
Longer Sperm Swim More Slowly in the Canary Islands Chiffchaff

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Procedures
2.2. Sperm Analysis
2.3. Statistical Analysis on Sperm
2.4. Genetic Analyses
3. Results
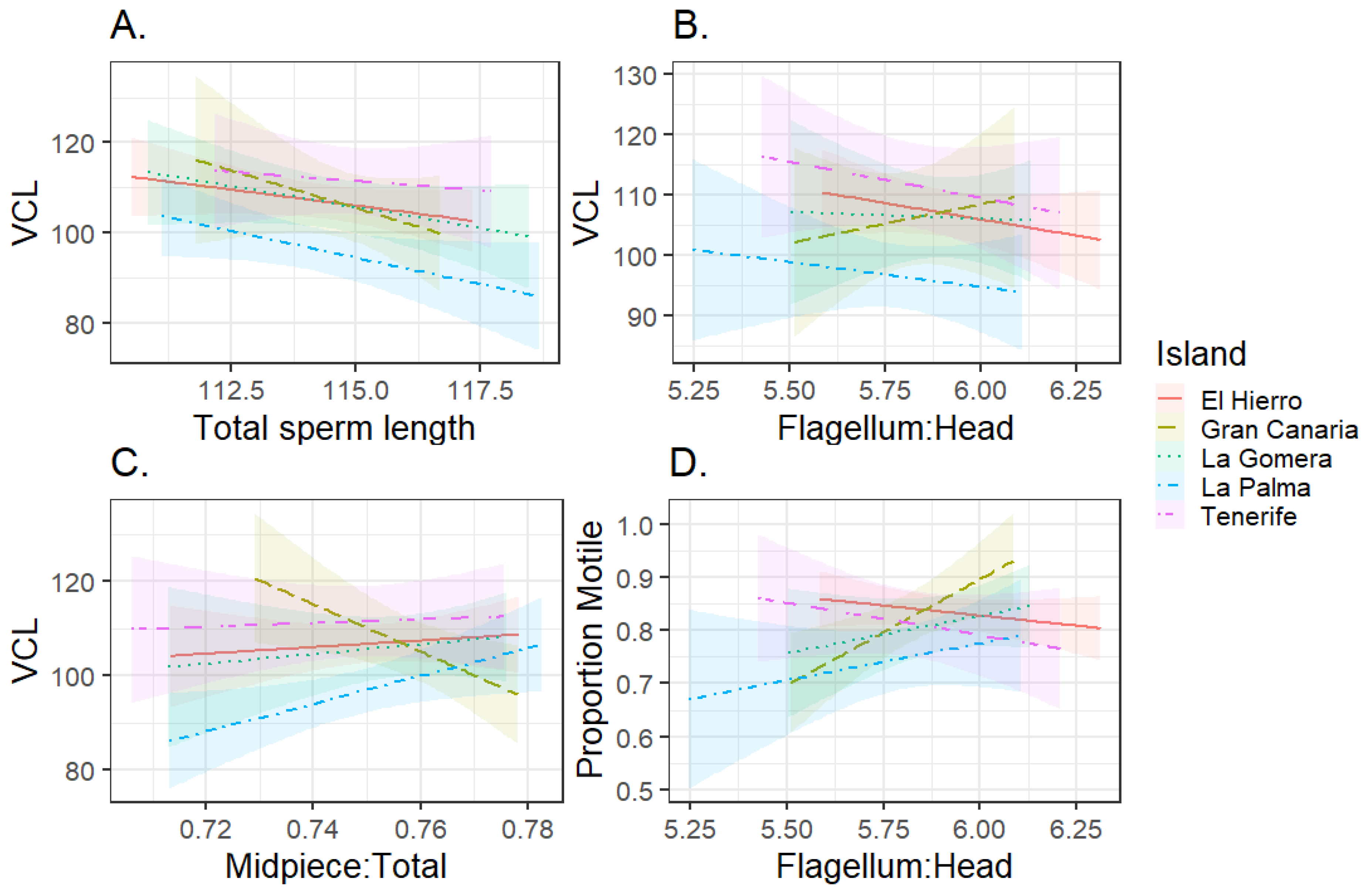
3.1. Sperm Morphology–Function Relationships
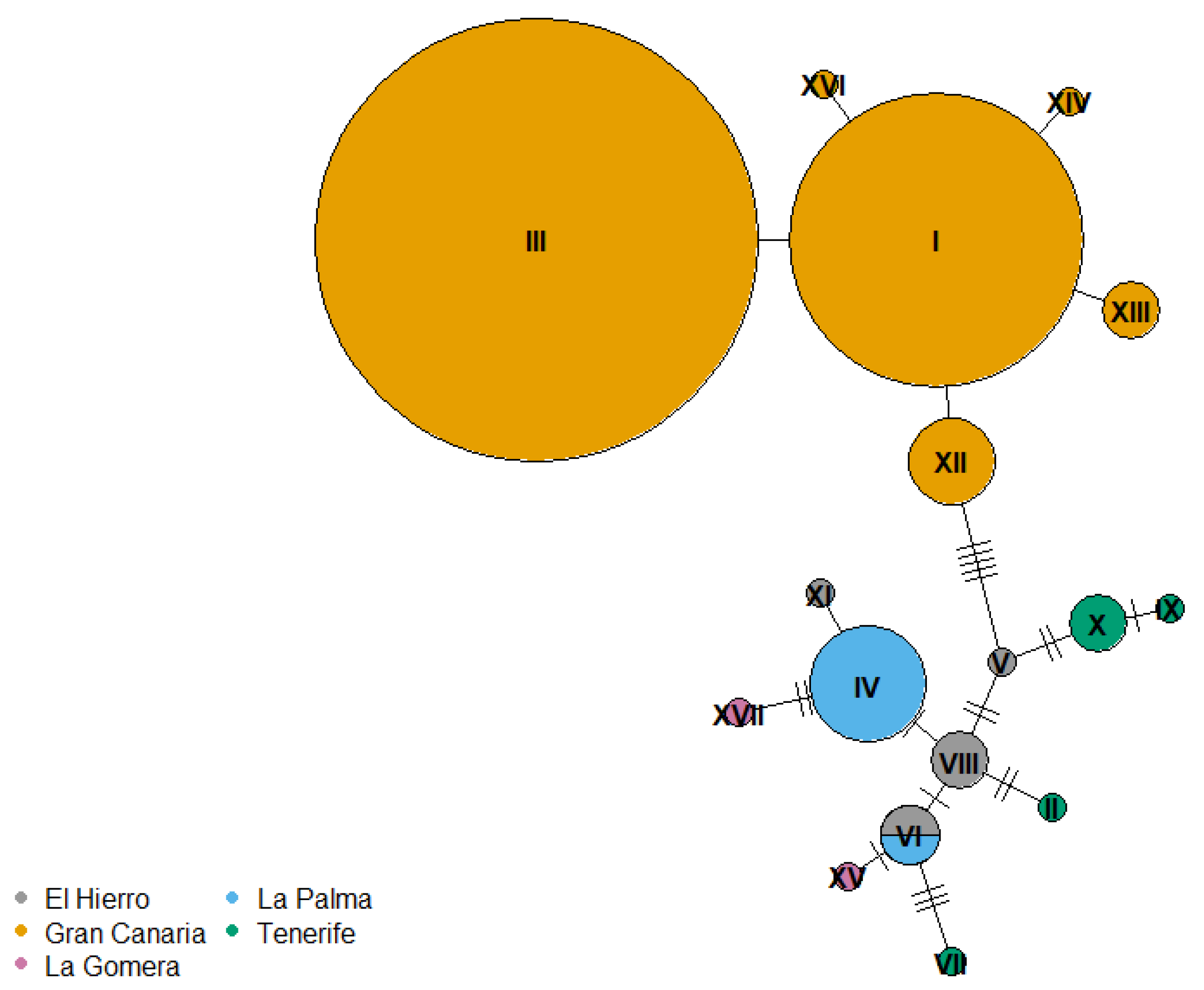
3.2. Among-Island Comparison of Sperm Phenotype and Genetic Divergence
4. Discussion
4.1. Sperm Morphology–Motility Relationships in Passerines
4.2. Sperm Morphology–Motility Relationships in the Canary Islands Chiffchaff
4.3. Sperm Evolution in the Canary Islands Chiffchaff
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parker, G. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970, 45, 525–567. [Google Scholar] [CrossRef]
- Parker, G.A. Sperm competition games: Raffles and roles. Proc. R. Soc. Lond. B 1990, 242, 120–126. [Google Scholar]
- Wedell, N.; Gage, M.J.G.; Parker, G.A. Sperm competition, male prudence and sperm-limited females. Trends Ecol. Evol. 2002, 17, 313–320. [Google Scholar] [CrossRef]
- Pizzari, T.; Parker, G.A. Sperm competition and sperm phenotype. In Sperm Biology: An Evolutionary Perspective; Elsevier: Amsterdam, The Netherlands, 2009; pp. 207–245. [Google Scholar]
- Parker, G.A. Sperm competition and the evolution of ejaculates: Towards a theory base. In Sperm Competition and Sexual Selection; Birkhead, T.R., Møller, A.P., Eds.; Academic Press: London, UK, 1998; pp. 3–54. [Google Scholar]
- Snook, R.R. Sperm in competition: Not playing by the numbers. Trends Ecol. Evol. 2005, 20, 46–53. [Google Scholar] [CrossRef]
- Simmons, L.W.; Fitzpatrick, J.L. Sperm wars and the evolution of male fertility. Reproduction 2012, 144, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Birkhead, T.R.; Martínez, J.G.; Burke, T.; Froman, D.P. Sperm mobility determines the outcome of sperm competition in the domestic fowl. Proc. R. Soc. B Biol. Sci. 1999, 266, 1759–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, J.L.; Humphries, S.; Evans, J.P.; Simmons, L.W.; Fitzpatrick, J.L. Relationships between sperm length and speed differ among three internally and three externally fertilizing species. Evolution 2014, 68, 92–104. [Google Scholar] [CrossRef]
- Fisher, H.S.; Jacobs-Palmer, E.; Lassance, J.M.; Hoekstra, H.E. The genetic basis and fitness consequences of sperm midpiece size in deer mice. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Cramer, E.R.A.; Laskemoen, T.; Stensrud, E.; Rowe, M.; Haas, F.; Lifjeld, J.T.; Sætre, G.-P.; Johnsen, A. Morphology-function relationships and repeatability in the sperm of Passer sparrows. J. Morphol. 2015, 276, 370–377. [Google Scholar] [CrossRef]
