CO2 capture, was also successfully immobilized as a m-CLEA [208]. Finally, glucose isomerase, the largest volume immobilized enzyme (see earlier) has been immobilized as a m-CLEA [209]. The temperature and pH optima shifted in the m-CLEA and a significant increase in activity with 80% retention after 6 recycles was observed which led the authors to conclude that the “m-CLEAs of glucose isomerase clearly have practical utility in HFCS production”.
7. Conclusions and Outlook
In the preceding decade the efficacy and cost-effectiveness of the cross-linked enzyme aggregates (CLEA) methodology in the immobilization of a wide variety of enzymes, thereby enabling many applications of biocatalysis, has been amply demonstrated. With regard to the eight requirements that Turner and coworkers [40] recently outlined for the commercial viability of an immobilized enzyme in the pharmaceutical industry, CLEAs exhibit (i) high enzyme loading (10 wt%); (ii) high activity recovery (50%); (iii) no leaching under reaction conditions; (iv) good tolerance to organic solvents; (v) are eminently recyclable (20 cycles); and (vi) result in a lower cost of good compared with alternatives. Mass transport of substrates is sometimes an issue, especially with macromolecular substrates, but techniques have been developed to alleviate this problem. Mechanical stability in batch and flow reactors is also an issue with CLEAs but methods have been developed to address these problems. In contrast, carrier-bound enzymes have low enzyme loadings, often low activity recoveries and, generally higher cost contributions. Depending on the method of binding to the carrier, leaching can also be an issue but they tend to have better mechanical stability than CLEAs. Both mass transport and mechanical stability issues can be circumvented by employing smart m-CLEAs with small particle size in conjunction with recovery using permanent magnets in commercially available separation equipment.
In short, the CLEA technology has proven its practical utility and industrial viability. It has been particularly successful in the immobilization of a variety of enzymes for use in the conversion of carbohydrates, ranging from mono- to polysaccharides, in aqueous media, thus enabling multiple recycling of the enzymes. Applications in industrial biocatalysis are currently being further stimulated by the ongoing transition to a more sustainable bio-based, circular economy in which there is a growing trend towards the use of carbohydrates as a substitute for hydrocarbon feedstocks. We expect this trend to continue in the future and that combi-CLEAs and “smart” mCLEAs will be increasingly utilized in carbohydrate conversion processes in both traditional food and beverages processing and in the production of biofuels, commodity chemicals and bio-based polyesters and polyamides from waste renewable biomass, including food supply chain waste.
Funding
This research received no external funding.
Acknowledgments
The important and lasting contributions of my co-workers at Delft University of Technology and CLEA Technologies B.V., who are mentioned in the references, are gratefully acknowledged.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Kazlaukas, R.J. Biocatalysis for Green Chemistry and Chemical Process Development; Wiley: Hoboken, NJ, USA, 2011. [Google Scholar]
- Sheldon, R.A.; Brady, D. The limits to biocatalysis: Pushing the envelope. Chem. Commun. 2018, 54, 6088–6104. [Google Scholar] [CrossRef]
- Wohlgemuth, R. Biocatalysis-key to sustainable chemistry. Curr. Opin. Biotechnol. 2010, 21, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Hu, J.-H. Biocatalysis in developments of green pharmaceutical processes. Curr. Opin. Chem. Biol. 2009, 13, 43–50. [Google Scholar] [CrossRef]
- Patel, R.N. Biocatalysis for the synthesis of pharmaceuticals. Bioorg. Med. Chem. 2018, 26, 1252–1274. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, J.-B.; Reetz, M.T. Biocatalysts for the pharmaceutical industry created by structure-guided directed evolution of stereoselective enzymes. Bioorg. Med. Chem. 2018, 26, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Zhang, H.; Ang, E.L.; Zhao, H. Biocatalysis for the synthesis of pharmaceuticals and pharmaceutical intermediates. Bioorg. Med. Chem. 2018, 26, 1275–1284. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, K.; Lütz, S. Recent developments and challenges of biocatalytic processes in the pharmaceutical industry. Curr. Opin. Green Sustain. Chem. 2018, 11, 58–64. [Google Scholar] [CrossRef]
- Huisman, G.W.; Collier, S.J. On the development of new biocatalytic processes for practical pharmaceutical synthesis. Curr. Opin. Chem. Biol. 2013, 17, 284–292. [Google Scholar] [CrossRef]
- Lalonde, J. Highly engineered biocatalysts for efficient small molecule pharmaceutical synthesis. Curr. Opin. Biotechnol. 2016, 42, 152–158. [Google Scholar] [CrossRef]
- Truppo, M.D. Biocatalysis in the Pharmaceutical Industry: The Need for Speed. ACS Med. Chem. Lett. 2017, 8, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Heux, S.; Meynial-Salles, I.; O’Donohue, M.J.; Dumon, C. White biotechnology: State of the art strategies for the development of biocatalysts for biorefining. Biotechnol. Adv. 2015, 33, 1653–1670. [Google Scholar] [CrossRef]
- Sheldon, R.A. Biocatalysis and Biomass Conversion in Alternative Reaction Media. Chem. Eur. J. 2016, 22, 1–17. [Google Scholar] [CrossRef]
- Sheldon, R.A. Chemicals from Renewable Biomass: A Renaissance in Carbohydrate Chemistry. Curr. Opin. Green Sustain. Chem. 2018, 14, 89–95. [Google Scholar] [CrossRef]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular biological access to the chemistry of unknown soil microbes: A new frontier for natural products. Chem. Biol. 1998, 5, R245–R249. [Google Scholar] [CrossRef]
- Reetz, M.T. Biocatalysis in Organic Chemistry and Biotechnology: Past, Present, and Future. J. Am. Chem. Soc. 2013, 135, 12480–12496. [Google Scholar] [CrossRef]
- Sun, Z.; Witmark, Y.; Bäckvall, J.-E.; Reetz, M.T. New Concepts for Increasing the Efficiency in Directed Evolution of Stereoselective Enzymes. Chem. Eur. J. 2016, 22, 5046–5054. [Google Scholar] [CrossRef]
- Madhavan, A.; Sindhu, R.; Binod, P.; Sukumaran, R.K.; Pandey, A. Strategies for design of improved biocatalysts for industrial applications. Bioresour. Technol. 2017, 245, 1304–1313. [Google Scholar] [CrossRef]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef]
- Bommarius, A.S.; Paye, M.F. Stabilizing Biocatalysts. Chem. Soc. Rev. 2013, 42, 6534–6565. [Google Scholar] [CrossRef]
