
Oncolytic Herpes Simplex Virus-Based Therapies for Cancer
by
 , , , and
, , , and
Norah Aldrak
1 ,
,
Sarah Alsaab
1,2,
Aliyah Algethami
1,
Deepak Bhere
3,4,5,
Hiroaki Wakimoto
3,4,5,6,
Khalid Shah
3,4,5,7,
Mohammad N. Alomary
1,2,* and
Nada Zaidan
1,2,* 1
Center of Excellence for Biomedicine, Joint Centers of Excellence Program, King Abdulaziz City for Science and Technology, P.O. Box 6086, Riyadh 11451, Saudi Arabia
2
National Center for Biotechnology, Life Science and Environmental Research Institute, King Abdulaziz City for Science and Technology, P.O. Box 6086, Riyadh 11451, Saudi Arabia
3
Center for Stem Cell Therapeutics and Imaging (CSTI), Brigham and Women’s Hospital, Harvard Medical School, Boston, MA 02115, USA
4
Department of Neurosurgery, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA 02115, USA
5
BWH Center of Excellence for Biomedicine, Brigham and Women’s Hospital, Harvard Medical School, Boston, MA 02115, USA
6
Department of Neurosurgery, Massachusetts General Hospital, Harvard Medical School, Boston, MA 02114, USA
7
Harvard Stem Cell Institute, Harvard University, Cambridge, MA 02138, USA
*
Authors to whom correspondence should be addressed.
Cells 2021, 10(6), 1541; https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061541
Submission received: 13 April 2021
/
Revised: 22 May 2021
/
Accepted: 26 May 2021
/
Published: 18 June 2021
Abstract
:With the increased worldwide burden of cancer, including aggressive and resistant cancers, oncolytic virotherapy has emerged as a viable therapeutic option. Oncolytic herpes simplex virus (oHSV) can be genetically engineered to target cancer cells while sparing normal cells. This leads to the direct killing of cancer cells and the activation of the host immunity to recognize and attack the tumor. Different variants of oHSV have been developed to optimize its antitumor effects. In this review, we discuss the development of oHSV, its antitumor mechanism of action and the clinical trials that have employed oHSV variants to treat different types of tumor.
1. Introduction
Cancer remains a serious global burden that threatens millions of lives worldwide. It is considered the second most common cause of death, with an annual worldwide incidence of 10 million cases [1]. The most common cancer therapies include surgery, chemotherapy, and radiotherapy [2]. Such traditional therapeutic approaches have detrimental consequences such as hematological toxicity, gastrointestinal toxicity, ototoxicity, hepatotoxicity, cardiotoxicity, neurotoxicity, and endocrine fluctuations [3]. Therefore, searching for alternative approaches that specifically target cancerous cells and are safer with no or moderate side effects is a priority for researchers around the globe. In particular, the use of engineered oncolytic viruses (OVs) is a promising alternative approach to specific targeting of tumors due to their ability to efficiently replicate in tumor cells [4]. Additionally, OVs promote a state of anti-tumor immunity [5]. Several OVs have been tested in clinical trials, such as herpesviruses, retroviruses, adenoviruses, vaccinia virus, and poliovirus [6]. These viruses vary in their sizes, genome nature, and replication efficiency [7].
Oncolytic herpes simplex viruses (oHSV) are among the few oncolytic viruses that were moved to phase III clinical trials. Herpes simplex virus (HSV), which belongs to the Herpesviridae family, has a large dsDNA (152 kb) which is encapsulated within an enveloped icosahedral capsid. HSV is a neurotropic virus with two variants: HSV-1 and HSV-2. They possess two significant features, which make them suitable candidates for targeting tumors. Firstly, the genome has approximately 30 kb encoding for nonessential genes [8]. This feature allows for genetic manipulation by adding or replacing genes through genetic recombination. Secondly, herpesviruses have a good safety profile, since they replicate in the nucleus without causing insertional mutagenesis [6]. According to their replication strategies, genetically engineered HSV-1 vectors are categorized into three groups: (i) replication defective, (ii) conditionally replicating and (iii) amplicons. Of these, conditionally replicating HSV-1 vectors are designed to specifically target cancerous cells. Moreover, they are engineered to have therapeutic transgenes to enhance anticancer immunity [9]. Several genetically modified variants of oHSV-1, such as NV1020 [10], G207 [11], T-VEC [12] and HSV1716 (Seprehvir) [13], have been tested extensively in clinical studies. Among these, FDA-approved T-VEC (trade name: Imlygic™) is the most widely used OV. As of December 2019, the clinical trials portal (https://clinicaltrials.gov/) has listed more than 38 clinical trials assessing the safety and effectiveness of T-VEC against several types of cancers. T-VEC was genetically engineered to delete the neurovirulence genes and the genes required for inhibition of antigen presentation [14,15]. Kaufman et al. reported that T-VEC was associated with high levels of melanoma-specific CD8+ T cells and decreased levels of immunosuppressive cells such as CD4+ FoxP3+ regulatory T cells [16]. Although T-VEC showed an excellent safety profile, common side effects include fever, nausea, vomiting and headache [17]. To provoke the antitumor immunity, two copies of granulocyte-macrophage colony-stimulating factor (GM-CSF) gene were engineered to replace the deleted neurovirulence gene [18]. Hence, in the current review, the following topics will be covered: (i) the major mechanisms by which oHSV induces tumor regression, (ii) the ongoing preclinical and clinical trials of oHSV, (iii) the role of oHSV in immunovirotherapy, and (iv) the limitations and ethical concerns regarding oHSV.
2. Mechanism of Oncolytic Herpes Simplex Virus-1 Antitumor Activity
2.1. Tumor-Selective Replication
HSV-1 is a double-stranded DNA virus that has a large genome containing 152 kb, 30 kb of which are dispensable for viral infection [19]. This makes HSV-1 an attractive candidate for genetic manipulation to enhance tumor-selectivity and patient safety. Although its site of replication is the nucleus, HSV-1 does not cause insertional mutagenesis and is sensitive to antiviral drugs such as acyclovir and ganciclovir [20]. Some of the most frequently modified non-essential viral genes in HSV-1 variants that have shown promise in preclinical and clinical studies include γ34.5, UL39, and α47, encoding infected-cell protein (ICP) 34.5, ICP6 and ICP47, respectively. These genes have evolved to provide wild type (WT) HSV-1 with the abilities to evade the host antiviral response and continue its replication cycle. In this section, we will discuss the function of these genes, and how manipulating them can play a role in conferring tumor selectivity.
γ34.5 is one of the most commonly mutated genes for creating oHSVs. It encodes ICP34.5, which allows WT HSV-1 to overcome the host cell protein synthesis shut-off response to viral infection. In WT HSV-1 infection, a normal cell activates protein kinase R (PKR) which inactivates eukaryotic initiation factor-2α (eIF-2α), leading to a shut down in protein synthesis. However, the presence of the ICP34.5 protein allows WT HSV-1 to restore protein synthesis by restoring eIF-2α function [21]. Therefore, deleting both copies of ICP34.5 hinders the HSV-1 ability to synthesize protein and subsequent propagation in normal cells. In contrast, the activation of PKR in tumor cells is often impaired, providing a preferential environment in which γ34.5-lacking HSV-1 can continue its life cycle. To improve safety and reduce the chance of reversion to WT, deletion of γ34.5 can be accompanied with UL39 inactivation (see below) to create G207, the first oHSV to be tested in a clinical trial in the US [19]. ICP6 is the large subunit in viral ribonuclease reductase, which converts ribonucleotides into deoxyribonucleotides that are utilized in viral genome synthesis [22]. Lacking ICP6 restricts viral replication to dividing cells, as mature, postmitotic cells lack ribonucleotide reductase expression and sufficient amounts of deoxyribonucleotides [19].
As an immune-evasion mechanism, HSV-1 infection leads to a down-regulation of MHC class I expression via binding of ICP47 to the transporter associated with antigen presentation (TAP), blocking the antigenic peptide transport in the endoplasmic reticulum and subsequent loading onto MHC class I molecules and presentation on the cell surface [23]. This attenuates the overall immune response to infection through attenuating CD8+ T cell recognition of infected cells [24]. Designing a multimutated oHSV and deleting the α47 gene prevents the down-regulation of MHC class I which would allow CD8+ T cells to recognize infected tumor cells and hinder viral immune-evasion. G47Δ is a multimutated oHSV variant built from G207, in which, in addition to the deletion of both copies of the γ34.5 gene, an inactivation of ICP6, it contains a deletion of the α47 gene [25]. G47Δ has shown significantly higher efficacy in vivo compared to its precursor G207 at inhibiting tumor growth in immune-competent and immune-deficient animal models [25].
2.2. Activation of Innate Immune Responses
Exploiting the immune system to fight cancer by activating the innate and adaptive immune pathways has been studied widely over the last years. Immunotherapy is found to be a promising approach for many forms of cancer in preclinical and clinical trials [19]. Such immunotherapies include chimeric antigen receptor T cell (CAR-T cell), bispecific killer cell engager (BiKE) [26] and oncolytic virotherapy [27]. Their most important mechanism is to activate immune cells such as T cells, natural killer (NK) cells and dendritic cells (DC). Oncolytic virus’ ability to selectively replicate inside tumor cells and stimulate innate immune cells could override the suppressive tumor microenvironment by inducing both antiviral and antitumor activity [20].
Oncolytic herpes simplex virus has been identified to induce a dual mechanism inside tumor cells. These mechanisms, as shown in Figure 1, involve the activation of antiviral pathways by triggering cell death signaling cascades and the induction of host antitumor immune responses by recruiting and activating the surrounding immune cells, which ultimately leads to tumor killing [20].
Recent research has shown that oncolytic viruses, particularly herpes simplex virus, induce cell death that elicits an immune response, identified as immunogenic cell death (ICD) [28]. The level of immunogenicity is associated with the activation of danger signaling pathways and the expression of damage-associated molecular patterns (DAMPs) in the tumor microenvironment [29]. One study has shown that the infection of oncolytic HSV-1 within squamous cell carcinoma (SCC) cells, in vitro, increased the release of ATP and high mobility group box 1 (HMGB1) compared to uninfected SCC. In addition, it induced the translocation of calreticulin (CRT) to the cell surface to act as an “eat me” signal [29].
During viral infection by oHSV, dying tumor cells consequentially release molecular components including ATP and HMGB1, in addition to the surface exposure CRT to prepare the cell for phagocytosis. The recognition of DAMPs by immature dendritic cells (DCs) facilitates their maturation and infiltration. Dendritic cells mature as they infiltrate into the tumor microenvironment in response to inflammatory chemokines such as CC-chemokine ligand 4 (CLL4) [28]. While DAMPs can prompt DCs maturation, the chemokine receptor expression and chemokine responsiveness can play an important part in DCs’ activation, cross-presentation, and migration to the draining lymph nodes [27]. The stimulation and recruitment of local macrophages and dendritic cells triggers them to engulf dying tumor cells and to process internalized tumor antigens to be presented to naive T cells. This results in priming the cytotoxic effect by T cells, and therefore leads to an antitumor response [28]. Additionally, the recognition of DAMPs and viral components by the innate sensing via pattern recognition receptors (PRRs) stimulates type I interferon production via stimulator interferon genes (STING) signaling pathway. In a recent study, it was found that the STING-dependent pathway has a critical role in the spontaneous priming of antitumor T cells [30]. This provides insights into cancer immunotherapy indications, which can be used for the development of current cancer immunotherapies.
