
Novel Technique for Retinal Nerve Cell Regeneration with Electrophysiological Functions Using Human Iris-Derived iPS Cells

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunohistochemistry of Human Iris Tissue
2.2. Cell Isolation and Culturing of Human Iris Tissue
2.3. Cell Sorting of Iris Tissue Stem/Progenitor Cells
2.4. Preparation and Verification Experiment of iPS Cells
2.5. Formation of Teratomas and Embryoid Bodies
2.6. Differentiation into Nerve Cells and Recoverin-Positive Cells
2.7. Differentiation into Retinal Ganglion Cells
2.8. Electrophysiology
3. Results
3.1. p75NTR-Positive Cells Observed in Human Iris Tissue and Cultured Cells
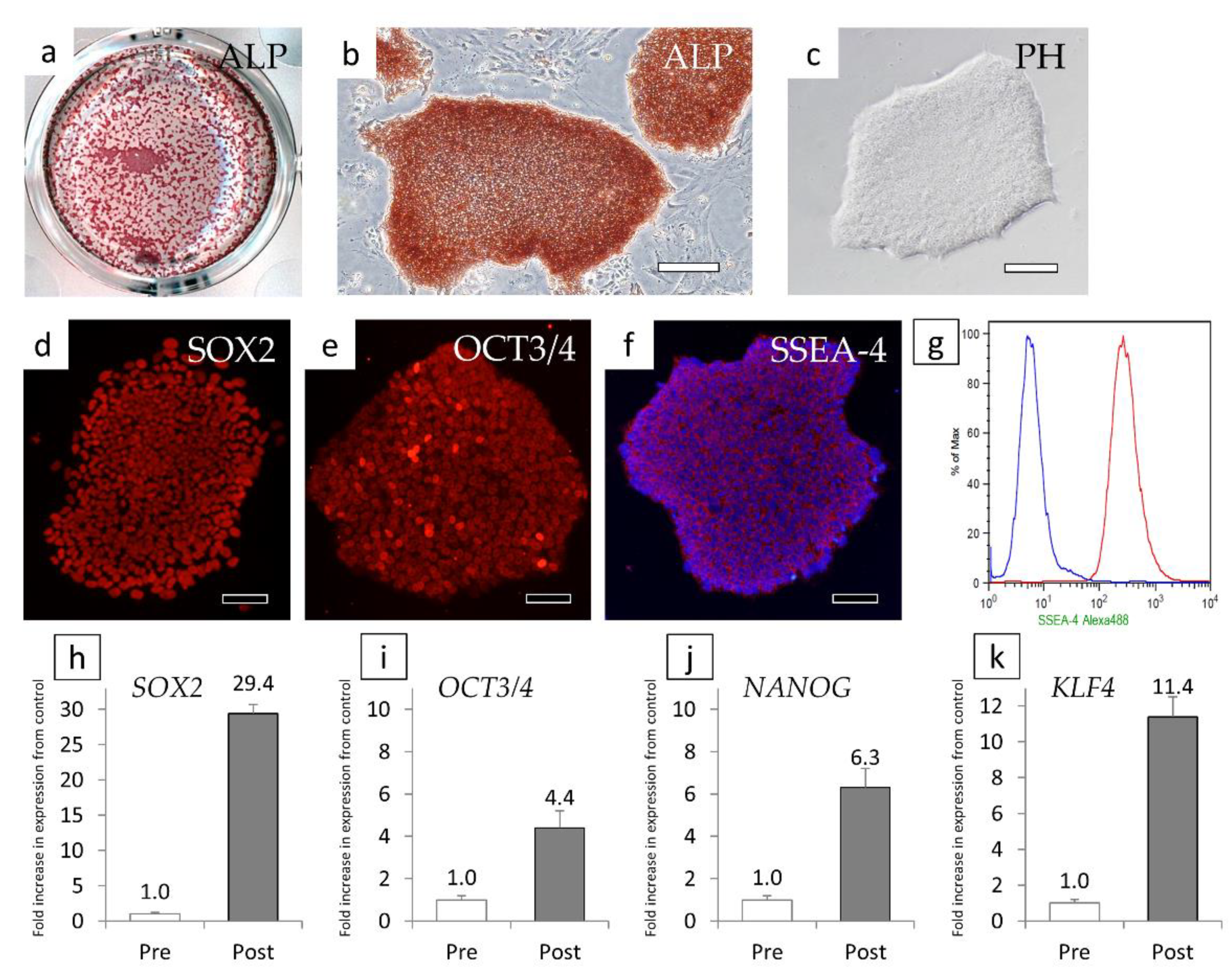
3.2. Preparation and Verification of iPS Cells Using Human Iris Tissue Stem/Progenitor Cells
3.3. Verification of Formed Teratomas and EBs
3.4. Differentiation into Nerve Cells and Recoverin-Positive Cells
3.5. Differentiation into Retinal Ganglion Cells
3.6. Electrophysiological Recording
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bianconi, E.; Piovesan, A.; Facchin, F.; Beraudi, A.; Casadei, R.; Frabetti, F.; Vitale, L.; Pelleri, M.C.; Tassani, S.; Piva, F.; et al. An estimation of the number of cells in the human body. Ann. Hum. Biol. 2013, 40, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen-Jones, S.M.; Occelli, L.M.; Winkler, P.A.; Lee, W.; Sparrow, J.R.; Tsukikawa, M.; Boye, S.L.; Chiodo, V.; Capasso, J.E.; Becirovic, E.; et al. Patients and animal models of CNGβ1-deficient retinitis pigmentosa support gene augmentation approach. J. Clin. Investig. 2018, 128, 190–206. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, A.; Shihabeddin, E.; Atkinson, J.A.; Nguyen, D.; Lin, Y.P.; O’Brien, J. A zebrafish model of retinitis pigmentosa shows continuous degeneration and regeneration of rod photoreceptors. Cells 2020, 9, 2242. [Google Scholar] [CrossRef] [PubMed]
- Morizane, Y.; Morimoto, N.; Fujiwara, A.; Kawasaki, R.; Yamashita, H.; Ogura, Y.; Shiraga, F. Incidence and causes of visual impairment in Japan: The first nation-wide complete enumeration survey of newly certified visually impaired individuals. Jpn. J. Ophthalmol. 2019, 63, 26–33. [Google Scholar] [CrossRef]
- Takahashi, M.; Palmer, T.D.; Takahashi, J.; Gage, F.H. Widespread integration and survival of adult-derived neural progenitor cells in the developing optic retina. Mol. Cell Neurosci. 1998, 12, 340–348. [Google Scholar] [CrossRef]
- Nishida, A.; Takahashi, M.; Tanihara, H.; Nakano, I.; Takahashi, J.B.; Mizoguchi, A.; Ide, C.; Honda, Y. Incorporation and differentiation of hippocampus-derived neural stem cells transplanted in injured adult rat retina. Investig. Ophthalmol. Vis. Sci. 2000, 41, 4268–4274. [Google Scholar]