- Rojas Mora, A.; Meniri, M.; Ciprietti, S.; Helfenstein, F. Is sperm morphology functionally related to sperm swimming ability? A case study in a wild passerine bird with male hierarchies. BMC Evol. Biol. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Lifjeld, J.T.; Laskemoen, T.; Kleven, O.; Pedersen, A.T.M.; Lampe, H.M.; Rudolfsen, G.; Schmoll, T.; Slagsvold, T. No evidence for pre-copulatory sexual selection on sperm length in a passerine bird. PLoS ONE 2012, 7, e32611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomendio, M.; Roldan, E.R. Sperm competition influences sperm size in mammals. Proc. Biol. Sci. 1991, 243, 181–185. [Google Scholar] [CrossRef]
- Humphries, S.; Evans, J.P.; Simmons, L.W. Sperm competition: Linking form to function. BMC Evol. Biol. 2008, 8, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, E.M. Life at low Reynolds number. Am. J. Phys. 1977, 45, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Simons, J.E.; Olson, S.D. Sperm motility: Models for dynamic behavior in complex environments. In Cell Movement, Modeling and Simulation in Science, Engineering and Technology; Stolarska, M., Tarfulea, N., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2018; pp. 169–209. ISBN 9783319968421. [Google Scholar]
- Lüpold, S.; Calhim, S.; Immler, S.; Birkhead, T.R. Sperm morphology and sperm velocity in passerine birds. Proc. R. Soc. B Biol. Sci. 2009, 276, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Tourmente, M.; Gomendio, M.; Roldan, E.R.S. Sperm competition and the evolution of sperm form and function in mammals. BMC Evol. Biol. 2011, 11, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mossman, J.; Slate, J.; Humphries, S.; Birkhead, T. Sperm morphology and velocity are genetically codetermined in the zebra finch. Evolution 2009, 63, 2730–2737. [Google Scholar] [CrossRef] [PubMed]
- Helfenstein, F.; Podevin, M.; Richner, H. Sperm morphology, swimming velocity, and longevity in the house sparrow Passer domesticus. Behav. Ecol. Sociobiol. 2010, 64, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Kleven, O.; Fossøy, F.; Laskemoen, T.; Robertson, R.J.; Rudolfsen, G.; Lifjeld, J.T. Comparative evidence for the evolution of sperm swimming speed by sperm competition and female sperm storage duration in passerine birds. Evolution 2009, 63, 2466–2473. [Google Scholar] [CrossRef] [PubMed]
- Cardullo, R.A.; Baltz, J.M. Metabolic regulation in mammalian sperm: Mitochondrial volume determines sperm length and flagellar beat frequency. Cell Motil. Cytoskelet. 1991, 19, 180–188. [Google Scholar] [CrossRef]