- Thompson, M.P.; Peñafiel, I.; Cosgrove, S.C.; Turner, N.J. Biocatalysis Using Immobilized Enzymes in Continuous Flow for the Synthesis of Fine Chemicals. Org. Process Res. Dev. 2019, 23, 9–18. [Google Scholar] [CrossRef]
- Tamborini, L.; Fernandes, P.; Paradisi, F.; Molinari, F. Flow Bioreactors as Complementary Tools for Biocatalytic Process Intensification. Trends Biotechnol. 2018, 36, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Enzyme Immobilization: The Quest for Optimum Performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzym. Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme Immobilization: Why, What and How? Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [PubMed]
- DiCosimo, R.; McAuliffe, J.; Poulose, A.J.; Bohlmann, G. Industrial use of immobilized enzymes. Chem. Soc. Rev. 2013, 42, 6437–6474. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Galan, C.; Berenguer-Murcia, A.; Fernandez-Lafuente, R.; Rodrigues, R.C. Potential of Different Immobilization Strategies to Improve the Performance of Enzymes. Adv. Synth. Catal. 2011, 353, 2885–2904. [Google Scholar] [CrossRef]
- Prasad, S.; Roy, I. Converting Enzymes into Tools of Industrial Importance. Recent Pat. Biotechnol. 2018, 12, 33–56. [Google Scholar] [CrossRef]
- Brady, D.; Jordaan, J. Advances in enzyme immobilization. Biotechnol. Lett. 2009, 31, 1639–1650. [Google Scholar] [CrossRef]
- Sheldon, R.A. Cross-linked Enzyme Aggregates(CLEAs) as Industrial Biocatalysts. In Biocatalysis: An Industrial Perspective; de Gonzalo, G., de Maria, P.D., Eds.; Royal Society of Chemistry: London, UK, 2018; Chapter 14. [Google Scholar]
- Sheldon, R.A. Multi-Step Enzyme Catalysis: Biotransformations and Chemoenzymatic Synthesi; Garcia-Junceda, E., Ed.; Wiley-VCH: Weinheim, Germany, 2008; pp. 109–135. [Google Scholar]
- Mateo, C.; Chmura, A.; Rustler, S.; van Rantwijk, F.; Stolz, A.; Sheldon, R.A. Synthesis of enantiomerically pure (S)-mandelic acid using an oxynitrilase-nitrilase bienzymatic cascade: A nitrilase surprisingly shows nitrile hydratase activity. Tetrahedron Asymmetry 2006, 17, 320–323. [Google Scholar] [CrossRef]
- Cao, L. Carrier-Bound Immobilized Enzymes, Principles, Applications, and Design; Wiley-VCH: Weinheim, Germany, 2005. [Google Scholar]
- Truppo, M.D.; Strotman, H.; Hughes, G. Development of an Immobilized Transaminase Capable of Operating in Organic Solvent. ChemCatChem 2012, 4, 1071–1074. [Google Scholar] [CrossRef]
- Cao, L.; van Langen, L.; Sheldon, R.A. Immobilized enzymes: Carrier-bound or carrier-free? Curr. Opin. Biotechnol. 2003, 14, 387–394. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Rantwijk, F.; van Langen, L.M.; Wegman, M.A.; Cao, L.; Janssen, M.H.A. Biocatalysts and Biocatalysis in the Synthesis of ß-Lactam Antibiotics. In Synthesis of β-Lactam Antibiotics: Chemistry, Biocatalysis and Process Integration; Bruggink, A., Ed.; Kluwer Academic: Dordrecht, The Netherlands, 2001; pp. 102–148. ISBN 0-7923-7060-0. [Google Scholar]
- Kallenberg, A.I.; van Rantwijk, F.; Sheldon, R.A. Immobilization of Penicillin G acylase: The Key to Optimum Performance. Adv. Synth. Catal. 2005, 347, 905–926. [Google Scholar] [CrossRef]
- Kosseva, M.R.; Panesar, P.S.; Kaur, G.; Kennedy, J.F. Use of immobilised biocatalysts in the processing of cheese whey. Int. J. Biol. Macromol. 2009, 45, 437–447. [Google Scholar] [CrossRef]
- Thompson, M.P.; Derrington, S.R.; Heath, R.S.; Porter, J.L.; Mangas-Sanchez, J.; Devine, P.N.; Truppo, M.D.; Turner, N.J. A generic platform for the immobilisation of engineered biocatalysts. Tetrahedron 2019, 75, 327–334. [Google Scholar] [CrossRef]
- Margolin, A.; Navia, M.A. Protein crystals as novel catalytic materials. Angew. Chem. Int. Ed. 2001, 40, 2204–2222. [Google Scholar] [CrossRef]
- Cao, L.; van Rantwijk, F.; Sheldon, R.A. Cross-Linked Enzyme Aggregates: A Simple and Effective Method for the Immobilization of Penicillin Acylase. Org. Lett. 2000, 2, 1361–1364. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Cross-linked enzyme aggregates (CLEAs): Stable and recyclable biocatalysts. Biochem. Soc. Trans. 2007, 35, 1583. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Cross-Linked Enzyme Aggregates as Industrial Biocatalysts. Org. Proc. Res. Dev. 2011, 15, 213–223. [Google Scholar] [CrossRef]
- Sheldon, R.A. Characteristic features and biotechnological applications of cross-linked enzyme aggregates (CLEAs). Appl. Microbiol. Biotechnol. 2011, 92, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Kiyota, Y.; Miyazaki, M. Techniques for Preparation of Cross-Linked Enzyme Aggregates and Their Applications in Bioconversions. Catalysts 2018, 8, 174. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; van Langen, L.M.; van Rantwijk, F.; Sheldon, R.A. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol. Bioeng. 2004, 86, 273–276. [Google Scholar] [CrossRef] [PubMed]
- Talekar, S.; Nadar, S.; Joshi, A.; Joshi, G. Pectin cross-linked enzyme aggregates (pectin-CLEAs) of glucoamylase. RSC Adv. 2014, 4, 59444–59453. [Google Scholar] [CrossRef]
- Weiser, D.; Varga, A.; Kovaca, K.; Nagy, F.; Szilagy, A.; Vertessy, B.G.; Paizs, C.; Poppe, L. Bisepoxide Cross-Linked Enzyme Aggregates—New Immobilized Biocatalysts for Selective Biotransformations. ChemCatChem 2014, 6, 1463–1469. [Google Scholar] [CrossRef]
- Pchelintsev, N.A.; Youshko, M.I.; Svedas, V.K. Quantitative characteristic of the catalytic properties and microstructure of cross-linked enzyme aggregates of penicillin acylase. J. Mol. Catal. B Enzym. 2009, 56, 202–207. [Google Scholar] [CrossRef]
- López-Gallego, F.; Betancor, L.; Hidalgo, A.; Alonso, N.; Fernández-Lafuente, R.; Guisán, J.M. Co-aggregation of Enzymes and Polyethyleneimine: A Simple Method to Prepare Stable and Immobilized Derivatives of Glutaryl Acylase. Biomacromolecules 2005, 6, 1839–1842. [Google Scholar] [CrossRef]
- Wilson, L.; Illanes, A.; Abian, O.; Pessela, B.C.C.; Fernandez-Lafuenta, R.; Guisan, J.M. Co-Aggregation of Penicillin G Acylase and Polyionic Polymers: An Easy Methodology To Prepare Enzyme Biocatalysts Stable in Organic Media. Biomacromolecules 2005, 5, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, B.K.; Kumar, S.S.; Golegaonkar, S.B.; Nene, S.N. Preparation of cross-linked enzyme aggregates of L-aminoacylase via co-aggregation with polyethyleneimine. J. Mol. Catal. B Enzym. 2012, 74, 184–191. [Google Scholar] [CrossRef]
- Yan, J.; Gui, X.; Wang, G.; Yan, Y. Improving and Activity of Cross-linked Enzyme Aggregates Based on Polyethylenimine in Hydrolysis of Fish Oil for Enrichment of Polyunsaturated Fatty Acids. Appl. Biochem. Biotechnol. 2012, 166, 925–932. [Google Scholar] [CrossRef]
