
Analysis of Programmed Cell Death and Senescence Markers in the Developing Retina of an Altricial Bird Species

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
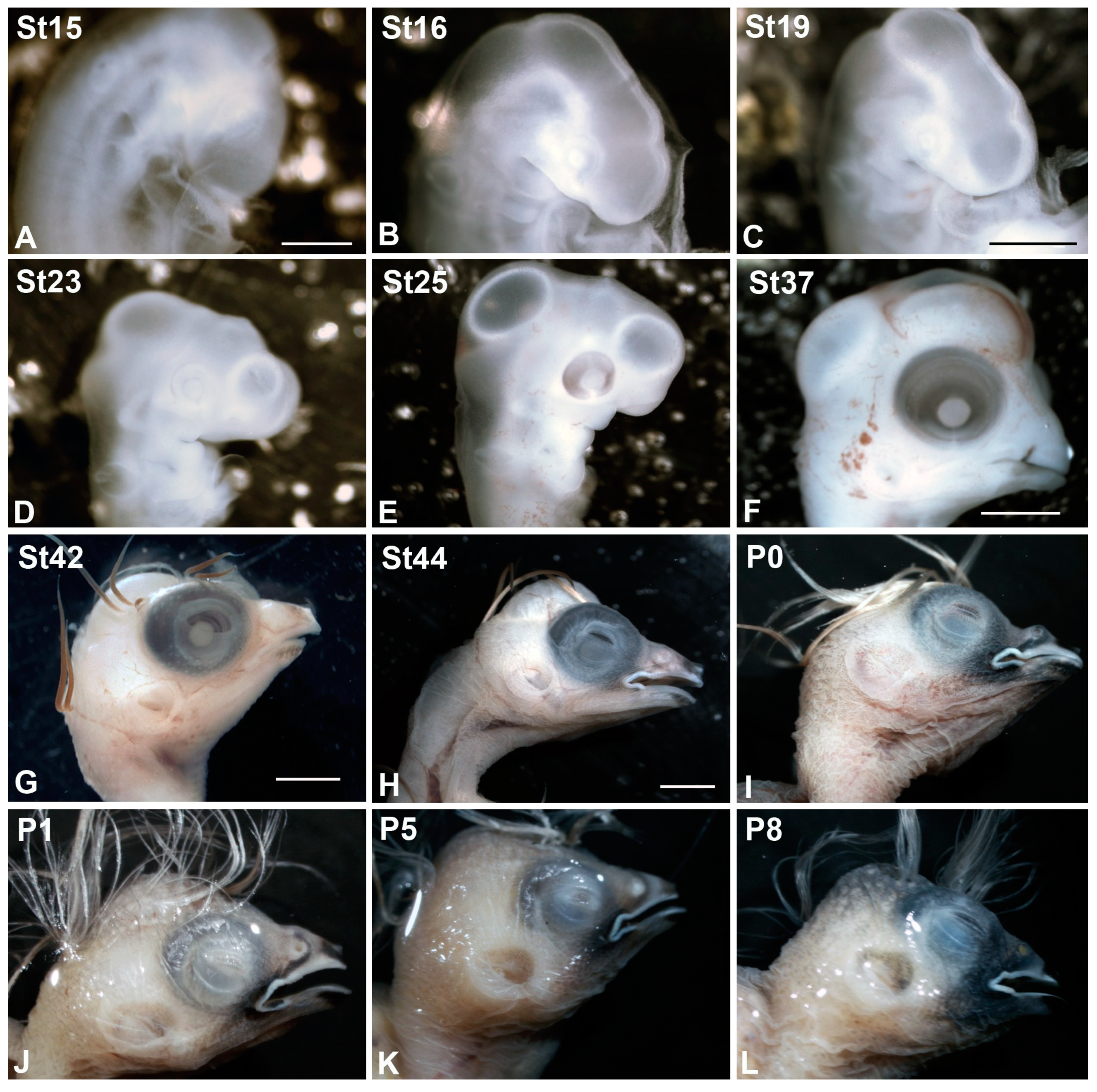
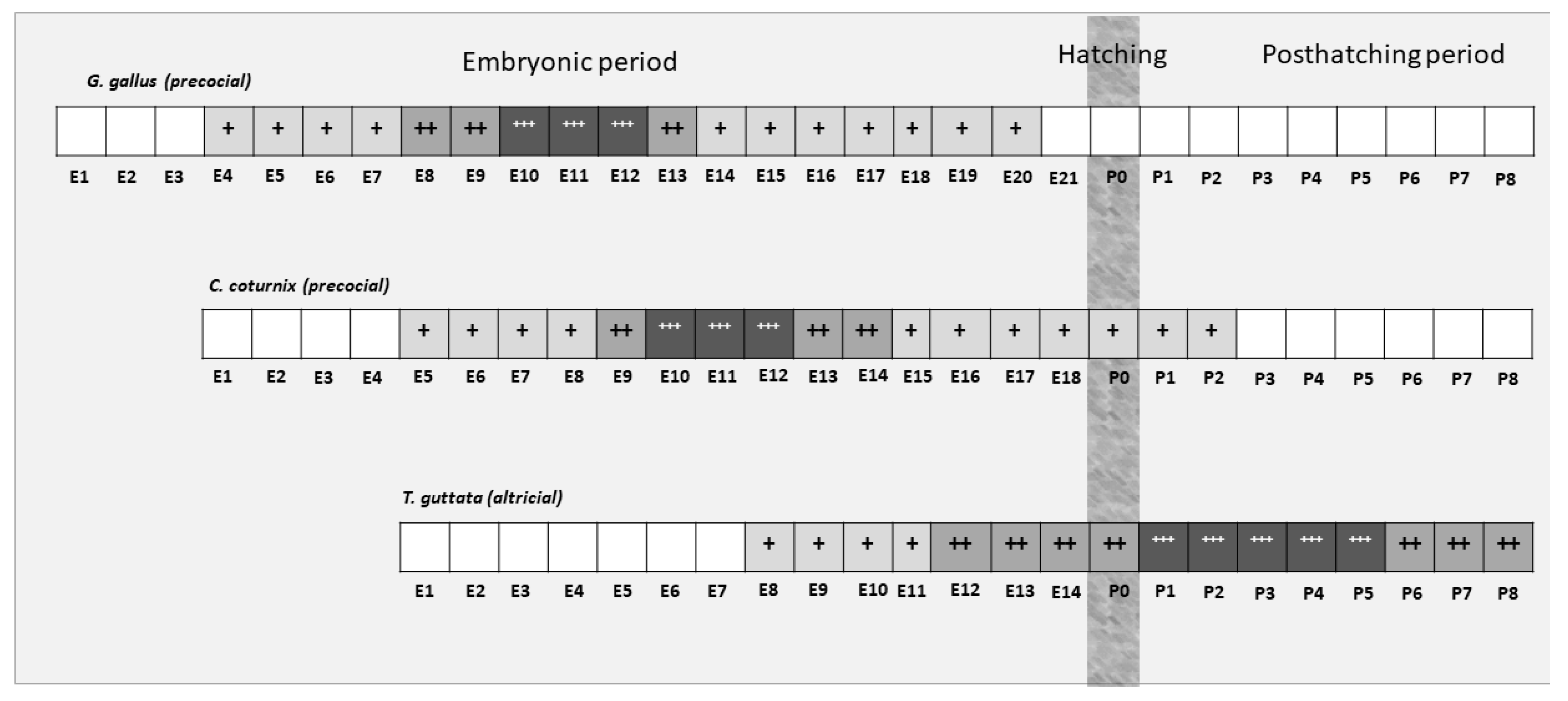
2.1. Animal and Tissue Processing
2.2. Toluidine Blue Staining
2.3. Detection of β-Galactosidase Activity
2.4. Immunohistochemistry
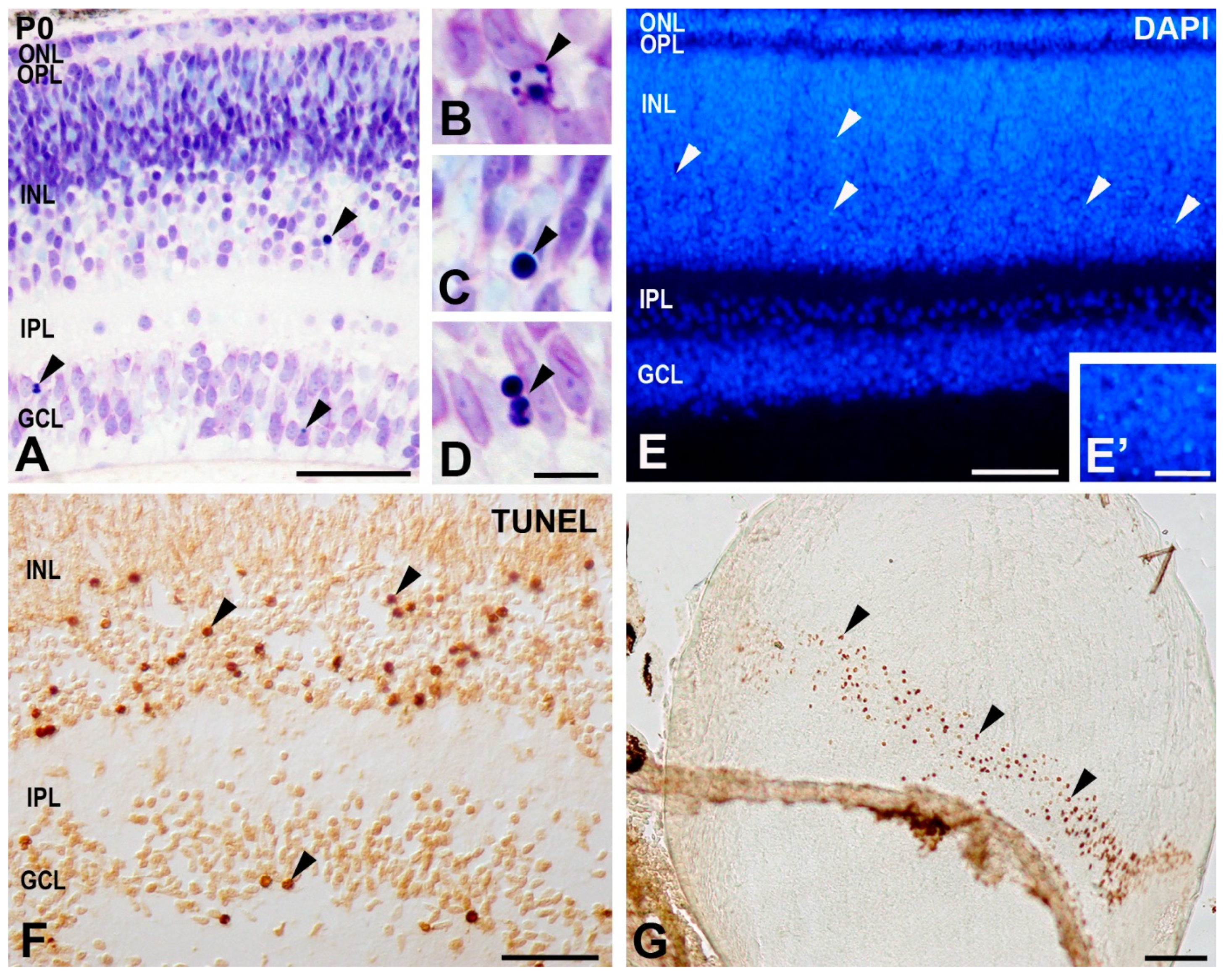
2.5. TUNEL Technique
2.6. Quantification of TUNEL-Positive Nuclei
2.7. Image Acquisition and Processing
3. Results
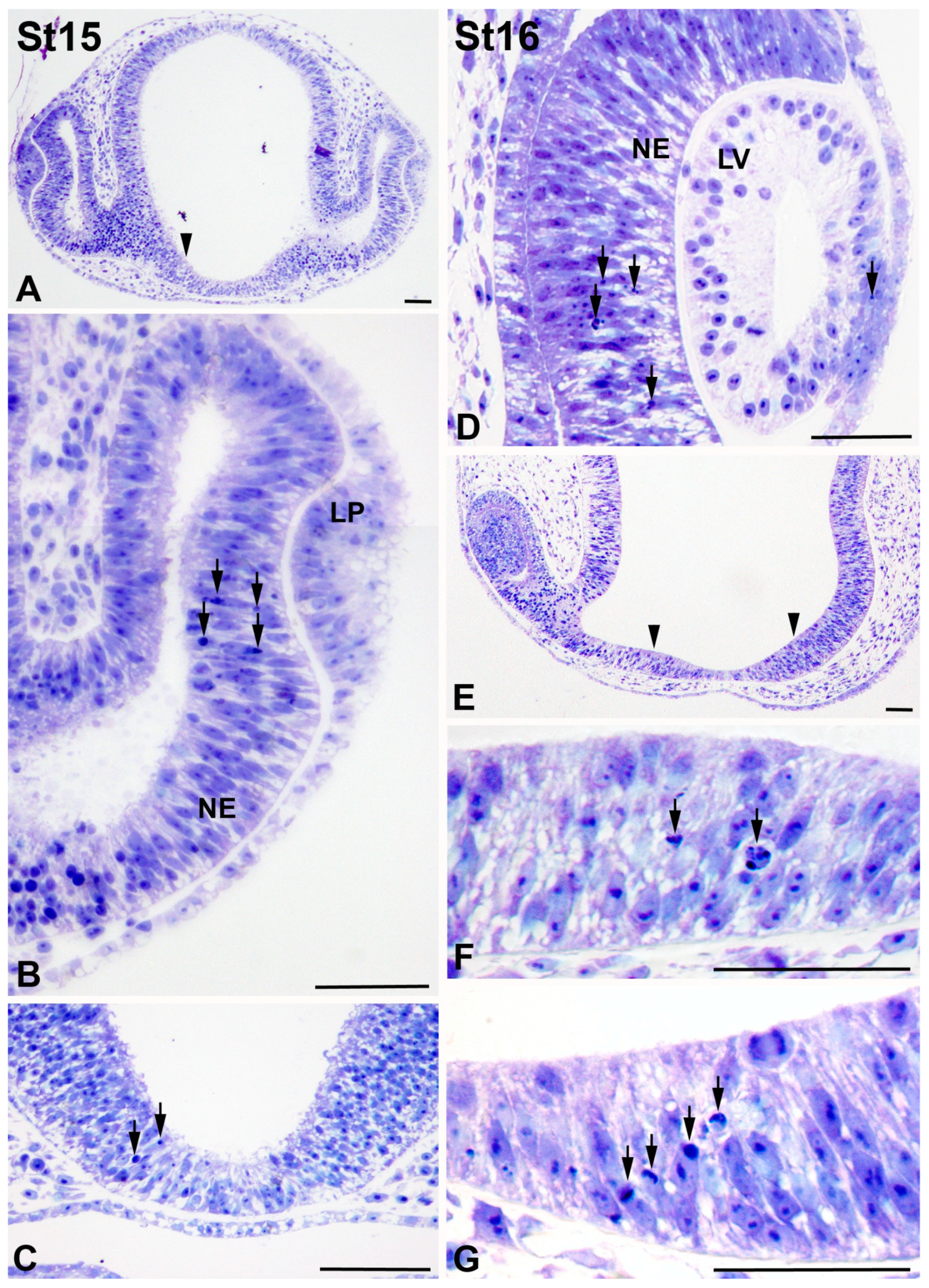
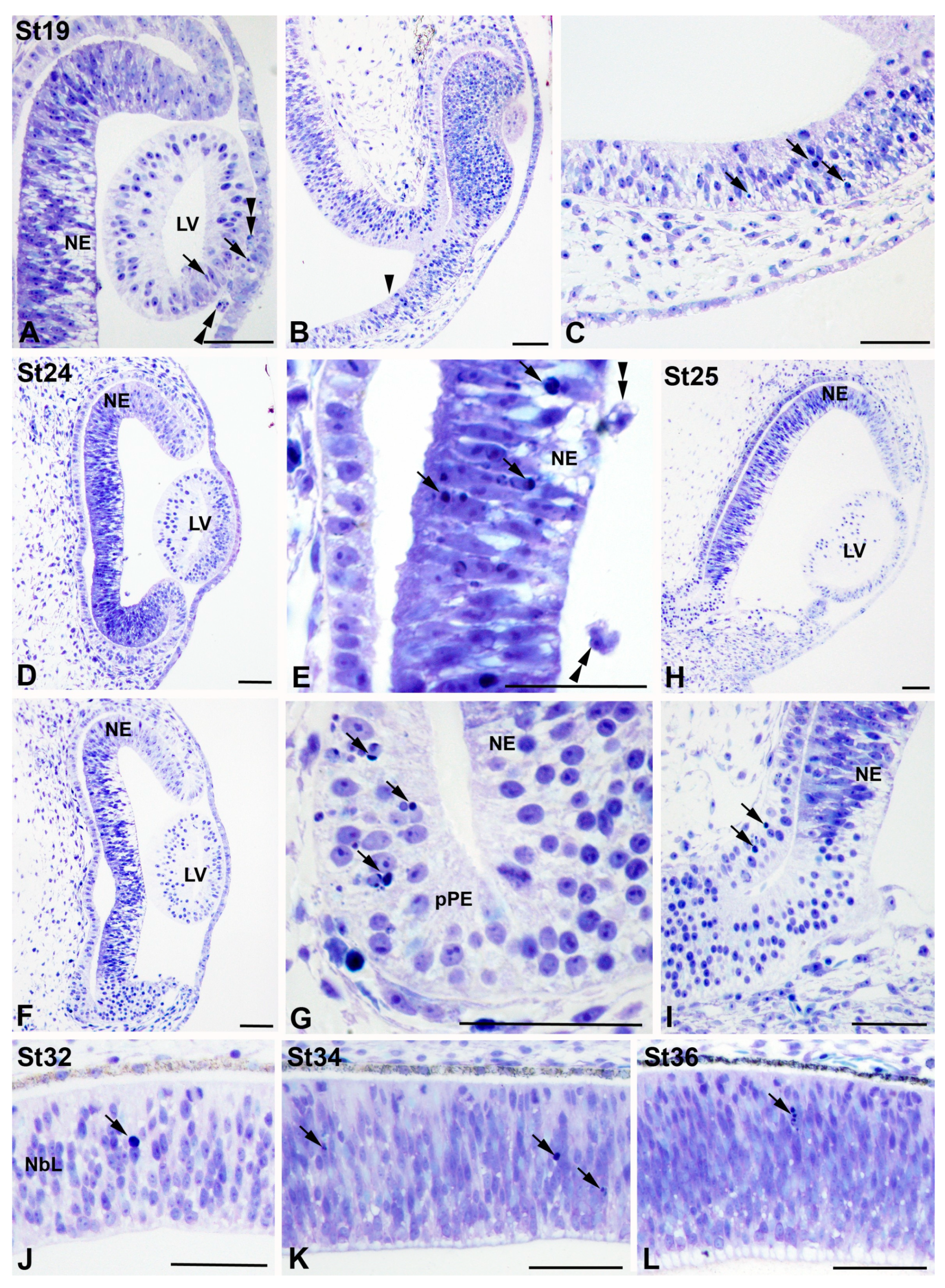
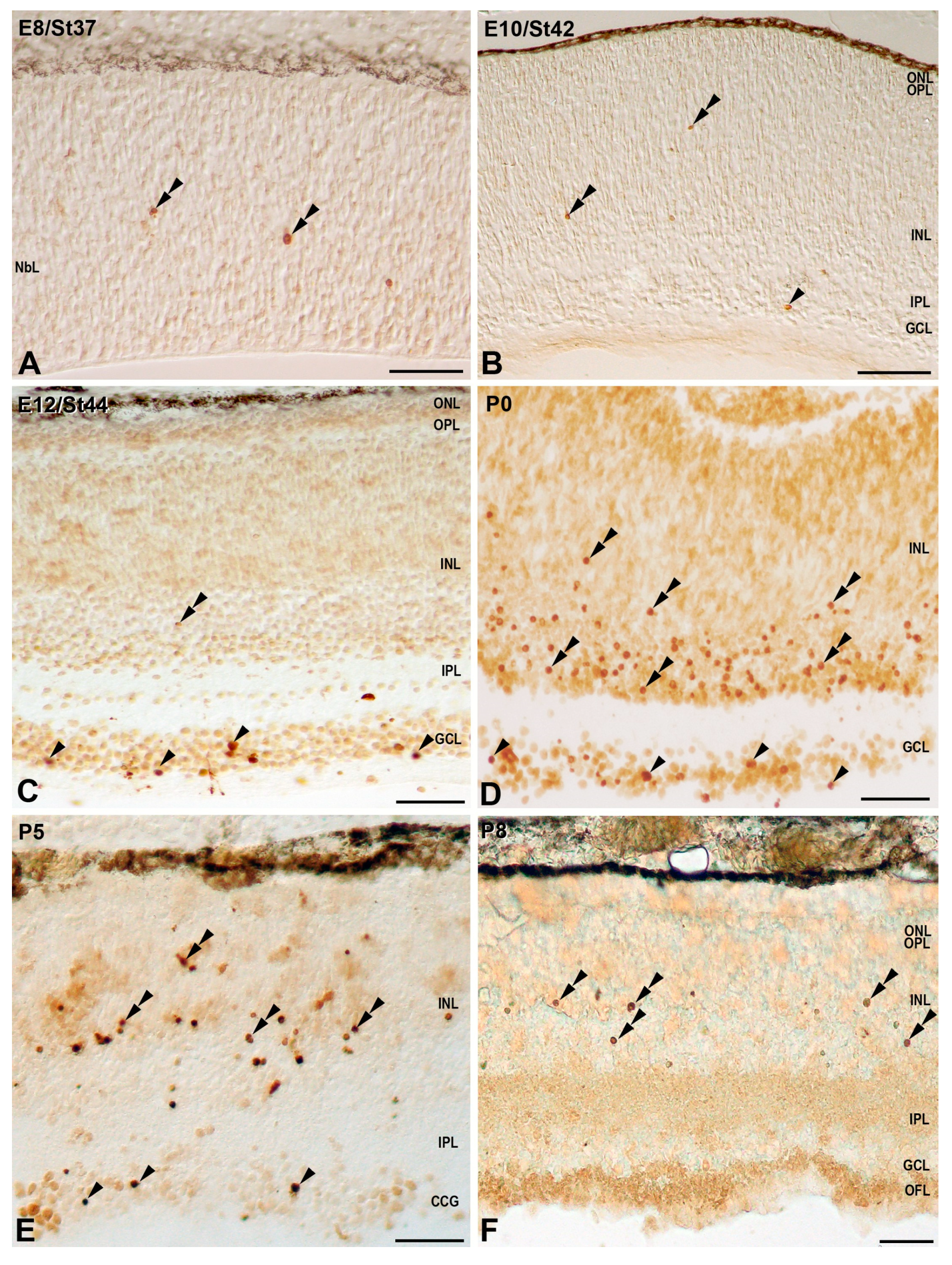
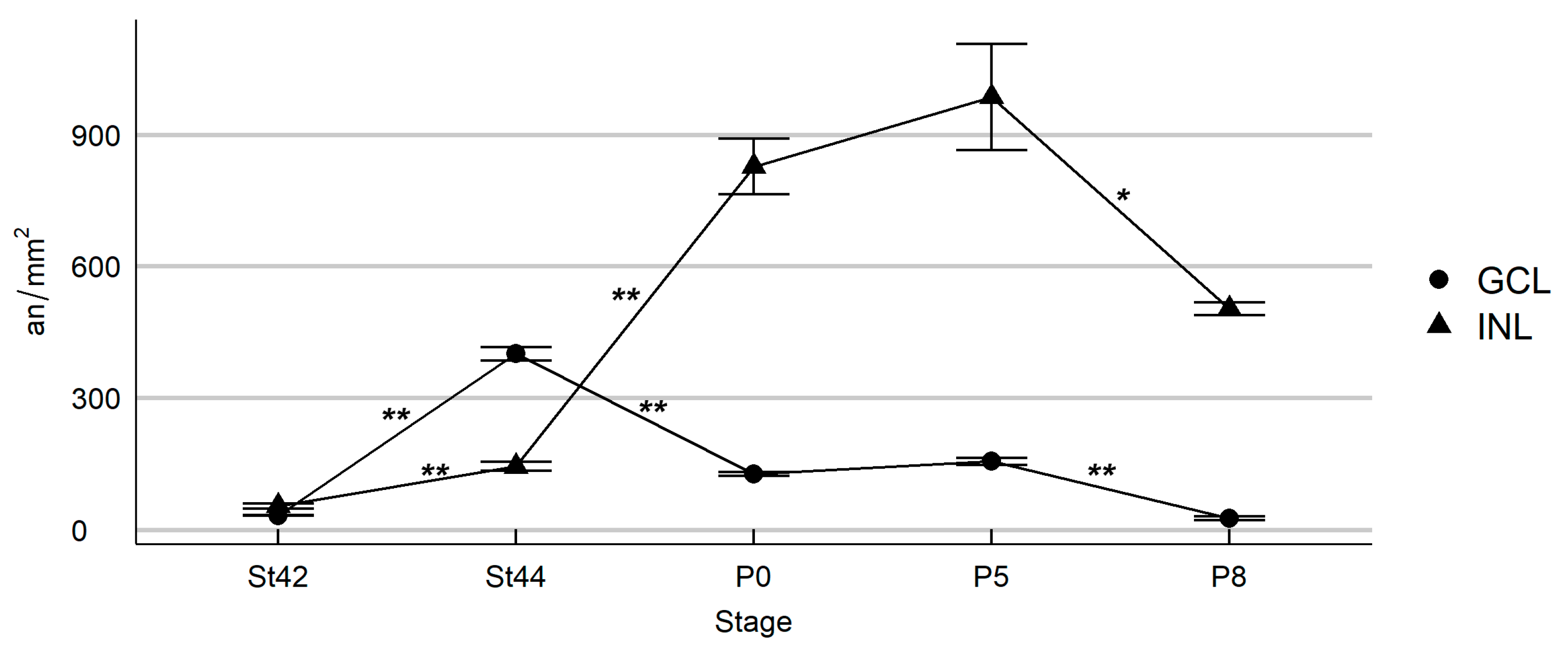
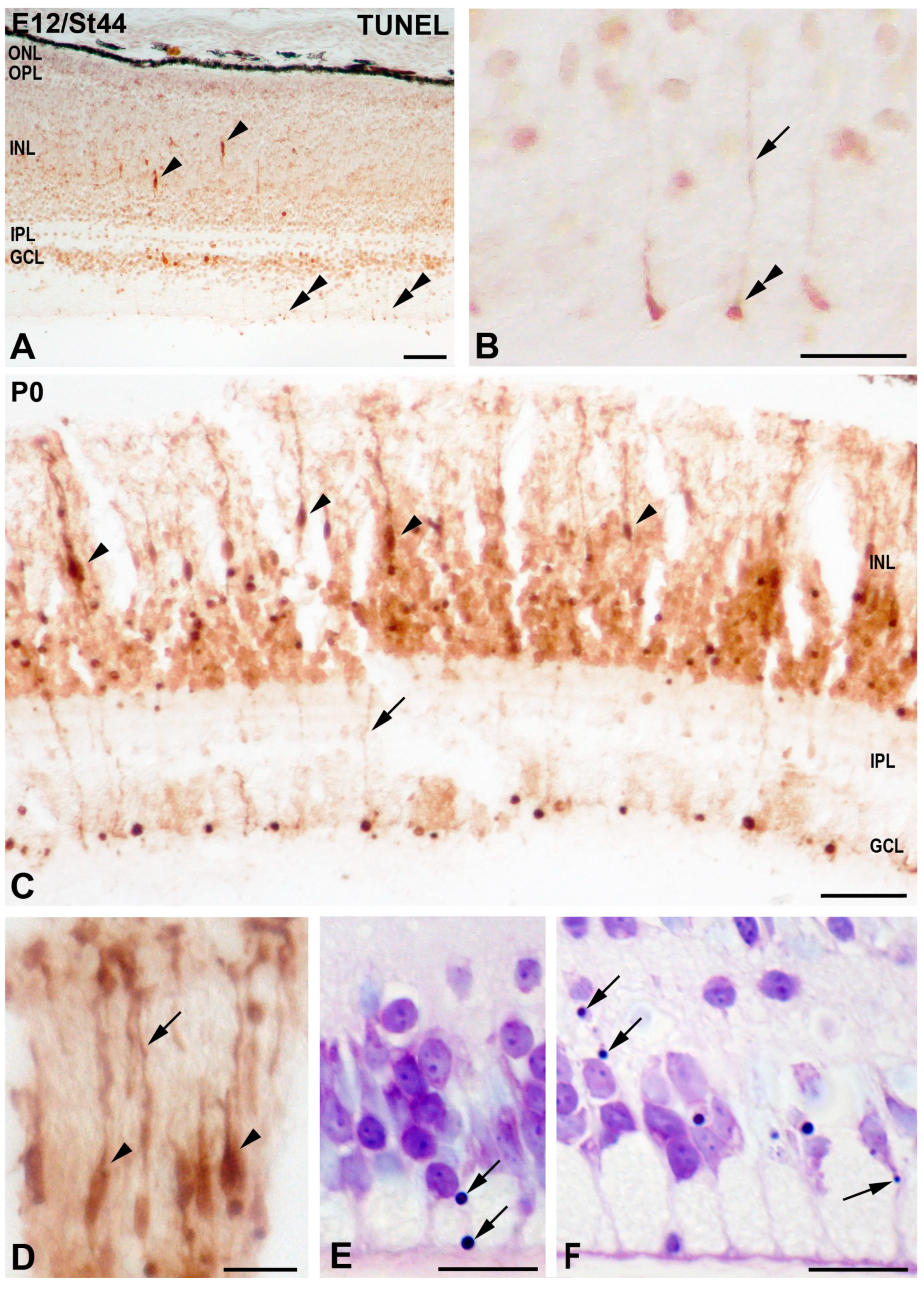
3.1. Programmed Cell Death in the Developing T. guttata Visual System
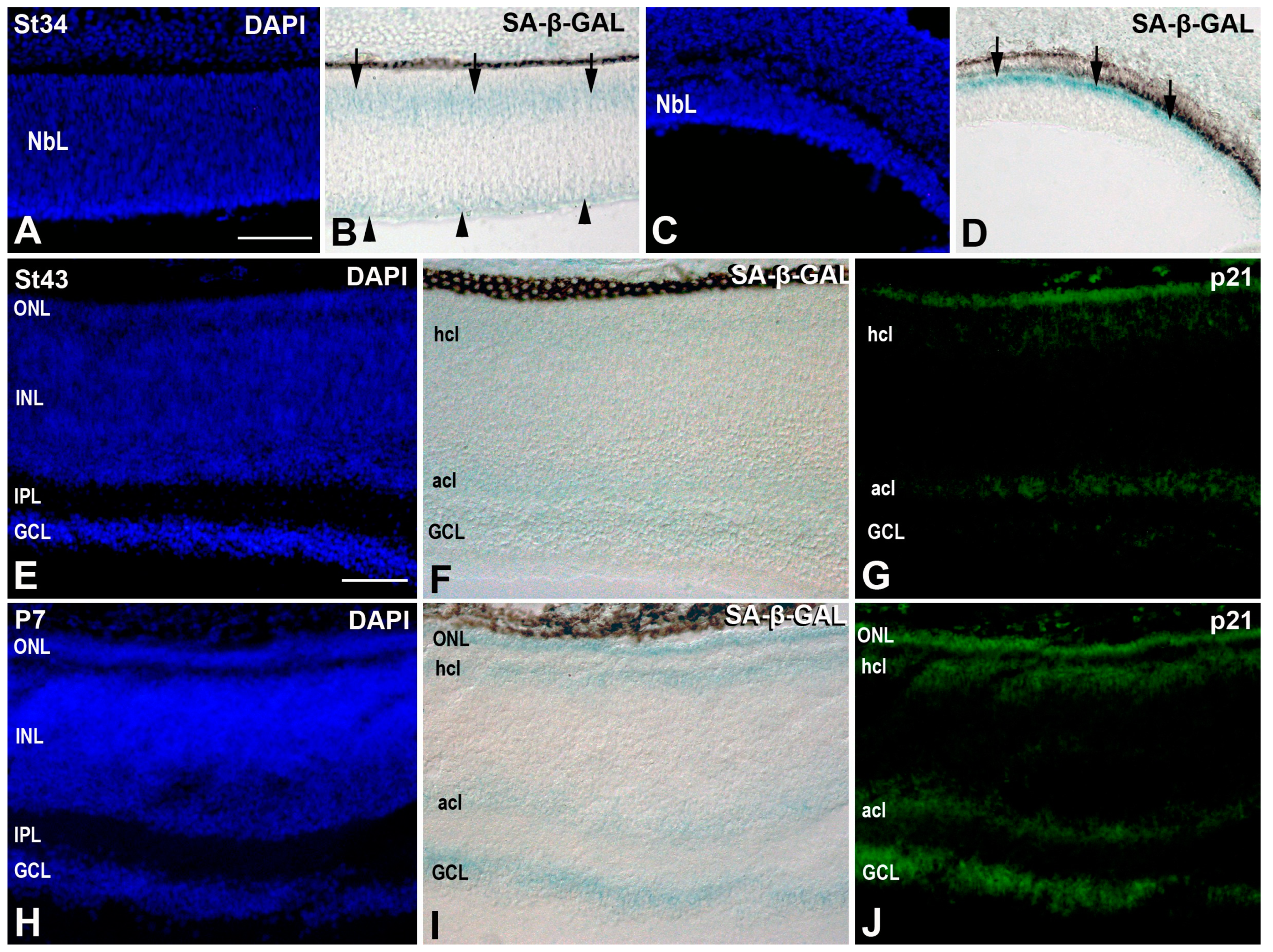
3.2. Senescence Markers in the Developing T. guttata Visual System
4. Discussion
4.1. Cell Death during Early Visual System Morphogenesis in T. guttata
4.2. Cell Death during the Period of Cell Differentiation in the T. guttata Retina
4.3. TUNEL Labeling in the Cytoplasm of Radially Oriented Cells
4.4. Senescence Markers in the Developing Retina of T. guttata
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muñoz-Espín, D.; Cañamero, M.; Maraver, A.; Gómez-López, G.; Contreras, J.; Murillo-Cuesta, S.; Rodríguez-Baeza, A.; Varela-Nieto, I.; Ruberte, J.; Collado, M.; et al. Programmed Cell Senescence during Mammalian Embryonic Development. Cell 2013, 155, 1104–1118. [Google Scholar] [CrossRef] [Green Version]
- Storer, M.; Mas, A.; Robert-Moreno, A.; Pecoraro, M.; Ortells, M.C.; Di Giacomo, V.; Yosef, R.; Pilpel, N.; Krizhanovsky, V.; Sharpe, J.; et al. Senescence Is a Developmental Mechanism that Contributes to Embryonic Growth and Patterning. Cell 2013, 155, 1119–1130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco-Morcillo, J.; Bejarano-Escobar, R.; Rodríguez-León, J.; Navascués, J.; Martín-Partido, G. Ontogenetic Cell Death and Phagocytosis in the Visual System of Vertebrates: Cell Death and Phagocytosis during Ontogeny. Dev. Dyn. 2014, 243, 1203–1225. [Google Scholar] [CrossRef]
- Lorda-Diez, C.I.; Montero, J.A.; Garcia-Porrero, J.A.; Hurle, J.M. Interdigital tissue regression in the developing limb of vertebrates. Int. J. Dev. Biol. 2015, 59, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Montero, J.A.; Sanchez-Fernandez, C.; Diez, C.I.L.; Garcia-Porrero, J.A.; Hurle, J.M. DNA damage precedes apoptosis during the regression of the interdigital tissue in vertebrate embryos. Sci. Rep. 2016, 6, 35478. [Google Scholar] [CrossRef] [PubMed]
