
Transcriptomic Profile Reveals Deregulation of Hearing-Loss Related Genes in Vestibular Schwannoma Cells Following Electromagnetic Field Exposure

 , ,
, ,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. EMF Treatment
2.3. Immunofluorescence (IFL) and Cell Characterization
2.4. In Vitro Cell Proliferation, Viability, and Migration Assays
2.5. RNA Preparation and qRT-PCR Analysis
2.6. Statistical Analysis
2.7. NGS Technology RNA-Seq
2.8. Bioinformatic Analysis of Transcriptome
3. Results
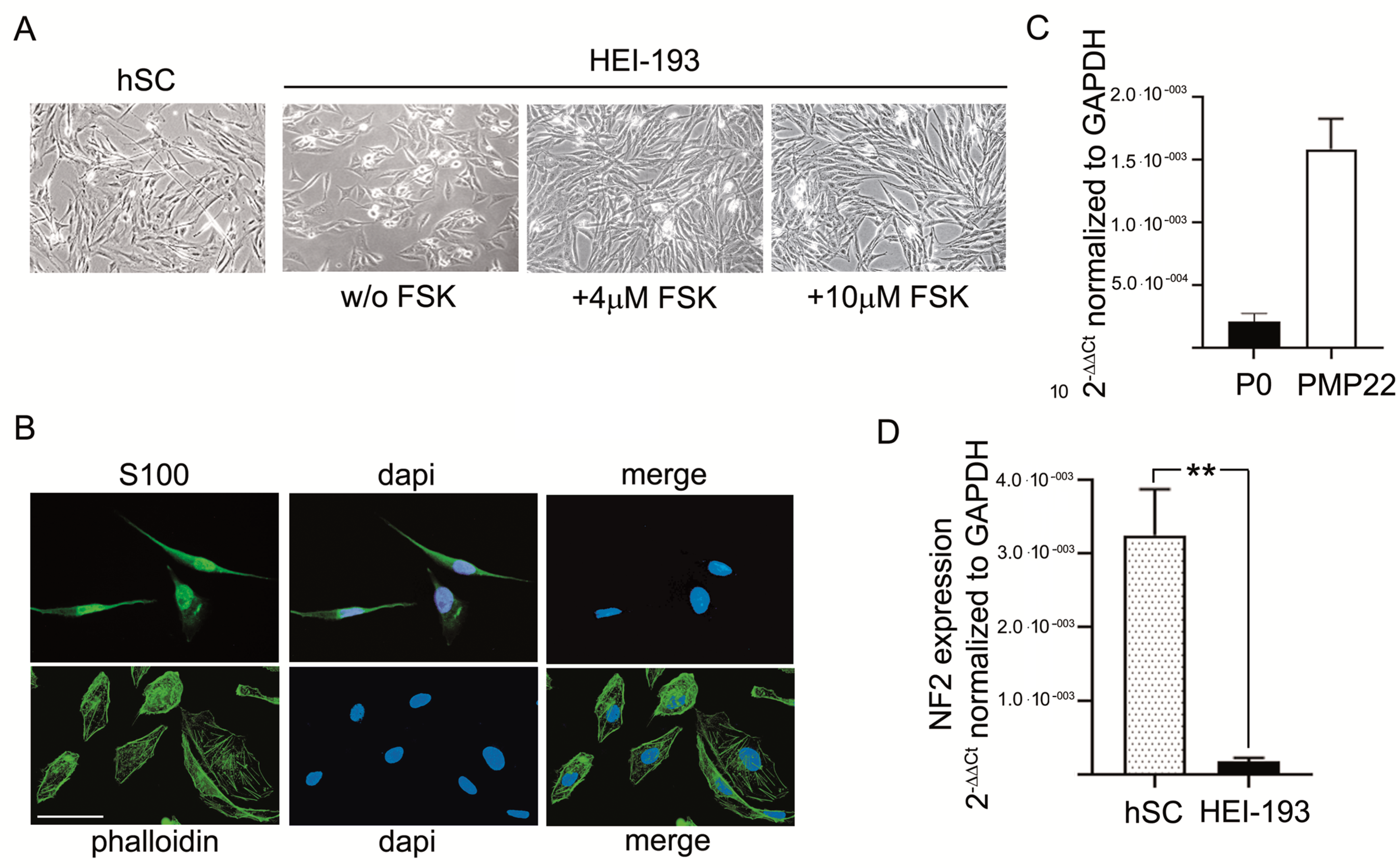
3.1. Characterization of Human Schwannoma-Derived Cells
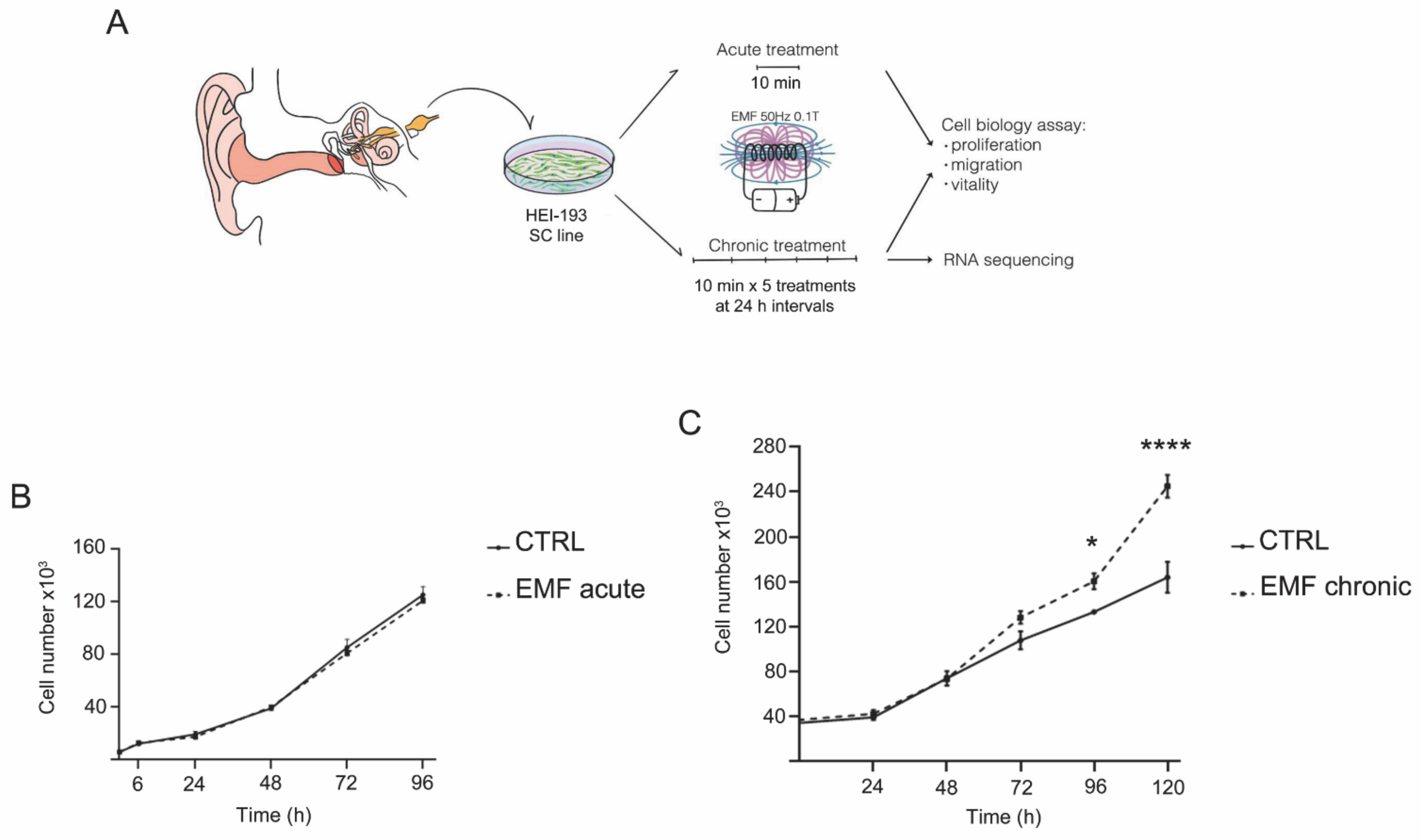
3.2. Chronic Exposure to EMF Induces Proliferative Changes in VS Cells
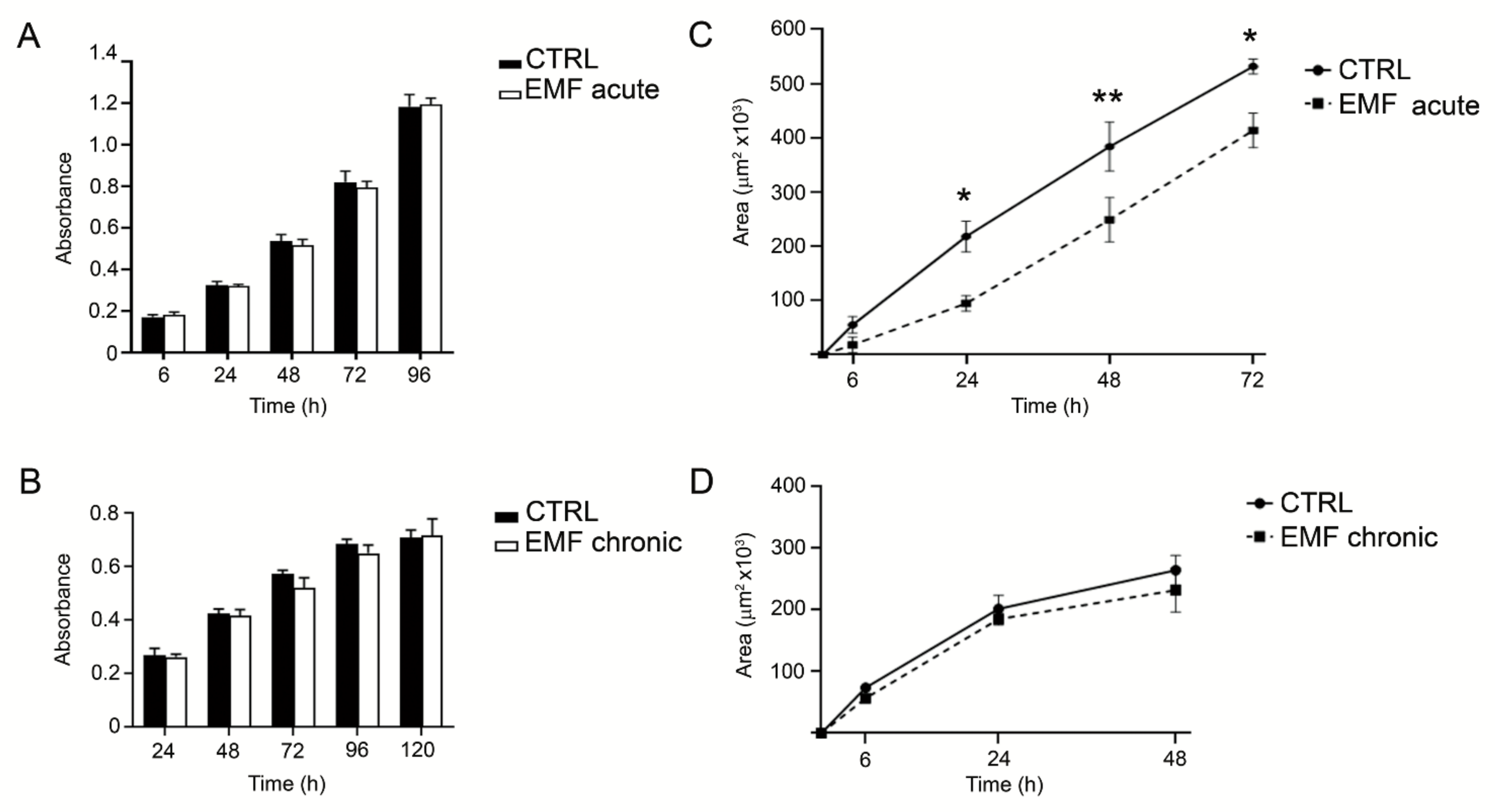
3.3. Chronic EMF Exposure Effect on Viability and Migration of VS Cells
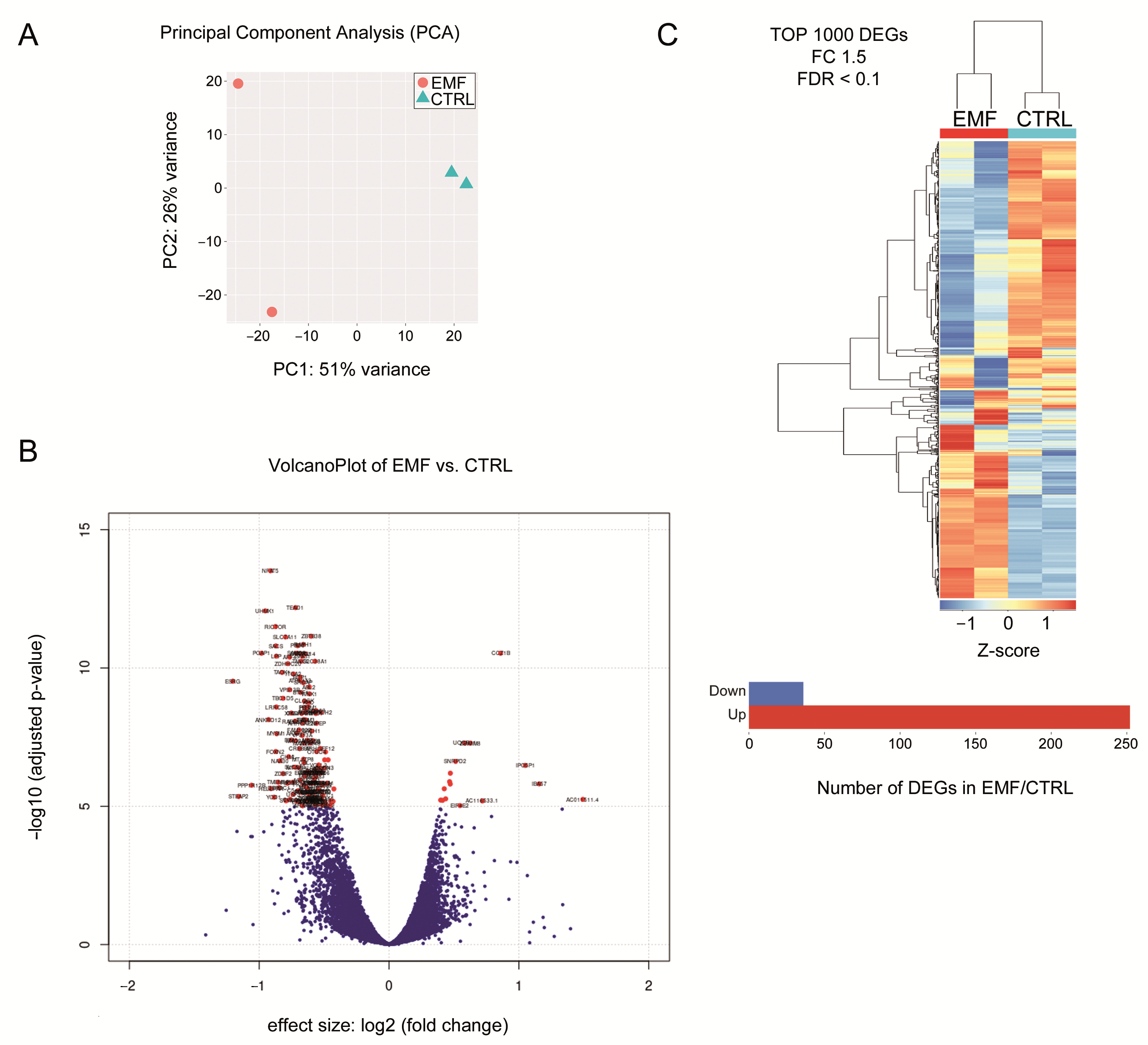
3.4. Identification of Differentially Expression Genes (DEG) in HEI-193 Cells Following Chronic EMF Exposure
3.5. Chronic EMF Exposure Changed Metabolic Pathways of HEI-193 Cells
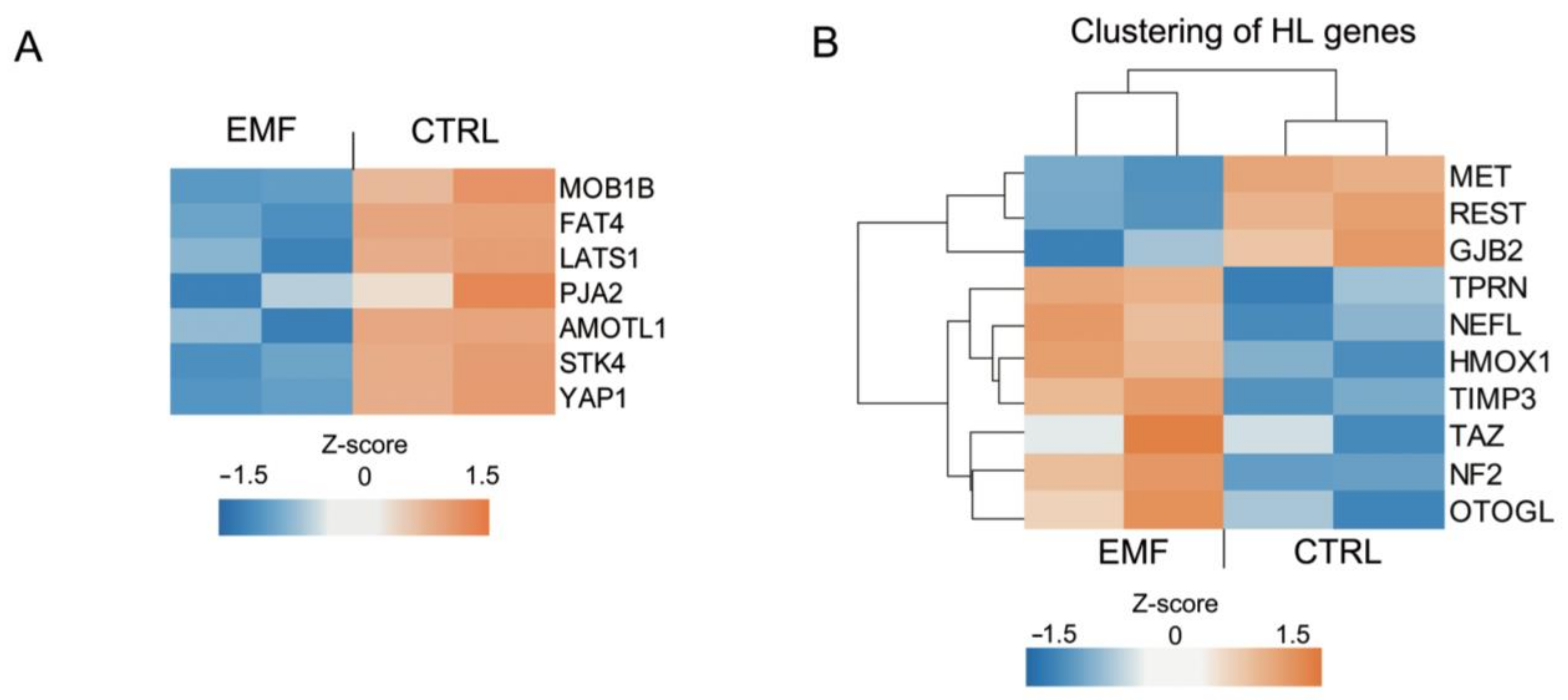
3.6. Hippo Pathway- and HL-Related Gene Expression Changes in HEI-193 Cells Following Chronic EMF Exposure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, V.K.; Thakker, A.; Gupta, K.K. Vestibular Schwannoma: What We Know and Where We are Heading. Head. Neck Pathol. 2020, 14, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Harner, S.G.; Fabry, D.A.; Beatty, C.W. Audiometric findings in patients with acoustic neuroma. Am. J. Otol. 2000, 21, 405–411. [Google Scholar] [CrossRef]
- Evans, D.G.; Moran, A.; King, A.; Saeed, S.; Gurusinghe, N.; Ramsden, R. Incidence of vestibular schwannoma and neurofibromatosis 2 in the North West of England over a 10-year period: Higher incidence than previously thought. Otol. Neurotol. 2005, 26, 93–97. [Google Scholar] [CrossRef]