- Perez, D.I.; van Rantwijk, F.; Sheldon, R.A. Cross-Linked Enzyme Aggregates of Chloroperoxidase: Synthesis, Optimization and Characterization. Adv. Synth. Catal. 2009, 351, 2133–2139. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Miyazaki, M.; Asanomi, Y.; Maeda, H. Poly-lysine supported cross-linked enzyme aggregates with efficient enzymatic activity and high operational stability. Catal. Sci. Technol. 2011, 1, 1256–1261. [Google Scholar] [CrossRef]
- Shah, S.; Sharma, A.; Gupta, M.N. Preparation of cross-linked enzyme aggregates by using bovine serum albumin as a proteic feeder. Anal. Biochem. 2006, 351, 207–213. [Google Scholar] [CrossRef]
- Dong, T.; Zhao, L.; Huang, Y.; Tan, X. Preparation of cross-linked aggregates of aminoacylase fron Aspergillus melleus by using bovine serum albumin as an inert additive. Bioresour. Technol. 2010, 101, 6569–6571. [Google Scholar] [CrossRef] [PubMed]
- Karimpil, J.J.; Melo, J.S.; D‘Souza, S.F. Hen egg white as a feeder protein for lipase immobilization. J. Mol. Catal. B Enzym. 2011, 71, 113–118. [Google Scholar] [CrossRef]
- Goetze, D.; Foletto, E.F.; da Silva, H.B.; Vitória, C.C.; Silveira, V.C.C.; Dal Magro, L.; Rodrigues, R.C. Effect of feather meal as a proteic feeder on combi-CLEA preparation for grape fruit clarification. Proc. Biochem. 2017, 62, 122–127. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S.; Kanbak-Aksu, S.; Rasmussen, J.; Janssen, M.H.A. Cross-Linked Enzyme Aggregates in Organic Synthesis. Aldrichim. Acta 2013, 46, 81–93. [Google Scholar]
- Sheldon, R.A. Industrial Applications of Asymmetric Synthesis using Cross-Linked Enzyme Aggregates. In Comprehensive Chirality; Carreira, E.M., Yamamoto, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; Volume 9, pp. 353–366. [Google Scholar]
- GuauqueTorres, M.P.; Foresti, M.L.; Ferreira, M.L. Cross-linked enzyme aggregates (CLEAs) of selected lipases: A procedure for the proper calculation of their recovered activity. AMB Express 2013, 3, 25. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Jami, S.; Khanahmadi, S. Optimizing the preparation conditions and characterization of a stable and recyclable cross-linked enzyme aggregate (CLEA)-protease. Bioresour. Bioprocess. 2016, 3, 3. [Google Scholar] [CrossRef]
- Montoro-Garcia, S.; Gil-Ortiz, F.; Navarro-Fernandez, J.; Rubio, V.; Garcia-Carmona, F.; Sanchez-Ferrer, A. Improved cross-linked enzyme aggregates for the production of desacetyl beta-lactam antibiotics intermediates. Bioresour. Technol. 2009, 101, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gonzalez, L.; Garcia-Carmona, F. Characterization of cross-linked immobilized arylesterase from Gluconobacter oxydans 621H with activity toward cephalosporin C and 7-aminocephalosporanic acid. Biotechnol. Prog. 2016, 32, 36–42. [Google Scholar] [CrossRef]
- Park, H.J.; Uhm, K.N.; Kim, H.K. Biotransformation of amides to acids using a co-cross-linked enzyme aggregate of Rhodococcus erythropolis amidase. J. Microbiol. Biotechnol. 2010, 20, 325–331. [Google Scholar]
- Malandra, A.; Cantarella, M.; Kaplan, O.; Vejvoda, V.; Uhnakova, B.; Stepankova, B.; Kubac, D.; Martinkova, L. Continuous hydrolysis of 4-cyanopyridine by nitrilases from Fusarium solani O1 and Aspergillus niger K10. Appl. Microbiol. Biotechnol. 2009, 85, 277–284. [Google Scholar] [CrossRef]
- Yu, C.-Y.; Li, X.-F.; Lou, W.-Y.; Zong, M.H. Cross-linked enzyme aggregates of Mung bean epoxide hydrolases: A highly active, stable and recyclable biocatalyst for asymmetric hydrolysis of epoxides. J. Biotechnol. 2013, 166, 12–19. [Google Scholar] [CrossRef]
- Cruz, J.; Barbosa, O.; Rodrigues, R.C.; Fernandez-Lafuente, R.; Rodrigo Torres, R.; Claudia Ortiz, C. Optimized preparation of CALB-CLEAs by response surface methodology: The necessity to employ a feeder to have an effective crosslinking. J. Mol. Catal. B Enzym. 2012, 80, 7–14. [Google Scholar] [CrossRef]
- Guauque Torres, M.P.; Foresti, M.L.; Ferreira, M.L. CLEAs of Candida antarctica lipase B (CaLB) with a bovine serum albumin (BSA) cofeeder core: Study of their catalytic activity. Biochem. Eng. J. 2014, 90, 36–43. [Google Scholar] [CrossRef]
- Guauque Torres, M.P.; Foresti, M.L.; Ferreira, M.L. Effect of different parameters on the hydrolytic activity of cross-linked enzyme aggregates (CLEAs) of lipase from Thermomyces lanuginose. Biochem. Eng. J. 2013, 72, 18–23. [Google Scholar] [CrossRef]
- Lai, J.-Q.; Hu, Z.-L.; Sheldon, R.A.; Yang, Z. Catalytic performance of cross-linked enzyme aggregates of Penicillium expansum lipase and their use as catalyst for biodiesel production. Proc. Biochem. 2012, 47, 2058–2063. [Google Scholar] [CrossRef]
- Rehman, S.; Bhatti, H.N.; Bilal, M.; Asgher, M. Cross-linked enzyme aggregates (CLEAs) of Pencilluim notatum lipaseenzyme with improved activity, stability and reusabilitycharacteristics. Int. J. Biol. Macromol. 2016, 91, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Lozano, S.; López-Gallego, F.; Rocha-Martin, J.; Guisán, J.M.; Favela-Torres, E. Improving enantioselectivity of lipase from Candida rugosa bycarrier-bound and carrier-free immobilization. J. Mol. Catal. B Enzym. 2016, 130, 32–39. [Google Scholar] [CrossRef]
- Chavez, G.; Rasmussen, J.-A.; Janssen, M.H.A.; Mamo, G.; Hatti-Kaul, R.; Sheldon, R.A. Baeyer-Villiger Oxidation of Cyclohexanone in Aqueous Medium with In Situ Generation of Peracid Catalyzed by Perhydrolase CLEA. Top. Catal. 2014, 57, 349–355. [Google Scholar] [CrossRef]
- Schroeder, H.; Strohmeier, G.A.; Leypold, M.; Nuijens, T.; Quaedflieg, P.J.L.M.; Breinbauer, R. Racemization-Free Chemoenzymatic Peptide Synthesis Enabled by the Ruthenium-Catalyzed Synthesis of Peptide Enol Esters via Alkyne-Addition and Subsequent Conversion Using Alcalase-Cross-Linked Enzyme Aggregates. Adv. Synth. Catal. 2013, 355, 1799–1807. [Google Scholar] [CrossRef]
- Illanes, A.; Wilson, L.; Caballero, E.; Fernandez-Lafuenta, R.; Guisan, J.M. Crosslinked penicillin acylase aggregates for synthesis of β-lactam antibiotics in organic medium. Appl. Biochem. Biotechnol. 2006, 133, 189–202. [Google Scholar] [CrossRef]
- Illanes, A.; Wilson, L.; Altamirano, C.; Cabrera, Z.; Alvarez, L.; Aguirre, C. Production of cephalexin in organic medium at high substrate concentrations with CLEA of penicillin acylase and PGA-450. Enzym. Microb. Technol. 2007, 40, 195–203. [Google Scholar] [CrossRef]
- Contesini, F.; de Alencar Figueira, J.; Kawaguti, H.; de Barros Fernandes, P.; de Oliveira Carvalho, P.; da Graça Nascimento, M.; Sato, H. Potential Applications of Carbohydrases Immobilization in the Food Industry. Int. J. Mol. Sci. 2013, 14, 1335–1369. [Google Scholar] [CrossRef] [Green Version]