- Montero, J.A.; Lorda-Diez, C.I.; Hurle, J.M. Confluence of Cellular Degradation Pathways During Interdigital Tissue Remodeling in Embryonic Tetrapods. Front. Cell Dev. Biol. 2020, 8, 593761. [Google Scholar] [CrossRef]
- Lorda-Diez, C.I.; Solis-Mancilla, M.E.; Sanchez-Fernandez, C.; Garcia-Porrero, J.A.; Hurle, J.M.; Montero, J.A. Cell senescence, apoptosis and DNA damage cooperate in the remodeling processes accounting for heart morphogenesis. J. Anat. 2019, 234, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Nacher, V.; Carretero, A.; Navarro, M.; Armengol, C.; Llombart, C.; Rodríguez, A.; Herrero-Fresneda, I.; Ayuso, E.; Ruberte, J. The Quail Mesonephros: A New Model for Renal Senescence? J. Vasc. Res. 2006, 43, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Davaapil, H.; Brockes, J.P.; Yun, M.H. Conserved and novel functions of programmed cellular senescence during vertebrate development. Development 2016, 144, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Villiard, É.; Denis, J.F.; Hashemi, F.S.; Igelmann, S.; Ferbeyre, G.; Roy, S. Senescence gives insights into the morphogenetic evolution of anamniotes. Biol. Open 2017, 6, 891–896. [Google Scholar] [CrossRef] [Green Version]
- Da Silva-Álvarez, S.; Lamas-González, O.; Ferreirós, A.; González, P.; Gómez, M.; García-Caballero, T.; Barcia, M.G.; García-González, M.A.; Collado, M. Pkd2 deletion during embryo development does not alter mesonephric programmed cell senescence. Int. J. Dev. Biol. 2018, 62, 637–640. [Google Scholar] [CrossRef]
- Gibaja, A.; Aburto, M.R.; Pulido, S.; Collado, M.; Hurle, J.M.; Varela-Nieto, I.; Magariños, M. TGFβ2-induced senescence during early inner ear development. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Varela-Nieto, I.; Palmero, I.; Magariños, M. Complementary and distinct roles of autophagy, apoptosis and senescence during early inner ear development. Hear. Res. 2019, 376, 86–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magariños, M.; Barajas-Azpeleta, R.; Varela-Nieto, I.; Aburto, M.R. Otic Neurogenesis Is Regulated by TGFβ in a Senescence-Independent Manner. Front. Cell. Neurosci. 2020, 14, 217. [Google Scholar] [CrossRef]
- Biehlmaier, O.; Neuhauss, S.C.; Kohler, K. Onset and time course of apoptosis in the developing zebrafish retina. Cell Tissue Res. 2001, 306, 199–207. [Google Scholar] [CrossRef]
- Candal, E.; Anadón, R.; DeGrip, W.J.; Rodríguez-Moldes, I. Patterns of cell proliferation and cell death in the developing retina and optic tectum of the brown trout. Brain Res. Dev. Brain Res. 2005, 154, 101–119. [Google Scholar] [CrossRef]
