
Characterization of RIN Isoforms and Their Expression in Tomato Fruit Ripening


Institute of Bioengineering, Research Center of Biotechnology, Russian Academy of Sciences, Leninsky Ave. 33, bld. 2, 119071 Moscow, Russia
*
Author to whom correspondence should be addressed.
Cells 2021, 10(7), 1739; https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071739
Submission received: 10 June 2021
/
Revised: 2 July 2021
/
Accepted: 7 July 2021
/
Published: 9 July 2021
(This article belongs to the Special Issue Molecular Regulation of Fruit-Set, Development, Maturation, Ripening and Shelf-Life)
Abstract
:Ripening of tomato fleshy fruit is coordinated by transcription factor RIN, which triggers ethylene and carotenoid biosynthesis, sugar accumulation, and cell wall modifications. In this study, we identified and characterized complete sequences of the RIN chromosomal locus in two tomato Solanum lycopersicum cultivars, its rin/RIN genotype, and three wild green-fruited species differing in fruit color and composition. The results reveal that S. lycopersicum cultivars and some wild species (S. pennellii, S. habrochaites, and S. huaylasense) had a 3′-splicing site enabling the transcription of RIN1i and RIN2i isoforms. The other wild species (S. arcanum, S. chmielewskii, S. neorickii, and S. peruvianum) had a 3′-splicing site only for RIN2i, which was consistent with RIN1i and RIN2i expression patterns. The genotype rin/RIN, which had an extended 3′-terminal deletion in the rin allele, mainly expressed the chimeric RIN–MC transcript, which was also found in cultivars (RIN/RIN). The RIN1, but not RIN2, protein is able to induce the transcription of the reporter gene in the Y2H system, which positively correlated with the transcription profile of RIN1i and RIN target genes. We suggest that during fruit ripening, RIN1 activates ripening-related genes, whereas RIN2 and RIN–MC act as modulators by competing for RIN-binding sites in gene promoters, which should be confirmed by further studies on the association between RIN-splicing mechanisms and tomato fruit ripening.
1. Introduction
Tomato Solanum lycopersicum L. is one of the most valuable crop plants in the world, producing fruit of high nutritional value. In addition, tomato is a good model to study the ripening of fleshy fruit, which is a complex process involving changes in color, texture, flavor, and taste [1]. The ripening process begins after the fruits have already reached their maximum size and transit from the mature green (MG) stage to fully heterotrophic stage, which is triggered by a climacteric boost in respiration and ethylene production [1]. At the molecular level, fruit ripening is regulated by three key transcription factors: MADS-domain RIPENING INHIBITOR (RIN or SlMADS-RIN), NAC NON-RIPENING (NOR), and SPL COLORLESS NON-RIPENING (CNR) [1,2]. Fruit-ripening regulation also includes transcription factors APETALA2a (AP2a), FRUITFULL (FUL1 and FUL2), and TOMATO AGAMOUS-LIKE1 (TAGL1) [2].
RIN is the main regulator of tomato fruit ripening. It controls the transcription of numerous genes involved in all stages of the process, including genes of the other two major transcription factors NOR and CNR, genes of the ethylene-dependent pathway, and structural genes associated with the ethylene-independent ripening network [3,4,5,6]. The list also includes genes responsible for the biosynthesis of carotenoids and aroma compounds, sucrose hydrolysis, and cell wall changes [3,4,5,6]. The accumulation of carotenoids, which define S. lycopersicum fruit color (yellow, orange, red, or brown), is already noticeable at the breaker (Br) stage and reaches a maximum at the red ripe (RR) stage [7]. In turn, carotenoids are converted to apocarotenoids, which serve as precursors for the synthesis of aroma compounds [8]. Sucrose hydrolysis and subsequent accumulation of monosaccharides define sweetness, whereas cell wall modifications—softness of ripe fruit [1].
S. lycopersicum belongs to the section Lycopersicon, which also includes 12 related wild tomato species mostly producing green fruit, except for Solanum cheesmaniae (L. Riley) Fosberg (yellow fruit), Solanum galapagense S.C. Darwin and Peralta (yellow-orange fruit), and Solanum pimpinellifolium B. Juss. (red fruit) [9,10]. The difference in fruit color between wild green-fruited (GF) tomatoes and S. lycopersicum cultivars is defined by carotenoid content [11], and that in taste by preferential accumulation of sucrose instead of glucose and fructose, respectively [12]. However, S. lycopersicum variants carrying the rin mutation are similar to GF species in fruit morphology and biochemical composition [2]. Thus, the fruit of both GF and rin plants retain their green color, do not accumulate moisture as they ripen, and remain firm for a longer time than cultivars. Such effects of the rin mutation on S. lycopersicum fruit are underlain by the suppression of cellular respiration and the associated ethylene release, which inhibits carotenoid accumulation, fruit softening, and production of aromatic compounds [2].
The rin mutation is caused by the deletion of a DNA region on chromosome 5, which results in the fusion of the adjacent truncated RIN and MACROCALYX (MC) genes (RIN–MC) [2,13]. The translated chimeric protein lacks a part of the RIN C-terminal activator domain and represents a gain-of-function mutant that actively represses ripening; in the nucleus, it interacts with other MADS-domain transcription factors and downregulates the expression of many ripening-related genes [13,14]. Therefore, it can be hypothesized that the differences in fruit ripening between wild GF tomato species and S. lycopersicum could be associated with the modifications of the RIN gene and its protein product. Understanding the role of the RIN and its isoforms in fruit ripening may be useful for breeding programs to increase fruit set quality.
In the current study, we tested this hypothesis by identifying and comparing complete sequences of the RIN chromosomal locus in wild-type S. lycopersicum cultivars, its rin/RIN genotype, and wild GF tomato species. We also performed expression profiling of the RIN isoforms in parallel with that of the RIN target genes involved in carotenoid and ethylene biosynthesis, sucrose hydrolysis, and cell wall metabolism, and tested the ability of the RIN1 and RIN2 isoforms to activate target gene transcription and interact with the known MADS-domain partners.
2. Materials and Methods
2.1. Plant Material
Seeds of S. lycopersicum cultivars, wild GF tomato species, and S. lycopersicum rin/RIN genotypes NN-25 and NN-21 (Table 1) were kindly provided by the N.I. Vavilov Institute of Plant Genetic Resources (VIR, St. Petersburg, Russia), the Federal Scientific Vegetable Center (FSVC, Moscow region, Russia), and the C.M. Rick Tomato Genetics Resource Center (TGRC, Davis, CA, USA). The NN-25 and NN-21 genotypes were progeny from the crossing of the cv. Zemba (VIR, St-Petersburg, Russia) with a line LA1795 homozygous for the rin mutation (unknown background; TGRC, Davis, CA, USA). Plants were grown in a greenhouse at 28/23 °C during the 16 h/8 h day/night light cycle (light intensity, 300–400 μmol m−2 s−1). Roots, leaves, flowers (anthesis), and fruit were collected in three biological replicates, homogenized in liquid nitrogen, and stored at −80 °C.
Fruits, forming red/red-to-violet (S. lycopersicum cultivars and NN-21 genotype) and pale yellow (NN-25 genotype) ripe fruits (Table 1), were harvested in planta at immature green (IG), MG, Br, and RR developmental stages. Fruits of wild GF tomato species were harvested in planta at MG and ripe final (RF) stages; fruit, both MG and RF, were green, of the final size, and hard and soft, respectively (i.e., the RR stage was defined by softness of the green fruit of a maximal size).
2.2. Identification and Structural Analysis of the RIN Locus
The sequences of the RIN chromosomal locus were amplified using genome-specific primers designed so that the neighboring amplicons overlapped (Supplementary Table S1). RIN–MC genomic sequences of S. lycopersicum cv. Heinz (Gene ID: 101252851) and S. pennellii (Gene ID: 107014532) were retrieved from NCBI GenBank (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/; genome annotation releases (accessed on 1 November 2020)), and primers were manually designed considering sequence polymorphism and checked using Primer3 (http://frodo.wi.mit.edu/primer3/ (accessed on 11 August 2020)). Multiple sequence alignments were conducted with MEGA 7.0.26 [16]. RIN target gene-specific primers were designed based on the S. lycopersicum mRNA sequences available in NCBI GenBank: PSY1 (NM_001247883.2), PSY2 (NM_001247742.2), Z-ISO (NM_001347622.1), ZDS (NM_001247454.2), PG2a (NM_001247092.2), EXP1 (NM_001247029.2), and E4 (NM_001320202.1) (Supplementary Table S1).
Genomic DNA was isolated from young leaves of a single plant of each tomato accession as previously described [17] and used as a template (100 ng) for PCR amplification at the following conditions: initial denaturation at 94 °C for 5 min, 35 cycles of denaturation at 94 °C for 30 s, primer annealing at 55 °C for 30 s, extension at 65 °C for 2 min, and final extension at 65 °C for 5 min. The amplified PCR products of the expected size were purified by using the QIAEX® II Gel Extraction kit (QIAGEN, Hilden, Germany), cloned in the pGEM®-T Easy (Promega, Madison, WI, USA), and sequenced (3–5 clones for each accession) on ABI Prism 377 DNA Sequencer (Applied Biosystems, Waltham, MA, USA) using the designed primers (Supplementary Table S1).