- Tourmente, M.; Rowe, M.; González-Barroso, M.M.; Rial, E.; Gomendio, M.; Roldan, E.R.S. Postcopulatory sexual selection increases ATP content in rodent spermatozoa. Evolution 2013, 67, 1838–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, R.M. Tales from the tail: What do we really know about sperm motility? J. Androl. 2003, 24, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marki, P.Z. Diversification, Geographic Expansion and Trait Evolution among Passerine Birds. Ph.D. Thesis, University of Copenhagen and University of Oslo, Oslo, Norway, 2017. [Google Scholar]
- Laskemoen, T.; Kleven, O.; Fossøy, F.; Robertson, R.J.; Rudolfsen, G.; Lifjeld, J.T. Sperm quantity and quality effects on fertilization success in a highly promiscuous passerine, the tree swallow Tachycineta bicolor. Behav. Ecol. Sociobiol. 2010, 64, 1473–1483. [Google Scholar] [CrossRef]
- Knief, U.; Forstmeier, W.; Pei, Y.; Ihle, M.; Wang, D.; Martin, K.; Opatová, P.; Albrechtová, J.; Wittig, M.; Franke, A.; et al. A sex-chromosome inversion causes strong overdominance for sperm traits that affect siring success. Nat. Ecol. Evol. 2017, 1, 1177–1184. [Google Scholar] [CrossRef]
- Losdat, S.; Helfenstein, F. Relationships between sperm morphological traits and sperm swimming performance in wild Great Tits (Parus major). J. Ornithol. 2018, 159, 805–814. [Google Scholar] [CrossRef]
- Sætre, C.L.C. Sperm morphology, sperm motility and paternity success in the bluethroat (Luscinia s. svecica). PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennison, C.; Hemmings, N.; Brookes, L.; Slate, J.; Birkhead, T. Sperm morphology, adenosine triphosphate (ATP) concentration and swimming velocity: Unexpected relationships in a passerine bird. Proc. R. Soc. B Biol. Sci. 2016, 283, 1558. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, H.; Wang, S.; Yang, W.; Ding, J.; Zhang, Y. Variation in sperm morphology and performance in tree sparrow (Passer montanus) under long-term environmental heavy metal pollution. Ecotoxicol. Environ. Saf. 2020, 197, 110622. [Google Scholar] [CrossRef] [PubMed]
- Rowe, M.; Laskemoen, T.; Johnsen, A.; Lifjeld, J.T. Evolution of sperm structure and energetics in passerine birds. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122616. [Google Scholar] [CrossRef] [Green Version]
- Cramer, E.R.A.; Rowe, M.; Eroukhmanoff, F.; Lifjeld, J.T.; Sætre, G.P.; Johnsen, A. Measuring sperm swimming performance in birds: Effects of dilution, suspension medium, mechanical agitation, and sperm number. J. Ornithol. 2019, 160, 1053–1063. [Google Scholar] [CrossRef]
- Cramer, E.R.A.; Stensrud, E.; Marthinsen, G.; Hogner, S.; Johannessen, L.E.; Laskemoen, T.; Eybert, M.-C.; Slagsvold, T.; Lifjeld, J.; Johnsen, A. Sperm performance in conspecific and heterospecific female fluid. Ecol. Evol. 2016, 6, 1363–1377. [Google Scholar] [CrossRef] [Green Version]
- Vernon, G.G.; Woolley, D.M. Three-dimensional motion of avian spermatozoa. Cell Motil. Cytoskelet. 1999, 42, 149–161. [Google Scholar] [CrossRef]