- Iijima, N.; Yokoyama, T. Apoptosis in the Medaka Embryo in the Early Developmental Stage. Acta Histochem. Cytochem. 2007, 40, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejarano-Escobar, R.; Blasco, M.; DeGrip, W.J.; Oyola-Velasco, J.A.; Martín-Partido, G.; Francisco-Morcillo, J. Eye development and retinal differentiation in an altricial fish species, the senegalese sole (Solea senegalensis, Kaup 1858). J. Exp. Zoöl. Part B Mol. Dev. Evol. 2010, 314, 580–605. [Google Scholar] [CrossRef] [PubMed]
- Bejarano-Escobar, R.; Blasco, M.; Durán, A.C.; Martín-Partido, G.; Francisco-Morcillo, J. Chronotopographical distribution patterns of cell death and of lectin-positive macrophages/microglial cells during the visual system ontogeny of the small-spotted catshark Scyliorhinus canicula. J. Anat. 2013, 223, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Gaze, R.M.; Grant, P. Spatio-temporal patterns of retinal ganglion cell death duringXenopus development. J. Comp. Neurol. 1992, 315, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Hensey, C.; Gautier, J. Programmed Cell Death duringXenopusDevelopment: A Spatio-temporal Analysis. Dev. Biol. 1998, 203, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Hutson, L.D.; Bothwell, M. Expression and function of Xenopus laevis p75(NTR) suggest evolution of developmental regulatory mechanisms. J. Neurobiol. 2001, 49, 79–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francisco-Morcillo, J.; Hidalgo-Sánchez, M.; Martín-Partido, G. Spatial and temporal patterns of apoptosis during differentiation of the retina in the turtle. Anat. Embryol. 2004, 208, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo-Sánchez, M.; Francisco-Morcillo, J.; Navascués, J.; Martín-Partido, G. Developmental changes in the fibre population of the optic nerve follow an avian/mammalian-like pattern in the turtle Mauremys leprosa. Brain Res. 2006, 1113, 74–85. [Google Scholar] [CrossRef]
- Hidalgo-Sánchez, M.; Francisco-Morcillo, J.; Martín-Partido, G. Changes in fiber arrangement in the retinofugal pathway of the turtle Mauremys leprosa: An evolutionarily conserved mechanism. Brain Res. 2007, 1186, 124–128. [Google Scholar] [CrossRef]
- Young, R.W. Cell death during differentiation of the retina in the mouse. J. Comp. Neurol. 1984, 229, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Knabe, W.; Kuhn, H.J. Pattern of cell death during optic cup formation in the tree shrew Tupaia belangeri. J. Comp. Neurol. 1998, 401, 352–366. [Google Scholar] [CrossRef]