- Hadfield, K.D.; Smith, M.J.; Urquhart, J.E.; Wallace, A.J.; Bowers, N.L.; King, A.T.; Rutherford, S.A.; Trump, D.; Newman, W.G.; Evans, D.G. Rates of loss of heterozygosity and mitotic recombination in NF2 schwannomas, sporadic vestibular schwannomas and schwannomatosis schwannomas. Oncogene 2010, 29, 6216–6221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamenkovic, I.; Yu, Q. Merlin, a “magic” linker between extracellular cues and intracellular signaling pathways that regulate cell motility, proliferation, and survival. Curr. Protein Pept. Sci. 2010, 11, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, K.A.; Shevde, L.A. Merlin: The wizard requires protein stability to function as a tumor suppressor. Biochim. Biophys. Acta 2012, 1826, 400–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colciago, A.; Melfi, S.; Giannotti, G.; Bonalume, V.; Ballabio, M.; Caffino, L.; Fumagalli, F.; Magnaghi, V. Tumor suppressor Nf2/merlin drives Schwann cell changes following electromagnetic field exposure through Hippo-dependent mechanisms. Cell Death Discov. 2015, 1, 15021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, V. Merlin and the ERM proteins in Schwann cells, neurons and growth cones. Nat. Rev. Neurosci. 2004, 5, 462–470. [Google Scholar] [CrossRef]
- Agrawal, Y.; Carey, J.P.; Della Santina, C.C.; Schubert, M.C.; Minor, L.B. Disorders of balance and vestibular function in US adults: Data from the National Health and Nutrition Examination Survey, 2001–2004. Arch. Intern Med. 2009, 169, 938–944. [Google Scholar] [CrossRef]
- Stangerup, S.E.; Tos, M.; Thomsen, J.; Caye-Thomasen, P. True incidence of vestibular schwannoma? Neurosurgery 2010, 67, 1335–1340. [Google Scholar] [CrossRef]
- Hardell, L.; Carlberg, M.; Hansson Mild, K. Epidemiological evidence for an association between use of wireless phones and tumor diseases. Pathophysiology 2009, 16, 113–122. [Google Scholar] [CrossRef]