2.3. Activation of Adaptive Immune Responses
The activation of T cells is a crucial part of a cell-mediated adaptive immune response. Sufficient activation of innate immunity and the cross-presentation, which is the process of presenting foreign antigens on MHC class I molecules, are essential for the activation of CD8+ T cells [20,31]. Activated CD8+ T cells clonally expand in secondary lymphoid organs such as the spleen and lymph nodes, and then, aided by chemokine gradients, migrate to the sites of inflammation [27,28,32]. As the tumor microenvironment (TME) suppresses CD8+ T cell trafficking and function due to multiple effects, oncolytic virotherapy offers the potential to overcome this by eliciting an adaptive immune response both against the viral infection and the tumor [33].
Tumors have evolved multiple immune-evasive mechanisms to interfere with the stimulation of a tumor-specific T cell response [34]. In oncolytic virotherapy, the initial T cell response may be instigated by oncolytic viral activity; however, the presence of tumor-associated antigens and tumor cell debris in the inflammatory milieu of the TME promotes the cross-presentation of tumor antigens to CD8+ T cells [28]. As certain oncolytic herpes simplex viruses are engineered to have a deletion in both copies of ICP47, which is a protein that interferes with the normal processing and presentation pathways of MHC class I, levels of MHC class I in infected cells are enhanced [19,35], which prevents viral inhibition of tumor-associated antigen processing and presentation [28]. In a study by Benencia et al., 2008, it was shown that ICP47-lacking oHSV significantly increases tumor antigen uptake by DCs, leading to an enhanced effect of tumor-associated NK and CD8+ T cells [36].
Oncolytic viruses can be engineered to express genes that enhance the immune response. Most commonly, oncolytic viruses may be armed with cytokines, which are immunomodulatory genes that play an essential role in the recruitment and homeostasis of T cells [28]. Cytokine can also sustain CD8+ T cell activation, contributing to an enhanced local and distant antitumor response [37]. In a phase II clinical trial using OncovexGM-CSF for melanoma, 26% of the patients with unresectable stage III or IV melanoma demonstrated objective clinical response, which included regression of infected and distant noninfected lesions [16].
As opposed to innate immunity, adaptive immunity induces immune memory, meaning that recurrent exposure to the same antigen will generate a stronger response [20]. Fully functional CD8+ T cells maintain ongoing tumor-specific surveillance against distant tumors or potential relapse [34]. As a result, when using oncolytic virotherapy, it is important to take antiviral memory into consideration, as it may hinder retreatment [20]. Oncolytic viruses may naturally induce an antiviral immune response, which may lead to clearance of the viruses prior to them performing their intended function [6]. Therefore, the balance between viral immunogenicity and antitumor immunity is crucial in determining the efficacy of oncolytic virotherapy [28].
3. oHSV Derivatives in Pre-Clinical Models
Various genetically engineered oHSVs have been developed for the treatment of different cancers [38]. Multiple approaches have been employed to enhance the effects of oHSVs [28]. Such approaches include arming oHSV with pro-drug metabolizing transgenes such as thymidine kinase [39], proapoptotic genes such as TNF-related apoptosis-inducing ligand (TRAIL) [40,41], or immunostimulatory transgenes such as GM-CSF [14,16]. Indeed, various preclinical studies have demonstrated the therapeutic efficacy of different oHSV variants in treating multiple types of tumors [38]. Table 1 summarizes several oHSV variants, their genetic modification and the preclinical models they have been tested on.
rRp450, an oHSV variant with an ICP6 deletion and an insertion of rat cytochrome P450 2B1 (CYP2B1), a pro-drug enzyme for cyclophosphamide (CPA), has initially shown therapeutic efficacy in treating preclinical animal models of glioblastoma [42]. In 2002, Pawlik et al. studied the efficacy of rRp450 in treating diffuse colon carcinoma liver metastases. The study showed that treatment with rRp450 and CPA significantly decreases the tumor burden from uncountable metastatic nodules in the control group to five in the treated group [43]. In an aggressive sarcoma mouse model, the administration of rRp450 and CPA significantly increased mouse survival compared to the control group [44]. In an orthotopic mouse model of atypical teratoid/rhabdoid tumors, treatment with rRp450 and CPA significantly prolonged the median survival to 84.5 days in tumor-bearing mice compared to 50 days in the vehicle-treated controls [57].
Another oHSV variant is rQNestin34.5v.2, which is an oHSV that retains ICP34.5 expression under the nestin promoter/enhancer elements [58]. This variant has demonstrated preclinical therapeutic efficacy for neurological tumors. Upon treatment with rQNestin34.5v.2, 77.8% of athymic mice bearing intracerebral human U87dEGFR glioma tumors survived >90 days, whereas mice injected with vehicle only survived up to day 21 post-tumor implantation [45].
Initial studies involving the second generation oHSV variant G207, which is genetically modified with a diploid deletion in γ34.5 and an inactivation in UL39, showed efficacious treatment of gliomas [46]. Combining G207 with fractionated ionizing radiation resulted in a synergistic action, as seen with reduced tumor sizes and prolonged survival in mice bearing high-grade gliomas [47]. In a xenogeneic flank mouse model of cervical cancer, a single injection of G207 led to the reduction in tumor burden by 50% [48].
Preclinical studies using G47Δ, which is a third generation oHSV carrying a γ34.5 diploid deletion, UL39 inactivation and an α47 deletion, demonstrated its efficacy and safety in different tumor models including brain [49], gastric [50], liver [51], breast [52], thyroid [51], and urological [52] cancers. In a study conducted by Nigim et al., mice bearing patient-derived malignant meningioma were treated with two injections of G47Δ. This treatment significantly prolonged survival, with 20% of mice surviving >160 days. Furthermore, the authors reported a lack of signs of encephalitic associated with G47Δ treatment, which confirms the safety of this treatment [49]. Several studies have tested the efficacy of G47Δ in treating types of breast cancer [52]. A study by J. Wang et al. reported a 9-fold reduction in the number of metastatic breast cancer nodules in the lungs of G47Δ-treated group compared to the control-treated group [59].
oHSV can be engineered to express proapoptotic or immunostimulatory genes to enhance the antitumor effect [28]. A G47Δ oHSV variant that expresses TRAIL was shown to significantly inhibit glioblastoma multiform tumor growth and invasion, and prolonged mice survival [41]. oHSV expressing immunostimulatory genes such as IL-12 or GM-CSF have been shown to have therapeutic efficacy via stimulating an antitumor immune response. For instance, IL12-expressing oHSV was shown to promote tumor-specific CD8+ T cell responses in the peritoneal cavity and omentum in ovarian cancer models [55]. Ghouse et al. reported that treatment of mice bearing triple-negative breast cancer with oHSV-IL12 resulted in the induction of local and abscopal immune effects, in addition to significantly prolonging the survival of treated mice compared to the control group [56]. GM-CSF-expressing oHSV have demonstrated tremendous potential in treating advanced melanoma, which was later approved by the FDA to treat stage III unresectable advanced melanoma [18].
4. oHSV Route of Delivery
4.1. Systemic Versus Local Delivery
Whether it is intratumoral or systemic administration, certain hurdles pose a challenge to oHSV delivery [60]. Although intratumoral injection delivers virus particles directly to the tumor, there are multiple limitations that prevent optimal delivery and spread within the tumor microenvironment. For instance, certain tumors may be comprised of several nodules located over a large area, or in an anatomical position inaccessible or inconvenient for local injection [61]. For example, the rates of response in phase I and Ib clinical trials using local injection of oHSV for glioblastoma have been suboptimal, potentially due to the influx of blood and cerebrospinal fluid into the cavity after surgical intervention and tumor resection, that resulting in washing out the virus [40]. Intravenous administration allows the virus to reach both primary and metastatic tumors simultaneously; however, rapid clearance of the viruses from the circulation before they reach their targets by antibodies, antiviral cytokines, and immune blood cells, in addition to nonspecific tissue, are among the main hurdles for systemic administration [62]. To overcome some of these challenges, researchers have explored the use of cell carriers to “cloak” the virus particles from the host’s defense. Some of the tested oHSV cell carriers include neural precursor cells [63], lymphocytes [64] and mesenchymal stem cells [65]. In the next section, we will explore the use of such carrier cells to deliver oHSV.
4.2. Mesenchymal Stem Cells as OV Carrier Cells
Mesenchymal stem cells (MSCs) have been explored as potential vehicles for gene therapy in tumors as they have demonstrated preferential integration into sites of tumor development [66,67]. The source of MSCs can be autologous, from patient’s bone marrow or adipose tissues, or allogeneic, from placenta and umbilical cord, which can be used as “off-the-shelf” cells [68]. Preclinical and clinical data indicate the safety of using MSCs, as there were no major health concerns reported, suggesting the relative safety of MSC therapies [69]. Another feature of MSCs is their low immunogenicity due to the lack of co-stimulatory molecules expression, eliminating the need for immunosuppression during allogenic transplantation [70]. All of these features make MSCs an excellent candidate to be used for oncolytic virus delivery to tumor sites.
MSCs have been extensively explored for their ability to “cloak” oncolytic viruses, providing them with effective protection from host neutralizing immunity and effective delivery to the tumor microenvironment [71]. Among the studied oncolytic viruses in conjunction with MSCs are Newcastle disease viruses [68], adenoviruses [72], measles viruses [73], and herpes simplex viruses [74,75]. oHSV has been frequently studied in conjunction with MSCs, and has shown promising results in treating gliomas, metastatic melanomas, breast and ovarian cancers, whether delivered systemically [74,75] or locally [40].
5. Updates on Recent Preclinical and Clinical oHSV Immunotherapies
At the beginning of the 1990s, genetically engineered oncolytic viruses were first reported in a pre-clinical mouse model showing selective antitumor effects [76]. Since then, different oHSV mutants have been established, including Oncovex GM-CSF (T-VEC), G207, HSV1716, HF10, NV1020 and G47Δ, to increase its safety and efficacy [11,17,77,78]. Many oHSV variants have entered either phase I, II, or III clinical trials or have completed them to treat different types and degrees of tumor, such as breast cancer, glioma and melanoma [79].