- Knabe, W.; Süss, M.; Kuhn, H.-J. The patterns of cell death and of macrophages in the developing forebrain of the tree shrew Tupaia belangeri. Anat. Embryol. 2000, 201, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Péquignot, M.; Provost, A.C.; Sallé, S.; Taupin, P.; Sainton, K.M.; Marchant, D.; Martinou, J.C.; Ameisen, J.C.; Jais, J.-P.; Abitbol, M. Major role of BAX in apoptosis during retinal development and in establishment of a functional postnatal retina. Dev. Dyn. 2003, 228, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Gallardo, L.; Lineros-Domínguez, M.D.C.; Francisco-Morcillo, J.; Martín-Partido, G. Macrophages during retina and optic nerve development in the mouse embryo: Relationship to cell death and optic fibres. Anat. Embryol. 2005, 210, 303–316. [Google Scholar] [CrossRef]
- Bejarano-Escobar, R.; Holguín-Arévalo, M.S.; Montero, J.A.; Francisco-Morcillo, J.; Martín-Partido, G. Macrophage and microglia ontogeny in the mouse visual system can be traced by the expression of Cathepsins B and D. Dev. Dyn. 2011, 240, 1841–1855. [Google Scholar] [CrossRef] [PubMed]
- Cuadros, M.A.; Ríos, A. Spatial and temporal correlation between early nerve fiber growth and neuroepithelial cell death in the chick embryo retina. Anat. Embryol. 1988, 178, 543–551. [Google Scholar] [CrossRef]
- Martín-Partido, G.; Rodríguez-Gallaro, L.; Alvarez, I.S.; Navascués, J. Cell death in the ventral region of the neural retina during the early development of the chick embryo eye. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1988, 222, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Cook, B.; Portera-Cailliau, C.; Adler, R. Developmental neuronal death is not a universal phenomenon among cell types in the chick embryo retina. J. Comp. Neurol. 1998, 396, 12–19. [Google Scholar] [CrossRef]
- Trousse, F.; Esteve, P.; Bovolenta, P. BMP4 Mediates Apoptotic Cell Death in the Developing Chick Eye. J. Neurosci. 2001, 21, 1292–1301. [Google Scholar] [CrossRef]
- Mayordomo, R.; Valenciano, A.I.; De La Rosa, E.J.; Hallböök, F. Generation of retinal ganglion cells is modulated by caspase-dependent programmed cell death. Eur. J. Neurosci. 2003, 18, 1744–1750. [Google Scholar] [CrossRef] [Green Version]
- Chavarría, T.; Valenciano, A.I.; Mayordomo, R.; Egea, J.; Comella, J.X.; Hallböök, F.; De Pablo, F.; De La Rosa, E.J. Differential, age-dependent MEK-ERK and PI3K-Akt activation by insulin acting as a survival factor during embryonic retinal development. Dev. Neurobiol. 2007, 67, 1777–1788. [Google Scholar] [CrossRef]