- Allen, T.E.; Grigg, G.W. Sperm transport in the fowl. Aust. J. Agric. Res. 1957, 8, 788–799. [Google Scholar] [CrossRef]
- Bakst, M.R.; Wishart, G.; Brillard, J.-P. Oviductal sperm selection, transport, and storage in poultry. Poult. Sci. Rev. 1994, 5, 117–143. [Google Scholar]
- Bennison, C.; Hemmings, N.; Slate, J.; Birkhead, T. Long sperm fertilize more eggs in a bird. Proc. R. Soc. B Biol. Sci. 2015, 282, 20141897. [Google Scholar] [CrossRef] [PubMed]
- Hemmings, N.; Bennison, C.; Birkhead, T.R. Intra-ejaculate sperm selection in female zebra finches. Biol. Lett. 2016, 12, 4–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donoghue, A.M.; Kirby, J.D.; Froman, D.P.; Lerner, S.P.; Crouch, A.N.; King, L.M.; Donoghue, D.J.; Sonstegard, T.S. Field testing the influence of sperm competition based on sperm mobility in breeder turkey toms. Br. Poult. Sci. 2003, 44, 498–504. [Google Scholar] [CrossRef]
- Denk, A.G.; Holzmann, A.; Peters, A.; Vermeirssen, E.L.M.; Kempenaers, B. Paternity in mallards: Effects of sperm quality and female sperm selection for inbreeding avoidance. Behav. Ecol. 2005, 16, 825–833. [Google Scholar] [CrossRef] [Green Version]
- Rowe, M.; Albrecht, T.; Cramer, E.R.A.; Johnsen, A.; Laskemoen, T.; Weir, J.T.; Lifjeld, J.T. Postcopulatory sexual selection is associated with accelerated evolution of sperm morphology. Evolution 2015, 69, 1044–1052. [Google Scholar] [CrossRef] [PubMed]
- Lifjeld, J.T.; Gohli, J.; Albrecht, T.; Garcia-Del-Rey, E.; Johannessen, L.E.; Kleven, O.; Marki, P.Z.; Omotoriogun, T.C.; Rowe, M.; Johnsen, A. Evolution of female promiscuity in Passerides songbirds. BMC Evol. Biol. 2019, 19, 7–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleven, O.; Laskemoen, T.; Fossøy, F.; Robertson, R.J.; Lifjeld, J.T. Intraspecific variation in sperm length is negatively related to sperm competition in passerine birds. Evolution 2008, 62, 494–499. [Google Scholar] [CrossRef]
- Calhim, S.; Immler, S.; Birkhead, T.R. Postcopulatory sexual selection is associated with reduced variation in sperm morphology. PLoS ONE 2007, 2, e413. [Google Scholar] [CrossRef]
- Immler, S.; Calhim, S.; Birkhead, T.R. Increased postcopulatory sexual selection reduces the intramale variation in sperm design. Evolution 2008, 62, 1538–1543. [Google Scholar] [CrossRef]
- Cramer, E.R.A.; Rowlison, T.; Comizzoli, P.; Ryder, T.B. Uniform sperm morphology in the lek-breeding wire-tailed manaking (Pipra filicauda). Ornitol. Neotrop. 2019, 30, 135–139. [Google Scholar]
- Garcia-del-Rey, E. Birds of the Canary Islands; Sociedad Ornitologica Canaria: Santa Cruz de Tenerife, Spain, 2015. [Google Scholar]
- Gohli, J.; Leder, E.H.; Garcia-del-Rey, E.; Johannessen, L.E.; Johnsen, A.; Laskemoen, T.; Popp, M.; Lifjeld, J.T. The evolutionary history of Afrocanarian blue tits inferred from genomewide SNPs. Mol. Ecol. 2015, 24, 180–191. [Google Scholar] [CrossRef]