2.3. Gene Expression Analysis
Total RNA was extracted using the RNeasy Plant Mini Kit (QIAGEN, Hilden, Germany), purified from genomic DNA (RNase free DNasy set; QIAGEN, Hilden, Germany), qualified by gel electrophoresis, and used for first-strand cDNA synthesis (GoScript Reverse Transcription System; Promega, Madison, WI, USA) with an oligo-dT primer. RNA and cDNA concentrations were quantified by fluorimetry (Qubit® Fluorometer, Thermo Fisher Scientific, Waltham, MA, USA).
Reverse transcription PCR was performed using 3.0 ng of cDNA and specific primers (Supplementary Table S1) at the following cycling conditions: initial denaturation at 95 °C for 5 min and 40 cycles of denaturation at 95 °C for 15 s and annealing/extension at 60 °C for 40 s. To confirm the amplification of the expected DNA fragment, PCR products were purified and sequenced. To ensure the specificity of the primers, a melting curve analysis was performed.
Quantitative (q) real-time (RT)-PCR was performed in 96-well plates with a CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, USA), 3.0 ng of cDNA, SYBR Green RT-PCR mixture (Syntol, Moscow, Russia), and specific primers (Supplementary Table S1) at the following cycling conditions: initial denaturation at 95 °C for 5 min and 40 cycles of denaturation at 95 °C for 15 s and annealing/extension at 60 °C for 40 s.
To normalize the levels of gene expression, two reference tomato genes, Expressed (SGN-U346908) and Actin 2/7 (NM_001330119.1) [18,19], were used. The qRT-PCR results were statistically analyzed with Graph Pad Prism version 8 (GraphPad Software Inc., San Diego, CA, USA; https://www.graphpad.com/scientific-software/prism/ (accessed on 1 March 2021)). The data were expressed as the mean ± standard deviation (SD) based on three technical replicates of three biological replicates for each combination of cDNA samples and primer pairs. The unequal variance (Welch’s) t-test was applied to assess the statistical significance (p-value < 0.05) of the differences in gene expression between samples.
2.4. In Vivo Analysis of Protein–Protein Interactions
Two-hybrid analysis was performed according to HybriZAP-2.1 Two-Hybrid cDNA Synthesis kit protocol (Stratagene, La Jolla, CA, USA) at room temperature and 30 °C, as previously described [20,21]. The bait and prey constructs were generated by cloning full-length cDNAs of the tomato genes into pAD-GAL4 and pBD-GAL4-Cam vectors (Stratagene, La Jolla, CA, USA).
2.5. Carotenoid Content
Total carotenoid content was measured by spectrophotometry in two biological and three technical replicates using a modified Folch method [22,23]. Briefly, 0.2 g of plant tissue was homogenized in Folch solution (chloroform:methanol 2:1 (v/v)) in the presence of trace Mg2CO3 amounts [23], incubated at 4 °C for 1 h, and centrifuged at 4000× g rpm for 10 min at 4 °C. The lower chloroform phase was collected and used for spectrophotometric assay of chlorophyll, lycopene, and β-carotene. Total carotenoid contents were measured in acetone–hexane solutions as previously described [24] using a spectrophotometer (Basic, Eppendorf, Hamburg, Germany) and quantified according to the following equations:
where A is the absorbance at the indicated wavelengths and x + c is the sum of xanthophylls and carotenes.
Chlorophyll a (µg/mL) = 11.47 (A666 − A750) − 2 (A648 − A750)
Chlorophyll b (µg/mL) = 21.85 (A648 − A750) − 4.53 (A666 − A750)
Total carotenoids (x + c) (µg/mL) = [1000 (A480 − A750) − 1.33 Chla − 23.93 Chlb]/202
Lycopene (mg/100 mL) = 0.204 A645 − 0.0458 A663 + 0.372 A505 − 0.0806 A453
β-Carotene (mg/100 mL) = 0.216 A663 − 1.22 A645 − 0.304 A505 + 0.452 A453
3. Results
3.1. Identification of the RIN Locus in the Analyzed Tomato Accessions
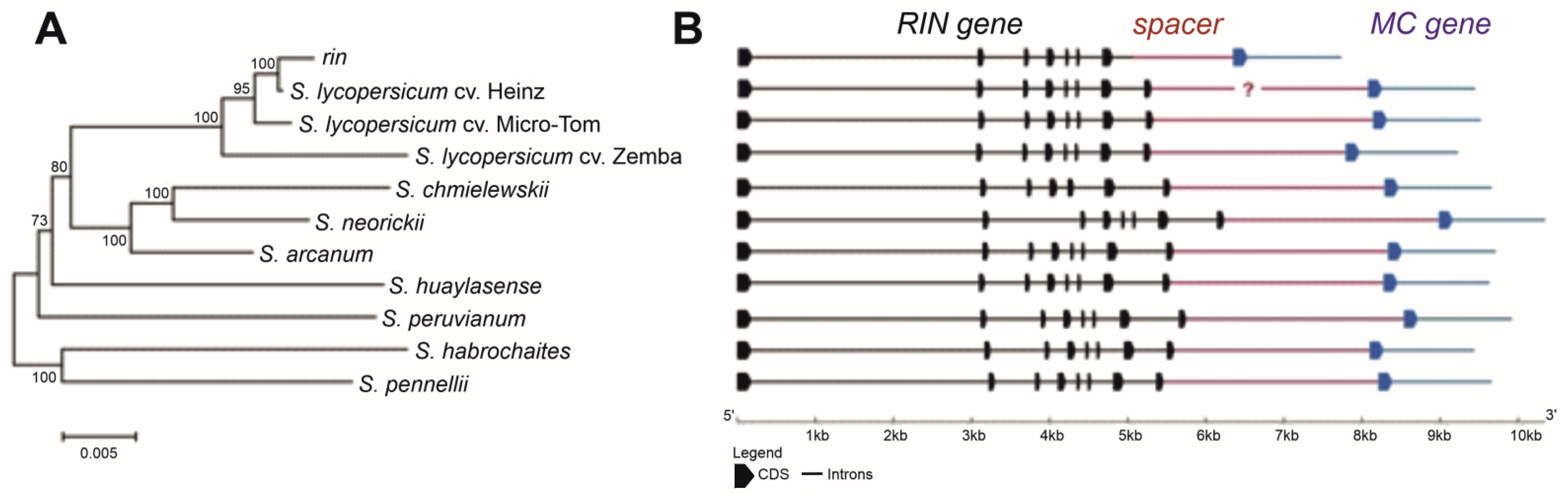
The RIN genomic locus was identified (cloned and sequenced, or identified in silico) in three S. lycopersicum cultivars, its rin/RIN genotype, and seven wild GF tomato species (Table 1 and Table 2). The analyzed fragments included the complete RIN gene, RIN–MC intergenic spacer, and 5′-terminal fragment comprising exon I and partially intron I of the MC gene (Figure 1). The total length of this genomic region varied among the accessions: from 7724 bp in rin/RIN to 10,348 bp in S. neorickii (Figure 1).
All analyzed tomato accessions had the same structure of the RIN gene, which contained eight exons and seven introns; the gene length varied from 5288 (cv. Heinz) to 6230 bp (S. neorickii) (Figure 1 and Table 2). In the NN-25 genotype, both mutant (rin) and wild-type (RIN) alleles were identified, confirming the heterozygous state of the gene (rin/RIN); the rin allele (5089 bp) was characterized by the absence of the 3′-fragment of the intron VII and the whole exon VIII (Figure 1 and Table 2).
The level of RIN polymorphism among the analyzed tomato accessions was 10.43%: in total, 714 single nucleotide polymorphisms (SNPs) were identified; among them, 48 (0.7%) were found in S. lycopersicum cultivars and 638 (9.32%) in wild GF species. Three SNPs were specific to the NN-25 rin allele (compared to S. lycopersicum cultivars and wild GF species), 30 to wild GF species (compared to S. lycopersicum cultivars and NN-25) (Supplementary Figure S1), and none to S. lycopersicum cultivars compared to the rin mutant or wild GF species. Among exon SNPs, 14 were non-synonymous in wild species compared to cultivars; five amino acid substitutions were located in the conserved DNA-binding MADS domain, but none in the RIN gene of the analyzed GF species.
The analyzed accessions significantly differed in the RIN–MC spacer, whose length varied from 1395 (NN-25) to 2816 bp (cv. Micro-Tom) (Supplementary Figure S2). Despite good assembly and coverage of the S. lycopersicum cv. Heinz genome, it has an undefined region inside the RIN–MC spacer, which is also present in the genomes of other S. lycopersicum cultivars (such as M82 and I-3) available in the NCBI database. To determine the missing sequence, we tried to amplify it from the genomes of Heinz, All Round, Moneymaker, Ailsa Craig, and Yellow Peach cultivars using different primer pairs specific for the flanking sequences (Supplementary Table S1); however, there was no product, probably because of the presence of mobile elements or other extended sequences inside the RIN–MC spacer, which prevented its amplification. In other accessions analyzed in this study, the missing region was successfully amplified and sequenced (Supplementary Figure S2).
Thus, the structure of the analyzed RIN genomic fragment was similar in three S. lycopersicum cultivars and seven wild GF tomato species but differed from that of the rin allele. The rin locus had an extended deletion, which started in intron VII and included the entire exon VIII and a fragment of the RIN–MC intergenic spacer. The RIN–MC intergenic spacer was 1700 bp in cv. Micro-Tom, 1487 bp in cv. Zemba, 1491 bp in S. habrochaites, and 1673–1790 bp in the other wild GF species.