- Hardell, L.; Carlberg, M.; Hansson Mild, K. Use of mobile phones and cordless phones is associated with increased risk for glioma and acoustic neuroma. Pathophysiology 2013, 20, 85–110. [Google Scholar] [CrossRef]
- Hardell, L.; Carlberg, M.; Hansson Mild, K. Mobile phone use and the risk for malignant brain tumors: A case-control study on deceased cases and controls. Neuroepidemiology 2010, 35, 109–114. [Google Scholar] [CrossRef]
- Melfi, S.; Colciago, A.; Giannotti, G.; Bonalume, V.; Caffino, L.; Fumagalli, F.; Magnaghi, V. Stressing out the Hippo/YAP signaling pathway: Toward a new role in Schwann cells. Cell Death Dis. 2015, 6, e1915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, G.; Faudoa, R.; Li, X.; Xeu, Z.; Brackmann, D.E.; Hitselberg, W.; Saleh, E.; Lee, F.; Gutmann, D.H.; Slattery, W., 3rd; et al. Establishment of primary vestibular schwannoma cultures from neurofibromatosis type-2 patients. Int. J. Oncol. 1999, 14, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Hung, G.; Li, X.; Faudoa, R.; Xeu, Z.; Kluwe, L.; Rhim, J.S.; Slattery, W.; Lim, D. Establishment and characterization of a schwannoma cell line from a patient with neurofibromatosis 2. Int. J. Oncol. 2002, 20, 475–482. [Google Scholar] [CrossRef]
- Piovesana, R.; Faroni, A.; Tata, A.M.; Reid, A.J. Functional Characterization of Muscarinic Receptors in Human Schwann Cells. Int. J. Mol. Sci. 2020, 21, 6666. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Dilwali, S.; Patel, P.B.; Roberts, D.S.; Basinsky, G.M.; Harris, G.J.; Emerick, K.S.; Stankovic, K.M. Primary culture of human Schwann and schwannoma cells: Improved and simplified protocol. Hear. Res. 2014, 315, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearer, A.E.; DeLuca, A.P.; Hildebrand, M.S.; Taylor, K.R.; Gurrola, J., 2nd; Scherer, S.; Scheetz, T.E.; Smith, R.J. Comprehensive genetic testing for hereditary hearing loss using massively parallel sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 21104–21109. [Google Scholar] [CrossRef] [Green Version]