- Chavarría, T.; Baleriola, J.; Mayordomo, R.; De Pablo, F.; De La Rosa, E.J. Early Neural Cell Death Is an Extensive, Dynamic Process in the Embryonic Chick and Mouse Retina. Sci. World J. 2013, 2013, 627240. [Google Scholar] [CrossRef] [Green Version]
- Marín-Teva, J.L.; Cuadros, M.A.; Calvente, R.; Almendros, A.; Navascués, J. Naturally Occurring Cell Death and Migration of Microglial Precursors in the Quail Retina during Normal Development. J. Comp. Neurol. 1999, 412, 255–275. [Google Scholar] [CrossRef]
- Navascuéeas, J.; Martíian-Partido, G.; Alvarez, I.S.; Rodríiaguez-Gallardo, L. Cell death in suboptic necrotic centers of chick embryo diencephalon and their topographic relationship with the earliest optic fiber fascicles. J. Comp. Neurol. 1988, 278, 34–46. [Google Scholar] [CrossRef]
- Navascués, J.; Martín-Partido, G. Glial cells in the optic chiasm arise from the suboptic necrotic centers of the diencephalon floor: Morphological evidence in the chick embryo. Neurosci. Lett. 1990, 120, 62–65. [Google Scholar] [CrossRef]
- Dimri, G.P.; Lee, X.; Basile, G.; Acosta, M.; Scott, G.; Roskelley, C.; Medrano, E.E.; Linskens, M.; Rubelj, I.; Pereira-Smith, O.; et al. A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc. Natl. Acad. Sci. USA 1995, 92, 9363–9367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mera-Rodríguez, J.A.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J.; Mera-Rodríguez, J.A. Senescence-associated β-galactosidase activity in the developing avian retina. Dev. Dyn. 2019, 248, 850–865. [Google Scholar] [CrossRef]
- De Mera-Rodríguez, J.A.; Álvarez-Hernán, G.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Is Senescence-Associated β-Galactosidase a Reliable in vivo Marker of Cellular Senescence During Embryonic Development? Front. Cell Dev. Biol. 2021, 9, 623175. [Google Scholar] [CrossRef] [PubMed]
- Francisco-Morcillo, J.; Sánchez-Calderón, H.; Kawakami, Y.; Belmonte, J.C.I.; Hidalgo-Sánchez, M.; Martín-Partido, G. Expression of Fgf19 in the developing chick eye. Dev. Brain Res. 2005, 156, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Calaza, K.D.C.; Gardino, P.F. Neurochemical phenotype and birthdating of specific cell populations in the chick retina. Anais Acad. Bras. Ciências 2010, 82, 595–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, L.M.; Mitchell, M.A.; Ramírez, Y.M.; McNeil, R. Comparative Analysis of Retina Structure and Photopic Electro-retinograms in Developing Altricial Pigeons (Columba Livia) and Precocial Japanese Quails (Coturnix Coturnix Japonica). Neotrop. Ornitholog. Soc. 2007, 18, 503–518. [Google Scholar]