- Lifjeld, J.T.; Anmarkrud, J.A.; Calabuig, P.; Cooper, J.E.J.; Johannessen, L.E.; Johnsen, A.; Kearns, A.M.; Lachlan, R.F.; Laskemoen, T.; Marthinsen, G.; et al. Species-level divergences in multiple functional traits between the two endemic subspecies of Blue Chaffinches Fringilla teydea in Canary Islands. BMC Zool. 2016, 1, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Illera, J.C.; Ramírez, Á.; Rodríguez, L.; Polypathellis, K.; Pérez-Tris, J. Maternal Genetic Structure Reveals an Incipient Differentiation in the Canary Islands Chiffchaff Phylloscopus canariensis. Ardeola 2020, 67, 401–414. [Google Scholar] [CrossRef]
- Laskemoen, T.; Kleven, O.; Johannessen, L.E.; Fossøy, F.; Robertson, R.J.; Lifjeld, J.T. Repeatability of sperm size and motility within and between seasons in the Barn Swallow (Hirundo rustica). J. Ornithol. 2013, 154, 955–963. [Google Scholar] [CrossRef]
- Laskemoen, T.; Kleven, O.; Fossøy, F.; Lifjeld, J.T. Intraspecific variation in sperm length in two passerine species, the Bluethroat Luscinia svecica and the Willow Warbler Phylloscopus trochilus. Ornis Fenn. 2007, 84, 131–139. [Google Scholar]
- Sokal, R.; Rohlf, F. Biometry: The Principles and Practice of Statistics in Biological Research; WH Freeman and Co.: San Francisco, CA, USA, 1995. [Google Scholar]
- Schielzeth, H. Simple means to improve the interpretability of regression coefficients. Methods Ecol. Evol. 2010, 1, 103–113. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fitzpatrick, J.L.; Baer, B. Polyandry reduces sperm length variation in social insects. Evolution 2011, 65, 3006–3012. [Google Scholar] [CrossRef]
- Packard, G.C.; Boardman, T.J. The use of percentages and size-specific indices to normalize physiological data for variation in body size: Wasted time, wasted effort? Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 1999, 122, 37–44. [Google Scholar] [CrossRef]
- van de Pol, M.; Wright, J. A simple method for distinguishing within- versus between-subject effects using mixed models. Anim. Behav. 2009, 77, 753–758. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. Linear and Nonlinear Mixed-Effects Models, R package version 3.1-113. 2020; R. Foundation for Statistical Computing: Vienna, Austria, 2007. [Google Scholar]
- Cramer, E.R.A.; Laskemoen, T.; Kleven, O.; Lifjeld, J.T. Sperm length variation in House Wrens Troglodytes aedon. J. Ornithol. 2013, 154, 129–138. [Google Scholar] [CrossRef]