3.2. Identification of Splicing Sites in the RIN Homologs
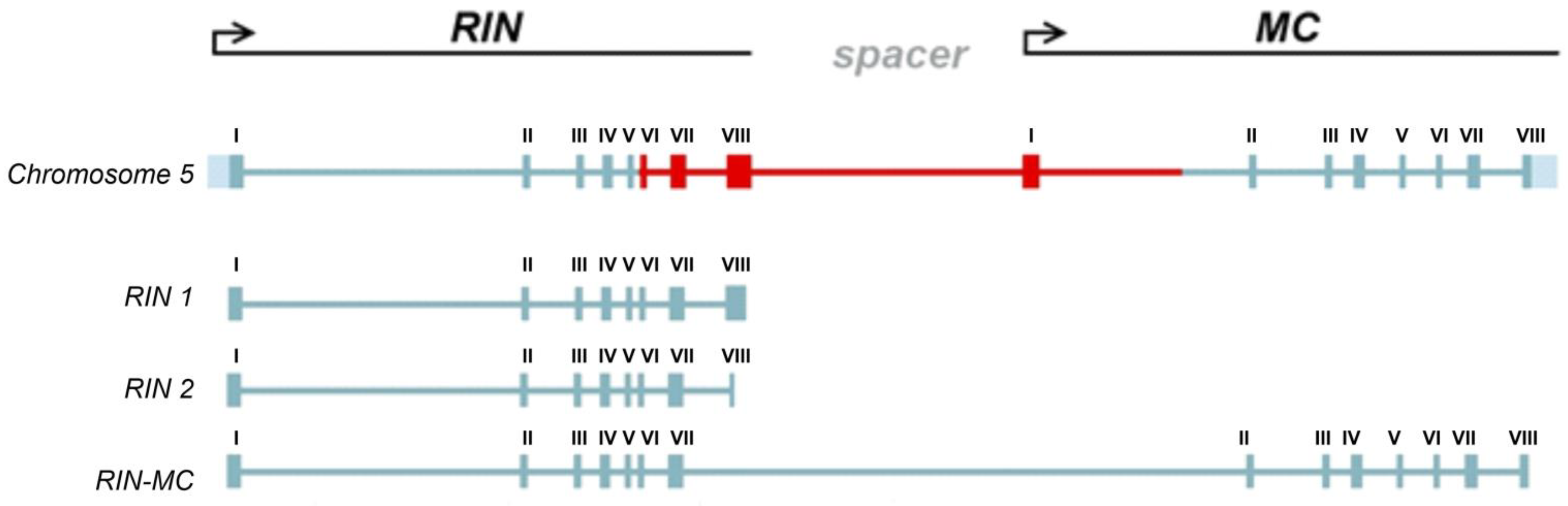
The RIN locus produces three different transcripts: isoforms RIN1i and RIN2i, and chimeric RIN–MC, which was previously identified in the S. lycopersicum rin/rin mutant [13] (Figure 2). All intron sequences of the identified RIN homologs contained a consensus of the main 3′-splicing site (NNNYAG) present in dicots [25] as well as an additional site TGCTAG at the beginning of exon IV; in S. chmielewskii, intron V 3′-splicing site contained an SNP (Figure 3). All GF species had a double 3′-splicing site in intron VII before exon VIII; another splicing site in this region was present in S. lycopersicum, S. huaylasense, S. habrochaites, and S. pennellii as well as in all S. lycopersicum cultivars (Figure 3). The NN-25 rin allele did not have the corresponding region of intron VII and exon VIII.
3.3. Expression of the Three RIN Transcripts in S. lycopersicum, rin/RIN, and Wild GF Species
To analyze the expression of different RIN transcripts, we used one common forward primer and three reverse primers specific for RIN1i (NM_001247741.2), RIN2i (NM_001315495.1), and RIN–MC (NM_001247047.2) (Supplementary Table S1), which allowed the amplification of full-length transcripts with the expected lengths of 729 (RIN1i), 660 (RIN2i), and 1194 bp (RIN–MC) (according to S. lycopersicum cv. Heinz RIN–MC sequence).
We determined the accumulation of the three transcripts in various organs of S. lycopersicum cv. Heinz, including roots, leaves, and flowers, as well as fruit pulp and skin at MG, Br, and RR developmental stages. The results reveal that RIN1i and RIN2i were expressed in all analyzed organs and tissues (except for the leaves which lacked RIN2i mRNA); the maximum level was observed in fruit, where it was increased at the Br and RR stages in the pulp and peel (Supplementary Figure S3). Surprisingly, the chimeric RIN–MC transcript was also found, albeit at a level much lower than those of RIN1i and RIN2i, in ripening fruit (Br and RR stages) (Supplementary Figure S3).
We compared the fruit-specific expression of RIN1i, RIN2i, and RIN–MC mRNA in S. lycopersicum, its rin/RIN genotype, and wild GF species. In the fruit of S. lycopersicum cv. Heinz and Zemba, the mRNA level of RIN1i was rather high, whereas that of RIN2i was lower; both isoforms were accumulated during fruit ripening, reaching a maximum at the Br and RR stages (Supplementary Figure S4). The RIN–MC transcript was expressed in fruit flesh of both cv. Zemba and Heinz at a level much lower than that of RIN1i and RIN2i (Supplementary Figure S4). In the rin/RIN genotype, the expression of RIN1i and RIN2i was weaker but still detected in all fruit stages (except for RIN2i at Br), thus confirming the heterozygous (rin/RIN) status of the RIN gene; the transcription of chimeric RIN–MC was first detected at the MG stage and was upregulated with fruit ripening, reaching a maximum at the RF stage (Supplementary Figure S4).
In the wild GF species S. habrochaites and S. peruvianum, the expression of RIN1i was observed in both hard and soft fruit of the final size (MG and RF, respectively), whereas that of RIN2i was much weaker and detected only in S. habrochaites, and that of RIN–MC was absent (Supplementary Figure S4). All the amplified PCR fragments were sequenced to confirm the amplification of the RIN1i, RIN2i, and RIN–MC transcripts (Supplementary Figures S5 and S6). The sequence of the RIN–MC transcript was the same for all analyzed accessions (Supplementary Figure S5). Semi-quantitative PCR allows only an approximate estimate of the levels of gene transcription, so we performed a quantitative analysis of gene expression.
3.4. RIN Isoform Expression Profiling in Developing Fruit of S. lycopersicum Cultivars, the rin/RIN Genotype, and Wild GF Species
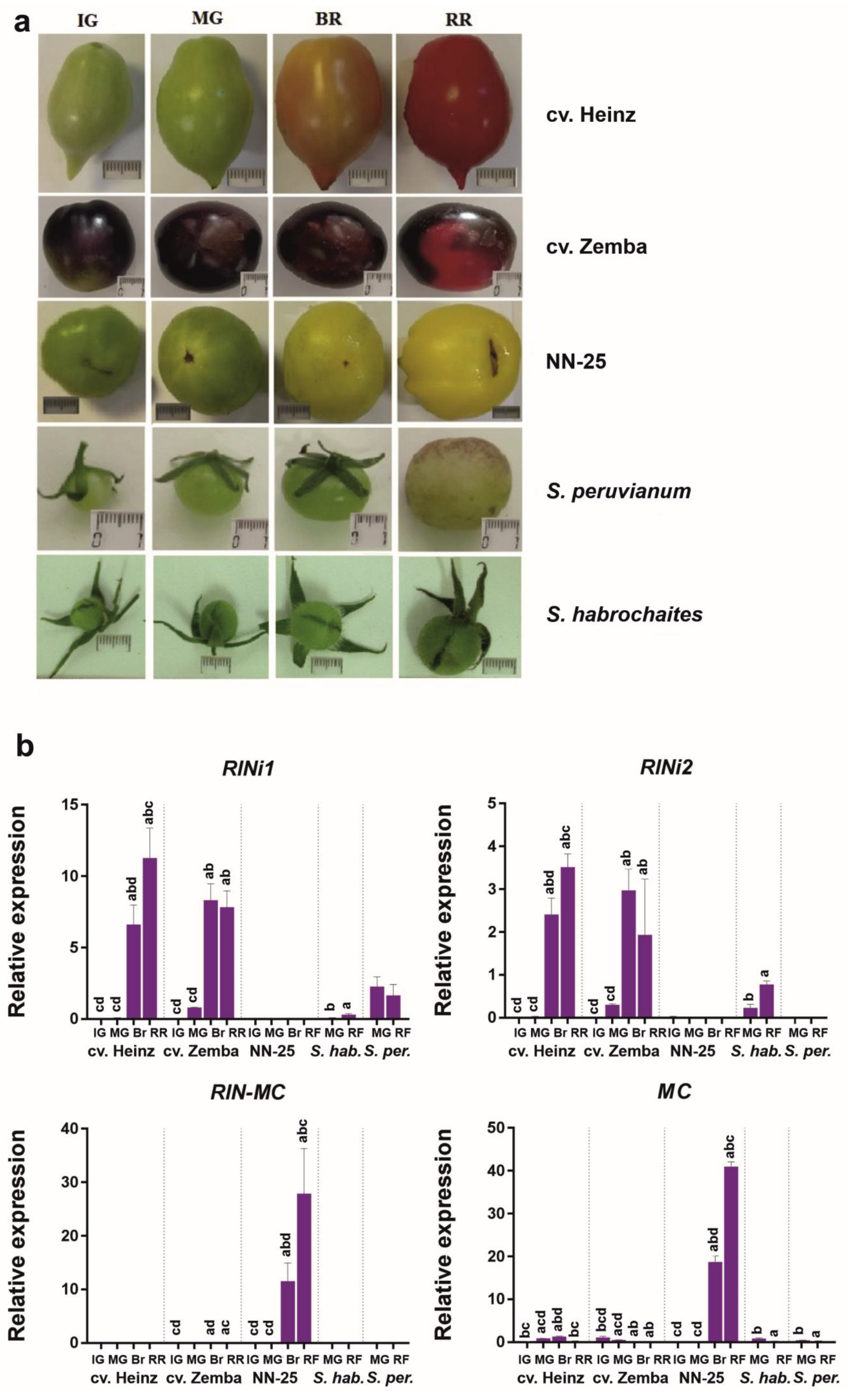
Quantitative expression analysis by qRT-PCR confirmed that the fruit of S. lycopersicum cultivars had very weak expression of RIN–MC and quite strong expression of RIN1i and RIN2i; the mRNA levels of the RIN1i were almost three times higher than those of the RIN2i (Figure 4). The opposite expression pattern was revealed in the rin/RIN genotype: trace amounts of RIN1i and RIN2i (similar to RIN1i and RIN2i in cv. Zemba; Supplementary Table S2), and high amounts of RIN–MC mRNA. Between the analyzed GF species, the expression of RIN1i and RIN2i was detected in the fruit of S. habrochaites, although at a much lower level than in S. lycopersicum, whereas in those of S. peruvianum, only RIN1i was expressed (Figure 4), which corresponded to the absence of the splicing site required for RIN2i transcription (Figure 3). Except for the rin/RIN genotype, the expression of chimeric RIN–MC and MC mRNA (NM_001247736.1) was absent or negligible in all other species (Figure 4). In the rin/RIN genotype, MC mRNA significantly exceeds that in the other accessions, primarily due to the fact that the primer pair for MC is suitable for the RIN–MC; nevertheless, the difference between MC and RIN–MC levels suggests that the levels of MC transcription exceeds that in the other accessions by over 100 times (Figure 4). These results indicate that the transcriptional mechanisms underlying the expression of the RIN locus differed among wild GF species, S. lycopersicum, and its rin/RIN genotype. Considering that the rin/RIN plant should form red fruits due to the functional RIN allele, CNR gene expression was analyzed in mature green and ripe NN-25 and NN-21 fruits, and an almost 4–9-fold reduction in CNR transcription was found in NN-25 compared to NN-21 (Supplementary Figure S7).