5.1. Talimogene Laherparepvec (T-VEC)
T-VEC is the first oncolytic virus to be FDA-approved for unresectable stage III advanced melanoma. It is a modified type 1 herpes simplex virus JS1 strain with ICP 34.5 genes deletion, reducing the neurovirulence of the virus and elevating the selectivity of infection for cancer cell killing [15,18,80]. Additionally, the ICP47 gene is deleted, allowing antigen presentation which increases the expression of US11, causing an increase in ICP 34.5-deleted HSV-1 replication without loss of its tumor specificity [81]. The addition of a human granulocyte–macrophage colony-stimulating factor (GM-CSF) permits local expression, increasing the activation of antigen-presenting cells, enhancing the antitumor immune response [15]. The precise mechanism is still unknown [82].
T-VEC is the only variant that has entered a phase III clinical trial, where it is administered in intralesional sites in advanced melanoma patients, causing a reduction in tumor growth and a systemic anti-tumor effect, prolonging survival rate [83]. Biopsies obtained from patients who received T-VEC showed a lower level of immune-suppressive cells such as CD4+ FoxP3+ regulatory T cells, CD8+ FoxP3+ T cells and myeloid-derived suppressor cells (MDSCs) in the TME compared to the untreated controls [84].
5.2. Clinical Trials of oHSV
oHSV has been studied extensively and tested in clinical setting with approximately 86 clinical trials to study and test for future clinical translation. Those clinical trials were registered at ClinicalTrials.gov and were accessed on February 13th, 2021. T-VEC is one of the most extensively investigated oHSV variants in clinical settings, as shown in Figure 2.
T-VEC is mostly studied as monotherapy or in combination with immune check inhibitors, radiotherapy, or chemotherapy to treat different types of cancer, although several investigate its effect on melanoma, due to the ease of lesion accessibility [85]. In a phase Ib trial of T-VEC in combination with immune check inhibitor ipilimumab (anti-CTLA4 antibody) for treating unresectable stage IIIB-IV melanoma, combination treatment showed higher efficacy compared to monotherapy, with a 50% objective response rate and no dose-limiting toxicities [86]. In this trial, durable response for over 6 months was seen in 44% of the patients. In a phase I trial assessing T-VEC in combination with pembrolizumab (anti-PD1 monoclonal antibody) for treating melanoma, 62% of the patients showed an objective response and a third exhibited an extraordinary complete response. A randomized phase III trial of this combination is currently under investigation [28]. Other oHSV variants such as G207, C134, and rQNestin34.5 are currently under clinical investigation for their safety and efficacy to treat high-grade glioma [87,88]. Table 2 summarizes most of the clinical trials employing such variants and their clinical studies.
RP1 is an oHSV variant modified to contain two deletions of γ34.5 and ICP47, and expression of GM-CSF and GALV-GP-R-, a fusogenic glycoprotein membrane isolated from gibbon ape leukemia virus [94,95]. Expression of GALV-GP-R- was shown to have tumor cell killing ability and an immunogenic response [95]. This variant is currently under evaluation as monotherapy for treating squamous cell carcinoma (NCT04349436), and in combination with PD-1 inhibitors such as nivolumab (NCT03767348) or cemiplimab (NCT04050436) [94].
ONCR-177 is an oHSV variant that contains mutations in the UL37 and ICP47 genes, which prevents replication and neuropathic activity in normal cells [96]. It is armed with five immune-modulatory agents, FLT3LG, IL-12, CCL4, anti-CLTA-4 and anti-PD-1, to increase T cell and NK activation in addition facilitating CD8+ T and dendritic cell recruitment [94,96]. ONCR-177 is in clinical trials to assess its safety and the preliminary anti-tumor efficacy as a monotherapy or in combination with pembrolizumab (anti-PD-1) for treating metastatic solid tumors (NCT04348916).
While most clinical trials investigate variants of oHSV-1, some clinical trials are investigating the safety and therapeutic efficacy of oncolytic variants of herpes simplex virus type-2 (HSV-2). OH2 is a potent oncolytic variant of HSV-2 with ICP34.5 and ICP47 deletion and human GM-CSF expression [79]. Certain variants of OH2 may include a deletion in the protein kinase (PK) domain of the ICP10 gene. The PK domain activates the Ras/MEK/MAPK mitogenic pathways, which facilitates HSV-2 replication. Deletion of this domain prevents HSV-2 replication in normal cells and restricts it to cells with aberrantly activated Ras pathways, such as tumor cells [97]. OH2 expressing GM-CSF has demonstrated significant therapeutic efficacy against metastatic ovarian cancer in preclinical models [98]. OH2 is currently undergoing clinical trials for treating different solid tumors in humans as a monotherapy (NCT04637698) or in combination with other drugs such as PD-1 inhibitor pembrolizumab (NCT04386967) or HX008 (NCT03866525 and NCT04616443).
6. Limitations
The route of delivery and the presence of pre-existing immunity are two important factors that may affect the efficiency of oHSV therapeutics. Delivery of oHSV is usually achieved by direct intertumoral injections or by locoregional means. However, these delivery routes are not suitable for several tumors, including metastatic cancers. In vivo studies with animal models found no correlation between pre-existing immunity and the therapeutic potential of oHSV [99,100]. For efficient systemic delivery of oHSV, the host is usually administered with immunosuppression agents [101]. Although administration of immunosuppression agents such as corticosteroid had no effect on the oncolytic viral activity, they inhibited the establishment of antitumor immunity [102]. Another important issue is the need for combined use of oHSV chemotherapy and/or radiotherapy. These two types of traditional therapy usually cause tissue necrosis and/or induce an inflammatory response that can limit virus spread [103].
Other important issues that should be considered while evaluating the safety profile of oHSV include: (i) the risk of reversion, (ii) latency, and (iii) modulation of the host immune response. Genetically modified variants of oHSV can restore their neurovirulence activity by recombination with WT HSV-1 strain in vivo [103]. The risk of acquiring neurovirulence will lead to a loss of specificity and the infection of healthy tissues. Latent infection is a feature for herpesviruses, and hence oHSV enter latency and reappear later on [104]. oHSVs usually carry one or more of the transgenes that act to modulate the host immune response. Transgenes are added to express several cytokines such as GM–CSF and IL-12, which may cause an inflammatory response against self-antigens and hence autoimmunity [103].
7. Conclusions and Perspectives
There has been a marked increase in oHSV research in the last two decades, as genetically modified HSV was the first oncolytic virus to be tested [19]. HSV, in particular, is an attractive agent for oncolytic virotherapy, as its genome is susceptible to modification, providing a promising platform to generate a safe and potent antitumor drug. The success or failure of oncolytic virotherapy is largely dependent on the interaction of antitumor and antiviral immune responses between host and virus [60]. In addition to lysing the tumor cells, oHSVs induce cytokine production and recruitment of immune cells to the tumor microenvironment and enhance antitumor immunity [20]. oHSV has shown efficacy in treating human tumors, and more clinical data support the beneficial role of oHSV therapy in combination with other cancer therapeutics, especially the immunotherapeutic agents [28].
Understanding the interactions between host immunity, the oncolytic viral infection and the tumor are essential for developing better strategies to combat cancer [34]. As oncolytic viral infections activate innate and adaptive immunity, the therapeutic efficacy largely depends on the balance between antiviral immunity, where the immune system clears out the viral infection, and antitumor immunity, where the immune system eliminates tumor cells [28]. The immune responses accompanying oncolytic virotherapy and unique to this system, and special consideration must be taken [34]. Early clearance of oHSV may be circumvented by taking the route and mode of delivery into consideration [61].
Effective delivery of such oncolytic viral agents to tumor sites remains a challenge [105]. Identifying the best route of delivery and vehicle is crucial to optimally engage the patient immune response to mediate effective antitumor immunity [106]. Improving engineered oncolytic viral constructs and testing combined immunotherapeutic agents, coupled with the “Trojan horse” concept, offer various therapeutic possibilities [71]. With more preclinical research going into clinical application, researchers are more likely to achieve more success in understanding the best combination of oHSV variants and other immunetherapeutics, as well as the most suitable route and vehicle for delivery that would ultimately help in winning the fight against cancer.
Funding
This work was supported by a grant from King Abdulaziz City for Science and Technology (KACST) through the Center of Excellence for Biomedicine (CEBM).
Conflicts of Interest
K.S. owns equity in and is a member of the Board of Directors of AMASA Therapeutics, a company developing stem cell-based therapies for cancer. K.S.’s interests were reviewed and are managed by Brigham and Women’s Hospital and Partners HealthCare in accordance with their conflict of interest policies. The other authors declare that they have no competing interests.