- Kurimoto, Y.; Shibuki, H.; Kaneko, Y.; Ichikawa, M.; Kurokawa, T.; Takahashi, M.; Yoshimura, N. Transplantation of adult rat hippocampus-derived neural stem cells into retina injured by transient ischemia. Neurosci. Lett. 2001, 306, 57–60. [Google Scholar] [CrossRef]
- Akita, J.; Takahashi, M.; Hojo, M.; Nishida, A.; Haruta, M.; Honda, Y. Neuronal differentiation of adult rat hippocampus-derived neural stem cells transplanted into embryonic rat explanted retinas with retinoic acid pretreatment. Brain Res. 2002, 954, 286–293. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Ooto, S.; Akagi, T.; Amemiya, K.; Igarashi, R.; Mizushima, Y.; Takahashi, M. Effects of prolonged delivery of brain-derived neurotrophic factor on the fate of neural stem cells transplanted into the developing rat retina. Biochem. Biophys. Res. Commun. 2003, 309, 843–847. [Google Scholar] [CrossRef]
- Reh, T.A.; Tully, T. Regulation of tyrosine hydroxylase-containing amacrine cell number in larval frog retina. Dev. Biol. 1986, 114, 463–469. [Google Scholar] [CrossRef]
- Turner, D.L.; Cepko, C.L. A common progenitor for neurons and glia persists in rat retina late in development. Nature 1987, 328, 131–136. [Google Scholar] [CrossRef]
- Reh, T.A.; Levine, E.M. Multipotential stem cells and progenitors in the vertebrate retina. J. Neurobiol. 1998, 36, 206–220. [Google Scholar] [CrossRef]
- Ahmad, I.; Dooley, C.M.; Thoreson, W.B.; Rogers, J.A.; Afiat, S. In vitro analysis of a mammalian retinal progenitor that gives rise to neurons and glia. Brain Res. 1999, 831, 1–10. [Google Scholar] [CrossRef]
- Tropepe, V.; Coles, B.L.; Chiasson, B.J.; Horsford, D.J.; Elia, A.J.; McInnes, R.R.; van der Kooy, D. Retinal stem cells in the adult mammalian eye. Science 2000, 287, 2032–2036. [Google Scholar] [CrossRef]
- Fischer, A.J.; Reh, T.A. Muller glia are a potential source of neural regeneration in the postnatal chicken retina. Nat. Neurosci. 2001, 4, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Ooto, S.; Akagi, T.; Kageyama, R.; Akita, J.; Mandai, M.; Honda, Y.; Takahashi, M. Potential for neural regeneration after neurotoxic injury in the adult mammalian retina. Proc. Natl. Acad. Sci. USA 2004, 101, 13654–13659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, H.; Osakada, F.; Watanabe, K.; Mizuseki, K.; Haraguchi, T.; Miyoshi, H.; Kamiya, D.; Honda, Y.; Sasai, N.; Yoshimura, N.; et al. Generation of Rx+/Pax6+ neural retinal precursors from embryonic stem cells. Proc. Natl. Acad. Sci. USA 2005, 102, 11331–11336. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Kamiya, D.; Nishiyama, A.; Katayama, T.; Nozaki, S.; Kawasaki, H.; Watanabe, Y.; Mizuseki, K.; Sasai, Y. Directed differentiation of telencephalic precursors from embryonic stem cells. Nat. Neurosci. 