- Gauer, R.; Paul, H.; Jain, R.; Khare, S.K. Galacto-oligosaccharide synthesis by immobilized Aspergillus oryzae β-galactosidase. Food Chem. 2006, 97, 426–430. [Google Scholar] [CrossRef]
- Li, L.; Li, G.; Cao, L.; Ren, G.; Kong, W.; Wang, S.; Guo, G.; Liu, Y. Characterization of the cross-linked enzyme aggregates of a novel β-galactosidase, a potential catalyst for the synthesis of galacto-oligosaccharides. J. Agric. Food Chem. 2015, 63, 894–901. [Google Scholar] [CrossRef]
- Ribeiro, M.H.L.; Rabaça, M. Cross-Linked Enzyme Aggregates of Naringinase: Novel Biocatalysts for Naringin Hydrolysis. Enzym. Res. 2011, 2011, 851272. [Google Scholar] [CrossRef]
- Chen, L.; Hu, Y.-D.; Li, N.; Zong, M.-H. Cross-linked enzyme aggregates of β-glucosidase from Prunus domestica seeds. Biotechnol. Lett. 2012, 34, 1673–1678. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Soto, M.E.; Rudiño-Piñera, E.; Rodriguez-Alegria, M.E.; Manguia, A.L. Evaluation of cross-linked aggregates from purified Bacillus subtilis levansucrase mutants for transfructosylation reactions. BMC Biotechnol. 2009, 9, 68. [Google Scholar] [CrossRef]
- Cerdobbel, A.; De Winter, K.; Desmet, T.; Soetaert, W. Sucrose phosphorylase as cross-linked enzyme aggregate: Improved thermal stability for industrial applications. Biotechnol. J. 2010, 5, 1192–1197. [Google Scholar] [CrossRef] [Green Version]
- De Winter, K.; Soetaert, W.; Desmet, T. An Imprinted Cross-Linked Enzyme Aggregate (iCLEA) of Sucrose Phosphorylase: Combining Improved Stability with Altered Specificity. Int. J. Mol. Sci. 2012, 13, 11333–11342. [Google Scholar] [CrossRef] [Green Version]
- Talekar, S.; Ghodake, V.; Kate, A.; Samant, N.; Kumar, C.; Gadagkar, S. Preparation and Characterization of Cross-linked Enzyme Aggregates of Saccharomyces Cerevisiae Invertase. Aust. J. Basic Appl. Sci. 2010, 4, 4760–4765. [Google Scholar]
- Diaz-Rodriguez, A.; Lavandera, I.; Kanbak-Aksu, S.; Sheldon, R.A.; Gotor, V.; Gotor-Fernandez, V. From Diols to Lactones under Aerobic Conditions Using a Laccase/TEMPO Catalytic System in Aqueous Medium. Adv. Synth. Catal. 2012, 354, 3405–3408. [Google Scholar] [CrossRef]
- Matijosyte, I.; Arends, I.W.C.E.; de Vries, S.; Sheldon, R.A. Preparation and use of cross-linked enzyme aggregates (CLEAs) of laccase. J. Mol. Catal. B Enzym. 2010, 62, 142–148. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, Y.-W. Optimizing the preparation conditions and characterization of cross-linked enzyme aggregates of a monoamine oxidase. Food Sci. Biotechnol. 2016, 25, 1421–1425. [Google Scholar] [CrossRef]
- Roberge, C.; Amos, D.; Pollard, D.; Devine, P. Preparation and application of cross-linked aggregates of chloroperoxidase with enhanced hydrogen peroxide tolerance. J. Mol. Catal. B Enzym. 2009, 56, 41–45. [Google Scholar] [CrossRef]
- Sulek, F.; Perez Fernandez, D.; Kneza, Z.; Habulina, M.; Sheldon, R.A. Immobilization of horseradish peroxidase as crosslinked enzyme aggregates (CLEAs). Process Biochem. 2011, 46, 765–769. [Google Scholar] [CrossRef]
- Morales, A.; Barbosa, O.; Rueda, N.; Fonseca, Z.; Torres, R.; Rodriguez, R.C.; Ortiz, C.; Fernandez-Lafuenta, R. Optimization and characterization of CLEAs of the very thermostable dimeric peroxidase from Roystonea regia. RSC Adv. 2015, 5, 53047–53053. [Google Scholar] [CrossRef]
- Kim, M.H.; Park, S.; Kim, Y.H.; Won, K.; Lee, S.H. Immobilization of formate dehydrogenase from Candida boidinii through cross-linked enzyme aggregates. J. Mol. Catal. B Enzym. 2013, 97, 209–214. [Google Scholar] [CrossRef]
- Sinirlioglu, Z.A.; Sinirlioglu, D.; Akbas, F. Preparation and characterization of stable cross-linked enzyme aggregates of novel laccase enzyme from Shewanella putrefaciens and using malachite green decolorization. Bioresour. Technol. 2013, 146, 807–811. [Google Scholar] [CrossRef]
- Ba, S.; Arsenault, A.; Hassani, T.; Jones, J.P.; Cabana, H. Laccase immobilization and insolubilization: From fundamentals to applications for the elimination of emerging contaminants in wastewater treatment. Crit. Rev. Biotechnol. 2013, 33, 404–418. [Google Scholar] [CrossRef]
- Cabana, H.; Jones, J.P.; Agathos, S.N. Preparation and characterization of cross-linked laccase aggregates and their application to the elimination of endocrine disrupting chemicals. J. Biotechnol. 2007, 132, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Cabirol, F.L.; Lim, A.E.C.; Hanefeld, U.; Sheldon, R.A. Straightforward Enzymatic Process Based on HNL CLEA-Catalysis towards Cyanohydrin Derivatives. Org. Proc. Res. Dev. 2010, 14, 114–118. [Google Scholar] [CrossRef]
- Alagoz, D.; Tukel, S.S.; Yildirim, D. Enantioselective Synthesis of Various Cyanohydrins Using Covalently Immobilized Preparations of Hydroxynitrile Lyase from Prunus dulcis. Appl. Biochem. Biotechnol. 2015, 177, 1348–1363. [Google Scholar] [CrossRef]
- Yildirim, D.; Tuekel, S.S.; Alagoez, D. Crosslinked enzyme aggregates of hydroxynitrile lyase partially purified from Prunus dulcis seeds and its application for the synthesis of enantiopure cyanohydrins. Biotechnol. Prog. 2014, 30, 818–826. [Google Scholar] [CrossRef]
- Cabirol, F.L.; Hanefeld, U.; Sheldon, R.A. Immobilized hydroxynitrile lyases for enantioselective ynthesis of cyanohydrins: Sol-gels and cross-linked enzyme aggregates. Adv. Synth. Catal. 2006, 348, 1645–1654. [Google Scholar] [CrossRef]
- Chmura, A.; van der Kraan, G.M.; Kielar, F.; van Langen, L.M.; van Rantwijk, F.; Sheldon, R.A. Cross-linked aggregates of the hydroxynitrile lyase from Manihot esculenta: Highly active and robust Biocatalysts. Adv. Synth. Catal. 2006, 348, 1655–1661. [Google Scholar] [CrossRef]
- Cabirol, F.L.; Tan, P.L.; Tay, B.; Cheng, S.; Hanefeld, U.; Sheldon, R.A. Linum usitatissimum hydroxynitrile lyase cross-linked enzyme aggregates: A recyclable enantioselective catalysis. Adv. Synth. Catal. 2008, 350, 2329–2338. [Google Scholar] [CrossRef]
- Roberge, C.; Fleitz, F.; Pollard, D.; Devine, P. Asymmetric synthesis of cyanohydrin derived from pyridine aldehyde with cross-linked aggregates of hydroxynitrile lyases. Tetrahedron Lett. 2007, 48, 1473–1477. [Google Scholar] [CrossRef]
- Lanfranchi, E.; Grill, B.; Raghoeber, Z.; Van Pelt, S.; Sheldon, R.A.; Steiner, K.; Glieder, A.; Winkler, M. Production of hydroxynitrile lyase from davallia tyermannii (DtHNL) in Komagataella phaffii and its immobilization as a CLEA to generate a robust biocatalyst. ChemBioChem 2018, 19, 312–316. [Google Scholar] [CrossRef]