- Tekin, D.; Yan, D.; Bademci, G.; Feng, Y.; Guo, S.; Foster, J., 2nd; Blanton, S.; Tekin, M.; Liu, X. A next-generation sequencing gene panel (MiamiOtoGenes) for comprehensive analysis of deafness genes. Hear. Res. 2016, 333, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Schularick, N.M.; Clark, J.J.; Hansen, M.R. Primary culture of human vestibular schwannomas. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hanz, S.; Fainzilber, M. Retrograde signaling in injured nerve--the axon reaction revisited. J. Neurochem. 2006, 99, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ducommun Priest, M.; Navarro, M.F.; Bremer, J.; Granato, M. Dynein promotes sustained axonal growth and Schwann cell remodeling early during peripheral nerve regeneration. PLoS Genet. 2019, 15, e1007982. [Google Scholar] [CrossRef]
- Langworthy, M.M.; Appel, B. Schwann cell myelination requires Dynein function. Neural. Dev. 2012, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, E.; Ko, P.; Jeong, J.; Keum, S.; Kim, J.W.; Seo, Y.J.; Song, W.K.; Rhee, S. Dynein-mediated nuclear translocation of yes-associated protein through microtubule acetylation controls fibroblast activation. Cell Mol. Life Sci. 2020, 77, 4143–4161. [Google Scholar] [CrossRef]
- Marcinkiewicz, M.; Savaria, D.; Marcinkiewicz, J. The pro-protein convertase PC1 is induced in the transected sciatic nerve and is present in cultured Schwann cells: Comparison with PC5, furin and PC7, implication in pro-BDNF processing. Brain Res. Mol. Brain Res. 1998, 59, 229–246. [Google Scholar] [CrossRef]
- Ren, Y.; Stankovic, K.M. The Role of Tumor Necrosis Factor Alpha (TNFalpha)in Hearing Loss and Vestibular Schwannomas. Curr. Otorhinolaryngol. Rep. 2018, 6, 15–23. [Google Scholar] [CrossRef]