2005, 8, 288–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakada, F.; Ikeda, H.; Mandai, M.; Wataya, T.; Watanabe, K.; Yoshimura, N.; Akaike, A.; Sasai, Y.; Takahashi, M. Toward the generation of rod and cone photoreceptors from mouse, monkey and human embryonic stem cells. Nat. Biotechnol. 2008, 26, 215–224. [Google Scholar] [CrossRef]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell-Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, G.; Abe, S.I.; Watanabe, K. Differentiation of lens-like structures from newt iris epithelial cells in vitro. Proc. Natl. Acad. Sci. USA 1974, 71, 5052–5056. [Google Scholar] [CrossRef] [Green Version]
- Kodama, R.; Eguchi, G. From lens regeneration in the newt to in-vitro transdifferentiation of vertebrate pigmented epithelial cells. Semin. Cell Biol. 1995, 6, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, M.; Kodama, R.; Eguchi, G. In vitro culture system for iris-pigmented epithelial cells for molecular analysis of transdifferentiation. Exp. Cell Res. 1998, 245, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Takeda, Y.; Yamada, K.; Akaishi, K.; Tomita, H.; Sato, M.; Tamai, M. Cytokine gene expression after subretinal transplantation. Tohoku J. Exp. Med. 1999, 189, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, N.; Tanikawa, A.; Horiguchi, M. Basic study of retinal stem/progenitor cell separation from mouse iris tissue. Med. Mol. Morphol. 2010, 43, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Asami, M.; Ohta, H.; Kosaka, J.; Kosaka, M. Retinal stem/progenitor properties of iris pigment epithelial cells. Dev. Biol. 2006, 289, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Takeda, S.; Hatsusaka, N.; Hiramatsu, N.; Nagai, N.; Deguchi, S.; Nakazawa, Y.; Takata, T.; Kodera, S.; Hirata, A.; et al. Effect of a Lens Protein in Low-Temperature Culture of Novel Immortalized Human Lens Epithelial Cells (iHLEC-NY2). Cells 2020, 9, 2670. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu, N.; Nagai, N.; Kondo, M.; Imaizumi, K.; Sasaki, H.; Yamamoto, N. Morphological comparison between three-dimensional structure of immortalized human lens epithelial cells and Soemmering’s ring. Med. Mol. Morphol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Isogai, S.; Yamamoto, N.; Hiramatsu, N.; Goto, Y.; Hayashi, M.; Kondo, M.; Imaizumi, K. Preparation of Induced Pluripotent Stem Cells Using Human Peripheral Blood Monocytes. Cell Reprogram. 2018, 20, 347–355. [Google Scholar] [CrossRef]