- Jangir, N.; Padhi, S.K. Immobilized Baliospermum montanum hydroxynitrile lyase catalyzed suynthesis of chiral cyanohydrins. Bioorg. Chem. 2019, 84, 32–40. [Google Scholar] [CrossRef]
- Garcia-Garcia, M.I.; Sola-Carvajal, A.; Garcia Carmona, F.; Sanchez Ferrer, A. Improved production of 2-keto-3-deoxy-d-glycero-galactononulosonic acid (KDN) using FastPrep-CLEAs. Process Biochem. 2014, 49, 90–94. [Google Scholar] [CrossRef]
- Garcia-Garcia, M.I.; Sola-Carvajal, A.; Sanchez-Carron, G.; Garcia-Carmona, F.; Sanchez-Ferrer, A. New stabilized FastPrep-CLEAs for sialic acid synthesis. Bioresour. Technol. 2011, 102, 6186–6191. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.D.; Zhang, S.; Sun, L.M. Cross-Linked Enzyme Aggregates of Phenylalanine Ammonia Lyase: Novel Biocatalysts for Synthesis of L-Phenylalanine. Appl. Biochem. Biotechnol. 2012, 167, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.D.; Sun, L.M.; Li, L.L. A Simple Technique of Preparing Stable CLEAs of Phenylalanine Ammonia Lyase Using Co-aggregation with Starch and Bovine Serum Album. Appl. Biochem. Biotechnol. 2013, 170, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, S.; van Rantwijk, F.; Sheldon, R.A. Nitrile hydratases in synthesis. Chim. Oggi 2008, 26, 2. [Google Scholar]
- Van Pelt, S.; Quignard, S.; Kubac, D.; Sorokin, D.Y.; van Rantwijk, F.; Sheldon, R.A. Nitrile hydratase CLEAs: The immobilization and stabilization of an industrially important enzyme. Green Chem. 2008, 10, 395–400. [Google Scholar] [CrossRef]
- Van Langen, L.M.; Selassa, R.P.; van Rantwijk, F.; Sheldon, R.A. Cross-Linked Aggregates of (R)-Oxynitrilase: A Stable, Recyclable Biocatalyst for Enantioselective Hydrocyanation. Org. Lett. 2005, 7, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Betancor, L.; Fernandez-Lorente, G.; Fuentes, M.; Hidalgo, A.; Guisan, J.M.; Pessela, B.C.C.; Fernandez-Lafuente, R. Cross-Linked Aggregates of Multimeric Enzymes: A Simple and Efficient Methodology To Stabilize Their Quaternary Structure. Biomacromolecules 2004, 5, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Du, X.; Jiang, W.; Tong, Y.; Zhao, Z.; Fang, R.; Feng, J. Cross-Linked Enzyme Aggregates (CLEAs) of halohydrin dehalogenase from Agrobacterium radiobacter AD1: Preparation, Characterization and application as a biocatalyst. J. Biotechnol. 2018, 272–273, 48–55. [Google Scholar] [CrossRef]
- Schoevaart, R.; Wolbers, M.W.; Golubovic, M.; Ottens, M.; Kieboom, P.G.; van Rantwijk, F.; Van der Wielen, L.A.M.; Sheldon, R.A. Preparation, optimization, and structures of cross-linked enzyme aggregates (CLEAs). Biotechnol. Bioeng. 2004, 87, 754–762. [Google Scholar] [CrossRef]
- Khanahmadi, S.; Yusof, F.; Amid, A.; Mahmod, S.S.; Mahat, M.K. Optimized preparation and characterization of CLEA-lipase from cocoa pod husk. J. Biotechnol. 2015, 202, 153–161. [Google Scholar] [CrossRef]
- Talekar, S.; Joshi, A.; Joshi, G.; Kamat, P.; Haripurkar, R.; Kambale, S. Parameters in preparation and characterization of cross linked enzyme aggregates (CLEAs). RSC Adv. 2013, 3, 12485–12511. [Google Scholar] [CrossRef]
- Cui, J.D.; Ja, S.R. Optimization protocols and improved strategies of cross-linked enzyme aggregates technology: Current development and future challenges. Crit. Rev. Biotechnol. 2015, 35, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Valesco-Lozano, S.; Lopez-Gallego, F.; Mateoes-Diaz, J.C.; Favela-Torres, E. Cross-linked enzyme aggregates (CLEA) in enzyme improvement—A review. Biocatalysis 2015, 1, 166–177. [Google Scholar] [CrossRef]
- Talekar, S.; Shah, V.; Patil, S.; Nimbalkar, M. Porous cross linked enzyme aggregates (p-CLEAs) of Saccharomyces cerevisiae invertase. Catal. Sci. Technol. 2012, 2, 1575–1579. [Google Scholar] [CrossRef]
- Wang, M.; Jia, C.; Qi, W.; Yu, Q.; Peng, X.; Su, R.; He, Z. Porous-CLEAs of papain: Application to enzymatic hydrolysis of macromolecules. Bioresour. Technol. 2010, 102, 3541–3545. [Google Scholar] [CrossRef] [PubMed]
- Vinoth Kumar, V.; Prem Kumar, M.P.; Thiruvenkadaravi, K.V.; Baskaralingram, P.; Senthil Kumar, P.; Sivanesan, S. Preparation and characterization of porous cross linked laccase aggregates for the decolorization of triphenyl methane and reactive dyes. Bioresour. Technol. 2012, 119, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Q.; Wang, M.; Qi, W.; Su, R.; He, Z. Preparation of β-mannanase CLEAs using macromolecular cross-linkers. Catal. Sci. Technol. 2013, 3, 1937–1941. [Google Scholar] [CrossRef]
- Brady, D.; Jordaan, J.; Simpson, C.; Chetty, A.; Arumugam, C.; Moolman, F. Spherezymes: A novel structured self-immobilisation enzyme technology. BMC Biotechnol. 2008, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Mbanjwa, M.B.; Land, K.J.; Windvoel, T.; Papala, P.M.; Korvink, J.G.; Visser, D.; Brady, D. Production of self-immobilised enzyme microspheres using microfluidics. Proc. Biochem. 2018, 69, 75–81. [Google Scholar] [CrossRef]
- Lopez-Serrano, P.; Cao, L.; van Rantwijk, F.; Sheldon, R.A. Cross-linked enzyme aggregates with enhanced activity: Application to lipases. Biotechnol. Lett. 2002, 24, 1379–1383. [Google Scholar] [CrossRef]
- Sorgedrager, M.J.; Verdoes, D.; van der Meer, H.; Sheldon, R.A. Cross-linked enzyme aggregates in a membrane slurry reactor. Continuous production of 6-APA by enzymatic hydrolysis of penicillin. Chim. Oggi 2008, 26, 23–25. [Google Scholar]
- Cabana, H.; Jones, J.P.; Agathos, S.N. Utilization of cross-linked laccase aggregates in a perfusion basket reactor for the continuous elimination of endocrine-disrupting chemicals. Biotechnol. Bioeng. 2009, 102, 1582–1592. [Google Scholar] [CrossRef]
- Hilterhaus, L.; Thum, O.; Liese, A. Reactor Concept for Lipase-Catalyzed Solvent-Free Conversion of Highly Viscous Reactants Forming Two-Phase Systems. Org. Process Res. Dev. 2008, 12, 618–625. [Google Scholar] [CrossRef]
- Hickey, A.M.; Marle, L.; McCreedy, T.; Watts, P.; Greenway, G.M.; Littlechild, J.A. Immobilization of thermophilic enzymes in miniaturized flow reactors. Biochem. Soc. Trans. 2007, 35, 1621–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.-Q.; Wang, S.S.; Li, L.-N.; Gao, J.; Zhang, Y.-W. Combined Cross-linked Enzyme Aggregates as Biocatalysts. Catalysts 2018, 8, 460. [Google Scholar] [CrossRef]
- Dalal, S.; Kapoor, M.; Gupta, M.N. Preparation and characterization of combi-CLEAs catalyzing multiple non-cascade reactions. J. Mol. Catal. B Enzym. 2007, 44, 128–132. [Google Scholar] [CrossRef]