- Dilwali, S.; Landegger, L.D.; Soares, V.Y.; Deschler, D.G.; Stankovic, K.M. Secreted Factors from Human Vestibular Schwannomas Can Cause Cochlear Damage. Sci. Rep. 2015, 5, 18599. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, S.; Gugel, I.; Remke, M.; Bornemann, A.; Pantazis, G.; Mack, S.C.; Shih, D.; Singh, S.K.; Sabha, N.; Taylor, M.D.; et al. Gene-expression profiling elucidates molecular signaling networks that can be therapeutically targeted in vestibular schwannoma. J. Neurosurg. 2014, 121, 1434–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, K.J.; Kiang, A.; Wang-Rodriguez, J.; Yu, M.A.; Doherty, J.K.; Ongkeko, W.M. EGF and bFGF promote invasion that is modulated by PI3/Akt kinase and Erk in vestibular schwannoma. Otol. Neurotol. 2011, 32, 308–314. [Google Scholar] [CrossRef] [PubMed]
- James, M.F.; Han, S.; Polizzano, C.; Plotkin, S.R.; Manning, B.D.; Stemmer-Rachamimov, A.O.; Gusella, J.F.; Ramesh, V. NF2/merlin is a novel negative regulator of mTOR complex 1, and activation of mTORC1 is associated with meningioma and schwannoma growth. Mol. Cell Biol. 2009, 29, 4250–4261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, M.F.; Stivison, E.; Beauchamp, R.; Han, S.; Li, H.; Wallace, M.R.; Gusella, J.F.; Stemmer-Rachamimov, A.O.; Ramesh, V. Regulation of mTOR complex 2 signaling in neurofibromatosis 2-deficient target cell types. Mol. Cancer Res. 2012, 10, 649–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Tekin, M.; Blanton, S.H.; Liu, X.Z. Next-generation sequencing in genetic hearing loss. Genet. Test Mol. Biomark. 2013, 17, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Meier, C.; Dermietzel, R.; Davidson, K.G.; Yasumura, T.; Rash, J.E. Connexin32-containing gap junctions in Schwann cells at the internodal zone of partial myelin compaction and in Schmidt-Lanterman incisures. J. Neurosci. 2004, 24, 3186–3198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, Y.; Kelly, M.C.; Rehman, A.U.; Boger, E.T.; Morell, R.J.; Kelley, M.W.; Friedman, T.B.; Banfi, B. Defects in the Alternative Splicing-Dependent Regulation of REST Cause Deafness. Cell 2018, 174, 536–548.e21. [Google Scholar] [CrossRef] [Green Version]