- Álvarez-Hernán, G.; Sánchez-Resino, E.; Hernández-Núñez, I.; Marzal, A.; Rodriguez-Leon, J.; Martín-Partido, G.; Francisco-Morcillo, J. Retinal histogenesis in an altricial avian species, the zebra finch (Taeniopygia guttata, Vieillot 1817). J. Anat. 2018, 233, 106–120. [Google Scholar] [CrossRef] [PubMed]
- Francisco-Morcillo, J.; Alvarez-Hernan, G.; De Mera-Rodríguez, J.A.; Gañán, Y.; Solana-Fajardo, J.; Martín-Partido, G.; Rodríguez-León, J. Development and postnatal neurogenesis in the retina: A comparison between altricial and precocial bird species. Neural Regen. Res. 2021, 16, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Hernán, G.; Hernández-Núñez, I.; Rico-Leo, E.M.; Marzal, A.; De Mera-Rodríguez, J.A.; Rodríguez-León, J.; Martín-Partido, G.; Francisco-Morcillo, J. Retinal differentiation in an altricial bird species, Taeniopygia guttata: An immunohistochemical study. Exp. Eye Res. 2020, 190, 107869. [Google Scholar] [CrossRef]
- Murray, J.R.; Varian-Ramos, C.W.; Welch, Z.S.; Saha, M.S. Embryological staging of the Zebra Finch, Taeniopygia guttata. J. Morphol. 2013, 274, 1090–1110. [Google Scholar] [CrossRef] [Green Version]
- Piechota, M.; Sunderland, P.; Wysocka, A.; Nalberczak, M.; Sliwinska, M.A.; Radwanska, K.; Sikora, E. Is senescence-associated β-galactosidase a marker of neuronal senescence? Oncotarget 2016, 7, 81099–81109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavrieli, Y.; Sherman, Y.; Ben-Sasson, S.A. Identification of programmed cell death in situ via specific labeling of nuclear DNA fragmentation. J. Cell Biol. 1992, 119, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Sanwal, M.; Muel, A.; Chaudun, E.; Courtois, Y.; Counis, M. Chromatin condensation and terminal differentiation process in embryonic chicken lens in vivo and in vitro. Exp. Cell Res. 1986, 167, 429–439. [Google Scholar] [CrossRef]
- Sengelaub, D.R.; Dolan, R.P.; Finlay, B.L. Cell generation, death, and retinal growth in the development of the hamster retinal ganglion cell layer. J. Comp. Neurol. 1986, 246, 527–543. [Google Scholar] [CrossRef] [PubMed]
- Cuadros, M.A.; Martin, C.; Ríos, A.; Martín-Partido, G.; Navascués, J. Macrophages of hemangioblastic lineage invade the lens vesicle-ectoderm interspace during closure and detachment of the avian embryonic lens. Cell Tissue Res. 1991, 266, 117–127. [Google Scholar] [CrossRef]
- Nishitani, K.; Sasaki, K. Macrophage localization in the developing lens primordium of the mouse embryo—An immunohistochemical study. Exp. Eye Res. 2006, 83, 223–228. [Google Scholar] [CrossRef]
- Valenciano, A.I.; Boya, P.; De La Rosa, E.J. Early neural cell death: Numbers and cues from the developing neuroretina. Int. J. Dev. Biol. 2009, 53, 1515–1528. [Google Scholar] [CrossRef] [Green Version]