References
- Ma, X.; Yu, H. Global Burden of Cancer. Yale J. Biol. Med. 2006, 79, 85–94. [Google Scholar]
- Urruticoechea, A.; Alemany, R.; Balart, J.; Villanueva, A.; Vinals, F.; Capella, G. Recent Advances in Cancer Therapy: An Overview. Curr. Pharm. Des. 2010, 16, 3–10. [Google Scholar] [CrossRef]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Mavani, H.J.; Wick, J.Y. Oncology’s Trojan Horse: Using Viruses to Battle Cancer. Consult. Pharm. 2016, 31, 676–684. [Google Scholar] [CrossRef]
- Scholl, S.M.; Balloul, J.M.; Le Goc, G.; Bizouarne, N.; Schatz, C.; Kieny, M.P.; von Mensdorff-Pouilly, S.; Vincent-Salomon, A.; Deneux, L.; Tartour, E.; et al. Recombinant Vaccinia Virus Encoding Human MUC1 and IL2 as Immunotherapy in Patients with Breast Cancer. J. Immunother. 2000, 23, 570–580. [Google Scholar] [CrossRef]
- Kaufman, H.L.; Kohlhapp, F.J.; Zloza, A. Oncolytic Viruses: A New Class of Immunotherapy Drugs. Nat. Rev. Drug Discov. 2015, 14, 642–662. [Google Scholar] [CrossRef] [PubMed]
- Lawler, S.E.; Speranza, M.-C.; Cho, C.-F.; Chiocca, E.A. Oncolytic Viruses in Cancer Treatment: A Review. JAMA Oncol. 2017, 3, 841–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macdonald, S.J.; Mostafa, H.H.; Morrison, L.A.; Davido, D.J. Genome Sequence of Herpes Simplex Virus 1 Strain KOS. J. Virol. 2012, 86, 6371–6372. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Nemunaitis, J. Herpes Simplex Virus 1 (HSV-1) for Cancer Treatment. Cancer Gene Ther. 2006, 13, 975–992. [Google Scholar] [CrossRef] [Green Version]
- Geevarghese, S.K.; Geller, D.A.; de Haan, H.A.; Horer, M.; Knoll, A.E.; Mescheder, A.; Nemunaitis, J.; Reid, T.R.; Sze, D.Y.; Tanabe, K.K.; et al. Phase I/II Study of Oncolytic Herpes Simplex Virus NV1020 in Patients with Extensively Pretreated Refractory Colorectal Cancer Metastatic to the Liver. Hum. Gene Ther. 2010, 21, 1119–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markert, J.M.; Razdan, S.N.; Kuo, H.-C.; Cantor, A.; Knoll, A.; Karrasch, M.; Nabors, L.B.; Markiewicz, M.; Agee, B.S.; Coleman, J.M.; et al. A Phase 1 Trial of Oncolytic HSV-1, G207, given in Combination with Radiation for Recurrent GBM Demonstrates Safety and Radiographic Responses. Mol. Ther. 2014, 22, 1048–1055. [Google Scholar] [CrossRef] [Green Version]
- Rehman, H.; Silk, A.W.; Kane, M.P.; Kaufman, H.L. Into the Clinic: Talimogene Laherparepvec (T-VEC), a First-in-Class Intratumoral Oncolytic Viral Therapy. J. Immunother. Cancer 2016, 4, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streby, K.A.; Geller, J.I.; Currier, M.A.; Warren, P.S.; Racadio, J.M.; Towbin, A.J.; Vaughan, M.R.; Triplet, M.; Ott-Napier, K.; Dishman, D.J.; et al. Intratumoral Injection of HSV1716, an Oncolytic Herpes Virus, Is Safe and Shows Evidence of Immune Response and Viral Replication in Young Cancer Patients. Clin. Cancer Res. 2017, 23, 3566–3574. [Google Scholar] [CrossRef] [Green Version]
- Toda, M.; Martuza, R.L.; Rabkin, S.D. Tumor Growth Inhibition by Intratumoral Inoculation of Defective Herpes Simplex Virus Vectors Expressing Granulocyte-Macrophage Colony-Stimulating Factor. Mol. Ther. 2000, 2, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.L.; Robinson, M.; Han, Z.-Q.; Branston, R.H.; English, C.; Reay, P.; McGrath, Y.; Thomas, S.K.; Thornton, M.; Bullock, P.; et al. ICP34.5 Deleted Herpes Simplex Virus with Enhanced Oncolytic, Immune Stimulating, and Anti-Tumour Properties. Gene Ther. 2003, 10, 292–303. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, H.L.; Kim, D.W.; DeRaffele, G.; Mitcham, J.; Coffin, R.S.; Kim-Schulze, S. Local and Distant Immunity Induced by Intralesional Vaccination with an Oncolytic Herpes Virus Encoding GM-CSF in Patients with Stage IIIc and IV Melanoma. Ann. Surg. Oncol. 2010, 17, 718–730. [Google Scholar] [CrossRef]
- Hu, J.C.C.; Coffin, R.S.; Davis, C.J.; Graham, N.J.; Groves, N.; Guest, P.J.; Harrington, K.J.; James, N.D.; Love, C.A.; McNeish, I.; et al. A Phase I Study of OncoVEXGM-CSF, a Second-Generation Oncolytic Herpes Simplex Virus Expressing Granulocyte Macrophage Colony-Stimulating Factor. Clin. Cancer Res. 2006, 12, 6737–6747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conry, R.M.; Westbrook, B.; McKee, S.; Norwood, T.G. Talimogene Laherparepvec: First in Class Oncolytic Virotherapy. Hum. Vaccines Immunother. 2018, 14, 839–846. [Google Scholar] [CrossRef]
- Peters, C.; Rabkin, S.D. Designing Herpes Viruses as Oncolytics. Mol. Ther. Oncolytics 2015, 2, 15010. [Google Scholar] [CrossRef]
- Yin, J.; Markert, J.M.; Leavenworth, J.W. Modulation of the Intratumoral Immune Landscape by Oncolytic Herpes Simplex Virus Virotherapy. Front. Oncol. 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- He, B.; Gross, M.; Roizman, B. The γ134.5 protein of herpes simplex virus 1 complexes with protein phosphatase 1 α to dephosphorylate the α subunit of the eukaryotic translation initiation factor 2 and preclude the shutoff of protein synthesis by double-stranded RNA-activated protein kinase. Proc. Natl. Acad. Sci. USA 1997, 94, 843–848. [Google Scholar] [CrossRef] [Green Version]
- Aghi, M.; Visted, T.; DePinho, R.A.; Chiocca, E.A. Oncolytic Herpes Virus with Defective ICP6 Specifically Replicates in Quiescent Cells with Homozygous Genetic Mutations in P16. Oncogene 2008, 27, 4249–4254. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.; Jugovic, P.; York, I.; Russ, G.; Bennink, J.; Yewdell, J.; Ploegh, H.; Johnson, D. Herpes Simplex Virus Turns off the TAP to Evade Host Immunity. Nature 1995, 375, 411–415. [Google Scholar] [CrossRef]
- Orr, M.T.; Edelmann, K.H.; Vieira, J.; Corey, L.; Raulet, D.H.; Wilson, C.B. Inhibition of MHC Class I Is a Virulence Factor in Herpes Simplex Virus Infection of Mice. PLoS Pathog. 2005, 1, 62–71. [Google Scholar] [CrossRef]
- Todo, T.; Martuza, R.L.; Rabkin, S.D.; Johnson, P.A. Oncolytic Herpes Simplex Virus Vector with Enhanced MHC Class I Presentation and Tumor Cell Killing. Proc. Natl. Acad. Sci. USA 2001, 98, 6396–6401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Song, X.; Wang, Y.; Liu, F.; Wei, J. Combining Oncolytic Viruses with Cancer Immunotherapy: Establishing a New Generation of Cancer Treatment. Front. Immunol. 2020, 11, 683. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Quintanilla, J.; Seah, I.; Chua, M.; Shah, K. Oncolytic Viruses: Overcoming Translational Challenges. J. Clin. Investig. 2019, 129, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Bommareddy, P.K.; Shettigar, M.; Kaufman, H.L. Integrating Oncolytic Viruses in Combination Cancer Immunotherapy. Nat. Rev. Immunol. 2018, 18, 1–16. [Google Scholar] [CrossRef]
- Takasu, A.; Masui, A.; Hamada, M.; Imai, T.; Iwai, S.; Yura, Y. Immunogenic Cell Death by Oncolytic Herpes Simplex Virus Type 1 in Squamous Cell Carcinoma Cells. Cancer Gene Ther. 2016, 23, 107–113. [Google Scholar] [CrossRef]
- Woo, S.-R.; Fuertes, M.B.; Corrales, L.; Spranger, S.; Furdyna, M.J.; Leung, M.Y.K.; Duggan, R.; Wang, Y.; Barber, G.N.; Fitzgerald, K.A.; et al. STING-Dependent Cytosolic DNA Sensing Mediates Innate Immune Recognition of Immunogenic Tumors. Immunity 2014, 41, 830–842. [Google Scholar] [CrossRef] [Green Version]
- Joffre, O.P.; Segura, E.; Savina, A.; Amigorena, S. Cross-Presentation by Dendritic Cells. Nat. Rev. Immunol. 2012, 12, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Bevan, M.J. CD8(+) T Cells: Foot Soldiers of the Immune System. Immunity 2011, 35, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellone, M.; Calcinotto, A. Ways to Enhance Lymphocyte Trafficking into Tumors and Fitness of Tumor Infiltrating Lymphocytes. Front. Oncol. 2013, 3, 231. [Google Scholar] [CrossRef] [Green Version]
- Gujar, S.A.; Lee, P.W.K. Oncolytic Virus-Mediated Reversal of Impaired Tumor Antigen Presentation. Front. Oncol. 2014, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Verweij, M.C.; Horst, D.; Griffin, B.D.; Luteijn, R.D.; Davison, A.J.; Ressing, M.E.; Wiertz, E.J.H.J. Viral Inhibition of the Transporter Associated with Antigen Processing (TAP): A Striking Example of Functional Convergent Evolution. PLoS Pathog. 2015, 11, e1004743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benencia, F.; Courrèges, M.C.; Fraser, N.W.; Coukos, G. Herpes Virus Oncolytic Therapy Reverses Tumor Immune Dysfunction and Facilitates Tumor Antigen Presentation. Cancer Biol. Ther. 2008, 7, 1194–1205. [Google Scholar] [CrossRef] [Green Version]
- Totsch, S.K.; Schlappi, C.; Kang, K.-D.; Ishizuka, A.S.; Lynn, G.M.; Fox, B.; Beierle, E.A.; Whitley, R.J.; Markert, J.M.; Gillespie, G.Y.; et al. Oncolytic Herpes Simplex Virus Immunotherapy for Brain Tumors: Current Pitfalls and Emerging Strategies to Overcome Therapeutic Resistance. Oncogene 2019, 38, 6159–6171. [Google Scholar] [CrossRef]
- Speranza, M.C.; Kasai, K.; Lawler, S.E. Preclinical Mouse Models for Analysis of the Therapeutic Potential of Engineered Oncolytic Herpes Viruses. ILAR J. 2016, 57, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Redaelli, M.; Franceschi, V.; Capocefalo, A.; Davella, D.; Denaro, L.; Cavirani, S.; Mucignat-Caretta, C.; Donofrio, G. Herpes Simplex Virus Type 1 Thymidine Kinasearmed Bovine Herpesvirus Type 4based Vector Displays Enhanced Oncolytic Properties in Immunocompetent Orthotopic Syngenic Mouse and Rat Glioma Models. Neuro-Oncology 2012, 14, 288–301. [Google Scholar] [CrossRef]
- Duebgen, M.; Martinez-Quintanilla, J.; Tamura, K.; Hingtgen, S.; Redjal, N.; Wakimoto, H.; Shah, K. Stem Cells Loaded with Multimechanistic Oncolytic Herpes Simplex Virus Variants for Brain Tumor Therapy. J. Natl. Cancer Inst. 2014, 106, dju090. [Google Scholar] [CrossRef]
- Tamura, K.; Wakimoto, H.; Agarwal, A.S.; Rabkin, S.D.; Bhere, D.; Martuza, R.L.; Kuroda, T.; Kasmieh, R.; Shah, K. Multimechanistic Tumor Targeted Oncolytic Virus Overcomes Resistance in Brain Tumors. Mol. Ther. 2013, 21, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Chase, M.; Chung, R.Y.; Antonio Chiocca, E. An Oncolytic Viral Mutant That Delivers the CYP2B1 Transgene and Augments Cyclophosphamide Chemotherapy. Nat. Biotechnol. 1998, 16, 444–448. [Google Scholar] [CrossRef]