- Dalal, S.; Sharma, A.; Gupta, M.N. A multipurpose immobilized biocatalyst with pectinase, xylanase and cellulase activities. Chem. Cent. J. 2007, 1, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Magro, L.; Hertz, P.F.; Fernandez-Lafuenta, R.; Klein, M.P.; Rodrigues, R.C. Preparation and characterization of a Combi-CLEA from pectinases and cellulases: Potential biocatalyst for grape juice clarification. RSC Adv. 2016, 6, 27242–27251. [Google Scholar] [CrossRef]
- Touahar, I.E.; Haroune, L.; Ba, S.; Belenger, J.-P.; Cabana, H. Characterization of combined cross-linked enzyme aggregates from laccase, versatile peroxidase and glucose oxidase and their utilization for the elimination of pharmaceuticals. Sci. Total Environ. 2014, 481, 90–99. [Google Scholar] [CrossRef]
- Ba, S.; Mialon, A.; Jones, J.P.; Cabana, H. Laccase insolubilized as combination of crosslinked enzyme aggregates: Characterization and potential application in wastewater treatment. J. Enzym. Res. 2012, 3, 23–28. [Google Scholar]
- Vafiadi, C.; Topakas, E.; Christakopoulos, P. Preparation of multipurpose cross-linked enzyme aggregates and their application to production of alkyl ferulates. J. Mol. Catal. B Enzym. 2008, 54, 35–41. [Google Scholar] [CrossRef]
- Mahmod, S.S.; Yusof, F.; Jami, M.S.; Khanahmadi, S.; Shah, H. Development of an immobilized biocatalyst with lipase and protease activities as a multipurpose cross-linked enzyme aggregate. Process. Biochem. 2015, 50, 2144–2157. [Google Scholar] [CrossRef]
- Muschiol, J.; Peters, C.; Oberleitner, N.; Mihovilovic, M.D.; Bornscheuer, U.T.; Rudroff, F. Cascade catalysis--strategies and challenges en route to preparative synthetic biology. Chem. Commun. 2015, 51, 5798–5811. [Google Scholar] [CrossRef]
- Schrittweiser, J.H.; Sattler, J.; Resch, V.; Mutti, F.G.; Kroutil, W. Recent biocatalytic oxidation-reduction cascades. Curr. Opin. Chem. Biol. 2011, 15, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Woodley, J.M. Process technology for multi-enzymatic reaction systems. Bioresour. Technol. 2012, 115, 183–195. [Google Scholar] [CrossRef]
- Santacoloma, P.A.D.; Sin, G.; Gernacy, K.V.; Woodley, J.M. Multienzyme-Catalyzed Processes: Next-Generation Biocatalysis. Org. Proc. Res. Dev. 2011, 15, 203–212. [Google Scholar] [CrossRef]
- Land, H.; Hendit-Forssell, P.; Martinelle, M.; Berglund, P. One-pot biocatalytic amine transaminase/acyl transferase cascade for aqueous formation of amides from aldehydes or ketones. Catal. Sci. Technol. 2016, 6, 2897–2900. [Google Scholar] [CrossRef] [Green Version]
- Quin, M.B.; Wallin, K.K.; Zhang, G.; Schmidt-Dannert, C. Spatial organization of multi-enzyme biocatalytic cascades. Org. Biomol. Chem. 2017, 15, 4260–4271. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Yang, K.-L. Combined cross-linked enzyme aggregates of horseradish peroxidase and glucose oxidase for catalyzing cascade chemical reactions. Enzym. Microb. Technol. 2017, 100, 52–59. [Google Scholar] [CrossRef]
- Taboada-Puig, R.; Junghanns, C.; Demarche, P.; Moreira, M.T.; Feijoo, G.; Lema, J.M.; Agathos, S.N. Combined cross-linked enzyme aggregates from versatile peroxidase and glucose oxidase: Production, partial characterization and application for the elimination of endocrine disruptors. Bioresour. Technol. 2011, 102, 6593–6599. [Google Scholar] [CrossRef]
- Ning, C.; Su, E.; Tian, Y.; Wei, D. Combined cross-linked enzyme aggregates (combi-CLEAs) for efficient integration of a ketoreductase and a cofactor regeneration system. J. Biotechnol. 2014, 184, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Liu, L.; Chen, D.; Wang, Y.; Zhang, J.; Shao, L. Co-expression of the recombined alcohol dehydrogenase and glucose dehydrogenase and cross-linked enzyme aggregates stabilization. Bioresour. Technol. 2017, 224, 531–535. [Google Scholar] [CrossRef]
- Li, H.; Xiao, W.; Xie, P.; Zheng, L. Co-immobilization of enoate reductase with a cofactor-recycling partner enzyme. Enzym. Microb. Technol. 2018, 109, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Chmura, A.; Rustler, S.; Paravidino, M.; van Rantwijk, F.; Stolz, A.; Sheldon, R.A. The combi-CLEA approach: Enzymatic cascade synthesis of enantiomerically pure (S)-mandelic acid. Tetrahedron Asymmetry 2013, 24, 1225–1232. [Google Scholar] [CrossRef]
- Van Rantwijk, F.; Stolz, A. Enzymatic cascade synthesis of (S)-2-hydroxycarboxylic amides and acids: Cascade reactions employing a hydroxynitrile lyase, nitrile-converting enzymes and an amidase. J. Mol. Catal. B Enzym. 2015, 114, 25–30. [Google Scholar] [CrossRef]
- Van Pelt, S.; van Rantwijk, F.; Sheldon, R.A. Synthesis of Aliphatic (S)-α-Hydroxycarboxylic Amides using a One-Pot Bienzymatic Cascade of Immobilised Oxynitrilase and Nitrile Hydratase. Adv. Synth. Catal. 2009, 351, 397–404. [Google Scholar] [CrossRef]
- Stressler, T.; Ewert, J.; Eisele, T.; Fischer, L. Cross-linked enzyme aggregates (CLEAs) of PepX and PepN-production, partial characterization and application of combi-CLEAs for milk protein hydrolysis. Biocatal. Agric. Biotechnol. 2015, 50, 752–760. [Google Scholar] [CrossRef]
- Jung, D.-H.; Jung, J.-H.; Seo, D.-H.; Ha, S.-J.; Kweon, D.-K.; Park, C.-S. One-pot conversion of sucrose to trehalose using enzymatic sequential reactions in combined cross-linked enzyme aggregates. Bioresour. Technol. 2013, 130, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Ahumada, K.; Urrutia, P.; Illanes, A.; Wilson, L. Production of combi-CLEAs of glycosidases utilized for aroma enhancement in wine. Food Bioprod. Process. 2015, 94, 555–560. [Google Scholar] [CrossRef]
- Talekar, S.; Waingade, S.; Gaikwad, V.; Patil, S.; Nagavekar, N. Preparation and characterization of crosslinked enzyme aggregates (CLEAs) of Bacillus amyloliquefaciens alpha amylase. J. Biochem. Technol. 2012, 3, 349–353. [Google Scholar]
- Sahutoglu, A.S.; Akgul, C. Immobilisation of Aspergillus oryzae α-amylase and Aspergillus niger glucoamylase enzymes as cross-linked enzyme aggregates. Chem. Pap. 2015, 69, 433–439. [Google Scholar] [CrossRef]
- Nadar, S.S.; Muley, A.B.; Ladole, M.R.; Joshi, P.U. Macromolecular cross-linked enzyme aggregates (m-CLEAs) of α-amylase. Int. J. Biol. Macromol. 2016, 84, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-D.; Wu, J.; Jia, D.-C.; Wan, Y.H.; Yang, N.; Qiao, M. Preparation of Cross-Linked Glucoamylase Aggregates Immobilization by Using Dextrin and Xanthan Gum as Protecting Agents. Catalysts 2016, 6, 77. [Google Scholar] [CrossRef]
- Talekar, S.; Desai, S.; Pillai, M.; Nagavekar, N.; Ambarkar, S.; Surnis, S.; Ladole, M.; Nadar, S.; Mulla, N. Carrier free co-immobilization of glucoamylase and pullulanase as combi-cross linked enzyme aggregates (combi-CLEAs). RSC Adv. 2013, 3, 2265–2271. [Google Scholar] [CrossRef]