3.5. Expression of RIN Target Genes during Tomato Fruit Development
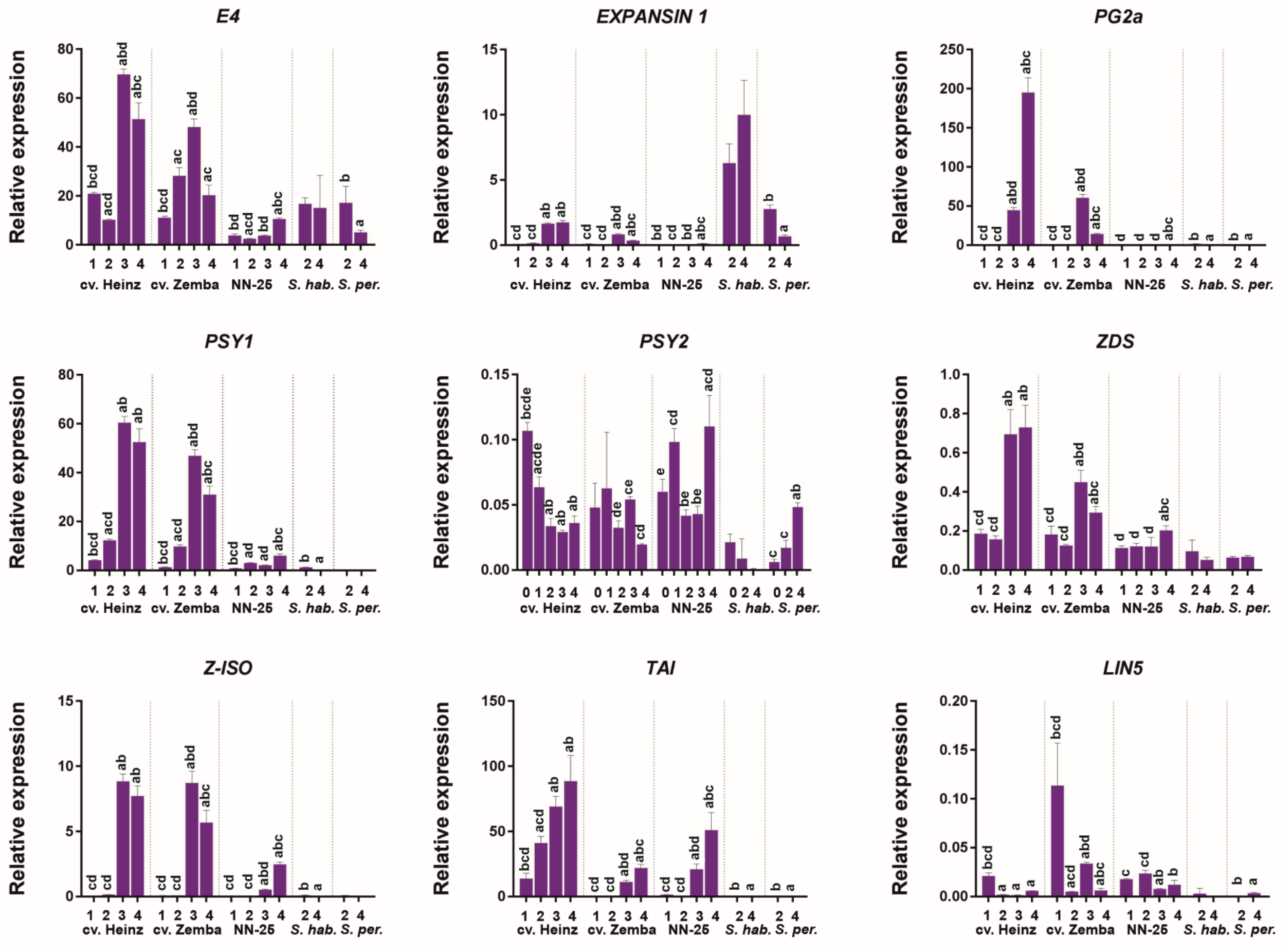
We analyzed the expression patterns of putative RIN target genes related to the fruit softening (PG2a, EXPANSIN 1 (EXP1), and E4), carotenoid biosynthesis (PSY1, PSY2, Z-ISO, and ZDS), and sucrose hydrolysis (TAI and LIN5). The expression of the EXP1 gene was the strongest in GF species, weakest in S. lycopersicum cultivars, and absent in the NN-25, whereas that of the E4 gene was the strongest in S. lycopersicum cultivars, weaker in GF species, and the weakest in the NN-25 (Figure 5). The expression of the PG2a gene was quite strong in Br and RR fruit of S. lycopersicum cultivars, but low in the other accessions (Figure 5).
The expression of PSY1 in S. lycopersicum cultivars was upregulated during fruit ripening and was quite strong at its final stages, but very weak in the rin/RIN genotype, and almost absent in GF species (Figure 5). At the same time, the PSY2 expression level was quite low both in S. lycopersicum cultivars and the rin/RIN genotype; it was the highest in the leaves of cv. Heinz and in RF of the rin/RIN genotype and the lowest in GF species (Figure 5). Z-ISO expression was detected only in Br and RR fruits of S. lycopersicum accessions; it was significantly stronger in wild-type plants than in the rin/RIN genotype (Figure 5). The expression of ZDS was found in all tested samples; it was the strongest in S. lycopersicum cultivars where it reached a maximum in Br and RR fruit, the weakest in S. peruvianum, and intermediate in rin/RIN where ZDS expression was slightly increased in RF fruit (Figure 5).
The expression of the TAI gene was detected in S. lycopersicum cultivars and rin/RIN, where it was upregulated with ripening, and was absent in S. habrochaites and S. peruvianum, whereas LIN5 expression was very low in all analyzed accessions, especially in GF species (Figure 5).
3.6. Carotenoid Content in Ripe Fruit of S. lycopersicum Cultivars, Its rin/RIN Genotype, and GF Species
In RR fruit of S. lycopersicum cultivars, the carotenoid content (x + c, lycopene, and β-carotene) was equally high, whereas in ripe fruit of the rin/RIN and GF species the total carotenoid (x + c) content was about 10 times lower, the level of β-carotene was similar (S. peruvianum) or 2–7 times lower (NN-25 and S. habrochaites), and lycopene was absent (Table 3).
3.7. Analysis of Protein–Protein Interactions of RIN1 and RIN2 Isoforms
The interactions of RIN1 and RIN2 proteins with the known RIN partners, the MADS-domain transcription factors FRUITFULL 2 (FUL2) and TOMATO AGAMOUS-LIKE 1 (TAGL1), were analyzed in vivo in yeast cells. The results indicate that both isoforms could interact with the two proteins, but such interactions were much stronger for RIN1 than for RIN2 and only RIN1 demonstrated the ability to activate the transcription of the reporter gene, whereas RIN2 did not (Table 4).