- Kelsell, D.P.; Dunlop, J.; Stevens, H.P.; Lench, N.J.; Liang, J.N.; Parry, G.; Mueller, R.F.; Leigh, I.M. Connexin 26 mutations in hereditary non-syndromic sensorineural deafness. Nature 1997, 387, 80–83. [Google Scholar] [CrossRef]
- Kim, M.; Kim, H.; Kim, D.; Huh, Y.; Park, C.; Chung, H.J.; Jung, J.; Jeong, N.Y. Heme Oxygenase 1 in Schwann Cells Regulates Peripheral Nerve Degeneration Against Oxidative Stress. ASN Neuro. 2019, 11, 1759091419838949. [Google Scholar] [CrossRef] [Green Version]
- Hardell, L.; Carlberg, M.; Soderqvist, F.; Mild, K.H. Pooled analysis of case-control studies on acoustic neuroma diagnosed 1997-2003 and 2007-2009 and use of mobile and cordless phones. Int. J. Oncol. 2013, 43, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Moon, I.S.; Kim, B.G.; Kim, J.; Lee, J.D.; Lee, W.S. Association between vestibular schwannomas and mobile phone use. Tumour Biol. 2014, 35, 581–587. [Google Scholar] [CrossRef] [Green Version]
- INTERPHONE Study Group. Acoustic neuroma risk in relation to mobile telephone use: Results of the INTERPHONE international case-control study. Cancer Epidemiol. 2011, 35, 453–464. [Google Scholar] [CrossRef]
- Kesari, K.K.; Siddiqui, M.H.; Meena, R.; Verma, H.N.; Kumar, S. Cell phone radiation exposure on brain and associated biological systems. Indian J. Exp. Biol. 2013, 51, 187–200. [Google Scholar]
- Funk, R.H.; Monsees, T.K. Effects of electromagnetic fields on cells: Physiological and therapeutical approaches and molecular mechanisms of interaction. A review. Cells Tissues Organs 2006, 182, 59–78. [Google Scholar] [CrossRef] [Green Version]