- Pawlik, T.M.; Nakamura, H.; Mullen, J.T.; Kasuya, H.; Yoon, S.S.; Chandrasekhar, S.; Chiocca, E.A.; Tanabe, K.K. Prodrug Bioactivation and Oncolysis of Diffuse Liver Metastases by a Herpes Simplex Virus 1 Mutant That Expresses the CYP2B1 Transgene. Cancer 2002, 95, 1171–1181. [Google Scholar] [CrossRef]
- Currier, M.A.; Gillespie, R.A.; Sawtell, N.M.; Mahller, Y.Y.; Stroup, G.; Collins, M.H.; Kambara, H.; Chiocca, E.A.; Cripe, T.P. Efficacy and Safety of the Oncolytic Herpes Simplex Virus RRp450 Alone and Combined with Cyclophosphamide. Mol. Ther. 2008, 16, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Kambara, H.; Okano, H.; Chiocca, E.A.; Saeki, Y. An Oncolytic HSV-1 Mutant Expressing ICP34.5 under Control of a Nestin Promoter Increases Survival of Animals Even When Symptomatic from a Brain Tumor. Cancer Res. 2005, 65, 2832–2839. [Google Scholar] [CrossRef] [Green Version]
- Mineta, T.; Rabkin, S.D.; Yazaki, T.; Hunter, W.D.; Martuza, R.L. Attenuated Multi-Mutated Herpes Simplex Virus-1 for the Treatment of Malignant Gliomas. Nat. Med. 1995, 1, 938–943. [Google Scholar] [CrossRef]
- Bradley, J.D.; Kataoka, Y.; Advani, S.; Chung, S.M.; Arani, R.B.; Gillespie, G.Y.; Whitley, R.J.; Markert, J.M.; Roizman, B.; Weichselbaum, R.R. Ionizing Radiation Improves Survival in Mice Bearing Intracranial High-Grade Gliomas Injected with Genetically Modified Herpes Simplex Virus. Clin. Cancer Res. 1999, 5, 1517–1522. [Google Scholar]
- Blank, S.V.; Rubin, S.C.; Coukos, G.; Amin, K.M.; Albelda, S.M.; Molnar-Kimber, K.L. Replication-Selective Herpes Simplex Virus Type 1 Mutant Therapy of Cervical Cancer Is Enhanced by Low-Dose Radiation. Hum. Gene Ther. 2002, 13, 627–639. [Google Scholar] [CrossRef]
- Nigim, F.; Esaki, S.I.; Hood, M.; Lelic, N.; James, M.F.; Ramesh, V.; Stemmer-Rachamimov, A.; Cahill, D.P.; Brastianos, P.K.; Rabkin, S.D.; et al. A New Patient-Derived Orthotopic Malignant Meningioma Model Treated with Oncolytic Herpes Simplex Virus. Neuro-Oncology 2016, 18, 1278–1287. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Iwai, M.; Yajima, S.; Tanaka, M.; Yanagihara, K.; Seto, Y.; Todo, T. Efficacy of a Third-Generation Oncolytic Herpes Virus G47Δ in Advanced Stage Models of Human Gastric Cancer. Mol. Ther. Oncolytics 2020, 17, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, L.; Zeng, W.; Hu, P.; Zeng, M.; Rabkin, S.D.; Liu, R. Treatment of Human Hepatocellular Carcinoma by the Oncolytic Herpes Simplex Virus G47delta. Cancer Cell Int. 2014, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Bryan, S.M.; Mathis, J.M. Oncolytic Virotherapy for Breast Cancer Treatment. Curr. Gene Ther. 2018, 18, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.N.; Xu, L.H.; Zeng, W.G.; Hu, P.; Rabkin, S.D.; Liu, R.R. Treatment of Human Thyroid Carcinoma Cells with the G47delta Oncolytic Herpes Simplex Virus. Asian Pac. J. Cancer Prev. 2015, 16, 1241–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taguchi, S.; Fukuhara, H.; Homma, Y.; Todo, T. Current Status of Clinical Trials Assessing Oncolytic Virus Therapy for Urological Cancers. Int. J. Urol. 2017, 24, 342–351. [Google Scholar] [CrossRef] [Green Version]
- Thomas, E.D.; Meza-Perez, S.; Bevis, K.S.; Randall, T.D.; Gillespie, G.Y.; Langford, C.; Alvarez, R.D. IL-12 Expressing Oncolytic Herpes Simplex Virus Promotes Anti-Tumor Activity and Immunologic Control of Metastatic Ovarian Cancer in Mice. J. Ovarian Res. 2016, 9, 70. [Google Scholar] [CrossRef]
- Ghouse, S.M.; Nguyen, H.M.; Bommareddy, P.K.; Guz-Montgomery, K.; Saha, D. Oncolytic Herpes Simplex Virus Encoding IL12 Controls Triple-Negative Breast Cancer Growth and Metastasis. Front. Oncol. 2020, 10, 384. [Google Scholar] [CrossRef] [Green Version]
- Studebaker, A.W.; Hutzen, B.J.; Pierson, C.R.; Haworth, K.B.; Cripe, T.P.; Jackson, E.M.; Leonard, J.R. Oncolytic Herpes Virus RRp450 Shows Efficacy in Orthotopic Xenograft Group 3/4 Medulloblastomas and Atypical Teratoid/Rhabdoid Tumors. Mol. Ther. Oncolytics 2017, 6, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Chiocca, E.A.; Nakashima, H.; Kasai, K.; Fernandez, S.A.; Oglesbee, M. Preclinical Toxicology of RQNestin34.5v.2: An Oncolytic Herpes Virus with Transcriptional Regulation of the ICP34.5 Neurovirulence Gene. Mol. Ther. Methods Clin. Dev. 2020, 17, 871–893. [Google Scholar] [CrossRef]
- Wang, J.; Hu, P.; Zeng, M.; Rabkin, S.D.; Liu, R. Oncolytic Herpes Simplex Virus Treatment of Metastatic Breast Cancer. Int. J. Oncol. 2012, 40, 757–763. [Google Scholar] [CrossRef]
- Ma, W.; He, H.; Wang, H. Oncolytic Herpes Simplex Virus and Immunotherapy. BMC Immunol. 2018, 19, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, J.; Roy, D. Cell Carriers for Oncolytic Viruses: Current Challenges and Future Directions. Oncolytic Virother. 2013, 2, 47–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, M.S.; Lemoine, N.R.; Wang, Y. Systemic Delivery of Oncolytic Viruses: Hopes and Hurdles. Adv. Virol. 2012, 2012, 805629. [Google Scholar] [CrossRef] [PubMed]
- Herrlinger, U.; Woiciechowski, C.; Sena-Esteves, M.; Aboody, K.S.; Jacobs, A.H.; Rainov, N.G.; Snyder, E.Y.; Breakefield, X.O. Neural Precursor Cells for Delivery of Replication-Conditional HSV-1 Vectors to Intracerebral Gliomas. Mol. Ther. 2000, 1, 347–357. [Google Scholar] [CrossRef]
- Kanzaki, A.; Kasuya, H.; Yamamura, K.; Sahin, T.T.; Nomura, N.; Shikano, T.; Shirota, T.; Tan, G.; Fukuda, S.; Misawa, M.; et al. Efficacy of Oncolytic Herpes Simplex Virus Adsorbed onto Antigen-Specific Lymphocytes. Cancer Gene Ther. 2012, 19, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Parker Kerrigan, B.C.; Shimizu, Y.; Andreeff, M.; Lang, F.F. Mesenchymal Stromal Cells for the Delivery of Oncolytic Viruses in Gliomas. Cytotherapy 2017, 19, 445–457. [Google Scholar] [CrossRef] [Green Version]
- Sonabend, A.M.; Ulasov, I.V.; Tyler, M.A.; Rivera, A.A.; Mathis, J.M.; Lesniak, M.S. Mesenchymal Stem Cells Effectively Deliver an Oncolytic Adenovirus to Intracranial Glioma. Stem Cells 2008, 26, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Kidd, S.; Spaeth, E.; Dembinski, J.L.; Dietrich, M.; Watson, K.; Klopp, A.; Battula, V.L.; Weil, M.; Andreeff, M.; Marini, F.C. Direct Evidence of Mesenchymal Stem Cell Tropism for Tumor and Wounding Microenvironments Using in Vivo Bioluminescent Imaging. Stem Cells 2009, 27, 2614–2623. [Google Scholar] [CrossRef] [Green Version]
- Kazimirsky, G.; Jiang, W.; Slavin, S.; Ziv-Av, A.; Brodie, C. Mesenchymal Stem Cells Enhance the Oncolytic Effect of Newcastle Disease Virus in Glioma Cells and Glioma Stem Cells via the Secretion of TRAIL. Stem Cell Res. Ther. 2016, 7, 149. [Google Scholar] [CrossRef] [Green Version]
- Herberts, C.A.; Kwa, M.S.G.; Hermsen, H.P.H. Risk Factors in the Development of Stem Cell Therapy. J. Transl. Med. 2011, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- Chulpanova, D.S.; Kitaeva, K.V.; Tazetdinova, L.G.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Application of Mesenchymal Stem Cells for Therapeutic Agent Delivery in Anti-Tumor Treatment. Front Pharmacol. 2018, 9, 259. [Google Scholar] [CrossRef]
- Hadryś, A.; Sochanik, A.; McFadden, G.; Jazowiecka-Rakus, J. Mesenchymal Stem Cells as Carriers for Systemic Delivery of Oncolytic Viruses. Eur. J. Pharmacol. 2020, 874, 172991. [Google Scholar] [CrossRef] [PubMed]
- Hakkarainen, T.; Särkioja, M.; Lehenkari, P.; Miettinen, S.; Ylikomi, T.; Suuronen, R.; Desmond, R.A.; Kanerva, A.; Hemminki, A. Human Mesenchymal Stem Cells Lack Tumor Tropism but Enhance the Antitumor Activity of Oncolytic Adenoviruses in Orthotopic Lung and Breast Tumors. Hum. Gene Ther. 2007, 18, 627–641. [Google Scholar] [CrossRef]
- Ong, H.-T.; Federspiel, M.J.; Guo, C.M.; Ooi, L.L.; Russell, S.J.; Peng, K.-W.; Hui, K.M. Systemically Delivered Measles Virus-Infected Mesenchymal Stem Cells Can Evade Host Immunity to Inhibit Liver Cancer Growth. J. Hepatol. 2013, 59, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Seah, I.; Bougazzoul, O.; Choi, G.; Meeth, K.; Bosenberg, M.W.; Wakimoto, H.; Fisher, D.; Shah, K. Stem Cell-Released Oncolytic Herpes Simplex Virus Has Therapeutic Efficacy in Brain Metastatic Melanomas. Proc. Natl. Acad. Sci. USA 2017, 114, E6157–E6165. [Google Scholar] [CrossRef] [Green Version]
- Leoni, V.; Gatta, V.; Palladini, A.; Nicoletti, G.; Ranieri, D.; Dall’Ora, M.; Grosso, V.; Rossi, M.; Alviano, F.; Bonsi, L.; et al. Systemic Delivery of HER2-Retargeted Oncolytic-HSV By Mesenchymal Stromal Cells Protects From Lung And Brain Metastases. Oncotarget 2015, 6, 34774–34787. [Google Scholar] [CrossRef] [Green Version]
- Martuza, R.L.; Malick, A.; Markert, J.M.; Ruffner, K.L.; Coen, D.M. Experimental Therapy of Human Glioma by Means of a Genetically Engineered Virus Mutant. Science 1991, 252, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Kanai, R.; Zaupa, C.; Sgubin, D.; Antoszczyk, S.J.; Martuza, R.L.; Wakimoto, H.; Rabkin, S.D. Effect of Γ34.5 Deletions on Oncolytic Herpes Simplex Virus Activity in Brain Tumors. J. Virol. 2012, 86, 4420–4431. [Google Scholar] [CrossRef] [Green Version]
- Israyelyan, A.; Chouljenko, V.N.; Baghian, A.; David, A.T.; Kearney, M.T.; Kousoulas, K.G. Herpes Simplex Virus Type-1(HSV-1) Oncolytic and Highly Fusogenic Mutants Carrying the NV1020 Genomic Deletion Effectively Inhibit Primary and Metastatic Tumors in Mice. Virol. J. 2008, 5, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Jin, J.; Wu, Z.; Hu, S.; Hu, H.; Ning, Z.; Li, Y.; Dong, Y.; Zou, J.; Mao, Z.; et al. Stability and Anti-Tumor Effect of Oncolytic Herpes Simplex Virus Type 2. Oncotarget 2018, 9, 24672–24683. [Google Scholar] [CrossRef] [Green Version]
- Roizman, B. The Function of Herpes Simplex Virus Genes: A Primer for Genetic Engineering of Novel Vectors. Proc. Natl. Acad. Sci. USA 1996, 93, 11307–11312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, S.S.; Hecht, J.R.; Chan, E. Talimogene Laherparepvec: Review of Its Mechanism of Action and Clinical Efficacy and Safety. Immunotherapy 2019, 11, 705–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IMLYGIC (Talimogene Laherparepvec) | FDA. Available online: https://www.fda.gov/vaccines-blood-biologics/cellular-gene-therapy-products/imlygic-talimogene-laherparepvec (accessed on 23 February 2021).