4. Discussion
The transcription factor RIN plays an important role in the regulation of the tomato fruit ripening; it does not participate in ripening induction but is strictly necessary for full ripening and prevention of over-ripening [1,2,3,4,5,6,27]. RIN activates gene transcription through its C-terminal domain and could also interact with different MADS-domain proteins through its other regions, forming multimeric complexes that bind to the promoters of target genes [27]. In the rin mutant, the RIN gene lacks exon VIII and part of exon VII (corresponding to the C-terminus) and is inactivated [27].
In the present study, we identified the RIN locus in S. lycopersicum cultivars, its rin/RIN genotype, and wild GF tomato species significantly differing in the morphology, taste and color of ripe fruit, which are red and soft in cultivars, green and soft in wild species, and pale yellow and firm in the rin/RIN genotype; such distictions depend on carotenoid accumulation, sugar content, and ethylene biosynthesis. Pale yellow ripe fruits of the NN-25 genotype are close in color to those of wild species, whereas the rin/RIN ripe fruits are reported to be red [28]. A significant decrease in CNR expression was observed in fruits of NN-25 compared to NN-21 (the same background as in NN-25) (Supplementary Figure S7). The Cnr allele is epistatic over the rin allele and masks the rin effects [28], and Cnr epimutation in the promoter is accompanied by a sharp decrease in CNR expression level and pale yellow fruit phenotype [29]. Given this, we may assume that the NN-25 fruits have a pale yellow color due to either the possible rin/RIN Cnr/CNR genotype, or very low (in comparison with red-fruited tomatoes) PSY1 gene expression (Figure 5).
In terms of ripening characteristics, the fruit of the NN-25 are closer to those of wild tomato species than to those of cultivars despite the fact that the rin locus of the NN-25 has a characteristic deletion [2], whereas the RIN structure in GF species is more similar to that of cultivars (Figure 1, Supplementary Figure S1). Thus, it can be suggested that the incomplete fruit ripening in wild GF tomato species and rin/RIN is caused by different mechanisms.
The S. lycopersicum RIN gene produces two mRNA isoforms (RIN1i and RIN2i) because of the presence of the alternative 3′-splice site; the respective proteins RIN1 (NP_001234670.1) and RIN2 (NP_001302424.1) differ in the C-terminus (Figure 2 and Figure 6) and the longer variant, RIN1, has been reported to act as a transcriptional activator [28,29,30,31,32,33,34]. The rin/rin mutant produces the RIN–MC fusion transcript, but the transcriptional activation potential of the RIN–MC protein is controversial: although it has been shown to induce the luciferase (LUC) gene, it rather does not activate the ripening genes [13,14].
The ability of RIN to perform transcriptional activation depends on its C-terminus [35], which in RIN2 is five residues longer (TLPISTINT) than in the RIN of the mutant rin allele or RIN–MC (TLPI) (Figure 6, Supplementary Figure S5), suggesting that, since RIN–MC may perform transcriptional activation, RIN2 could also do so through binding to the RIN target genes. However, our analysis of the RIN1 and RIN2 in a two-hybrid system indicated that RIN2 could dimerize with TAGL1 and FUL2 (Table 4), known as RIN partners [30,31,32,33,34], but could not activate the transcription of the reporter gene. Given these results and the fact that MC does not act as a transcriptional activator [35], the RIN–MC protein should also lack the ability to induce transcription.
Overall, these findings suggest that RIN1 activates fruit ripening-related genes, whereas RIN2 and RIN–MC regulate the RIN target genes by competing with RIN1 for the binding sites. It is possible that the plant has developed such a system for high-precision tuning of the fruit-ripening process.
The immaturity of the rin fruit is associated with the expression of RIN–MC and the absence of that of RIN (RIN1) [2,13,14]. This is consistent with our results showing that the identified S. lycopersicum RIN homologs could produce a significant amount of the RIN1i transcript, whereas the NN-25—only its traces; moreover, the NN-25 highly expresses the RIN–MC (Figure 4). At the same time, the RIN genes of GF species are split in terms of the presence of the splicing site for the RIN2i transcript: some of them had it (S. pennellii, S. habrochaites, and S. huaylasense) and some did not (S. arcanum, S. chmielewskii, S. neorickii, and S. peruvianum) (Figure 3). This is consistent with qRT-PCR results showing the low presence of RIN1i in both S. habrochaites and S. peruvianum fruits, and RIN2i only in S. habrochaites fruits (Figure 4). In S. habrochaites, the RIN2 may compete with RIN1 for target ripening genes, suppressing their expression, which may explain the formation of green-colored fruits. The green fruits of S. peruvianum are likely to be due to the fact that the expression of RIN1i is about 4–5 times lower than that of the S. lycopersicum cultivars (Figure 4).
Expression analysis confirmed that all three RIN transcripts were expressed in S. lycopersicum cultivars; however, the RIN2i expression level was 2–3 times lower than that of RIN1i and only trace amounts of the RIN–MC transcript were detected (Supplementary Figures S3 and S4, Figure 4). It should be noted that RIN–MC expression has not been previously reported in wild-type tomatoes but only in rin mutants [13]. It can be hypothesized that the RIN–MC transcript is expressed in S. lycopersicum accessions because of the presence of 3′-splicing site AAACAG upstream of the MC sequence involved in the fusion (Supplementary Figure S5). However, the MC genes of GF species S. peruvianum (KY471421.1) and S. habrochaites (KY471429.1) also had this 3′-splicing site but did not express RIN–MC mRNA (Supplementary Figure S4, Figure S5). Possibly, small amounts of the RIN–MC transcripts are normally synthesized at the RIN–MC locus using existing splice sites, whereas in the mutant rin allele, due to the deletion of the last exon of the RIN gene, the equilibrium is shifted towards the formation of RIN–MC.
In the rin/RIN genotype, the transcription levels of RIN1i and RIN2i were significantly lower, whereas those of chimeric RIN–MC were significantly higher than in the cultivars (Figure 5 and Figure 6), which could be due to the presence of only one wild-type allele and the absence of splice sites for RIN1i and RIN2i in the rin allele. The expression of both splicing variants in GF S. habrochaites and only RIN1i in S. peruvianum (Figure 5 and Figure 6; Supplementary Figure S1) was in accordance with the presence of the RIN2i splicing site in the S. peruvianum RIN and its absence in the S. habrochaites RIN (Figure 3).
RIN binds to over a thousand genomic regions, including promoters of numerous ripening-related genes, including those encoding transcription factors and proteins involved in ethylene production, carotenoid biosynthesis, and cell wall modification [3,4,5,35,36,37]. The RIN-exerted effects can be either transcriptional upregulation (RIN1), downregulation (RIN–MC), or more complex modulation including both RIN1 and RIN–MC [13], which is consistent with our results. Therefore, the differential transcription of the RIN isoforms in the analyzed tomato accessions should directly affect the expression of the target genes. Significantly lower RIN1i mRNA levels in GF species and rin/RIN may be correlated with the decreased production of carotenoids (Table 3) as well as ethylene biosynthesis, which should change the color and increase the hardness of the fruit, respectively [6]. Genes involved in the biosynthesis of carotenoids are differentially regulated by RIN isoforms. For example, the expression of the phytoene synthase gene PSY1 required for phytoene/lycopene synthesis during tomato ripening can be regulated by both RIN and RIN–MC, whereas that of PSY2 involved in the synthesis of carotenoids in immature green fruit is regulated by RIN–MC [3,13]. Z-ISO, which encodes carotene zeta isomerase converting colorless 15-cis phytoene to red trans-lycopene [38], is regulated by RIN and RIN–MC [3,13], whereas ZDS encoding zeta-carotene desaturase, which converts zeta-carotene to lycopene, is regulated by RIN but not RIN–MC [13]. Our results indicate that in the analyzed tomato accessions, the expression level of RIN1i (Supplementary Figures S3 and S4, Figure 4) was directly correlated with those of PSY1, Z-ISO, and ZDS (Figure 5) and carotenoid content (Table 3): all were high in red fruit of S. lycopersicum cultivars and low in pale yellow and green fruit of the rin/RIN and GF species, respectively, which is consistent with PSY1 expression levels in cultivars and the rin/RIN Cnr/CNR revealed in a previous study [28]. The expression of PSY2, which is mostly observed in chloroplasts, was similar in the leaves and fruit (Figure 5), suggesting that the chloroplast-specific carotenoid synthesis was maintained at the basic low level in both photosynthetic and reproductive tissues of the analyzed accessions and, furthermore, that the RIN–MC did not activate the PSY2 gene as has been proposed earlier [13].
The other RIN target genes, TAI and LIN5 [13,39], are responsible for the taste and texture of tomato fruit. TAI encodes vacuolar invertase, converting sucrose, which is accumulated in wild GF tomato species, to glucose and fructose, characteristic for ripe fruit of S. lycopersicum cultivars [12], whereas LIN5 encodes cell wall invertase, which affects the biosynthesis of cuticle components and sugar absorption by tomato fruit [40]. In this study, the expression of TAI and LIN5 was noticeable only in S. lycopersicum (cultivars and NN-25) (Figure 5) and corresponded to high hexose content in their fruit (Table 1). However, there was no correlation between TAI/LIN5 and RIN1i mRNA levels, suggesting that TAI and LIN5 may be competitively regulated by RIN1 and RIN2/RIN–MC, which exert positive and negative, respectively, transcriptional effects. A low level of LIN5 expression may indicate that this gene is involved in the control of sugar content only in the cuticle but not in the whole fruit.