- Tanaka, T.; Yokoi, T.; Tamalu, F.; Watanabe, S.; Nishina, S.; Azuma, N. Generation of retinal ganglion cells with functional axons from human induced pluripotent stem cells. Sci. Rep. 2015, 5, 8344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch. 1981, 391, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Kawai, F.; Horiguchi, M.; Suzuki, H.; Miyachi, E. Na(+) action potentials in human photoreceptors. Neuron 2001, 30, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Ohkuma, M.; Kawai, F.; Horiguchi, M.; Miyachi, E. Patch-clamp recording of human retinal photoreceptors and bipolar cells. Photochem. Photobiol. 2007, 83, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Akamatsu, H.; Hasegawa, S.; Yamada, T.; Nakata, S.; Ohkuma, M.; Miyachi, E.; Marunouchi, T.; Matsunaga, K. Isolation of multipotent stem cells from mouse adipose tissue. J. Dermatol. Sci. 2007, 48, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Ishimura, D.; Yamamoto, N.; Tajima, K.; Ohno, A.; Yamamoto, Y.; Washimi, O.; Yamada, H. Differentiation of adipose-derived stromal vascular fraction culture cells into chondrocytes using the method of cell sorting with a mesenchymal stem cell marker. Tohoku J. Exp. Med. 2008, 216, 149–156. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Akamatsu, H.; Hasegawa, S.; Yamamoto, N.; Yoshimura, T.; Hasebe, Y.; Inoue, Y.; Mizutani, H.; Uzawa, T.; Matsunaga, K.; et al. Age-related changes of p75 neurotrophin receptor-positive adipose-derived stem cells. J. Dermatol. Sci. 2010, 58, 36–42. [Google Scholar] [CrossRef]
- Kim, K.; Doi, A.; Wen, B.; Ng, K.; Zhao, R.; Cahan, P.; Kim, J.; Aryee, M.J.; Ji, H.; Ehrlich, L.I.; et al. Epigenetic memory in induced pluripotent stem cells. Nature 2010, 467, 285–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishizawa, M.; Chonabayashi, K.; Nomura, M.; Tanaka, A.; Nakamura, M.; Inagaki, A.; Nishikawa, M.; Takei, I.; Oishi, A.; Tanabe, K.; et al. Epigenetic Variation between Human Induced Pluripotent Stem Cell Lines Is an Indicator of Differentiation Capacity. Cell Stem. Cell 2016, 19, 341–354. [Google Scholar] [CrossRef] [Green Version]
- Sridhar, A.; Ohlemacher, S.K.; Langer, K.B.; Meyer, J.S. Robust Differentiation of mRNA-Reprogrammed Human Induced Pluripotent Stem Cells Toward a Retinal Lineage. Stem. Cells Transl. Med. 2016, 5, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Kamao, H.; Mandai, M.; Okamoto, S.; Sakai, N.; Suga, A.; Sugita, S.; Kiryu, J.; Takahashi, M. Characterization of human induced pluripotent stem cell-derived retinal pigment epithelium cell sheets aiming for clinical application. Stem. Cell Rep. 2014, 2, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, E.; Koide, N.; Hanzawa, H.; Kiyama, M.; Ohta, M.; Kuwabara, J.; Takeda, S.; Takahashi, M. Fabricating retinal pigment epithelial cell