- Albrecht, T.; Opletalová, K.; Reif, J.; Janoušek, V.; Piálek, L.; Cramer, E.R.A.; Johnsen, A.; Reifová, R. Sperm divergence in a passerine contact zone: Indication of reinforcement at the gametic level. Evolution 2019, 73, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Lüpold, S.; Birkhead, T.R.; Westneat, D.F. Seasonal variation in ejaculate traits of male red-winged blackbirds (Agelaius phoeniceus). Behav. Ecol. Sociobiol. 2012, 66, 1607–1617. [Google Scholar] [CrossRef]
- Edme, A.; Zobač, P.; Korsten, P.; Albrecht, T.; Schmoll, T.; Krist, M. Moderate heritability and low evolvability of sperm morphology in a species with high risk of sperm competition, the collared flycatcher Ficedula albicollis. J. Evol. Biol. 2019, 32, 205–217. [Google Scholar] [CrossRef]
- Schmoll, T.; Kleven, O.; Rusche, M. Individual phenotypic plasticity explains seasonal variation in sperm morphology in a passerine bird. Evol. Ecol. Res. 2018, 19, 561–574. [Google Scholar]
- Paradis, E. Pegas: An R package for population genetics with an integrated—Modular approach. Bioinformatics 2010, 26, 419–420. [Google Scholar] [CrossRef] [Green Version]
- Mickova, K. Postcopulatory Sexual Selection on Phenotypic Traits in European Barn Swallows; Charles University: Prague, Czech Republic, 2018. [Google Scholar]
- Immler, S.; Pryke, S.R.; Birkhead, T.R.; Griffith, S.C. Pronounced within-individual plasticity in sperm morphometry across social environments. Evolution 2010, 64, 1634–1643. [Google Scholar] [CrossRef] [PubMed]
- Støstad, H.N.; Johnsen, A.; Lifjeld, J.; Rowe, M. Sperm head morphology is associated with sperm swimming speed: A comparative study of songbirds using electron microscopy. Evolution 2018, 72, 1918–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hook, K.A.; Fisher, H.S. Methodological considerations for examining the relationship between sperm morphology and motility. Mol. Reprod. Dev. 2020, 87, 633–649. [Google Scholar] [CrossRef]
- Schmoll, T.; Rudolfsen, G.; Schielzeth, H.; Kleven, O. Sperm velocity in a promiscuous bird across experimental media of different viscosities: Sperm velocity in viscous media. Proc. R. Soc. B Biol. Sci. 2020, 287. [Google Scholar] [CrossRef]
- Takeuchi, S.; Diluzio, W.R.; Weibel, D.B.; Whitesides, G.M. Controlling the shape of filamentous cells of Escherichia coli. Nano Lett. 2005, 5, 1819–1823. [Google Scholar] [CrossRef] [Green Version]
- Martínez, L.E.; Hardcastle, J.M.; Wang, J.; Pincus, Z.; Tsang, J.; Hoover, T.R.; Bansil, R.; Salama, N.R. Helicobacter pylori strains vary cell shape and flagellum number to maintain robust motility in viscous environments. Mol. Microbiol. 2016, 99, 88–110. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Gulino, M.; Morse, M.; Tang, J.X.; Powers, T.R.; Breuer, K.S. Helical motion of the cell body enhances Caulobacter crescentus motility. Proc. Natl. Acad. Sci. USA 2014, 111, 11252–11256. [Google Scholar] [CrossRef] [Green Version]