- Caspers, B.A.; Hagelin, J.C.; Paul, M.; Bock, S.; Willeke, S.; Krause, E.T. Zebra Finch chicks recognise parental scent, and retain chemosensory knowledge of their genetic mother, even after egg cross-fostering. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Egensperger, R.; Maslim, J.; Bisti, S.; Holländer, H.; Stone, J. Fate of DNA from retinal cells dying during development: Uptake by microglia and macroglia (Müller cells). Dev. Brain Res. 1996, 97, 1–8. [Google Scholar] [CrossRef]
- Thummel, R.; Kassen, S.C.; Enright, J.M.; Nelson, C.M.; Montgomery, J.E.; Hyde, D.R. Characterization of Müller glia and neuronal progenitors during adult zebrafish retinal regeneration. Exp. Eye Res. 2008, 87, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.J.; Fossum, S.L.; Fimbel, S.M.; Montgomery, J.E.; Hyde, D.R. The inhibitor of phagocytosis, O-phospho-l-serine, suppresses Müller glia proliferation and cone cell regeneration in the light-damaged zebrafish retina. Exp. Eye Res. 2010, 91, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Bejarano-Escobar, R.; Blasco, M.; Martín-Partido, G.; Francisco-Morcillo, J. Light-induced degeneration and microglial response in the retina of an epibenthonic pigmented teleost: Age-dependent photoreceptor susceptibility to cell death. J. Exp. Biol. 2012, 215, 3799–3812. [Google Scholar] [CrossRef] [Green Version]
- Sakami, S.; Imanishi, Y.; Palczewski, K. Müller glia phagocytose dead photoreceptor cells in a mouse model of retinal degenerative disease. FASEB J. 2018, 33, 3680–3692. [Google Scholar] [CrossRef]
- Nomura-Komoike, K.; Saitoh, F.; Fujieda, H. Phosphatidylserine recognition and Rac1 activation are required for Müller glia proliferation, gliosis and phagocytosis after retinal injury. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bejarano-Escobar, R.; Sánchez-Calderón, H.; Otero-Arenas, J.; Martín-Partido, G.; Francisco-Morcillo, J. Müller glia and phagocytosis of cell debris in retinal tissue. J. Anat. 2017, 231, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Charlton-Perkins, M.; Almeida, A.D.; Macdonald, R.B.; Harris, W.A. Genetic control of cellular morphogenesis in Müller glia. Glia 2019, 67, 1401–1411. [Google Scholar] [CrossRef] [Green Version]
- Vasey, D.B.; Wolf, C.R.; Brown, K.; Whitelaw, C.B.A. Spatial p21 expression profile in the mid-term mouse embryo. Transgenic Res. 2011, 20, 23–28. [Google Scholar] [CrossRef]
- Dewulf, V.; Bottjer, S.W. Age and Sex Differences in Mitotic Activity within the Zebra Finch Telencephalon. J. Neurosci. 2002, 22, 4080–4094. [Google Scholar] [CrossRef]