- Messerli, M.A.; Graham, D.M. Extracellular electrical fields direct wound healing and regeneration. Biol. Bull. 2011, 221, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, K.R.; Glaser, R. Thermal mechanisms of interaction of radiofrequency energy with biological systems with relevance to exposure guidelines. Health Phys. 2007, 92, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Baan, R.; Grosse, Y.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Islami, F.; Galichet, L.; Straif, K. Carcinogenicity of radiofrequency electromagnetic fields. Lancet Oncol. 2011, 12, 624–626. [Google Scholar] [CrossRef]
- Gamboa, O.L.; Gutierrez, P.M.; Alcalde, I.; De la Fuente, I.; Gayoso, M.J. Absence of relevant effects of 5 mT static magnetic field on morphology, orientation and growth of a rat Schwann cell line in culture. Histol. Histopathol. 2007, 22, 777–780. [Google Scholar] [CrossRef]
- Cuccurazzu, B.; Leone, L.; Podda, M.V.; Piacentini, R.; Riccardi, E.; Ripoli, C.; Azzena, G.B.; Grassi, C. Exposure to extremely low-frequency (50 Hz) electromagnetic fields enhances adult hippocampal neurogenesis in C57BL/6 mice. Exp. Neurol. 2010, 226, 173–182. [Google Scholar] [CrossRef]
- Sisken, B.F.; Kanje, M.; Lundborg, G.; Herbst, E.; Kurtz, W. Stimulation of rat sciatic nerve regeneration with pulsed electromagnetic fields. Brain Res. 1989, 485, 309–316. [Google Scholar] [CrossRef]
- Kanje, M.; Rusovan, A.; Sisken, B.; Lundborg, G. Pretreatment of rats with pulsed electromagnetic fields enhances regeneration of the sciatic nerve. Bioelectromagnetics 1993, 14, 353–359. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Direction | NES | p adj | |
|---|---|---|---|---|
| Cotranslational protein targeting to membrane | Up | 7.859 | 5.30 × 10−10 | |
 | Protein targeting to ER | Up | 7.807 | 5.30 × 10−10 |
| SRP dependent cotranslational protein targeting to membrane | Up | 7.797 | 5.30 × 10−10 | |
| Establishment of protein localization to endoplasmic reticulum | Up | 7.585 | 5.30 × 10−10 | |
 | Mitochondrial translational elongation | Up | 6.081 | 6.10 × 10−6 |