- Talekar, S.; Pandharbale, A.; Ladole, M.; Nadar, S.; Mulla, M.; Japhalekar, K.; Pattankude, K.; Arage, D. Carrier free co-immobilization of alpha amylase, glucoamylase and pullulanase as combined cross-linked enzyme aggregates (combi-CLEAs). Bioresour. Technol. 2013, 147, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Bhattachariya, A.S.; Bhattachariya, A.; Pletschke, B.I. Synergism of fungal and bacterial cellulases and hemicellulases: A novel perspective for enhanced bio-ethanol production. Biotechnol. Lett. 2015, 37, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Bayer, E.A.; Belaich, J.-P.; Shoham, Y.; Lamed, R. The Cellulosomes: Multienzyme Machines for Degradation of Plant Cell Wall Polysaccharides. Ann. Rev. Microbiol. 2004, 58, 521–554. [Google Scholar] [CrossRef] [PubMed]
- Bhattachariya, A.; Pletschke, B.I. Strategic optimization of xylanase-mannanase combi-CLEAs for synergistic and efficient hydrolysis of complex lignocellulosic substrates. J. Mol. Catal. B Enzym. 2015, 115, 140–150. [Google Scholar] [CrossRef]
- Periyasamy, K.; Santhalembi, L.; Mortha, G.; Aurousseau, M.; Subramanian, S. Carrier-free co-immobilization of xylanase, cellulase and β-1,3-glucanase as combined cross-linked enzyme aggregates (combi-CLEAs) for one-pot saccharification of sugarcane bagasse. RSC Adv. 2016, 6, 32849–32857. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Sorgedrager, M.J.; Kondor, B. Non-Leachable Magnetic Cross-Linked Enzyme Aggregate. WO Patent 2012/023847 A2, 23 February 2012. [Google Scholar]
- Whitesides, G.M.; Hill, C.L.; Brunie, J.-C. Magnetic Filtration of Small Heterogeneous Catalyst Particles. Preparation of Ferrimagnetic Catalyst Supports. Ind. Eng. Chem. Process Des. Dev. 1976, 15, 226–227. [Google Scholar] [CrossRef]
- Cui, J.D.; Cui, L.L.; Zhang, S.P.; Zhang, Y.F.; Su, Z.G.; Ma, G.H. Hybrid Magnetic Cross-Linked Enzyme Aggregates of Phenylalanine Ammonia Lyase from Rhodotorula glutinis. PLoS ONE 2014, 9, e97221. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, S.; Janssen, M.H.A.; Rasmussen, J.-A.M.; Sorgedrager, M.J.; Sheldon, R.A.; Koning, P. Cross-Linked Enzyme Aggregates Comprising Magnetizable Particles. WO Patent 2018/004341 A1, 4 January 2018. [Google Scholar]
- Talekar, S.; Ghodake, V.; Ghotage, T.; Rathod, P.; Deshmukh, P.; Nadar, S.; Mulla, M.; Ladole, M. Novel magnetic cross-linked enzyme aggregates (magnetic CLEAs) of alpha amylase. Bioresour. Technol. 2012, 123, 542–547. [Google Scholar] [CrossRef]
- Nadar, S.S.; Rathod, V.K. Magnetic macromolecular cross linked enzyme aggregates (CLEAs) of glucoamylase. Enzym. Microb. Technol. 2016, 83, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Kumar Jana, A.; Kumar, S.; Maiti, M. Immobilization of amyloglucosidase from SSF of Aspergillus niger by crosslinked enzyme aggregate onto magnetic nanoparticles using minimum amount of carrier and characterizations. J. Mol. Catal. B Enzym. 2013, 98, 30–36. [Google Scholar] [CrossRef]
- Amaral-Fonseca, M.; Kopp, W.; de Lima Camargo Giordano, R.; Fernandez-Lafuenta, R.; Tardioli, P.W. Preparation of Magnetic Cross-Linked Amyloglucosidase Aggregates: Solving Some Activity Problems. Catalysts 2018, 8, 496. [Google Scholar] [CrossRef]
- Asgher, M.; Shahid, M.; Kamal, S.; Iqbal, H.M.N. Recent trends and valorization of immobilization strategies and ligninolytic enzymes by industrial biotechnology. J. Mol. Catal. B Enzym. 2014, 101, 56–66. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Pletschke, B.I. Magnetic cross-linked enzyme aggregates (CLEAs): A novel concept towards carrier free immobilization of lignocellulolytic enzymes. Enzym. Microb. Technol. 2014, 61–62, 17–27. [Google Scholar] [CrossRef]
- Khorshadi, K.J.; Lenjannezhadian, H.; Jamalan, M.; Zeinali, M. Preparation and characterization of nanomagnetic cross-linked cellulase aggregates for cellulose bioconversion. J. Chem. Technol. Biotechnol. 2016, 91, 539–546. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, W.; Yang, Z.; Yang, X.; Wang, N.; Yu, X. Novel Magnetic Cross-Linked Cellulase Aggregates with a Potential Application in Lignocellulosic Biomass Conversion. Molecules 2017, 22, 269. [Google Scholar] [CrossRef]
- Hwangbo, M.; Tran, J.L.; Chu, K.-H. Effective one-step saccharification of lignocellulosic biomass using magnetite-biocatalysts containing saccharifying enzymes. Sci. Total Environ. 2019, 647, 806–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaarani, S.M.; Jahim, J.M.; Rahman, R.A.; Idris, A.; Murad, A.M.A.; Illias, R.M. Silanized maghemite for cross-linked enzyme aggregates of recombinant xylanase from Trichoderma reesei. J. Mol. Catal. B Enzym. 2016, 133, 65–76. [Google Scholar] [CrossRef]
- Olofsson, K.; Bertilsson, M.; Liden, G. A short review on SSF—An interesting process option for ethanol production from lignocellulosic feedstocks. Biotechnol. Biofuels 2008, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Sojitra, U.V.; Nadar, S.S.; Rathod, V.K. A magnetic tri-enzyme nanobiocatalyst for fruit juice clarification. Food Chem. 2016, 213, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Pfaltzgraff, L.A.; De Bruyn, M.; Cooper, E.C.; Budarin, V.; Clark, J.H. Food waste biomass: A resource for high-value chemicals. Green Chem. 2013, 15, 307–314. [Google Scholar] [CrossRef]
- Mehde, A.A.; Mehdi, W.A.; Severgün, O.; Çakar, S.; Özacar, M. Lipase-based on starch material as a development matrix with magnetite cross-linked enzyme aggregates and its application. Int. J. Biol. Macromol. 2018, 120, 1533–1543. [Google Scholar] [CrossRef]
- Cruz-Izquierdo, A.; Pico, E.A.; Lopez, C.; Serra, J.L.; Lama, M.J. Magnetic Cross-Linked Enzyme Aggregates (mCLEAs) of Candida antarctica Lipase: An Efficient and Stable Biocatalyst for Biodiesel Synthesis. PLoS ONE 2014, 9, e115202. [Google Scholar] [CrossRef]
- Picó, E.A.; López, C.; Cruz-Izquierdo, A.; Munarriz, M.; Iruretagoyena, F.J.; Serra, J.L.; Llama, M.J. Easy reuse of magnetic cross-linked enzyme aggregates of lipase B from Candida antarctica to obtain biodiesel from Chlorella vulgaris lipids. J. Biosci. Bioeng. 2018, 126, 451–457. [Google Scholar] [CrossRef]
- Zhang, W.W.; Yang, X.L.; Jia, J.Q.; Wang, N.; Hu, C.J.; Yu, X.Q. Surfactant-activated magnetic cross-linked enzyme aggregates (magnetic CLEAs) of Thermomyces lanuginosus lipase for biodiesel production. J. Mol. Catal. B Enzym. 2015, 115, 83–89. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, H.; Liu, W.; Wang, N.; Yu, X. Improved Performance of Magnetic Cross-Linked Lipase Aggregates by Interfacial Activation: A Robust and Magnetically Recyclable Biocatalyst for Transesterification of Jatropha Oil. Molecules 2017, 22, 2157. [Google Scholar] [CrossRef]