- Andtbacka, R.H.I.; Ross, M.; Puzanov, I.; Milhem, M.; Collichio, F.; Delman, K.A.; Amatruda, T.; Zager, J.S.; Cranmer, L.; Hsueh, E.; et al. Patterns of Clinical Response with Talimogene Laherparepvec (T-VEC) in Patients with Melanoma Treated in the OPTiM Phase III Clinical Trial. Ann. Surg. Oncol. 2016, 23, 4169–4177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigg, C.; Blake, Z.; Gartrell, R.; Sacher, A.; Taback, B.; Saenger, Y. Talimogene Laherparepvec (T-Vec) for the Treatment of Melanoma and Other Cancers. Semin. Oncol. 2016, 43, 638–646. [Google Scholar] [CrossRef]
- Andtbacka, R.H.I.; Kaufman, H.L.; Collichio, F.; Amatruda, T.; Senzer, N.; Chesney, J.; Delman, K.A.; Spitler, L.E.; Puzanov, I.; Agarwala, S.S.; et al. Talimogene Laherparepvec Improves Durable Response Rate in Patients with Advanced Melanoma. J. Clin. Oncol. 2015, 33, 2780–2788. [Google Scholar] [CrossRef]
- Puzanov, I.; Milhem, M.M.; Minor, D.; Hamid, O.; Li, A.; Chen, L.; Chastain, M.; Gorski, K.S.; Anderson, A.; Chou, J.; et al. Talimogene Laherparepvec in Combination with Ipilimumab in Previously Untreated, Unresectable Stage IIIB-IV Melanoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 2619–2626. [Google Scholar] [CrossRef] [Green Version]
- Andtbacka, R.H.I.; Amatruda, T.; Nemunaitis, J.; Zager, J.S.; Walker, J.; Chesney, J.A.; Liu, K.; Hsu, C.-P.; Pickett, C.A.; Mehnert, J.M. Biodistribution, Shedding, and Transmissibility of the Oncolytic Virus Talimogene Laherparepvec in Patients with Melanoma. EBioMedicine 2019, 47, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Hua, L.; Wakimoto, H. Oncolytic Herpes Simplex Virus Therapy for Malignant Glioma: Current Approaches to Successful Clinical Application. Expert Opin. Biol. Ther. 2019, 19, 845–854. [Google Scholar] [CrossRef] [PubMed]
- Andtbacka, R.H.I.; Collichio, F.; Harrington, K.J.; Middleton, M.R.; Downey, G.; Öhrling, K.; Kaufman, H.L. Final Analyses of OPTiM: A Randomized Phase III Trial of Talimogene Laherparepvec versus Granulocyte-Macrophage Colony-Stimulating Factor in Unresectable Stage III-IV Melanoma. J. Immunother. Cancer 2019, 7, 145. [Google Scholar] [CrossRef] [Green Version]
- Chesney, J.; Puzanov, I.; Collichio, F.; Milhem, M.M.; Hauschild, A.; Chen, L.; Sharma, A.; Garbe, C.; Singh, P.; Mehnert, J.M. Patterns of Response with Talimogene Laherparepvec in Combination with Ipilimumab or Ipilimumab Alone in Metastatic Unresectable Melanoma. Br. J. Cancer 2019, 121, 417–420. [Google Scholar] [CrossRef]
- Harrington, K.J.; Kong, A.; Mach, N.; Chesney, J.A.; Fernandez, B.C.; Rischin, D.; Cohen, E.E.W.; Radcliffe, H.-S.; Gumuscu, B.; Cheng, J.; et al. Talimogene Laherparepvec and Pembrolizumab in Recurrent or Metastatic Squamous Cell Carcinoma of the Head and Neck (MASTERKEY-232): A Multicenter, Phase 1b Study. Clin Cancer Res. 2020, 26, 5153–5161. [Google Scholar] [CrossRef]
- Streby, K.A.; Currier, M.A.; Triplet, M.; Ott, K.; Dishman, D.J.; Vaughan, M.R.; Ranalli, M.A.; Setty, B.; Skeens, M.A.; Whiteside, S.; et al. First-in-Human Intravenous Seprehvir in Young Cancer Patients: A Phase 1 Clinical Trial. Mol. Ther. 2019, 27, 1930–1938. [Google Scholar] [CrossRef] [Green Version]
- Sze, D.Y.; Iagaru, A.H.; Gambhir, S.S.; De Haan, H.A.; Reid, T.R. Response to Intra-Arterial Oncolytic Virotherapy with the Herpes Virus NV1020 Evaluated by [18F]Fluorodeoxyglucose Positron Emission Tomography and Computed Tomography. Hum. Gene Ther. 2012, 23, 91–97. [Google Scholar] [CrossRef]
- Koch, M.S.; Lawler, S.E.; Chiocca, E.A. HSV-1 Oncolytic Viruses from Bench to Bedside: An Overview of Current Clinical Trials. Cancers 2020, 12, 3514. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Kuncheria, L.; Roulstone, V.; Kyula, J.N.; Mansfield, D.; Bommareddy, P.K.; Smith, H.; Kaufman, H.L.; Harrington, K.J.; Coffin, R.S. Development of a New Fusion-Enhanced Oncolytic Immunotherapy Platform Based on Herpes Simplex Virus Type 1. J. Immunother. Cancer 2019, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Haines, B.B.; Denslow, A.; Grzesik, P.; Lee, J.S.; Farkaly, T.; Hewett, J.; Wambua, D.; Kong, L.; Behera, P.; Jacques, J.; et al. ONCR-177, an Oncolytic HSV-1 Designed to Potently Activate Systemic Antitumor Immunity. Cancer Immunol. Res. 2021, 9, 291–308. [Google Scholar] [CrossRef]
- Fu, X.; Tao, L.; Cai, R.; Prigge, J.; Zhang, X. A Mutant Type 2 Herpes Simplex Virus Deleted for the Protein Kinase Domain of the ICP10 Gene Is a Potent Oncolytic Virus. Mol. Ther. 2006, 13, 882–890. [Google Scholar] [CrossRef]
- Fu, X.; Tao, L.; Zhang, X. An Oncolytic Virus Derived from Type 2 Herpes Simplex Virus Has Potent Therapeutic Effect against Metastatic Ovarian Cancer. Cancer Gene Ther. 2007, 14, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Lambright, E.S.; Kang, E.H.; Force, S.; Lanuti, M.; Caparrelli, D.; Kaiser, L.R.; Albelda, S.M.; Molnar-Kimber, K.L. Effect of Preexisting Anti-Herpes Immunity on the Efficacy of Herpes Simplex Viral Therapy in a Murine Intraperitoneal Tumor Model. Mol. Ther. 2000, 2, 387–393. [Google Scholar] [CrossRef]