- Pizzari, T.; Cornwallis, C.K.; Froman, D.P. Social competitiveness associated with rapid fluctuations in sperm quality in male fowl. Proc. Biol. Sci. 2007, 274, 853–860. [Google Scholar] [CrossRef] [Green Version]
- Mendonca, T.; Birkhead, T.R.; Cadby, A.J.; Forstmeier, W.; Hemmings, N. A trade-off between thickness and length in the zebra finch sperm mid-piece. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [Green Version]
- Hogner, S.; Laskemoen, T.; Lifjeld, J.T.; Pavel, V.; Chutny, B.; Fernandez, J.G.; Eybert, M.-C.; Matsyna, E.; Johnsen, A. Rapid sperm evolution in the bluethroat (Luscinia svecica) subspecies complex. Behav. Ecol. Sociobiol. 2013, 67, 1205–1217. [Google Scholar] [CrossRef]
- Lifjeld, J.T.; Laskemoen, T.; Kleven, O.; Albrecht, T.; Robertson, R.J. Sperm length variation as a predictor of extrapair paternity in passerine birds. PLoS ONE 2010, 5, e13456. [Google Scholar] [CrossRef] [Green Version]
- Bjørnstad, G.; Lifjeld, J.T. High frequency of extra-pair paternity in a dense and synchronous population of willow warblers Phylloscopus trochilus. J. Avian Biol. 1997, 28, 319–324. [Google Scholar] [CrossRef]
- Gil, D.; Slater, P.J.B.; Graves, J.A. Extra-pair paternity and song characteristics in the willow warbler Phylloscopus trochilus. J. Avian Biol. 2007, 38, 291–297. [Google Scholar] [CrossRef]
- Valente, L.; Illera, J.C.; Havenstein, K.; Pallien, T.; Etienne, R.S.; Tiedemann, R. Equilibrium bird species diversity in Atlantic islands. Curr. Biol. 2017, 27, 1660–1666.e5. [Google Scholar] [CrossRef] [Green Version]
- Raković, M.; Neto, J.M.; Lopes, R.J.; Koblik, E.A.; Fadeev, I.V.; Lohman, Y.V.; Aghayan, S.A.; Boano, G.; Pavia, M.; Perlman, Y.; et al. Geographic patterns of mtDNA and Z-linked sequence variation in the Common Chiffchaff and the ‘chiffchaff complex. PLoS ONE 2019, 14, e210268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briskie, J.V.; Montgomerie, R. Patterns of sperm storage in relation to sperm competition in passerine birds. Condor 1993, 95, 442–454. [Google Scholar] [CrossRef]
- Briskie, J.V.; Montgomerie, R.; Birkhead, T.R. The evolution of sperm size in birds. Evolution 1997, 51, 937–945. [Google Scholar] [CrossRef]
- Hemmings, N.; Birkhead, T. Differential sperm storage by female zebra finches Taeniopygia guttata. Proc. R. Soc. B Biol. Sci. 2017, 284. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.W.; Lüpold, S.; Fitzpatrick, J.L. Evolutionary trade-off between secondary sexual traits and ejaculates. Trends Ecol. Evol. 2017, 32, 964–976. [Google Scholar] [CrossRef]
- Supriya, K.; Price, T.D.; Rowe, M. Resource variation generates positive correlations between pre- and postcopulatory sexual traits. Behav. Ecol. 2019, 30, 341–347. [Google Scholar] [CrossRef]