- Dewulf, V.; Bottjer, S.W. Neurogenesis within the juvenile zebra finch telencephalic ventricular zone: A map of proliferative activity. J. Comp. Neurol. 2004, 481, 70–83. [Google Scholar] [CrossRef]
- Striedter, G.F.; Charvet, C.J. Developmental origins of species differences in telencephalon and tectum size: Morphometric comparisons between a parakeet (Melopsittacus undulatus) and a quail (Colinus virgianus). J. Comp. Neurol. 2008, 507, 1663–1675. [Google Scholar] [CrossRef]
- Striedter, G.F.; Charvet, C.J. Telencephalon enlargement by the convergent evolution of expanded subventricular zones. Biol. Lett. 2008, 5, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Charvet, C.J.; Striedter, G.F. Causes and consequences of expanded subventricular zones. Eur. J. Neurosci. 2011, 34, 988–993. [Google Scholar] [CrossRef]
- Sugiyama, T.; Yamamoto, H.; Kon, T.; Chaya, T.; Omori, Y.; Suzuki, Y.; Abe, K.; Watanabe, D.; Furukawa, T. The potential role of Arhgef33 RhoGEF in foveal development in the zebra finch retina. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage | n | Incubation Time (Approximate) |
|---|---|---|
| St11 | 2 | 54 h |
| St15 | 3 | 66 h |
| St16 | 3 | 3 days |
| St19 | 3 | 3.5 days |
| St20 | 3 | 3.5 days |
| St24 | 3 | 4.5 days |
| St25 | 3 | 5 days |
| St37 | 3 | 8 days |
| St42 | 3 | 11 days |
| St44 | 3 | 13 days |
| P0 | 3 | 14 days |
| P1 | 3 | 15 days |
| P5 | 3 | 19 days |
| P8 | 3 | 22 days |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-Hernán, G.; de Mera-Rodríguez, J.A.; Hernández-Núñez, I.; Marzal, A.; Gañán, Y.; Martín-Partido, G.; Rodríguez-León, J.; Francisco-Morcillo, J. Analysis of Programmed Cell Death and Senescence Markers in the Developing Retina of an Altricial Bird Species. Cells 2021, 10, 504. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10030504
Álvarez-Hernán G, de Mera-Rodríguez JA, Hernández-Núñez I, Marzal A, Gañán Y, Martín-Partido G, Rodríguez-León J, Francisco-Morcillo J. Analysis of Programmed Cell Death and Senescence Markers in the Developing Retina of an Altricial Bird Species. Cells. 2021; 10(3):504. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10030504
Chicago/Turabian StyleÁlvarez-Hernán, Guadalupe, José Antonio de Mera-Rodríguez, Ismael Hernández-Núñez, Alfonso Marzal, Yolanda Gañán, Gervasio Martín-Partido, Joaquín Rodríguez-León, and Javier Francisco-Morcillo. 2021. "Analysis of Programmed Cell Death and Senescence Markers in the Developing Retina of an Altricial Bird Species" Cells 10, no. 3: 504. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10030504