sheets derived from human induced pluripotent stem cells in an automated closed culture system for regenerative medicine. PLoS ONE 2019, 14, e0212369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, R.; Ishikawa, Y.; Sasamoto, Y.; Katori, R.; Nomura, N.; Ichikawa, T.; Araki, S.; Soma, T.; Kawasaki, S.; Sekiguchi, K.; et al. Co-ordinated ocular development from human iPS cells and recovery of corneal function. Nature 2016, 531, 376–380. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Person/Organization | Event |
|---|---|---|
| 1981 | Dr. M. Evans Dr. M. Kaufman | Created embryonic stem cells (ES cells) derived from the inner cell mass of mouse blastocysts [2] |
| 1998 | Prof. J. Thomson et al. | Successful production of human ES cells [3] |
| 2006 | Prof. S. Yamanaka et al. | Succeeded in producing iPS cells using skin cells of adult mice [4] |
| 2007 | Prof. S. Yamanaka et al. | Succeeded in producing iPS cells using human fibroblast cells [5] |
| 2009 | U.S. President B. Obama | Lifted the ban against the use of federal funding for human ES cell research |
| Cell Type | Cell Origin | Ethical Issues | Gene Transfer | Differentiation Ability |
|---|---|---|---|---|
| Embryonic stem (ES) cells | Fertilized egg (inner cell mass) | Yes | No | Versatile (every cell of adult tissue) |
| Induced pluripotent stem (iPS) cells | Body cells (autologous cells) | No | Yes | Versatile (every cell of adult tissue) |
| Somatic stem cells (tissue stem cells) | Body cells (autologous cells) | No | No | Limited (only limited cell types can be differentiated) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, N.; Hiramatsu, N.; Ohkuma, M.; Hatsusaka, N.; Takeda, S.; Nagai, N.; Miyachi, E.-i.; Kondo, M.; Imaizumi, K.; Horiguchi, M.; et al. Novel Technique for Retinal Nerve Cell Regeneration with Electrophysiological Functions Using Human Iris-Derived iPS Cells. Cells 2021, 10, 743. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10040743
Yamamoto N, Hiramatsu N, Ohkuma M, Hatsusaka N, Takeda S, Nagai N, Miyachi E-i, Kondo M, Imaizumi K, Horiguchi M, et al. Novel Technique for Retinal Nerve Cell Regeneration with Electrophysiological Functions Using Human Iris-Derived iPS Cells. Cells. 2021; 10(4):743. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10040743
Chicago/Turabian StyleYamamoto, Naoki, Noriko Hiramatsu, Mahito Ohkuma, Natsuko Hatsusaka, Shun Takeda, Noriaki Nagai, Ei-ichi Miyachi, Masashi Kondo, Kazuyoshi Imaizumi, Masayuki Horiguchi, and et al. 2021. "Novel Technique for Retinal Nerve Cell Regeneration with Electrophysiological Functions Using Human Iris-Derived iPS Cells" Cells 10, no. 4: 743. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10040743