| | Mitochondrial translational termination | Up | 6.022 | 6.70 × 10−6 |
| | Translational termination | Up | 5.668 | 1.50 × 10−5 |
| Mitochondrial respiratory chain complex assembly | Up | 4.546 | 9.00 × 10−4 | |
| Mitochondrial ATP synthesis coupled electron transport | Up | 4.404 | 1.20 × 10−3 | |
| Cytoplasmic translation | Up | 4.360 | 1.50 × 10−3 | |
| ATP synthesis coupled electron transport | Up | 4.234 | 1.90 × 10−3 | |
| NADH dehydrogenase complex assembly | Up | 3.955 | 6.60 × 10−3 | |
| Mitochondrial respiratory chain complex I assembly | Up | 3.955 | 6.60 × 10−3 | |
| Ribosomal large subunit biogenesis | Up | 3.920 | 6.60 × 10−3 | |
| Respiratory electron transport chain | Up | 3.858 | 6.60 × 10−3 | |
| | Ribosome assembly | Up | 3.414 | 3.10 × 10−2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colciago, A.; Audano, M.; Bonalume, V.; Melfi, V.; Mohamed, T.; Reid, A.J.; Faroni, A.; Greer, P.A.; Mitro, N.; Magnaghi, V. Transcriptomic Profile Reveals Deregulation of Hearing-Loss Related Genes in Vestibular Schwannoma Cells Following Electromagnetic Field Exposure. Cells 2021, 10, 1840. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071840
Colciago A, Audano M, Bonalume V, Melfi V, Mohamed T, Reid AJ, Faroni A, Greer PA, Mitro N, Magnaghi V. Transcriptomic Profile Reveals Deregulation of Hearing-Loss Related Genes in Vestibular Schwannoma Cells Following Electromagnetic Field Exposure. Cells. 2021; 10(7):1840. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071840
Chicago/Turabian StyleColciago, Alessandra, Matteo Audano, Veronica Bonalume, Valentina Melfi, Tasnim Mohamed, Adam J. Reid, Alessandro Faroni, Peter A. Greer, Nico Mitro, and Valerio Magnaghi. 2021. "Transcriptomic Profile Reveals Deregulation of Hearing-Loss Related Genes in Vestibular Schwannoma Cells Following Electromagnetic Field Exposure" Cells 10, no. 7: 1840. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10071840