- Parker, G.A.; Lessells, C.M.; Simmons, L.W. Sperm competition games: A general model for precopulatory male—male competition. Evolution 2013, 67, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A. Sperm competition games: Sperm size and sperm number under adult control. Proc. R. Soc. B 1993, 253, 245–254. [Google Scholar] [PubMed]
- Supriya, K.; Rowe, M.; Laskemoen, T.; Mohan, D.; Price, T.D.; Lifjeld, J.T. Early diversification of sperm size in the evolutionary history of the old world leaf warblers (Phylloscopidae). J. Evol. Biol. 2016, 29, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Støstad, H.N.; Rekdal, S.L.; Kleven, O.; Laskemoen, T.; Marthinsen, G.; Johnsen, A.; Lifjeld, J.T. Weak geographical structure in sperm morphology across the range of two willow warbler Phylloscopus trochilus subspecies in Scandinavia. J. Avian Biol. 2016, 47, 731–741. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Island (n Males Morphology, Motility) | ANOVA or Levene’s Test | |||||
|---|---|---|---|---|---|---|
| Sperm Trait | El Hierro (23, 21) | Gran Canaria (30, 13) | La Gomera (11, 9) | La Palma (22, 22) | Tenerife (19, 8) | |
| Total sperm length (µm) | 114.19 ± 2.10 | 114.52 ± 1.55 | 114.85 ± 2.21 | 114.19 ± 2.03 | 114.10 ± 2.92 | F4,101.75 = 0.33, p = 0.86 |
| F:H | 5.90 ± 0.21 | 5.89 ± 0.18 | 5.82 ± 0.27 | 5.80 ± 0.21 | 5.66 ± 0.35 | F4,100.25 = 0.98, p = 0.42 |
| Midpiece length (µm) | 85.59 ± 2.73 | 87.07 ± 1.96 | 86.40 ± 2.43 | 85.40 ± 1.85 | 83.61 ± 6.20 | F4,99.04 = 1.42, p = 0.23 |
| CVwm | 1.46 ± 0.66 | 1.52 ± 0.63 | 1.39 ± 0.78 | 1.44 ± 0.34 | 1.37 ± 0.44 | F4,97.15 = 0.83, p = 0.51 |
| CVam | 1.86 | 1.37 | 1.97 | 1.8 | 2.59 | F4,102 = 1.81, p = 0.13 |
| VCL (µm/s) | 106.77 ± 8.50 | 105.99 ± 12.50 | 106.22 ± 7.85 | 96.41 ± 11.59 | 111.44 ± 8.07 | F4,61.57 = 7.49, p < 0.001 |
| Proportion motile | 0.83 ± 0.06 | 0.82 ± 0.10 | 0.81 ± 0.07 | 0.75 ± 0.13 | 0.81 ± 0.07 | F4, 34.77 = 2.15, p = 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cramer, E.R.A.; Garcia-del-Rey, E.; Johannessen, L.E.; Laskemoen, T.; Marthinsen, G.; Johnsen, A.; Lifjeld, J.T. Longer Sperm Swim More Slowly in the Canary Islands Chiffchaff. Cells 2021, 10, 1358. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061358
Cramer ERA, Garcia-del-Rey E, Johannessen LE, Laskemoen T, Marthinsen G, Johnsen A, Lifjeld JT. Longer Sperm Swim More Slowly in the Canary Islands Chiffchaff. Cells. 2021; 10(6):1358. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061358
Chicago/Turabian StyleCramer, Emily R. A., Eduardo Garcia-del-Rey, Lars Erik Johannessen, Terje Laskemoen, Gunnhild Marthinsen, Arild Johnsen, and Jan T. Lifjeld. 2021. "Longer Sperm Swim More Slowly in the Canary Islands Chiffchaff" Cells 10, no. 6: 1358. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061358