- Delman, K.A.; Bennett, J.J.; Zager, J.S.; Burt, B.M.; McAuliffe, P.F.; Petrowsky, H.; Kooby, D.A.; Hawkins, W.G.; Horsburgh, B.C.; Johnson, P.; et al. Effects of Preexisting Immunity on the Response to Herpes Simplex-Based Oncolytic Viral Therapy. Hum. Gene Ther. 2000, 11, 2465–2472. [Google Scholar] [CrossRef]
- Ikeda, K.; Ichikawa, T.; Wakimoto, H.; Silver, J.S.; Deisboeck, T.S.; Finkelstein, D.; Harsh, G.R., IV; Louis, D.N.; Bartus, R.T.; Hochberg, F.H.; et al. Oncolytic Virus Therapy of Multiple Tumors in the Brain Requires Suppression of Innate and Elicited Antiviral Responses. Nat. Med. 1999, 5, 881–887. [Google Scholar] [CrossRef]
- Todo, T.; Rabkin, S.D.; Chahlavi, A.; Martuza, R.L. Corticosteroid Administration Does Not Affect Viral Oncolytic Activity, but Inhibits Antitumor Immunity in Replication-Competent Herpes Simplex Virus Tumor Therapy. Hum. Gene Ther. 1999, 10, 2869–2878. [Google Scholar] [CrossRef] [PubMed]
- Varghese, S.; Rabkin, S.D. Oncolytic Herpes Simplex Virus Vectors for Cancer Virotherapy. Cancer Gene Ther. 2002, 9, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Lou, E. Oncolytic Herpes Viruses as a Potential Mechanism for Cancer Therapy. Acta Oncol. 2003, 42, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Eissa, I.R.; Bustos-Villalobos, I.; Ichinose, T.; Matsumura, S.; Naoe, Y.; Miyajima, N.; Morimoto, D.; Mukoyama, N.; Zhiwen, W.; Tanaka, M.; et al. The Current Status and Future Prospects of Oncolytic Viruses in Clinical Trials against Melanoma, Glioma, Pancreatic, and Breast Cancers. Cancers 2018, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.; McFadden, G. Viruses for Tumor Therapy. Cell Host Microbe 2014, 15, 260–265. [Google Scholar] [CrossRef] [Green Version]
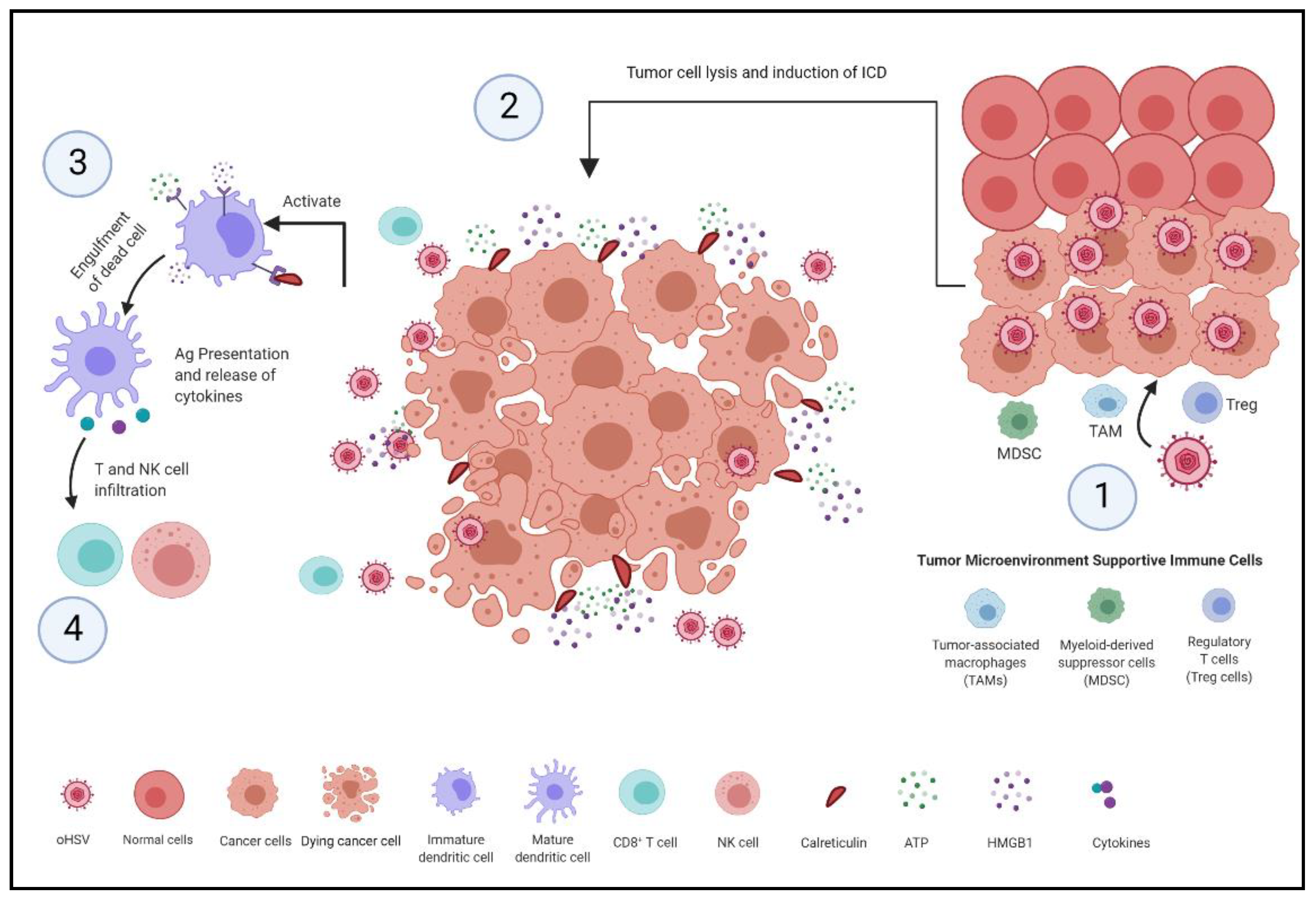
Figure 1.
Mechanism of inducing antitumor immunity. (1) oHSV selectively infects and replicates in tumor cells. (2) Infected tumor cells undergo cell lysis and immunogenic cell death (ICD) [28]. The ICD process involves alterations in composition of the cell surface, such as expression of calreticulin (CRT), and the sequential release of damage-associated molecular patterns (DAMPs) such as ATP and high mobility group protein B1 (HMGB1) [20,28]. (3) These alterations and released mediators recruit and interact with receptors expressed on DCs, which recruits them to the tumor site to engulf, process and present tumor antigens [20]. (4) DCs secrete pro-inflammatory cytokines that stimulate natural killer cells and tumor-specific CD8+ T cells, thereby inducing immune-mediated tumor cell killing [20].
Figure 1.
Mechanism of inducing antitumor immunity. (1) oHSV selectively infects and replicates in tumor cells. (2) Infected tumor cells undergo cell lysis and immunogenic cell death (ICD) [28]. The ICD process involves alterations in composition of the cell surface, such as expression of calreticulin (CRT), and the sequential release of damage-associated molecular patterns (DAMPs) such as ATP and high mobility group protein B1 (HMGB1) [20,28]. (3) These alterations and released mediators recruit and interact with receptors expressed on DCs, which recruits them to the tumor site to engulf, process and present tumor antigens [20]. (4) DCs secrete pro-inflammatory cytokines that stimulate natural killer cells and tumor-specific CD8+ T cells, thereby inducing immune-mediated tumor cell killing [20].

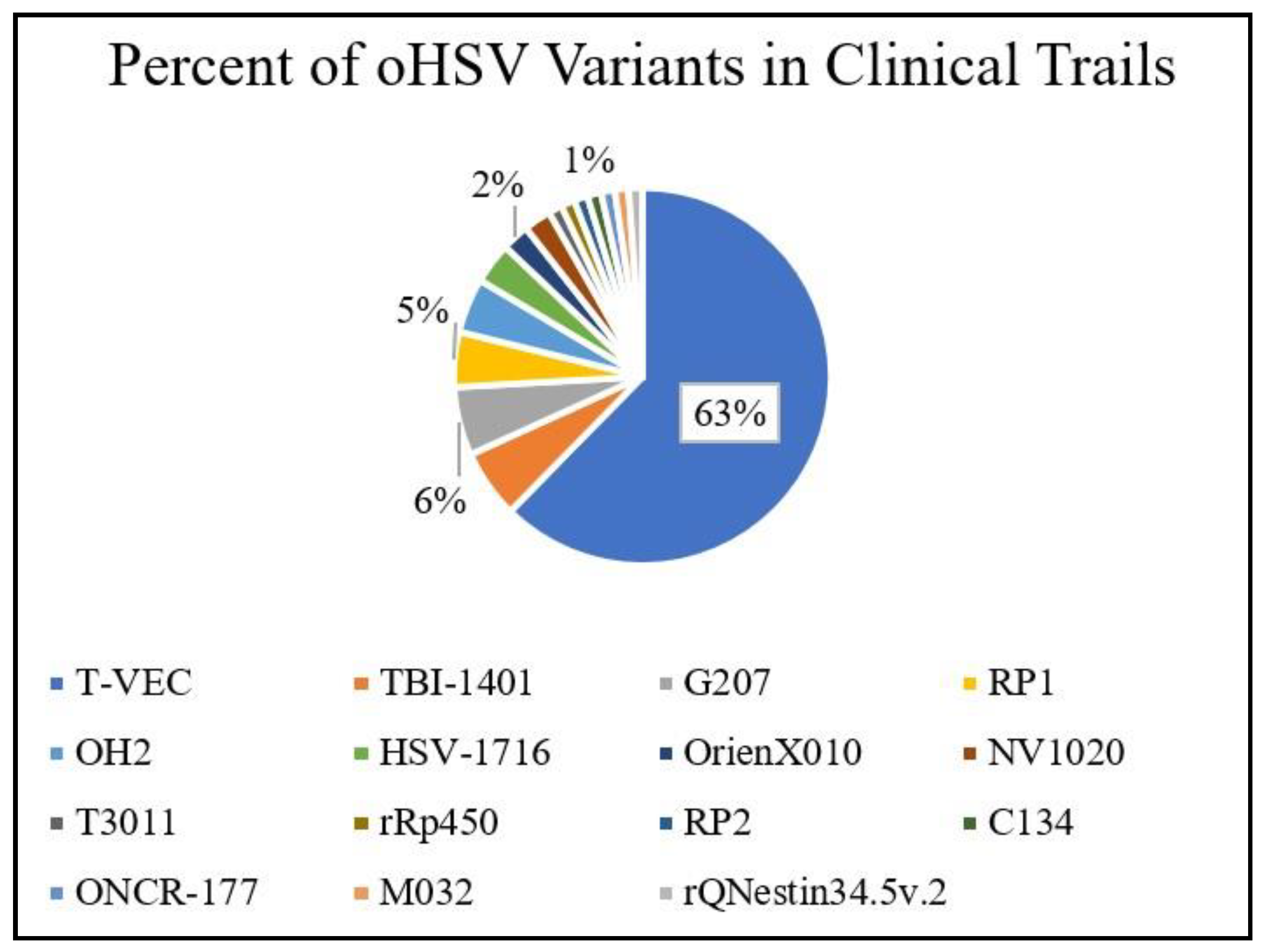
Figure 2.
A summary of clinical trials registered at ClinicalTrials.gov involving oHSV variants. Access date: 13 February 2021.
Figure 2.
A summary of clinical trials registered at ClinicalTrials.gov involving oHSV variants. Access date: 13 February 2021.


{kind=link}
{kind=link}
Table 1.
A summary of oHSV variants and their applications in preclinical models.
| oHSV Variant | Genetic Changes | Preclinical Models |
|---|---|---|
| rRp450 | ICP6 deletion, insertion of CYP2B1. | Glioblasoma [42] Colon carcinoma [43] Sarcoma [44] |
| rQNestin34.5v.2 | Nestin promoter drives the expression of ICP34.5. | Glioma [45] |
| G207 | Diploid deletion of γ34.5, inactivation of UL39. | Glioma [46,47] Cervical cancer [48] |
| G47Δ | Diploid deletion of γ34.5, inactivation of UL39, deletion of α47. | Meningioma [49] Gastic cancer [50] Liver cancer [51] Breast cancer [52] Thyroid carcinoma [53] Urological cancers [54] |
| G47Δ-TRAIL | G47Δ expressing TRAIL | Glioblastoma [41] |
| oHSV-IL12 | G207 expressing IL12 | Ovarian cancer [55] Breast cancer [56] |
| oHSV-GM-CSF | G207 expressing GM-CSF | Advanced melanoma [18] |
Table 2.
A summary of clinical trials registered at ClinicalTrials.gov involving oHSV variants. Access date: 13 February 2021.
Table 2.