The PG2a gene encodes polygalacturonase-2a associated with cell wall modification (polyuronide degradation) and texture. PG2a expression, which is induced by ethylene and reaches a maximum in ripe fruits [41,42], has been shown to be regulated by RIN–MC but not RIN (RIN1) [13], although the presence of RIN-binding sites in the promoter [3] implies that it should respond to all RIN isoforms. We observed strong PG2a expression only in Br and RR fruit of S. lycopersicum cultivars (Figure 5), suggesting that RIN1, rather than RIN–MC, may be involved in PG2a upregulation.
EXP1, which is one of the key genes defining fruit softness [43], is suggested to be regulated by both RIN (RIN1) and RIN–MC [13]. However, the RIN1 and RIN–MC expression profiles (Supplementary Figure S4, Figure 4) suggest that EXP1 transcription (Figure 5) can be repressed by RIN–MC.
The E4 gene encoding methionine sulfoxide reductase, a repair enzyme for oxidation-damaged proteins, is a target of RIN1 but not RIN–MC [13] and is upregulated after ethylene induction [44]. In this study, the E4 transcription levels were high in cultivars, intermediate in GF species, and low in the rin/Rin genotype (Figure 5), and were correlated with RIN1 expression (Figure 4), confirming the role of RIN1 in E4 activation.
5. Conclusions
In this study, we identified and characterized complete sequences of the RIN chromosomal locus in S. lycopersicum cultivars, its rin/RIN genotype, and wild GF tomato species, which differ in ripe fruit morphology and composition. Cultivars, as well as some wild species (S. pennellii, S. habrochaites, and S. huaylasense), had a splicing site in intron VII/exon VIII of the RIN gene, which enabled the transcription of two RIN isoforms, RIN1i and RIN2i, whereas the other GF species (S. arcanum, S. chmielewskii, S. neorickii, and S. peruvianum) did not have this splicing site. Accordingly, S. lycopersicum cultivars as well as GF S. habrochaites expressed RIN1i and RIN2i, whereas S. peruvianum—only RIN1i. The rin/RIN genotype had an extended deletion, including exon VIII and a fragment of the RIN–MC intergenic spacer, and mostly expressed the chimeric RIN–MC fragment, which was also detected in S. lycopersicum cultivars. Although both RIN1 and RIN2 could interact with MADS-domain-containing RIN partners, only RIN1 could activate the transcription of reporter genes, which was consistent with the positive correlation between the expression of putative RIN target genes and that of RIN1i. We assume the existence of a link between differences in fruit ripening of wild GF tomato species and S. lycopersicum cultivars with modifications of the RIN gene and its protein product. High expression of the functional RIN isoform, RIN1, which prevails over the expression of RIN2i and RIN–MC, in fruits of tomato cultivars leads to the activation of ripening genes that are RIN targets; as a result, the fruit turns red (or yellow, pink, brown, etc.—due to the accumulation of carotenoids, mainly lycopene). The extremely low expression of RIN1i and the high level of RIN–MC in the rin/RIN genotype correspond to a pale yellow color of the fruit due to the low presence of the RIN1 protein (competing with RIN–MC for partners and targets), which leads to a weak induction of the expression of carotenoid pathway genes, PSY1, ZDS, etc. Low amounts of RIN1i, PSY1, ZDS, and Z-ISO transcripts, as well as the presence of RIN2i and RIN–MC in wild tomato species, correspond to ripe green fruits without lycopene. These results provide further insights into the molecular mechanisms regulating tomato fruit ripening.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/cells10071739/s1, Figure S1: Alignment of the RIN gene from S. lycopersicum cultivars, rin/RIN genotype (rin allele), and wild GF tomato species; Figure S2: Alignment of the spacer between RIN and MC genes from S. lycopersicum cultivars, rin/RIN genotype (rin allele), and wild GF species; Figure S3: Expression of RIN1i, RIN2i, and RIN–MC in different organs and tissues of S. lycopersicum cv. Heinz. R, roots; L, leaves; F, opened flowers; MG, mature; Figure S4: Fruit-specific expression of RIN1i, RIN2i, and RIN–MC. NN-25, the rin/RIN genotype; S. hab., S. habrochaites; S. per., S. peruvianum; IG, immature green (flesh); MG, mature green (flesh); Br, breaker (flesh); RR, ripe red (flesh); RF, ripe final (flesh). The Actin gene was used as reference; Figure S5: Alignment of the RIN–MC transcript from S. lycopersicum cultivars and the rin/RIN genotype; Figure S6: Alignment of the RIN1i and RIN2i transcripts from S. lycopersicum cultivars, rin/RIN genotype, and wild tomato species; Figure S7: Expression of the CNR gene in mature green and ripe fruits of the NN-21 and NN-25 accessions. Lowercase letters above the bars indicate statistically significant differences (p < 0.005) between gene expression levels; Table S1: List of primers used for gene amplification, sequencing, and expression analysis; Table S2: RIN1i, RIN2i, and RIN–MC relative expression data in fruits of S. lycopersicum cultivars, rin/RIN genotype NN-25, and wild GF tomato species.
Author Contributions
Performed the experiments: M.A.S., A.V.S., and G.I.E. Analyzed the data: M.A.S., E.Z.K., and A.V.S. Wrote the paper: A.V.S., M.A.S., and E.Z.K. All authors have read and agreed to the published version of the manuscript.
Funding
The article was financially supported by the Russian Science Foundation (grant #19-16-00016) and by the Ministry of Science and Higher Education of the Russian Federation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Sequences of the RIN locus and the RIN1, RIN2 and RIN-MC transcripts were deposited in the NCBI database (listed in Table 2).
Acknowledgments
We would like to thank Marina Chuenkova for English language editing. This work was performed using the experimental climate control facility in the Institute of Bioengineering (Research Center of Biotechnology, Russian Academy of Sciences).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.P.; Lutts, S. Tomato Fruit Development and Metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlova, R.; Chapman, N.; David, K.; Angenent, G.C.; Seymour, G.B.; de Maagd, R.A. Transcriptional control of fleshy fruit development and ripening. J. Exp. Bot. 2014, 65, 4527–4541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, M.; Shima, Y.; Higuchi, N.; Nakano, T.; Koyama, Y.; Kasumi, T.; Ito, Y. Direct targets of the tomato-ripening regulator RIN identified by transcriptome and chromatin immunoprecipitation analyses. Planta 2012, 235, 1107–1122. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, M.; Nakano, T.; Shima, Y.; Ito, Y. A large-scale identification of direct targets of the tomato MADS box transcription factor RIPENING INHIBITOR reveals the regulation of fruit ripening. Plant Cell 2013, 25, 371–386. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Wang, Y.; Cao, B.; Wang, W.; Tian, S. Unraveling the regulatory network of the MADS box transcription factor RIN in fruit ripening. Plant J. 2012, 70, 243–255. [Google Scholar] [CrossRef]
- Fujisawa, M.; Ito, Y. The regulatory mechanism of fruit ripening revealed by analyses of direct targets of the tomato MADS-box transcription factor RIPENING INHIBITOR. Plant Signal. Behav. 2013, 8, e24357. [Google Scholar] [CrossRef] [Green Version]
- Flores, P.; Sánchez, E.; Fenoll, J.; Hellín, P. Genotypic variability of carotenoids in traditional tomato cultivars. Food Res. Int. 2017, 100, 510–516. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [Green Version]
- Peralta, I.E.; Knapp, S.; Spooner, D.M. Nomenclature for wild and cultivated tomatoes. Rep. Tomato Genet. Coop. 2006, 56, 6–10. [Google Scholar]
- Peralta, I.E.; Spooner, D.M. History, origin and early cultivation of tomato (Solanaceae). Genet. Improv. Solanaceous Crop. 2006, 2, 1–24. [Google Scholar]
- Efremov, G.I.; Slugina, M.A.; Shchennikova, A.V.; Kochieva, E.Z. Differential Regulation of Phytoene Synthase PSY1 During Fruit Carotenogenesis in Cultivated and Wild Tomato Species (Solanum section Lycopersicon). Plants 2020, 9, 1169. [Google Scholar] [CrossRef]
- Slugina, M.A.; Shchennikova, A.V.; Kochieva, E.Z. TAI vacuolar invertase orthologs: The interspecific variability in tomato plants (Solanum section Lycopersicon). Mol. Genet. Genom. 2017, 292, 1123–1138. [Google Scholar] [CrossRef]
- Li, S.; Xu, H.J.L.; Ju, Z.; Cao, D.Y.; Zhu, H.L.; Fu, D.Q.; Grierson, D.; Qin, G.Z.; Luo, Y.B.; Zhu, B.Z. The RIN-MC fusion of MADS-box transcription factors has transcriptional activity and modulates expression of many ripening genes. Plant Physiol. 2018, 176, 891–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Chen, K.; Grierson, D. A critical evaluation of the role of ethylene and MADS transcription factors in the network controlling fleshy fruit ripening. New Phytol. 2019, 221, 1724–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckles, D.M. Factors affecting the postharvest sugar content of tomato Solanum lycopersicum L. fruit. Postharvest Biol. Technol. 2012, 63, 129–140. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0. molecular biology and evolution. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchooa, D. A simple, rapid and efficient method for the extraction of genomic DNA from lychee (Litchi chinensis Sonn.). Afr. J. Biotechnol. 2004, 3, 253–255. [Google Scholar]