- Cui, J.; Cui, L.; Jia, S.; Su, Z.; Zhang, S. Hybrid Cross-Linked Lipase Aggregates with Magnetic Nanoparticles: A Robust and Recyclable Biocatalyst for the Epoxidation of Oleic Acid. J. Agric. Food Chem. 2016, 64, 7179–7187. [Google Scholar] [CrossRef]
- Tudorache, M.; Gheorghe, A.; Viana, A.S.; Parvulescu, V.I. Biocatalytic epoxidation of α-pinene to oxy-derivatives over cross-linked lipase aggregates. J. Mol. Catal. B Enzym. 2016, 134, 9–15. [Google Scholar] [CrossRef]
- Tudorache, M.; Nae, A.; Coman, S.; Parvulescu, V.I. Strategy of cross-linked enzyme aggregates onto magnetic particles adapted to the green design of biocatalytic synthesis of glycerol carbonate. RSC Adv. 2013, 3, 4052–4058. [Google Scholar] [CrossRef]
- Zheng, D.; Wang, S.; Qiu, S.; Lin, J.; Diao, X. Synthesis of butyl oleate catalyzed by cross-linked enzyme aggregates with magnetic nanoparticles in rotating magneto-micro-reactor. J. Biotechnol. 2018, 281, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Loos, K. Enzymatic Synthesis of Biobased Polyesters and Polyamides. Polymers 2016, 8, 243. [Google Scholar] [CrossRef]
- Stavila, E.; Arsyi, R.Z.; Petrovic, D.M.; Loos, K. Fusarium solani pisi cutinase-catalyzed synthesis of polyamides. Eur. Polym. J. 2013, 49, 834–842. [Google Scholar] [CrossRef]
- Steunenberg, P.; Uiterweerd, M.; Sijm, M.; Scott, E.L.; Zuilhof, H.; Sanders, J.P.M.; Franssen, M.C.R. Enzyme-Catalyzed Polymerization of β-alanine Esters, A Sustainable Route Towards the Formation of Poly-β-alanine. Curr. Org. Chem. 2013, 17, 682–690. [Google Scholar] [CrossRef]
- Kumar, V.V.; Sivaesan, S.; Cabana, H. Magnetic cross-linked laccase aggregates—Bioremediation tool for decolorization of distinct classes of recalcitrant dyes. Sci. Total Environ. 2014, 487, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, Z.; Lin, Y.; Ng, T.B.; Ye, X.; Lin, J. Immobilized Cerrena sp. Laccase: Preparation, thermal inactivation and operational stability in malachite green decolorization. Sci. Rep. 2017, 7, 16429. [Google Scholar] [CrossRef]
- Zhou, L.; Tang, W.; Jiang, Y.; Ma, L.; He, Y.; Gao, J. Magnetic combined cross-linked enzyme aggregates of horseradish peroxidase and glucose oxidase: An efficient biocatalyst for dye decolourization. RSC Adv. 2016, 6, 90061–90068. [Google Scholar] [CrossRef]
- Kumar, V.V.; Cabana, H. Towards high potential magnetic biocatalysts for on-demand elimination of pharmaceuticals. Bioresour. Technol. 2016, 200, 81–89. [Google Scholar] [CrossRef]
- Bedade, D.K.; Muley, A.B.; Singhal, R.S. Magnetic cross-linked aggregates of acrylamidase from Cupriavidus oxalaticus ICTDB921 for biodegradation of acrylamide from industrial waste water. Bioresour. Technol. 2019, 272, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Grooten, H.M.J.; Rijkers, M.P.W.M.; Moody, H.M.; Webbers, J.J.P.; De Vroom, E. Method for the Separation of a Solid Substance. WO Patent 98/56486, 17 December 1998. [Google Scholar]
- Kopp, W.; Da Costa, T.P.; Pereira, S.C.; Jafelicci, M.; Giordano, R.C.; Marques, R.F.C.; Araujo-Moreira, F.M.; Giordano, R.L.C. Easily handling penicillin G acylase magnetic cross-linked enzymes aggregates: Catalytic and morphological studies. Process Biochem. 2014, 49, 38–46. [Google Scholar] [CrossRef]
- Ling, X.-M.; Wang, X.-Y.; Ma, P.; Yang, Y.; Qin, J.-M.; Zhang, X.-J.; Zhang, Y.-W. Covalent Immobilization of Penicillin G Acylase onto Fe3O4@Chitosan Magnetic Nanoparticles. J. Microbiol. Biotechnol. 2016, 26, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Illanes, A.; Soler, L.; Henriquez, M.J. Effect of the degree of cross-linking on the properties of different CLEAs of penicillin acylase. J. Process Biochem. 2009, 44, 322–326. [Google Scholar] [CrossRef]
- Su, E.; Meng, Y.; Ning, C.; Ma, X.; Deng, S. Magnetic combined cross-linked enzyme aggregates (Combi-CLEAs) for cofactor regeneration in the synthesis of chiral alcohol. J. Biotechnol. 2018, 271, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Jiang, Q.; Sun, L.; Li, Q.; Zhou, L.; Chen, Q.; Li, S.; Yu, M.; Li, W. Magnetic Combined Cross-Linked Enzyme Aggregates of Ketoreductase and Alcohol dehydrogenase: An Efficient and Stable Biocatalyst for Asymmetric Synthesis of (R)-3-Quinuclidinol with Regeneration of Coenzymes In Situ. Catalysts 2018, 8, 334. [Google Scholar] [CrossRef]
- Peirce, S.; Russo, M.E.; De Luca, V.; Capasso, C.; Rossi, M.; Olivieri, G.; Salatino, P.; Marzocchella, A. Immobilization of carbonic anhydrase for biomimetic CO2 capture in a slurry absorber as cross-linked enzyme aggregates (CLEA). Chem. Eng. Trans. 2015, 43, 259–264. [Google Scholar] [CrossRef]
- Gupta, A.; Srivastava, S.K. Study of cross linked enzyme aggregate of glucose isomerase of Streptomyces thermonitrificans immobilised on magnetic particle. J. Biochem. Technol. 2016, 7, 1102–1106. [Google Scholar]
Figure 1.
Cross-linked enzyme aggregate (CLEA) preparation.

Figure 2.
Hydrolysis of naringin and synthesis of salidroside with glycoside CLEAs.

Figure 3.
CLEA-catalyzed transglycosidations.

Figure 4.
Asymmetric synthesis of cyanohydrins with hydroxynitrile lyase (HNL)-CLEAs.

Figure 5.
N-acetyl-d-neuraminic acid synthesis using a L. planatarum N-acetyl-d-neuraminic acid aldolase (NAL)-CLEA.
Figure 5.
N-acetyl-d-neuraminic acid synthesis using a L. planatarum N-acetyl-d-neuraminic acid aldolase (NAL)-CLEA.

Figure 6.
Phenylalanine ammonia lyase (PAL)-CLEA catalyzed synthesis of l-phenylalanine.

Figure 7.
Enoate reductase/glucose dehydrogenase (ERED/GDH)-combi-CLEA.

Figure 8.
HNL-Nlase-Amidase combi-CLEAand HNL-NHase combi-CLEA.

Figure 9.
One-pot conversion of sucrose to trehalose with a combi-CLEA.

Figure 10.
Wine aroma enhancement catalyzed by a combi-CLEA.

Figure 11.
One-pot starch hydrolysis with a combi-CLEA.

Figure 12.
Preparation of magnetic CLEAs.

Figure 13.
Lipase-m-CLEA catalyzed epoxidation and transesterification.

Figure 14.
Enzymatic synthesis of penicillins and cephalosporins.

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sheldon, R.A. CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts 2019, 9, 261. https://0-doi-org.brum.beds.ac.uk/10.3390/catal9030261
AMA Style
Sheldon RA. CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts. 2019; 9(3):261. https://0-doi-org.brum.beds.ac.uk/10.3390/catal9030261
Chicago/Turabian StyleSheldon, Roger A. 2019. "CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy" Catalysts 9, no. 3: 261. https://0-doi-org.brum.beds.ac.uk/10.3390/catal9030261
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.