A summary of clinical trials registered at ClinicalTrials.gov involving oHSV variants. Access date: 13 February 2021.
| Variant | Combination Therapy | Clinical Trial Number | Status | Virus | Malignant Type | Phase | Year Posted |
|---|---|---|---|---|---|---|---|
| T-VEC | - | NCT00289016 | Completed | HSV-1 | Stage IIIC and IV melanoma | II | 2006 |
| - | NCT00289016 | Completed | HSV-1 | Stage IIIC and IV melanoma | II | 2006 | |
| - | NCT00769704 | Completed [89] | HSV-1 | Melanoma | III | 2008 | |
| Cisplatin + Radiation | NCT01161498 | Terminated | HSV-1 | Head and neck cancer | III | 2010 | |
| GM-CSF | NCT01368276 | Completed | HSV-1 | Melanoma | III | 2011 | |
| Ipilimumab | NCT01740297 | Active, not recruiting [90] | HSV-1 | Melanoma | I, II | 2012 | |
| - | NCT02014441 | Completed [87] | HSV-1 | Melanoma | II | 2013 | |
| Pembrolizumab | NCT02263508 | Active, not recruiting | HSV-1 | Melanoma | III | 2014 | |
| - | NCT02173171 | Enrolling by invitation | HSV-1 | Any tumor type | Unspecified | 2014 | |
| Resection surgery | NCT02211131 | Active, not recruiting | HSV-1 | Melanoma | II | 2014 | |
| Pembrolizumab | NCT02263508 | Active, not recruiting | HSV-1 | Melanoma | III | 2014 | |
| - | NCT02297529 | No longer available | HSV-1 | Stage IIIB-IVM1c melanoma | III | 2014 | |
| Chemotherapy or PV-10 | NCT02288897 | Terminated | HSV-1 | Melanoma | III | 2014 | |
| - | NCT02366195 | Completed | HSV-1 | Stage IIIb-IVM1c melanoma | II | 2015 | |
| Radiation | NCT02453191 | Active, not recruiting | HSV-1 | Soft tissue sarcoma | I, II | 2015 | |
| Pembrolizumab | NCT02626000 | Completed [91] | HSV-1 | Squamous cell carcinoma of the head and neck | I | 2015 | |
| - | NCT02574260 | Completed | HSV-1 | Melanoma | II | 2015 | |
| Pembrolizumab | NCT02509507 | Recruiting | HSV-1 | Liver cancer | I | 2015 | |
| - | NCT02658812 | Active, not recruiting | HSV-1 | Breast cancer | II | 2016 | |
| - | NCT02756845 | Recruiting | HSV-1 | Advanced non-CNS tumors | I | 2016 | |
| Radiation | NCT02819843 | Recruiting | HSV-1 | Melanoma, Markel cell carcinoma, and other tumors | II | 2016 | |
| Radiation | NCT02923778 | Recruiting | HSV-1 | Soft tissue sarcoma | II | 2016 | |
| Nivolumab | NCT02978625 | Recruiting | HSV-1 | Melanoma, lymphoma, lung cancer, and other | II | 2016 | |
| Pembrolizumab | NCT02965716 | Recruiting | HSV-1 | Stage III and IV melanoma | II | 2016 | |
| Paclitaxel | NCT02779855 | Active, not recruiting | HSV-1 | Triple negative breast cancer | I, II | 2016 | |
| - | NCT02658812 | Completed | HSV-1 | Breast cancer | II | 2016 | |
| - | NCT02910557 | Recruiting | HSV-1 | Melanoma and herpetic infection | Unspecified | 2016 | |
| TTI-621 | NCT02890368 | Terminated | HSV-1 | Solid tumors and mycosis fungoides | I | 2016 | |
| - | NCT03086642 | Recruiting | HSV-1 | Pancreatic cancer | I | 2017 | |
| - | NCT03064763 | Active, not recruiting | HSV-1 | Stage IIIb- IV melanoma | I | 2017 | |
| Pembrolizumab | NCT03069378 | Recruiting | HSV-1 | Sarcoma | II | 2017 | |
| Atezolizumab | NCT03256344 | Active, not recruiting | HSV-1 | Metastatic colorectal and breast cancers | I | 2017 | |
| Chemotherapy + Radiation | NCT03300544 | Recruiting | HSV-1 | Rectal cancer | I | 2017 | |
| Dabrafenib + Trametinib | NCT03088176 | Active, not recruiting | HSV-1 | Melanoma | I | 2017 | |
| - | NCT03458117 | Unknown | HSV-1 | Non-melanoma skin cancer | I | 2018 | |
| - | NCT03555032 | Active, not recruiting | HSV-1 | Sarcoma and melanoma | I, II | 2018 | |
| - | NCT03663712 | Recruiting | HSV-1 | Stage IV peritoneal malignancy | I | 2018 | |
| Autologous CD1c (BDCA-1)+ myeloid dendritic cells | NCT03747744 | Active, not Recruiting | HSV-1 | Melanoma | I | 2018 | |
| Nivolumab | NCT03597009 | Recruiting | HSV-1 | Lung cancer | I, II | 2018 | |
| - | NCT03430687 | Withdrawn | HSV-1 | Bladder carcinoma | I | 2018 | |
| - | NCT03714828 | Recruiting | HSV-1 | Squamous cell carcinoma | II | 2018 | |
| Chemotherapy or endocrine therapy | NCT03554044 | Recruiting | HSV-1 | Breast cancer | I | 2018 | |
| - | NCT03921073 | Recruiting | HSV-1 | Skin angiosarcoma | II | 2019 | |
| Atezolizumab | NCT03802604 | Recruiting | HSV-1 | Breast cancer | I | 2019 | |
| Pembrolizumab | NCT03842943 | Recruiting | HSV-1 | Melanoma | II | 2019 | |
| Dabrafenib + Trametinib | NCT03972046 | Withdrawn | HSV-1 | Melanoma | II | 2019 | |
| Pembrolizumab | NCT04068181 | Recruiting | HSV-1 | Melanoma | II | 2019 | |
| Panitumumab | NCT04163952 | Recruiting | HSV-1 | Squamous cell carcinoma of the skin | I | 2019 | |
| Nivolumab + Trabectedin | NCT03886311 | Recruiting | HSV-1 | Sarcoma | II | 2019 | |
| - | NCT04065152 | Not yet recruiting | HSV-1 | Kaposi sarcoma | II | 2019 | |
| Ipilimumab + Nivolumab | NCT04185311 | Recruiting | HSV-1 | Breast cancer | I | 2019 | |
| - | NCT04330430 | Recruiting | HSV-1 | Stage III and IV melanoma | II | 2020 | |
| TBI-1401 | - | NCT01017185 | Completed | HSV-1 | Squamous cell carcinoma of the skin, breast carcinoma, melanoma, head and neck cancer | I | 2009 |
| Ipilimumab | NCT02272855 | Completed | HSV-1 | Melanoma | II | 2014 | |
| - | NCT02428036 | Completed | HSV-1 | Solid tumors | I | 2015 | |
| Ipilimumab | NCT03153085 | Completed | HSV-1 | Stage III and IV melanoma | II | 2017 | |
| Chemotherapy | NCT03252808 | Active, not recruiting | HSV-1 | Stage III and IV pancreatic cancer | I | 2017 | |
| G207 | - | NCT00028158 | Completed | HSV-1 | Brain cancer | I, II | 2001 |
| Radiation | NCT00157703 | Completed | HSV-1 | Malignant glioma | I | 2005 | |
| - | NCT02457845 | Active, not recruiting | HSV-1 | Brain cancer | I | 2015 | |
| - | NCT03911388 | Recruiting | HSV-1 | Brain cancers | I | 2019 | |
| - | NCT04482933 | Not yet recruiting | HSV-1 | High grade glioma | II | 2020 | |
| RP1 | Nivolumab | NCT03767348 | Recruiting | HSV-1 | Melanoma | I, II | 2018 |
| Cemiplimab | NCT04050436 | Recruiting | HSV-1 | Melanoma | II | 2019 | |
| - | NCT04349436 | Recruiting | HSV-1 | Squamous cell carcinoma | I | 2020 | |
| RP2 | Nivolumab | NCT04336241 | Recruiting | HSV-1 | Non-specified | I | 2020 |
| HSV-1716 | - | NCT00931931 | Completed [92] | HSV-1 | Non-CNS solid tumors | I | 2009 |
| - | NCT01721018 | Completed | HSV-1 | Malignant pleural mesothelioma | I, II | 2012 | |
| NV1020 | - | NCT00012155 | Completed | HSV-1 | Colorectal cancer | I | 2003 |
| - | NCT00149396 | Completed [93] | HSV-1 | Liver cancer and colorectal cancer | I, II | 2005 | |
| OrienX010 | - | NCT01935453 | Completed | HSV-1 | Melanoma, liver cancer, pancreatic cancer and lung cancer | I | 2013 |
| - | NCT03048253 | Unknown | HSV-1 | Melanoma | I-c | 2017 | |
| rRp450 | - | NCT01071941 | Recruiting | HSv-1 | Liver cancer | I | 2010 |
| M032 | - | NCT02062827 | Recruiting | HSV-1 | Brain cancers | I | 2014 |
| rQNestin | - | NCT03152318 | Recruiting | HSV-1 | Brain cancers | I | 2017 |
| C134 | - | NCT03657576 | Active, not recruiting | HSV-1 | Malignant glioma | I | 2018 |
| T3011 | NCT04370587 | Recruiting | HSV-1 | Head and neck cancer, Melanoma, Lung cancer, Soft tissue tumors and/or sarcoma, Solid tumors | I | 2020 | |
| ONCR-177 | Pembrolizumab | NCT04348916 | Recruiting | HSV-1 | Various tumors | I | 2020 |
| OH2 | HX008 | NCT03866525 | Recruiting | HSV-2 | Solid and GI tumors | I, II | 2019 |
| Pembrolizumab | NCT04386967 | Recruiting | HSV-2 | Solid tumors | I, II | 2020 | |
| - | NCT04637698 | Recruiting | HSV-2 | Pancreatic cancer | I, II | 2020 | |
| HX008 | NCT04616443 | Recruiting | HSV-2 | Melanoma | I, II | 2020 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aldrak, N.; Alsaab, S.; Algethami, A.; Bhere, D.; Wakimoto, H.; Shah, K.; Alomary, M.N.; Zaidan, N. Oncolytic Herpes Simplex Virus-Based Therapies for Cancer. Cells 2021, 10, 1541. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061541
AMA Style
Aldrak N, Alsaab S, Algethami A, Bhere D, Wakimoto H, Shah K, Alomary MN, Zaidan N. Oncolytic Herpes Simplex Virus-Based Therapies for Cancer. Cells. 2021; 10(6):1541. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061541
Chicago/Turabian StyleAldrak, Norah, Sarah Alsaab, Aliyah Algethami, Deepak Bhere, Hiroaki Wakimoto, Khalid Shah, Mohammad N. Alomary, and Nada Zaidan. 2021. "Oncolytic Herpes Simplex Virus-Based Therapies for Cancer" Cells 10, no. 6: 1541. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10061541
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.