- Expósito-Rodríguez, M.; Borges, A.A.; Borges-Pérez, A.; Pérez, J.A. Selection of internal control genes for quantitative real-time RT-PCR studies during tomato development process. BMC Plant Biol. 2008, 8, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bemer, M.; Karlova, R.; Ballester, A.R.; Tikunov, Y.M.; Bovy, A.G.; Wolters-Arts, M.; de Barros Rossetto, P.; Angenent, G.C.; de Maagd, R.A. The tomato FRUITFULL homologs TDR4/FUL1 and MBP7/FUL2 regulate ethylene-independent aspects of fruit ripening. Plant Cell 2012, 24, 4437–4451. [Google Scholar] [CrossRef] [Green Version]
- Immink, R.G.; Ferrario, S.; Busscher-Lange, J.; Kooiker, M.; Busscher, M.; Angenent, G.C. Analysis of the petunia MADS-box transcription factor family. Mol. Genet. Genom. 2003, 268, 598–606. [Google Scholar] [CrossRef]
- Ferrario, S.; Immink, R.G.; Shchennikova, A.; Busscher-Lange, J.; Angenent, G.C. The MADS box gene FBP2 is required for SEPALLATA function in petunia. Plant Cell 2003, 15, 914–925. [Google Scholar] [CrossRef] [Green Version]
- Solovchenko, A.E.; Chivkunova, O.B.; Merzlyak, M.N.; Reshetnikova, I.V. A spectrophotometric analysis of pigments in apples. Russ. J. Plant Physiol. 2001, 48, 693–700. [Google Scholar] [CrossRef]
- Nagata, M.; Yamashita, I. A simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. J. JPN Soc. Food Sci. Technol. 1992, 39, 925–928. [Google Scholar] [CrossRef] [Green Version]
- Meléndez-Martínez, A.J.; Mapelli-Brahm, P.; Benítez-González, A.; Stinco, C.M. A comprehensive review on the colorless carotenoids phytoene and phytofluene. Arch. Biochem. Biophys. 2015, 572, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, J.E.; Rose, A.B. Intron DNA Sequences Can Be More Important Than the Proximal Promoter in Determining the Site of Transcript Initiation. Plant Cell 2017, 29, 843–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shchennikova, A.V.; Shulga, O.A.; Immink, R.; Skryabin, K.G.; Angenent, G.C. Identification and characterization of four chrysanthemum MADS-box genes, belonging to the APETALA1/FRUITFULL and SEPALLATA3 subfamilies. Plant Physiol. 2004, 134, 1632–1641. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Sekiyama, Y.; Nakayama, H.; Nishizawa-Yokoi, A.; Endo, M.; Shima, Y.; Nakamura, N.; Kotake-Nara, E.; Kawasaki, S.; Hirose, S.; et al. Allelic Mutations in the Ripening-Inhibitor Locus Generate Extensive Variation in Tomato Ripening. Plant Physiol. 2020, 183, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lammers, M.; Tikunov, Y.; Bovy, A.G.; Angenent, G.C.; de Maagd, R.A. The rin, nor and Cnr spontaneous mutations inhibit tomato fruit ripening in additive and epistatic manners. Plant Sci. 2020, 294, 110436. [Google Scholar] [CrossRef]
- Manning, K.; Tör, M.; Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Giovannoni, J.J.; Seymour, G.B. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nat. Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef]
- Ito, Y.; Kitagawa, M.; Ihashi, N.; Yabe, K.; Kimbara, J.; Yasuda, J.; Ito, H.; Inakuma, T.; Hiroi, S.; Kasumi, T. DNA-binding specificity, transcriptional activation potential, and the rin mutation effect for the tomato fruit-ripening regulator RIN. Plant J. 2008, 55, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Itkin, M.; Seybold, H.; Breitel, D.; Rogachev, I.; Meir, S.; Aharoni, A. TOMATO AGAMOUS-LIKE 1 is a component of the fruit ripening regulatory network. Plant J. 2009, 60, 1081–1095. [Google Scholar] [CrossRef]
- Vrebalov, J.; Pan, I.L.; Arroyo, A.J.; McQuinn, R.; Chung, M.; Poole, M.; Rose, J.; Seymour, G.; Grandillo, S.; Giovannoni, J.; et al. Fleshy fruit expansion and ripening are regulated by the Tomato SHATTERPROOF gene TAGL1. Plant Cell 2009, 21, 3041–3062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, M.; Shima, Y.; Nakagawa, H.; Kitagawa, M.; Kimbara, J.; Nakano, T.; Kasumi, T.; Ito, Y. Transcriptional regulation of fruit ripening by tomato FRUITFULL homologs and associated MADS box proteins. Plant Cell 2014, 26, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Leseberg, C.H.; Eissler, C.L.; Wang, X.; Johns, M.A.; Duvall, M.R.; Mao, L. Interaction study of MADS-domain proteins in tomato. J. Exp. Bot. 2008, 59, 2253–2265. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, M.; Nakano, T.; Ito, Y. Identification of potential target genes for the tomato fruit-ripening regulator RIN by chromatin immunoprecipitation. BMC Plant Biol. 2011, 11, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, C.; Vrebalov, J.; Tafelmeyer, P.; Giovannoni, J.J. The tomato MADS-box transcription factor RIPENING INHIBITOR interacts with promoters involved in numerous ripening processes in a COLORLESS NONRIPENING-dependent manner. Plant Physiol. 2011, 157, 1568–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, S.; Fei, Z.; Chen, Y.R.; Zheng, Y.; Huang, M.; Vrebalov, J.; McQuinn, R.; Gapper, N.; Liu, B.; Xiang, J.; et al. Single-base resolution methylomes of tomato fruit development reveal epigenome modifications associated with ripening. Nat. Biotechnol. 2013, 31, 154–159. [Google Scholar] [CrossRef]
- Chen, Y.; Li, F.; Wurtzel, E.T. Isolation and characterization of the Z-ISO gene encoding a missing component of carotenoid biosynthesis in plants. Plant Physiol. 2010, 153, 66–79. [Google Scholar] [CrossRef] [Green Version]
- Qin, G.; Zhu, Z.; Wang, W.; Cai, J.; Chen, Y.; Li, L.; Tian, S. A Tomato Vacuolar Invertase Inhibitor Mediates Sucrose Metabolism and Influences Fruit Ripening. Plant Physiol. 2016, 172, 1596–1611. [Google Scholar] [CrossRef] [Green Version]
- Vallarino, J.G.; Yeats, T.H.; Maximova, E.; Rose, J.K.; Fernie, A.R.; Osorio, S. Postharvest changes in LIN5-down-regulated plants suggest a role for sugar deficiency in cuticle metabolism during ripening. Phytochemistry 2017, 142, 11–20. [Google Scholar] [CrossRef]
- Giovannoni, J.J.; DellaPenna, D.; Bennett, A.B.; Fischer, R.L. Expression of a chimeric polygalacturonase gene in transgenic rin (ripening inhibitor) tomato fruit results in polyuronide degradation but not fruit softening. Plant Cell 1989, 1, 53–63. [Google Scholar] [PubMed] [Green Version]
- Wang, D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco-Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Perez Fons, L.; et al. Characterization of CRISPR Mutants Targeting Genes Modulating Pectin Degradation in Ripening Tomato. Plant Physiol. 2019, 179, 544–557. [Google Scholar] [PubMed] [Green Version]
- Rose, J.K.; Lee, H.H.; Bennett, A.B. Expression of a divergent expansin gene is fruit-specific and ripening-regulated. Proc. Natl. Acad. Sci. USA 1997, 94, 5955–5960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Goldman, S.; Coupe, S.; Deikman, J. Ethylene control of E4 transcription during tomato fruit ripening involves two cooperative cis elements. Plant Mol. Biol. 1996, 31, 1117–1127. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic relatedness and structure of the RIN–MC genomic region in S. lycopersicum cultivars, its rin/RIN genotype, and wild tomato species. (A) Phylogenetic tree. (B) The exon–intron structure. ?—the sequence is not defined; bars (black—for RIN; red—for spacer; blue—for MC) correspond to introns, blocks (black—for RIN; blue—for MC)—to exons.
Figure 1.
Phylogenetic relatedness and structure of the RIN–MC genomic region in S. lycopersicum cultivars, its rin/RIN genotype, and wild tomato species. (A) Phylogenetic tree. (B) The exon–intron structure. ?—the sequence is not defined; bars (black—for RIN; red—for spacer; blue—for MC) correspond to introns, blocks (black—for RIN; blue—for MC)—to exons.

Figure 2.
The RIN–MC genomic organization and exon composition of RIN1i, RIN2i and RIN–MC transcripts. Exons are indicated by boxes with the numbers above. The region of the locus subject to changes through rin mutations and gene splicing is colored red.
Figure 2.
The RIN–MC genomic organization and exon composition of RIN1i, RIN2i and RIN–MC transcripts. Exons are indicated by boxes with the numbers above. The region of the locus subject to changes through rin mutations and gene splicing is colored red.

Figure 3.
3′-splicing sites (highlighted yellow) identified in the introns of the RIN gene.

Figure 4.
RIN1i, RIN2i, RIN–MC, and MC mRNA expression in the fruit of Solanum lycopersicum, its rin/RIN genotype, and wild tomato species. (a) Fruit images at different ripening stages; scale bar = 1 cm. (b) Quantitative expression analysis by qRT-PCR. NN-25, the rin/RIN genotype; S. habr., S. habrochaites; S. per., S. peruvianum; IG, immature green; MG, mature green; Br, breaker; RR, ripe red; RF, ripe final. Lowercase letters above the bars indicate statistically significant differences (p < 0.005) between gene expression in different tissues of the same accession: IG, a; MG, b; Br, c; RR (RF), d. For example, the gene expression in IG fruit was significantly different from that in other fruit, which is denoted by letters “bcd” above the bar.
Figure 4.
RIN1i, RIN2i, RIN–MC, and MC mRNA expression in the fruit of Solanum lycopersicum, its rin/RIN genotype, and wild tomato species. (a) Fruit images at different ripening stages; scale bar = 1 cm. (b) Quantitative expression analysis by qRT-PCR. NN-25, the rin/RIN genotype; S. habr., S. habrochaites; S. per., S. peruvianum; IG, immature green; MG, mature green; Br, breaker; RR, ripe red; RF, ripe final. Lowercase letters above the bars indicate statistically significant differences (p < 0.005) between gene expression in different tissues of the same accession: IG, a; MG, b; Br, c; RR (RF), d. For example, the gene expression in IG fruit was significantly different from that in other fruit, which is denoted by letters “bcd” above the bar.

Figure 5.
Expression of the RIN target genes during tomato fruit ripening in Solanum lycopersicum cultivars, its rin/RIN genotype, and wild GF tomato species. NN-25, the rin/RIN genotype; S. hab., S. habrochaites; S. per., S. peruvianum; L, leaves (0); IG, immature green (1); MG, mature green (2); Br, breaker (3); RR, red ripe (or RF, red final) (4). Lowercase letters above the bars indicate statistically significant differences (p < 0.005) between gene expression levels in different tissues of the same accession: IG, a; MG, b; Br, c; RR (RF), d.
Figure 5.
Expression of the RIN target genes during tomato fruit ripening in Solanum lycopersicum cultivars, its rin/RIN genotype, and wild GF tomato species. NN-25, the rin/RIN genotype; S. hab., S. habrochaites; S. per., S. peruvianum; L, leaves (0); IG, immature green (1); MG, mature green (2); Br, breaker (3); RR, red ripe (or RF, red final) (4). Lowercase letters above the bars indicate statistically significant differences (p < 0.005) between gene expression levels in different tissues of the same accession: IG, a; MG, b; Br, c; RR (RF), d.

Figure 6.
Alignment of the C-termini of RIN1 isoforms from S. lycopersicum cultivars, and wild GF species. NN-25, the product of the NN-25 rin allele; RIN2, S. lycopersicum cv. Heinz RIN2 isoform.
Figure 6.
Alignment of the C-termini of RIN1 isoforms from S. lycopersicum cultivars, and wild GF species. NN-25, the product of the NN-25 rin allele; RIN2, S. lycopersicum cv. Heinz RIN2 isoform.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the analyzed tomato accessions.
| Accession | Catalogue Number | Ripe Fruit Color | Primary Storage Sugars |
|---|---|---|---|
| S. lycopersicum cv. Micro-Tom | LA3911 | Red | Glucose, fructose [15] |
| S. lycopersicum cv. Heinz 1706-BG | LA4345 | Red | Glucose, fructose [15] |
| S. lycopersicum cv. Zemba | FSVC 11277 | Red to violet | Glucose, fructose [15] |
| rin/RIN | FSVC NN-25 | Pale yellow | Glucose, fructose 1 |
| RIN/RIN | FSVC NN-21 | Red | Glucose, fructose [15] |
| S. chmielewskii | LA 2663 | Green | Sucrose [15] |
| S. neorickii | LA1326 | Green | n/a |
| S. arcanum | LA2157 | Green | n/a |
| S. huaylasense | LA1365 | Green | Sucrose [15] |
| S. peruvianum | VIRR 4361 | Green | Sucrose [15] |
| S. habrochaites | LYC4 | Green | Sucrose [15] |
| S. pennellii | LA0716 | Green | n/a |
1 This study.
Table 2.
Characteristics of the RIN–MC locus in the analyzed tomato accessions.
| Species | RIN | RIN1i | RIN2i | RIN–MC | Intergenic Spacer bp | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Accession Number | Total bp | Transcript bp | Accession Number | Protein aa | Transcript bp | Accession Number | Protein aa | Transcript bp | Protein aa | Accession Number | ||
| S. lycopersicum cv. Micro-Tom | CM022786.1 (1792…7111) 1 | 5320 | 729 | n/a | 242 | 660 | n/a | 219 | 1194 | 397 | n/a | 2816 |
| S. lycopersicum cv. Heinz 1706 | NC_015442.3 (2081…7368) 1 | 5288 | 729 | MW889896 | 242 | 660 | MW889900 | 219 | 1194 | 397 | MW889904 | n/d |
| S. lycopersicum cv. Zemba | MW889907 | 5291 | 729 | MW889897 | 242 | 660 | MW889901 | 219 | 1194 | 397 | MW889905 | 2511 |
| rin/RIN genotype NN-25 | MW889911 | 5089 | RIN exon VIII is absent in the rin allele | 1194 | 397 | MW889906 | 1395 | |||||
| S. chmielewskii LA 2663 | MT228441 | 5543 | 729 | n/a | n/a | 660 | n/a | 219 | n/a | n/a | n/a | 2741 |
| S. neorickii LA1326 | MW889908 | 6230 | 729 | n/a | n/a | 660 | n/a | 219 | n/a | n/a | n/a | 2749 |
| S. arcanum LA2157 | CBYQ010009972.1 (20826…21107); CBYQ010009973.1 (288...5582) 1 | 5582 | 729 | n/a | n/a | 660 | n/a | 219 | n/a | n/a | n/a | 2748 |
| S. huaylasense LA1365 | MW889909 | 5536 | 729 | n/a | n/a | 660 | n/a | 219 | n/a | n/a | n/a | 2734 |
| S. peruvianum LA4361 | MW889910 | 5742 | 729 | MW889898 | 242 | 660 | n/d | 219 | 1194 | 397 | n/d | 2116 |
| S. habrochaites LYC4 | CBYS010023633.1 (12571…18159) 1 | 5589 | 729 | MW889899 | 242 | 660 | MW889902 | 219 | 1194 | 397 | n/d | 2505 |
| S. pennellii LA0716 | CCXL01022058.1 (2300…7750) 1 | 5451 | 729 | MW928510 | 242 | 660 | MW889903 | 219 | n/a | n/a | n/d | 2754 |
1 Sequences were extracted from the WGS NCBI database; n/d—not detected, n/a—not analyzed.
Table 3.
Carotenoid content in the analyzed tomato accessions.
| Accession | Carotenoid Content in Ripe Fruits (mg/g Wet Weight) | ||
|---|---|---|---|
| Total (x + c) | Lycopene | β-Carotene | |
| S. lycopersicum cv. Heinz 1706-BG | 0.2197 ± 0.0398 | 0.1592 ± 0.0386 | 0.0212 ± 0.0037 |
| S. lycopersicum cv. Zemba | 0.2701 ± 0.0822 | 0.1488 ± 0.0466 | 0.0219 ± 0.034 |
| rin/RIN | 0.0047 ± 0.0011 | 0.0055 ± 0.0027 | 0.0033 ± 0.0013 |
| S. peruvianum | 0.0195 ± 0.0018 | 0 | 0.0242 ± 0.0045 |
| S. habrochaites | 0.0189 ± 0.0065 | 0 | 0.0099 ± 0.0034 |
Table 4.
Protein–protein interactions of RIN1 and RIN2 isoforms 1.
| pAD_GAL4 | pBD_GAL4cam | ˗LH +10 mM 3AT | ˗LTH +10 mM 3AT | ˗LTA | X-gal Test |
|---|---|---|---|---|---|
| Autoactivation test | |||||
| RIN2 | − | − | − | ||
| RIN1 | ++ | + | + | ||
| CDM44 (+control) | + | + | + | ||
| CDM37 (−control) | − | − | − | ||
| Protein–protein interaction test | |||||
| RIN2 | TAGL1 | + | + − | − | |
| RIN2 | FUL2 | + | + − | − | |
| RIN1 | TAGL1 | ++ | ++ | ++ | |
| RIN1 | FUL2 | ++ | ++ | ++ | |
| CDM44 (+control) | CDM37 (+control) | + | + | + | |
| CDM44 (−control) | CDM111 (−control) | − | − | − | |
1 The experiment was carried out both at room temperature and 30 °C and the same results were obtained at both temperatures. L—L-leucine, H—L-histidine, T—L-tryptophan, A—L-adenine hemisulfate salt, 3AT—3-amino-1,2,4-triazole, ˗LH, ˗LTH and ˗LTA—medium devoid of these amino acids, X-gal—5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside. Chrysanthemum MADS-domain proteins CDM44, CDM37, and CDM111 were used as controls [26].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Slugina, M.A.; Efremov, G.I.; Shchennikova, A.V.; Kochieva, E.Z. Characterization of RIN Isoforms and Their Expression in Tomato Fruit Ripening. Cells 2021, 10, 1739. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071739
AMA Style
Slugina MA, Efremov GI, Shchennikova AV, Kochieva EZ. Characterization of RIN Isoforms and Their Expression in Tomato Fruit Ripening. Cells. 2021; 10(7):1739. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071739
Chicago/Turabian StyleSlugina, Maria A., Gleb I. Efremov, Anna V. Shchennikova, and Elena Z. Kochieva. 2021. "Characterization of RIN Isoforms and Their Expression in Tomato Fruit Ripening" Cells 10, no. 7: 1739. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071739
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.